")
Back to Journals » Infection and Drug Resistance » Volume 18
A New Composite Integrative and Conjugative Element Mediates Multiple Drug Resistance from Streptococcus anginosus to Streptococcus agalactiae
Authors Wang Y , Liu T , Yi S, Zhu Y
Received 25 December 2024
Accepted for publication 14 April 2025
Published 18 April 2025 Volume 2025:18 Pages 1941—1949
DOI https://doi.org/10.2147/IDR.S514281
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Sandip Patil
Yingting Wang,1,* Taoran Liu,1,* Sida Yi,2 Yuanting Zhu3,4
1Department of Critical Care Medicine, The First Affiliated Hospital, Sun Yat-Sen University, Guangzhou, 510080, People’s Republic of China; 2Institute of Antibiotics, Huashan Hospital, Fudan University, Shanghai, People’s Republic of China; 3NHC Key Laboratory of Assisted Circulation, The First Affiliated Hospital of Sun Yat-Sen University, Guangzhou, 510080, People’s Republic of China; 4Department of Cardiology, The Seventh Affiliated Hospital of Sun Yat-Sen University, Shenzhen, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Yuanting Zhu, Department of Cardiology, The Seventh Affiliated Hospital of Sun Yat-Sen University, Shenzhen, People’s Republic of China, Tel +86-18501230142, Email [email protected]
Introduction: Integrative and conjugative elements (ICEs) are a category of horizontal mobile genetic elements (MGEs) that play important roles in mediating the spread of antimicrobial resistance in Streptococci.
Methods: In this study, a novel ICE, namely ICESan 26_rplL, was identified from commensal Streptococcus anginosus through routine silico analyses. The genetic characterization of ICESan 26_rplL was explored based on a comparison with known ICEs. The cyclization and cross-species transferability (from Streptococcus anginosus to Streptococcus agalactiae) of ICESan 26_rplL were explored using inverse PCR and conjugation transfer experiments.
Results: ICESan 26_rplL is 61.618 kb in length at downstream of rplL and carries multiple antibiotic resistance genes, including erm(B) [for erythromycin, clindamycin, and streptococcin B (MLSB)], tetM-tetL (tetracycline), lnu and lsa(E) (clindamycin), and ant(6)-I (aminoglycosides). The comparative analysis results showed that ICESan 26_rplL was a composite ICE with a mosaic structure composed of modules in the ICESa2603 and TnGBS families. The inverse PCR results demonstrated that ICESan 26_rplL could be excised from the chromosome to form a circular intermediate. The conjugation transfer experiment and sequencing results confirmed the cross-species transfer of ICESan 26_rplL to S. agalactiae recipients at a relatively low frequency (2.13 × 10− 8). Moreover, core functional modules were retrieved from GenBank to search for any ICEs related to ICESan 26_rplL. Eventually, 53 putative ICEs were identified, including three composite ICEs with high similarity to ICESan 26_rplL, three ICEs in the TnGBS family, and 47 ICEs in the ICESa 2603 family.
Discussion: In this study, a novel mosaic ICE was reported for the first time. In addition, its transfer to the major pathogen Streptococcus agalactiae was characterized and proved. These findings suggest that this ICE is expected to become a vehicle for the dissemination of multiple antimicrobial resistance through the wider pathogen Streptococci.
Keywords: Streptococcus anginosus, Streptococcus agalactiae, ICEs, antibiotic resistance genes, conjugation transfer, composite
Introduction
Streptococcus anginosus, is an opportunistic pathogen and along with Streptococcus constellatus and Streptococcus intermedius belong to the Streptococcus anginosus group (SAG).1 The latest research has reported that, in addition to Helicobacter pylori, S. anginosus infection can also spontaneously induce chronic gastritis, parietal cell atrophy, mucinous metaplasia and dysplasia, increasing the risk of gastric cancer (GC).2,3 Streptococcus agalactiae, also known as group B Streptococcus (GBS), is a major pathogen inducing neonatal meningitis and sepsis.4
The prolonged administration of antibiotics has resulted in rising resistance of the streptococcal group to macrolides, lincosamides, streptogramin B (MLSB), tetracyclines, and aminoglycosides. Macrolide resistance in the streptococcal group can be mainly attributed to ERM methylase-mediated target modification, which is encoded by genes in the erm family.5 These methylases can modify 23S rRNA bacterial ribosomes, thus preventing the specific binding of macrolides, lincosamides, and streptogramin B antibiotics (MLSB) and rendering related resistance. Lnu transferases, encoded by the lnu gene, can inactivate lincosamides by modifying their third or fourth hydroxyl groups. This gene can only mediate lincomycin resistance. Lnu transferases include Lnu(A), Lnu(B), Lnu(C), Lnu(D), Lnu(E), and Lnu(F).6,7 Through the same mechanism, ant(6) can encode aminoglycoside nucleoside transferases to inactivate aminoglycoside antibiotics, such as streptomycin or gentamicin.8 lsa(E) can encode ATP-binding cassette (ABC) transporters, rendering resistance to lincosamides, plectin, and streptococcin B through efflux.9 The tet family, including tet(M) and tet(O), can encode ribosomal protection proteins (RPRs) to confer tetracycline resistance.10,11 These antibiotic resistance genes (ARGs) can be detected in pathogenic microorganisms or normal flora, and it is difficult to eliminate them. Under certain conditions, ARGs can be transmitted between microorganisms along with mobile genetic elements (MGEs), further leading to the spread of bacterial resistance.
