")
Back to Journals » Clinical, Cosmetic and Investigational Dermatology » Volume 18
A Novel Immune-Related Three-Gene Signature and Immune Infiltration Insights in Psoriasis and Chronic Kidney Disease
Received 24 October 2024
Accepted for publication 16 January 2025
Published 25 January 2025 Volume 2025:18 Pages 267—286
DOI https://doi.org/10.2147/CCID.S499202
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Prof. Dr. Rungsima Wanitphakdeedecha
Tingting Yin, Tingting Zhang, Lei Ma
College of Life Science, Shihezi University, Shihezi, Xinjiang, People’s Republic of China
Correspondence: Tingting Zhang; Lei Ma, Email [email protected]; [email protected]
Purpose: There are significant inflammatory correlations and common immune dysregulation features between psoriasis and chronic kidney disease, however, the inflammatory mechanisms of these two diseases have not been clarified. The aim of this study was to screen immunologically related biomarkers for psoriasis and chronic kidney disease with the objective of identifying specific molecular markers to improve the accuracy and sensitivity of disease diagnosis.
Patients and Methods: To achieve this objective, common differentially expressed genes between psoriasis and chronic kidney disease were first identified. Through further functional analysis, these genes were found to be primarily involved in the activation of inflammation and innate immune responses. Subsequently, six hub genes were determined using five topological algorithms. The responses of these two diseases exhibited similar changes in immune reactions. By cross-analyzing these key genes with known immune genes, three Immunity-Related Hub Genes (IRHGs) were identified, including MX1, DDX58, and ISG20.
Results: ROC curve analysis validated the excellent discriminative ability of MX1 and ISG20 in both diseases. Furthermore, immune infiltration analysis revealed a higher abundance of T cells in samples from both psoriasis and chronic kidney disease, suggesting that T cell-driven immune responses may play a crucial role in the association of these two diseases. Lastly, single-cell analysis observed a significant increase in the cell abundance of T cells and endothelial cells in psoriasis and chronic kidney disease, respectively. The differential expression of MX1, DDX58, and ISG20 in these cells suggests that they may be involved to varying degrees in the pathogenic mechanisms of the two diseases.
Conclusion: This study provides a theoretical foundation for prognosis assessment and treatment of psoriasis and chronic kidney disease, contributing to a deeper understanding of the immune mechanisms underlying these conditions.
Keywords: psoriasis, chronic kidney disease, WGCNA, immune infiltration, single cells
Introduction
Psoriasis, as a chronic autoimmune disease, is characterized pathologically by an abnormal proliferation of the epidermis with an accompanying inflammatory response, clinically manifested as erythema with silvery-white scales.1 Its pathological mechanism involves an overreaction of the immune system to the skin tissue, leading to abnormal proliferation and keratinization of keratinocytes, and is closely associated with inflammatory cell infiltration, vasodilatation, and aberrant release of cytokines (IL-1, IL-6, TNF-α).2,3
Chronic Kidney Disease (CKD) is caused by a variety of factors that lead to chronic damage to renal structure and function, culminating in renal failure and high mortality.4 The disease is closely associated with abnormal immune activity, which may trigger glomerular inflammation, thereby affecting renal structure and function.5 Epidemiologic studies have shown that the prevalence of CKD significantly higher in patients with psoriasis than in the general population, suggesting that psoriasis may be an important risk factor for CKD.6
Recent studies have shown that the inflammatory response to psoriasis skin lesions can induce renal injury as a potential trigger for CKD.7 Specifically, the inflammatory response to psoriatic lesions may lead to elevated levels of inflammatory factors in serum and renal tissues. These factors may induce inflammation in renal tubular epithelial cells and mesangial cells, leading to renal dysfunction.8 In addition, the inflammatory response in psoriatic lesions may activate Toll-like receptors (TLRs) and MyD88 (myeloid differentiation factor 88) signaling pathways, promoting increased expression of NF-κB-related proteins (especially NF-κBp65) and decreased IκBα protein. These changes may promote further development of the inflammatory response and exacerbate renal injury.9
Despite the significant association between psoriasis and CKD, the underlying pathogenesis between the two is unclear. In this study, we used Weighted Gene Co-expression Network Analysis (WGCNA) combined with five Cytoscape algorithms with the aim of identifying hub genes associated with psoriasis and CKD. By comparing these genes with known immune genes, we identified three immunity-related hub genes (IRHGs). The diagnostic efficacy of these IRHGs was evaluated in the training and validation sets using Receiver Operating Characteristic (ROC) curves. Furthermore, CIBERSORT analysis was applied to explore the association between IRHGs and immune cells, and the results were validated at the single-cell transcriptome level, as shown in the flow in Figure 1. Through this study, a deeper understanding of the potential association between psoriasis and CKD was achieved, with the hope that it will provide new insights into the treatment of patients with the coexistence of psoriasis and CKD.