Integrative and conjugative elements (ICEs) are a category of self-transmitted MGEs that have been shown to reside in the bacterial genome chromosome.12 They have a typical modular structure, including integrases, relaxases, and modules in Type IV secretion systems (T4SSs) as well as diverse cargos. Active ICEs can be excised from chromosomes to form a circular intermediate, and one of the DNA strands is transferred mediated by T4SSs to a new host through rolling replication. Single-stranded ICEs can synthesize double-stranded genes via relaxases and integrases into specific sites in the chromosome through the integrase module.13 In this process, ARGs,14,15 virulence,16 or other niche adaptation genes harbored by ICEs are transmitted. As the most well-studied ICE group in streptococci, ICESa2603 has been widely reported in studies on Streptococcus agalactiae, Streptococcus pyogenes, Streptococcus pneumoniae, and zoonotic Streptococcus suis.17 They share similar tyrosine integrases, which can recognize the 3′ -end of the 50S ribosomal subunit protein L7/L12 gene (rplL). Multiple ARGs, such as erm(B) (MLSB), aphA3, aadE, sat, ant6 (aminoglycoside), tetM, and tetO (tetracycline) have been reported in the ICE group. TnGBS is another broad and diverse ICE family of streptococci, and it is characterized by a DDE transposon containing an active triad of acidic amino acids (D.D.E).18 Currently, ARG-related ICEs in S. anginosus are currently underrepresented in existing studies. In addition, the transfer or cross-species transfer of ARGs mediated by the ICESa2603 or TnGBS family has not been reported in studies on S. anginosus, and their epidemiology and genetic characterization have not been fully clarified.
In this study, a new ICE from the clinical isolate S. anginosus San26 was identified and characterized, and its transfer from S. anginosus to the major pathogen S. agalactiae was also proved. The ICE, named ICESan26_rplL, carried multiple ARGs that conferred host MLSB, tetracycline, and aminoglycoside resistance. Moreover, the ICE had a unique mosaic structure composed of modules from different ICE families. However, these aspects in streptococci have never been fully investigated.
Materials and Methods
Bacterial Strains and Antimicrobial Susceptibility Tests
In this study, the next-generation sequencing (NGS) drafts of clinical S. anginosus San26 (JAXIIF000000000) were available. The recipient bacterial S. agalactiae SagR331 was the clinical isolate preserved in Huashan Hospital. ATCC13813 was used as the quality control strain for the antimicrobial susceptibility test, and it was preserved in The First Affiliated Hospital, Sun Yat-sen University. The breakpoint complied with the standards of Clinical and Laboratory Standards Institute (CLSI). The clinical sample used in this study was partially collected from routine hospital laboratory procedures, and no other personal information was involved in this study.
PCR Amplifications
The PCR amplifications were performed to verify the ICE extrachromosomal circular intermediate, which was key evidence for ICE transferability. Designed primer pairs P1-P4 were used for ICESan26_rplL and PI1-PI4 for internal TnGBS-like ICE. Primers were listed in Table S1.
Conjugation Transfer Experiments
Recipient bacterial need to display opposite resistance phenotype to donor and contained at least one unoccupied rplL genes site for ICESan26_rplL. Therefore, the S. agalactiae SagR331 (CP138369) which resistant to levofloxacin but sensitive to erythromycin and clindamycin and the genome contained a rplL gene as a putative target for ICESan26_rplL.