 |
Figure 1 The primary process for exploring Immunity-Related Hub Genes (IRHGs) in psoriasis and chronic kidney disease. |
Materials and Methods
Data Source
To obtain the raw gene expression profile data, we conducted searches in the GEO database (https://www.ncbi.nlm.nih.gov/geo/) using the keywords “psoriasis” and “chronic kidney disease”. Ultimately, we selected the GSE30999 and GSE109248 datasets as the training sets for psoriasis, including 102 non-lesional samples and 99 psoriatic lesion samples. Simultaneously, the GSE15072 and GSE66494 datasets were chosen as the training sets for CKD, comprising a total of 62 CKD samples and 8 healthy samples. To validate the robustness of the model, we utilized the GSE201827 and GSE70528 datasets as independent validation sets for psoriasis and CKD, respectively. Additionally, for single-cell analysis, we also chose the single-cell transcriptome datasets for psoriasis (GSE151177) and CKD (GSE199711). The detailed information of the research samples has been summarized in Supplementary Table 1.
Identification of Differentially Expressed Genes in Psoriasis and Chronic Kidney Disease (CKD)
In this study, the “limma” package in R was employed to identify differentially expressed genes (DEGs) in the psoriasis dataset and CKD dataset. Differentially expressed genes were selected using a threshold of P value < 0.05 and |log FC| > 1.5, and the results were visualized using volcano plots.
Identification and Pathway Enrichment of Common Genes
To investigate the biological functional categories and potential mechanisms of pathogenic genes in psoriasis and CKD, this study conducted Gene Ontology (GO)10 and Kyoto Encyclopedia of Genes and Genomes (KEGG) analyses. Pathways with a p-value < 0.05 were considered significant.
WGCNA Identifies Key Modules Associated With Psoriasis and Chronic Kidney Disease (CKD)
We utilized the “WGCNA” package in the R programming language to construct gene co-expression networks for psoriasis and CKD. Firstly, by utilizing the gene-gene correlation matrix and soft thresholding, we established an adjacency matrix to depict the levels of association between nodes. Subsequently, using gene hierarchical clustering, a dendrogram was generated to identify co-expressed modules. Finally, we employed hierarchical clustering and the dynamic tree-cutting function to detect modules, calculated the module eigengenes (ME) for each module, and explored the correlation between modules and patient clinical features through Pearson correlation analysis (P < 0.05).
Identification of Immunity-Related Hub Genes (IRHGs)
This study employed five different topological analysis methods to screen for important hub genes, including Degree, EPC, MNC, Radiality, and Closeness. For each method, we selected the top 10 genes with the highest scores as important hub genes.11 Subsequently, the candidate genes obtained from these five algorithms were intersected. Finally, these candidate genes were compared with known immune genes to identify three immunity-related hub genes (IRHGs) associated with the pathogenesis of psoriasis and CKD.
Expression Level and Diagnostic Value of IRHGs
We extracted the gene expression profiles of IRHGs from the psoriasis dataset (GSE30999) and CKD dataset (GSE66494). Using the ggplot2 package in R, we created boxplots to observe the expression levels of these genes across different samples (P < 0.05). To validate the results, GSE201827 (comprising 44 psoriasis samples and 49 control samples) and GSE70528 (comprising 7 CKD samples and 8 control samples) were used as validation sets to predict the performance of IRHGs. Receiver Operating Characteristic (ROC) curves were drawn, and Area Under the Curve (AUC) values were calculated using the “pROC” package in R. Typically, an AUC greater than 0.7 is considered to indicate relatively good predictive performance, suggesting that IRHGs can effectively discriminate between patients and non-patients.
Evaluation of Immune Cell Infiltration and Its Correlation With IRHGs
In this study, we investigated the relationship between 22 immune cells and IRHGs by Spearman correlation analysis using the CIBERSORT tool.12 The R package “corrplot” was employed to visualize the expression matrix of Spearman correlation coefficients among 22 immune cells in the datasets of psoriasis and CKD. Additionally, the study utilized R packages such as “reshape2”, “ggpubr”, “ggExtra”, and “ggstatsplot” to calculate the correlation between the expression of three IRHGs and immune cells. The results were visualized using the “ggplot2” package.
Candidate Small Molecular Compounds Prediction
IRHGs were searched against the Connectivity Map database (https://clue.io) to identify potential small molecule compounds for the treatment of psoriasis combined with CKD. Ultimately, the 10 compounds with the highest score were identified.
GSEA Analysis
We conducted Gene Set Enrichment Analysis (GSEA) on the expression levels of each IRHGs using the R package “clusterProfiler”. For this purpose, we utilized gene sets from MSigDB (https://www.gsea-msigdb.org/gsea/msigdb) and considered a standardized enrichment score with p < 0.05 as the significance threshold.13 Through these criteria, we validated the association between the expression of IRHGs and signaling pathways in both psoriasis and CKD.
Single Cell Analysis
This study analyzed single-cell RNA sequencing data from psoriasis and CKD using the “Seurat” package.14 First, low quality or dead cells were excluded by setting a screening threshold (min.cells=3, min.features=250). Subsequently, the data were normalized using the LogNormalize method and unsupervised clustering was achieved by principal component analysis (PCA) and UMAP to visualize the cell population distribution on a 2D map.15 Next, cells were annotated using SingleR,16 and differentially expressed genes in different cell populations were screened using the “FindAllMarkers” function. Finally, the “Monocle” package was used to simulate the dynamics of temporal development based on key gene expression patterns.
Statistical Analysis
All statistical analyses were conducted using R software (v4.2.2), with all images generated in R Studio. Statistical tests, including Student’s t test, Wilcoxon rank-sum test, and Kruskal–Wallis test were used where indicated. P-values > 0.05 were considered not statistically significant (n.s)., while P-values ≤ 0.05 were categorized as follows: *p ≤ 0.05, **p ≤ 0.01, ***p ≤ 0.001, and ****p ≤ 0.0001. For multiple-hypothesis testing in GSEA analysis, p-values were adjusted using the false discovery rate (FDR).
Results
Identification of DEGs in Psoriasis and Chronic Kidney Disease (CKD)
By comparing the transcriptomic data between psoriasis and control samples, we identified a total of 488 differentially expressed genes, comprising 351 upregulated and 137 downregulated genes (Figure 2A). Simultaneously, upon comparing the transcriptomic data of CKD samples with control samples, we identified 154 differentially expressed genes, including 74 upregulated and 80 downregulated genes (Figure 2B).
 |
Figure 2 Identification of common differentially expressed genes. (A) Volcano plot of differentially expressed genes (DEGs) in psoriasis. (B) Volcano plot of DEGs in chronic kidney disease. (C) Heat map illustrating DEGs in psoriasis (n=40). (D) Heat map illustrating DEGs in chronic kidney disease (n=40). (E) Differentially expressed genes common to psoriasis and chronic kidney disease. |
Subsequently, we filtered the top 20 upregulated genes and the top 20 downregulated genes in both psoriasis and CKD datasets (Supplementary Table 2). The results indicated that differentially expressed genes effectively distinguished between the two groups (Figure 2C and D). Furthermore, we observed 13 commonly differentially expressed genes in both psoriasis and CKD datasets (Figure 2E), suggesting potential similarities in their pathogenic mechanisms.
Function of Common DEGs
The common DEGs in psoriasis and CKD samples involve several immune reactions and immune pathways, which may be regulated differently during disease progression. They are involved in various biological processes (eg, positive regulation of toll-like receptor signaling pathway, innate immune response, and negative regulation of viral process), cellular components (eg, inflammasome complex, secretory granule lumen, and cytoplasmic vesicle lumen), and KEGG pathways (eg, Influenza A, Renin−angiotensin system, Terpenoid backbone biosynthesis) (Figure 3A and B). These findings suggest that inflammatory pathways and immune activation play crucial roles in the pathogenesis of psoriasis and CKD.