Conjugation transfer experiment was performed as described previously.19,20 Donors and recipients were thawed and grown to OD600=0.5, and mixed by 1:10 (donor and recipient). The mixed bacterial solution was uniformly coated on nitrocellulose membrane and incubated at 37 °C for 4–6 hours. Then, 10 mg/mL DNase was added to eliminate potential effects of DNA transformation. After the incubation, the mixed bacteria were diluted and cultured in the medium containing 50 μg/mL erythromycin, 50 μg/mL clindamycin, 20 μg/mL levofloxacin for 2–5 days. Selected clones were further verified by PCR and sequencing.
DNA Sequencing and Comparison Analysis
Conjugation transfer of ICE was confirmed through NGS sequencing of recipients and relative transconjugants. DNA was extracted using QIAGEN Midi Kit (Qiagen, Hilden, Germany). The whole-genome sequencing was performed at BGI Genomics (HiSeq X; Illumina, San Diego, CA, USA). High-quality reads were assembled by Spades v3.9.03421 and uploaded to the online annotation tool RAST22 (rast.nmpdr.org).
To visualize the obtained ICE and excluded potential influences of other elements, the sequences of recipient and transconjugant were aligned using Mauve v2.4.0.23 Firstly, NGS sequences of recipient and relative transconjugant were arranged using the function of order-contig of Mauve v2.4.0 basing on a reference complete genome S. agalactiae NGBS128 (CP012480).24 Then, the rearranged genome of transconjugant was aligned with recipient, and difference regions were location of ICESan26_rplL.
Bioinformatics Identification of ICE Similar with ICESan26_rplL
Although ICEs vary in genetic organization and sequence, many share similar core functional genes including, integrases, relaxases and T4SSs. As previously described,20,25,26 the core functional genes (integraseICESan26-I, integraseICESan26-E, relaxaseICESan26-I, relaxaseICESan26-E, and T4SSs) of ICESan26_rplL were used as query terms for blastN to identify similar ICEs. More specifically, ICEs could be classified into the same family, namely the ICESan26_rplL family, if all their core function genes showed >60% sequence identity to ICESan26_rplL; ICEs could be classified as ICESan26_rplL-related ICEs, if their integrases showed >60% sequence identity and at least one of the other core function genes showed >60% sequence identity to ICESan26_rplL. The boundaries were manually delimited, and the ARGs carried by ICEs were predicted using ResFinder.27
Results
Identification and Characterization of ICESan26_rplL
The clinical isolate S. anginosus San26 exhibited high resistance to erythromycin, clindamycin, tetracycline, and gentamycin. The sequence analysis results of San26 revealed multiple ARGs, including erm(B), tetM, tetL, ant(6)-I, lnu, and lsa(E), within a small genetic range. Through the prediction using the online tool ICEFinder (bioinfo-mml.sjtu.edu.cn/ICEfinder/index.php), it was found that these ARGs were carried by a putative ICE at the downstream segment of rplL, an integrating hotspot of the ICESa2603 family in the Streptococcal group.17 The newly discovered ICE was named ICESan26_rplL according to its host number, abbreviation, and integration site.
ICESan26_rplL (JAXIIF010000002, 92,631–154249) was found to be 61.618 kb in length and had 64 open reading frames (ORFs) with a direct repeat sequence 5ʹ-TTATTTAAGAGTAAC-3ʹ. The backbones of ICESan26_rplL were highly related to five reported ICEs, including ICESa2603 in the Streptococcus agalactiae strain 2603 V/R;17 ICESsuYS388, an ICE carrying erm(B) and tetO from zoonotic Streptococcus suis;28 ICESpy009, an ICE carrying mef(E) and mel originally identified in S. pyogenes;29 ICESp1116, a genetic element responsible for erm(B)-mediated ICEs in S. pyogenes;30 TnGBS2.6, an ARGs-free ICE from S. agalactiae18 (Figure 1a). ICESa2603, ICESsuYS388, and ICESpy009 shared related backbones and could be classified into the streptococcal ICESa2603 family, whereas ICESp1116 and TnGBS2.6 could be classified into the TnGBS family. The in silico comparison results indicated ICESan26_rplL had a typical mosaic structure, and it was composed of three regions. The first region included the integraseICESan26-E, relaxaseICESan26-E, and virB1 of T4SSs at both ends of ICESan26_rplL, which were closely related to the corresponding modules of the ICESa2603 family. The second region, ~28 kb in length, included the relaxaseICESan26-I, most T4SS genes, and other backbones, and it showed high identity with TnGBS. The third region was a resistance gene cluster comprising up to six ARGs and other uncharacterized ORFs. However, the internal element of ICESan26_rplL was not an ICE in the TnGBS family, because the integraseICESan26-I differed from the characteristic DDE transposase of TnGBS, which may have been altered during module exchanges. Hence, it was designated a TnGBS-like ICE in this study. These results suggested that ICESan26_rplL was a mosaic ICE with module combinations of ICEs in the ICESa2603 family and TnGBS-like ICEs.
 |
Figure 1 Comparison analysis of ICESan26_rplL with known ICEs. (a) comparison of ICESan26_rplL with ICESa2603, ICESsuYS388, ICESpy009, ICESp1116 and TnGBS2.6. (b) comparison of resistance genes cluster of ICESan26_rplL with plasmid pNUITMV6_2. Integrase genes are showed by blue arrows, relaxase by purple arrows, T4SSs genes by yellow arrows, antibiotics resistance genes by red arrows, putative ISs sequence by green arrows and others by white arrows. Areas shaded French grey represent regions of >60% identity. |
ICESan26_rplL carried resistance genes for multiple antibiotics that were clustered within a ~19 kb genetic region. A similar region was also observed in the multi-resistance plasmid pNUITMV6_2 (AP031235) from Enterococcus faecium, indicating that this resistance region of ICESan26_rplL may be derived from the same ancestor as that of this plasmid (Figure 1b).
Excision/Circular form of ICESan26_rplL and Internal TnGBS-Like ICEs
Reverse pair primers (P1-P4 and PI1-PI4) were used to detect the presence of the intermediate circularization form of ICESan26_rplL as well as internal TnGBS-like ICEs (Figure 2). The results showed that the PCR bands of the integration (P1/P2 and P3/P4), excision (P1/P4), and circulation (P2/P3) of ICESan26_rplL were positive, whereas the internal TnGBS-like ICE was positive for integration (PI1/PI2 and PI3/PI4), but negative for excision (PI1/PI4) and circulation (PI2/PI3). This indicated that a significant proportion of ICESan26_rplL could be excised from the chromosome and circularized. However, the TnGBS-like ICE only acted as a part of ICESan26_rplL rather than being excised independently.
 |
Figure 2 Diagram and PCR detection of integration and excision/circularization of ICESan26_rplL. Black rectangles represent TnGBS-like ICE within ICESan26_rplL, the white combinate black rectangles represent composite ICESan26_rplL, Primers used are indicated by black arrows. The bottom panel shows the PCR results and PCR marker is Vazyme DL2000 DNA. |
Cross-Species Transfer of ICESan26_rplL to S. agalactiae
In the conjugation transfer experiments, S. anginosus San26 (ERYR, CLIR, TETR, and LEVS) was used as the donor, and S. agalactiae SagR331 (ERYS, CLIS, TETR, and LEVR) was used as the recipient. The results showed that a transconjugant (S. agalactiae SagR331_TC) was obtained at a frequency of 2.13×10−8. The drug susceptibility test results showed that the transconjugant exhibited high-level resistance (>128 μg·mL−1) to erythromycin and clindamycin, indicating ARGs had been transferred to MLSB-sensitive recipients (Table 1). The whole-genome sequencing of the recipient and transconjugant confirmed that the ICESan26_rplL (61.618 kb) was integrated at the downstream of rplL (JBKOIP010000001, 35,600–96,868) (Figure 3).
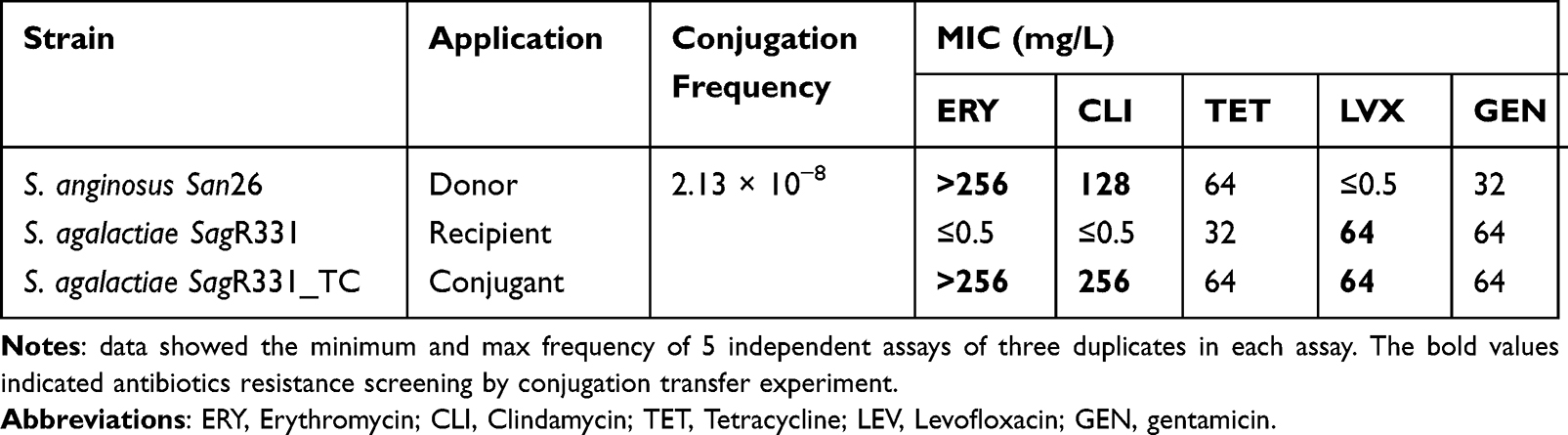 |
Table 1 Characteristics of the Donor, Recipient and Transconjugant |
 |
Figure 3 (a) Schematic of ICE conjugation transfer from donor to recipient and obtained transconjugant in cell perspective. (b) Alignment of genome drafts of recipient S. agalactiae SagR331 and transconjugant SagR331_TC. The different regions between transconjugant and recipient represents transferred ICESan26_rplL. |
Identification of ICESan26-Like ICEs in NCBI