 |
Figure 3 Functional enrichment analysis of common DEGs. (A) Results of GO enrichment analysis of common DEGs in psoriasis and chronic kidney disease. (B) Results of KEGG enrichment analysis of common DEGs in psoriasis and chronic kidney disease. |
WGCNA Identifies Co-Expression Modules for Psoriasis and Chronic Kidney Disease (CKD)
We employed WGCNA to explore co-expressed gene modules and networks in psoriasis and CKD. When processing the psoriasis dataset, a soft threshold (β) of 10 was selected, while for the CKD dataset, the chosen β value was 13 (Figure 4A and D). In constructing the co-expression network for psoriasis samples, thirteen modules were successfully identified (Figure 4B and C). When constructing the co-expression network for CKD samples, eleven modules were identified (Figure 4E and F).
 |
Figure 4 Identification of modules associated with clinical features of psoriasis and chronic kidney disease by WGCNA. (A and D) Based on the results of the scale-free topological model and the average connectivity, the soft threshold β = 10 was chosen in the psoriasis dataset; in the chronic kidney disease dataset, the soft threshold was chosen to be 13. (B and E) Clustering tree of gene expression levels in psoriasis and chronic kidney disease, the upper panel shows the clustering relationship of genes at the transcriptional level, and the lower panel illustrates the different cluster clusters with different colors. (C and F) Pearson correlation analysis of each module (clustering clusters in (B) and (E) with different psoriasis clinical samples and chronic kidney disease samples. Horizontal is the first principal component value of the network module. Longitudinal is the trait data of the samples. In the heatmap table, the correlation and significance (p_value) between the modules and the different traits of psoriasis and chronic kidney disease, respectively, are indicated. (G) Venn diagrams showing the intersection of key module genes in psoriasis and chronic kidney disease. |
Through the correlation analysis between modules and clinical phenotypes, it was observed that in the psoriasis dataset, the red module exhibited the strongest positive correlation with psoriasis samples (LS) (r=0.88, P<0.05, Figure 4C), indicating a close association between the genes in the red module and the occurrence and development of psoriasis. In the CKD dataset, the greenyellow module showed the strongest positive correlation with CKD samples (CKD) (r=0.67, P<0.05, Figure 4F), suggesting a close association between the genes in the greenyellow module and the progression of CKD. Consequently, genes in the red and greenyellow modules may play a critical role in the progression of psoriasis and CKD.
Identification of Immunity-Related Hub Genes (IRHGs)
In this study, the intersection of key module genes of psoriasis and CKD was mapped by a Venn diagram, and a total of 69 crossover genes were obtained (Figure 4G), which were defined as PCGs (Psoriasis and CKD Gene set). Furthermore, we found that immune-related genes in PCGs may play a critical role in the development of psoriasis and CKD. To explored the role of these genes, we comprehensively assessed the importance of PCGs in the network through five pivotal metrics (Degree, EPC, MNC, Radiality, and Closeness). Among these genes, six genes (DDX58, ISG20, MX1, CD47, POLI, and PRIM1), showed high node centrality (Figure 5A) in Degree, EPC, MNC, Radiality, and Closeness (Supplementary Table 3), implying that these genes are in a critical position in the network and therefore identified as core targets in psoriasis and CKD. Upon searching the Immunology Database, MX1, DDX58, and ISG20 were the immunity-related hub genes (IRHGs) for PCGs (Figure 5B).
 |
Figure 5 Identification of IRHGs. (A) Identification of 6 hub genes using 5 centrality metrics in cytoHubba. The outermost circles in the figure represent the screened hub genes that scored high on each metric, thereby being considered centrally or critically important in the biological network. (B) The hub gene set for psoriasis and chronic kidney disease was analyzed in conjunction with the immune gene set to identify three IRHGs associated with psoriasis and chronic kidney disease. |
Expression Level and Diagnostic Value of IRHGs
Figure 6A and B illustrate the expression levels of three immunity-related hub genes (IRHGs), which were upregulated in both psoriasis and CKD samples compared to controls. The sensitivity and specificity of these IRHGs were further evaluated. In the psoriasis dataset, MX1 (AUC=0.969), DDX58 (AUC=0.870), and ISG20 (AUC=0.944) demonstrated superior diagnostic values (Figure 6C). Similarly, in the CKD dataset, MX1 (AUC=0.972), DDX58 (AUC=0.958), and ISG20 (AUC=0.833) exhibited high diagnostic value (Figure 6D). Subsequently, external validation of the aforementioned IRHGs was conducted in psoriasis (GSE201827) and CKD datasets (GSE70528), revealing their effective predictive performance in both datasets (Figure 6E and F).