To retrieve ICESan26-like ICEs from the available sequences in GenBank, the core functional genes of ICESan26_rplL were used as query terms for BLASTN. At the time of writing this manuscript, 53 putative ICEs related to ICESan26_rplL were identified in the available complete genome. Their host accession numbers, positions, and ARGs are listed in Table S2.
Among these 53 putative ICEs, three ICEs were highly consistent with ICESan26_rplL, and these ICEs were classified into the same ICE family. They had the same mosaic structures and identical ARG regions, and they were integrated at the downstream segment of rplL. They were found in the genomes of the anaerobic bacteria Streptococcus gallolyticus and Streptococcus pasteurianus (Figure S1). The other 50 ICEs were classified as ICESan26-related ICEs. Among the 50 ICEs, three ICEs were almost identical to TnGBS-like ICEs. These ICEs appeared to have randomly inserted sites, and they were identified in two S. pyogenes genomes and one S. anginosus genome. The remaining 47 putative ICEs were classified as the ICEs in the ICESa2603 family with various genetic characteristics and cargos. In addition to the above-mentioned ARGs, catQ (for chloramphenicol resistance), mel and mef(A) (macrolides), and aac(6ʹ)-aph(2ʹ) (aminoglycosides) were classified into the newly identified ICESa2603 family.
Discussion
According to a recent report, S. anginosus can transmit ARGs to major streptococcal and enterococcal pathogens through ICEs.20 Hence, S. anginosus is considered an important ARG reservoir. In general, there are fewer reports on the drug resistance and transmission of ARGs in S. anginosus, despite that S. anginosus has garnered significant attention owing to its increasing risk for gastric cancer.2 In this study, a novel ICE (ICESan26_rplL) carrying up to six ARGs was identified, and it conferred a host multi-resistance phenotype, including erm(B) for MLSB, tetM-tetL for tetracycline, lnu and lsa(E) for lincomycin, and ant(6)-I for aminoglycosides. ICESan26_rplL was integrated at the downstream segment of rplL by a tyrosine integrase, consistent with ICEs in the ICESa2603 family. However, different from ICEs in the ICESa2603 family, most backbones of ICESan26_rplL, especially T4SSs, are highly similar to those of the TnGBS family. In other words, ICESan26_rplL is a composite ICE with a mosaic structure, and the tyrosine integrase of the ICESa2603 family determines the position of ICESan26_rplL in the genome, whereas T4SS, consistent with the TnGBS family, enables the horizontal transfer of ICESan26_rpl.
Mosaic ICEs are known to be generated by a series of interaction events of mobile elements, such as accretion, imprecise excision, tandem array, and recombination.12,31,32 Therefore, ICESan26_rplL may originate from two accretion events and one recombination event. TnGBS and the plasmid pNUITMV6_2 were embedded and accreted within the ICEs in the ICESa2603 family, respectively, at the downstream segment of rplL. However, it is difficult for ICEs with a long sequence to maintain stabilization for a long time in the chromosome. Some similar sequences may become recombined, thus resulting in the deletion of some fragments during the accretion of ICEs to form the current relatively stable ICESan26_rplL. In this study, the circular intermediate forms of ICESan26_rplL and the internal TnGBS-like ICE were examined. However, the excision and circulation of the internal TnGBS-like ICE were not detected, suggesting that the TnGBS-like ICE lost its transfer activity and became a new conservative structure in ICESan26_rplL.
Most previous reports on ICEs have focused on three major streptococcal species, including S. agalactiae, S. pyogenes and S. pneumoniae, and the zoonotic S. suis.19,26 To our knowledge, this may be the first scientific attempt to demonstrate the transferability of ICESan26_rplL from S. anginosus to other species. Further, this may also be the first report on the ICE associated with ICESa2603 or TnGBS in S. anginosus. However, the conjugative transfer frequency of ICESan26_rplL (2.13×10−8) was much lower than that of previously reported ICEs, such as ICESpy009 (4.0×10−7~4.0×10−5),29 ICESsuYZDH1 (1.19×10−6~1.90×10−6),33 and ICESan95_hsdM (6.32×10−7~1.19×10−6).20 In contrast to other composite ICEs with significant mosaic structures, such as ICESagTR7 (3×10−9)34 in S. agalactiae and ICESsu32457/ICESa2603 (1.5×10−9)31 in S. suis, ICESan26_rplL was relatively more efficient, which may be attributed to its more complex structure and longer length, which affected its activity. The in silico analysis results revealed ICESan26-like ICEs in the anaerobic Streptococcus gallolyticus and Streptococcus pasteurianus. These results indicated that the presence of ICEs, similar to ICESan26_rplL, could benefit the dissemination of ARGs, thus increasing the risk of drug resistance in S. anginosus and displaying the potential to spread throughout the streptococcal groups. Given the cross-species transfer of ICEs, S. anginosus indirectly may serve as a reservoir and transit station for resistance genes of other streptococcal groups.
As revealed in previous studies, S. anginosus is often co-infected with other streptococci in the pleural cavity, vagina, and respiratory tract, but its role as a normal flora is often ignored.35–37 Our findings suggest that the cross-species transfer of ICESan26_rplL might be a new way for genes to undergo horizontal transfer between normal to major pathogenic bacteria. This may contribute to the resistance of susceptible pathogenic S. agalactiae during co-infection, thereby avoiding killing by antibiotics. According to a recent study, S. anginosus, similar to Helicobacter pylori, can also induce gastrointestinal cancer, which has attracted extensive concern.2 As a native ICE of S. anginosus, ICESan26_rplL may be an obstacle to the prevention and control of S. anginosus in the future due to its potential ability to transmit drug resistance.
However, ICESan26_rplL and highly similar ICEs appear to be less common in the genome compared with conventional ICEs, despite the fact that they confer more resistant phenotypes to the host. Moreover, the modules from different mobile elements are prone to inappropriate fitting, resulting in a decrease in the transfer frequency and limiting their further dissemination. Other reasons, such as multiple resistance genes or longer ICE fragments, may impose a heavy fitness cost, leading to a growth disadvantage for new host bacteria that cannot be excluded. Therefore, these considerations still need to be further investigated.
Conclusion
ICESan26_rplL, a composite ICE composed of modules in the ICESa2603 and TnGBS families, could mediate the dissemination of multiple ARGs from the commensal S. anginosus to the main pathogen S. agalactiae. The transfer of this ICE may represent a new method for the transmission of antimicrobial resistance among species. The unique structure of ICESan26_rplL implies potential interactions between different genetic elements, leading to the diverse evolution of ICEs.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
There is no funding to report.
Disclosure
The authors declare that there are no competing interests in this work.