 |
Figure 6 Validation of Immunity-Related Hub Genes (IRHGs). (A and B) Comparative analysis of the expression levels of IRHGs in the psoriasis dataset and the chronic kidney disease dataset using the Wilcoxon rank-sum test. Box line plots illustrate the distribution of expression levels across samples, presenting medians, quartiles, and outliers in the data. (C) ROC curves for common diagnostic genes in the psoriasis dataset. (D) ROC curves for common diagnostic genes in the chronic kidney disease dataset. (E) ROC curves for common diagnostic genes in the psoriasis validation set (GSE201827). (F) ROC curves for common diagnostic genes in the chronic kidney disease validation set (GSE70528). |
Immune Cell Infiltration Analyses and Their Relationship With IRHGs
To explore the immune landscapes of psoriasis and CKD, we assessed the proportions of 22 immune cells in the psoriasis and CKD datasets. The abundance of dendritic cells activated, CD4 memory T cells, resting dendritic cells, and M1 macrophages was significantly increased in psoriasis samples compared with controls (Figure 7A). In CKD samples, the abundance of naïve B cells, M0 macrophages, and CD4 memory T cells was significantly increased (Figure 7C).
 |
Figure 7 Immune infiltration analysis of 3 Immunity-Related Hub Genes (IRHGs). (A and C) Differences in the proportion of immune cells between the control (con) and psoriasis groups (treat) and between the control (con) and chronic kidney disease groups (treat) were compared within subgroups using the Wilcoxon rank-sum test. The horizontal axis represents 22 immune cells. “Fraction value” indicates the percentage of a particular type of immune cell in disease samples (blue markers) and control samples (pink markers), where *, **, ***, and **** denote P<0.05, 0.01, 0.001, and 0.0001, respectively. (B) In the psoriasis dataset, the Spearman correlation between MX1, DDX58, ISG20, and the 22 immune cell types is calculated. (D) In the chronic kidney disease dataset, the Spearman correlation between MX1, DDX58, ISG20, and the 22 immune cell types is calculated. The colors in the plot represent the strength of the correlation. |
The present study further evaluated the relationship between the three IRHGs and immune cells. Specifically, in psoriasis samples, the expression levels of MX1, DDX58, and ISG20 were mainly positively correlated with the infiltration of T follicular helper cells, CD4 memory T cells, γΔT cells, and dendritic cells, and negatively correlated with the infiltration of M2 macrophages, activated NK cells, and mast cells resting (Figure 7B). In addition, in CKD samples, the expression levels of DDX58 and ISG20 were mainly positively correlated with the infiltration of M0, M1 macrophages, γΔT cells and CD4 memory T cells activated and negatively correlated with the infiltration of activated NK cells, Monocytes, and mast cells resting (Figure 7D). MX1 is positively associated with the infiltration of resting CD4 memory T cells and regulatory T cells, while it is negatively correlated with M1 macrophages and dendritic cells.
Drug Sensitivity Analysis of Candidate Markers
To investigate the potential therapeutic effects of small-molecule drugs in patients with psoriasis complicated by CKD, we utilized the Connectivity Map (CMap) database to identify 10 compounds (Supplementary Table 4). Specifically, in the MCF7 cell line, we observed a significantly positive correlation between the gene expression patterns induced by Imidazoline receptor ligands and the expression patterns of ISG20 and DDX58. This suggests a potential association between ISG20 and DDX58 with Imidazoline receptor ligands in this cell line. Furthermore, in the HAE1 and PC3 cell lines, we observed a positive correlation between the expression patterns of ISG20 and MX1 and the gene expression induced by the Protein kinase C activator. This implies that these genes may be involved in the PKC signaling pathway or biological processes related to PKC in these two cell lines. These findings provide new insights into exploring therapeutic approaches for psoriasis in combination with CKD, emphasizing the potential therapeutic value of these small molecule compounds in future treatment strategies. However, further research and clinical experiments will be required to validate their efficacy and safety.