References
1. Jensen A, Hoshino T, Kilian M. Taxonomy of the anginosus group of the genus streptococcus and description of Streptococcus anginosus subsp. whileyi subsp. nov. and streptococcus constellatus subsp. viborgensis subsp. nov. Int J Syst Evol Microbiol. 2013;63(Pt 7):2506–2519. doi:10.1099/ijs.0.043232-0
2. Fu K, Cheung AHK, Wong CC, et al. Streptococcus anginosus promotes gastric inflammation, atrophy, and tumorigenesis in mice. Cell. 2024;187(23):6783. doi:10.1016/j.cell.2024.10.036
3. Guo F, Li L, Li L. Streptococcus anginosus: a new pathogen of superficial gastritis, atrophic gastritis and gastric cancer. Biomol Biomed. 2024;24(5):1040–1043. doi:10.17305/bb.2024.10705
4. Le Doare K, Heath PT. An overview of global GBS epidemiology. Vaccine. 2013;31(Suppl 4):D7–12. doi:10.1016/j.vaccine.2013.01.009
5. Leclercq R. Mechanisms of resistance to macrolides and lincosamides: nature of the resistance elements and their clinical implications. Clin Infect Dis. 2002;34(4):482–492. doi:10.1086/324626
6. Achard A, Villers C, Pichereau V, Leclercq R. New lnu(C) gene conferring resistance to lincomycin by nucleotidylation in streptococcus agalactiae UCN36. Antimicrob Agents Chemother. 2005;49(7):2716–2719. doi:10.1128/AAC.49.7.2716-2719.2005
7. Roberts MC. Update on macrolide-lincosamide-streptogramin, ketolide, and oxazolidinone resistance genes. FEMS Microbiol Lett. 2008;282(2):147–159. doi:10.1111/j.1574-6968.2008.01145.x
8. Vaziri F, Peerayeh SN, Nejad QB, Farhadian A. The prevalence of aminoglycoside-modifying enzyme genes (aac (6’)-I, aac (6’)-II, ant (2”)-I, aph (3’)-VI) in Pseudomonas aeruginosa. Clinics. 2011;66(9):1519–1522. doi:10.1590/s1807-59322011000900002
9. Uruen C, Gimeno J, Sanz M, Fraile L, Marin CM, Arenas J. Invasive streptococcus suis isolated in Spain contain a highly promiscuous and dynamic resistome. Front Cell Infect Microbiol. 2023;13:1329632. doi:10.3389/fcimb.2023.1329632
10. Roberts AP, Mullany P. Tn916-like genetic elements: a diverse group of modular mobile elements conferring antibiotic resistance. FEMS Microbiol Rev. 2011;35(5):856–871. doi:10.1111/j.1574-6976.2011.00283.x
11. Warburton PJ, Amodeo N, Roberts AP. Mosaic tetracycline resistance genes encoding ribosomal protection proteins. J Antimicrob Chemother. 2016;71(12):3333–3339. doi:10.1093/jac/dkw304
12. Wozniak RA, Waldor MK. Integrative and conjugative elements: mosaic mobile genetic elements enabling dynamic lateral gene flow. Nat Rev Microbiol. 2010;8(8):552–563. doi:10.1038/nrmicro2382
13. Johnson CM, Grossman AD. Integrative and conjugative elements (ICEs): what they do and how they work. Annu Rev Genet. 2015;49(1):577–601. doi:10.1146/annurev-genet-112414-055018
14. Khan UB, Portal EAR, Sands K, et al. Genomic analysis reveals new integrative conjugal elements and transposons in GBS conferring antimicrobial resistance. Antibiotics. 2023;12(3). doi:10.3390/antibiotics12030544
15. Botelho J, Schulenburg H. The role of integrative and conjugative elements in antibiotic resistance evolution. Trends Microbiol. 2021;29(1):8–18. doi:10.1016/j.tim.2020.05.011
16. Zhou K, Xie L, Han L, Guo X, Wang Y, Sun J. ICESag37, a novel integrative and conjugative element carrying antimicrobial resistance genes and potential virulence factors in Streptococcus agalactiae. Front Microbiol. 2017;8:1921. doi:10.3389/fmicb.2017.01921
17. Huang J, Liang Y, Guo D, et al. Comparative genomic analysis of the ICESa2603 family ICEs and spread of erm(B)- and tet(O)-carrying transferable 89K-subtype ICEs in swine and bovine isolates in China. Front Microbiol. 2016;7:55. doi:10.3389/fmicb.2016.00055
18. Guerillot R, Da Cunha V, Sauvage E, Bouchier C, Glaser P. Modular evolution of TnGBSs, a new family of integrative and conjugative elements associating insertion sequence transposition, plasmid replication, and conjugation for their spreading. J Bacteriol. 2013;195(9):1979–1990. doi:10.1128/JB.01745-12
19. Huang J, Ma J, Shang K, et al. Evolution and diversity of the antimicrobial resistance associated mobilome in Streptococcus suis: a probable mobile genetic elements reservoir for other streptococci. Front Cell Infect Microbiol. 2016;6:118. doi:10.3389/fcimb.2016.00118
20. Yi S, Zhou K, Xu X. Characterization of erm(B)-carrying integrative and conjugative elements transferred from Streptococcus anginosus to other streptococci and enterococci. Microb Drug Resist. 2024;30(6):243–253. doi:10.1089/mdr.2023.0342
21. Prjibelski A, Antipov D, Meleshko D, Lapidus A, Korobeynikov A. Using SPAdes de novo assembler. Curr Protoc Bioinformatics. 2020;70(1):e102. doi:10.1002/cpbi.102
22. Brettin T, Davis JJ, Disz T, et al. RASTtk: a modular and extensible implementation of the RAST algorithm for building custom annotation pipelines and annotating batches of genomes. Sci Rep Feb. 2015;10(5):8365. doi:10.1038/srep08365
23. Darling AC, Mau B, Blattner FR, Perna NT. Mauve: multiple alignment of conserved genomic sequence with rearrangements. Genome Res. 2004;14(7):1394–1403. doi:10.1101/gr.2289704
24. Teatero S, Ramoutar E, McGeer A, et al. Clonal complex 17 group B Streptococcus strains causing invasive disease in neonates and adults originate from the same genetic pool. Sci Rep. 2016;6(1):20047. doi:10.1038/srep20047
25. Rahman M, Nguyen SV, McCullor KA, King CJ, Jorgensen JH, McShan WM. Complete genome sequence of Streptococcus anginosus J4211, a clinical isolate. Genome Announc. 2015;3(6). doi:10.1128/genomeA.01440-15
26. Ambroset C, Coluzzi C, Guedon G, et al. New insights into the classification and integration specificity of Streptococcus integrative conjugative elements through extensive genome exploration. Front Microbiol. 2015;6:1483. doi:10.3389/fmicb.2015.01483
27. Florensa AF, Kaas RS, Clausen P, Aytan-Aktug D, Aarestrup FM. ResFinder - an open online resource for identification of antimicrobial resistance genes in next-generation sequencing data and prediction of phenotypes from genotypes. Microb Genom. 2022;8(1). doi:10.1099/mgen.0.000748
28. Wang J, Qi K, Bai X, et al. Characterization of integrative and conjugative elements carrying antibiotic resistance genes of Streptococcus suis isolated in China. Front Microbiol. 2022;13:1074844. doi:10.3389/fmicb.2022.1074844
29. Del Grosso M, Camilli R, Rizzi E, Pietrelli A, De Bellis G, Pantosti A. ICESpy009, a conjugative genetic element carrying mef(E) in Streptococcus pyogenes. Antimicrob Agents Chemother. 2016;60(7):3906–3912. doi:10.1128/AAC.03082-15
30. Brenciani A, Tiberi E, Morici E, Oryasin E, Giovanetti E, Varaldo PE. ICESp1116, the genetic element responsible for erm(B)-mediated, inducible resistance to erythromycin in Streptococcus pyogenes. Antimicrob Agents Chemother. 2012;56(12):6425–6429. doi:10.1128/AAC.01494-12
31. Marini E, Palmieri C, Magi G, Facinelli B. Recombination between Streptococcus suis ICESsu32457 and Streptococcus agalactiae ICESa2603 yields a hybrid ICE transferable to Streptococcus pyogenes. Vet Microbiol. 2015;178(1–2):99–104. doi:10.1016/j.vetmic.2015.04.013
32. Pavlovic G, Burrus V, Gintz B, Decaris B, Guedon G. Evolution of genomic islands by deletion and tandem accretion by site-specific recombination: iCESt1-related elements from Streptococcus thermophilus. Microbiology. 2004;150(Pt 4):759–774. doi:10.1099/mic.0.26883-0
33. Yi S, Huang J, Hu X, et al. Nonconservative integration and diversity of a new family of integrative and conjugative elements associated with antibiotic resistance in zoonotic pathogen Streptococcus suis. Vet Microbiol. 2021;254:109009. doi:10.1016/j.vetmic.2021.109009
34. Mingoia M, Morici E, Marini E, Brenciani A, Giovanetti E, Varaldo PE. Macrolide resistance gene erm(TR) and erm(TR)-carrying genetic elements in Streptococcus agalactiae: characterization of ICESagTR7, a new composite element containing IMESp2907. J Antimicrob Chemother Mar. 2016;71(3):593–600. doi:10.1093/jac/dkv408
35. Kanellakis NI, Wrightson JM, Gerry S, et al. The bacteriology of pleural infection (TORPIDS): an exploratory metagenomics analysis through next generation sequencing. Lancet Microbe. 2022;3(4):e294–e302. doi:10.1016/S2666-5247(21)00327-X
36. Li H, Dong M, Xie W, et al. Mixed vaginitis in the third trimester of pregnancy is associated with adverse pregnancy outcomes: a cross-sectional study. Front Cell Infect Microbiol. 2022;12:798738. doi:10.3389/fcimb.2022.798738
37. McDonald HM, Chambers HM. Intrauterine infection and spontaneous midgestation abortion: is the spectrum of microorganisms similar to that in preterm labor? Infect Dis Obstet Gynecol. 2000;8(5–6):220–227. doi:10.1155/S1064744900000314
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms.php
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms.php
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.