GSEA Identifies IRHGs Related Signaling Pathways
To explore the pathways associated with the three IRHGs in psoriasis and CKD, we performed GSEA functional enrichment analysis. The results showed that in the psoriasis dataset, upregulation of the RIG-I-like receptor signaling pathway and the IL-17 signaling pathway were significantly associated with high expression of MX1, DDX58, and ISG20 (Supplementary Figure 1A). Whereas, in the CKD dataset, the activation of Allograft rejection and Ascorbate and aldarate metabolism pathways was significantly associated with high expression of MX1, DDX58, and ISG20 (Supplementary Figure 1B). The above analyses suggest that these pathways may have played a significant role in the progression of these two diseases.
Identification of Different Cellular Subpopulations in Psoriasis and Chronic Kidney Disease (CKD)
We conducted a single-cell transcriptome analysis of psoriasis and CKD, revealing the complexity of cellular populations in both diseases. Specifically, we annotated the single-cell data for psoriasis and identified three cell types, including keratinocytes expressing IL-22, IL-1β, and INF-γ, T cells, and Dendritic cells (Figure 8B). Furthermore, this study found that in psoriasis samples, there was a higher proportion of keratinocytes expressing IL-22, dendritic cells, and T cells, indicating that these cells likely participated in the occurrence and development of psoriasis (Figure 8A). Additionally, we further explored three IRHGs to understand their relationship with the aforementioned three cell subgroups. The results showed higher levels of ISG20 expression in these three cellular subpopulations, suggesting that ISG20 may be involved in the development of psoriasis in these cellular subpopulation settings (Figure 8C).
 |
Figure 8 Three cell subpopulations were identified based on psoriasis single-cell RNA-seq data. (A and B) illustrates the distribution of different cell populations in psoriasis samples (Ps1, 2) and control samples (C1, 2). The horizontal axis (UMAP_1) represents the first principal feature in the low-dimensional space after dimensionality reduction, while the vertical axis (UMAP_2) represents the second principal feature in the low-dimensional space after dimensionality reduction. (C) Depicts the expression level distribution of MX1, DDX58, and ISG20 in three cell subpopulations. |
This study inferred the developmental trajectory of cells based on the dynamic changes in gene expression in psoriasis single-cell data and analyzed the heterogeneity of immune cells among patients. In the cell trajectory, keratinocytes initially expressing IL1β gradually transitioned to expressing IL-22 in keratinocytes. Subsequently, once the state of expressing IL-22 in keratinocytes was established, these cells quickly differentiated into two distinct branches, representing two major cell lineages in the late stages of reprogramming. Specifically, one branch differentiated into keratinocytes expressing IFN-γ, while the other branch differentiated into dendritic cells and T cells, ultimately developing towards T cell differentiation (Figure 9).
 |
Figure 9 Cell trajectory analysis of three psoriasis cell subpopulations with distinct differentiation patterns. The trajectory analysis reveals that the three subpopulations of psoriasis cells exhibit distinct differentiation patterns. One branch differentiates into a subpopulation of T cell, while another branch is dominated by keratinocytes expressing interferon IFN-γ. The left panel illustrates the differentiation process of different cell subpopulations, with different colors representing various immune cells. The lower right panel displays three states of cells during the trajectory differentiation process, each represented by a different color. The upper right panel presents a pseudotime plot of cell trajectory differentiation, arranging cells in the order of their differentiation or development based on time information from single-cell data. The color depth in the plot reflects the duration of the differentiation process, indicating the length of time for each differentiation state. |
In the single-cell data of CKD, we identified five cell subgroups, including B cells, endothelial cells, induced pluripotent stem cells, tissue stem cells, and neuronal cells (Figure 10B). Compared to the control group, we observed a higher proportion of endothelial cells, B cells, and induced pluripotent stem cells in CKD samples, suggesting their potential involvement in the occurrence and development of CKD (Figure 10A). Furthermore, this study investigated the expression of three IRHGs in these five cell subgroups. The results revealed that, among these five cell subgroups, MX1 and DDX58 exhibited higher expression levels, suggesting that these two genes may play a crucial role in the occurrence and development of CKD in specific biological processes (Figure 10C).
 |
Figure 10 Five cell subpopulations with different annotations were identified based on single-cell RNA-seq data from chronic kidney disease. (A and B) depict the distribution of different cell populations in chronic kidney disease samples (CKD1, 2) and control samples (C1, 2). The horizontal axis (UMAP_1) represents the first principal feature in the low-dimensional space after dimensionality reduction, while the vertical axis (UMAP_2) represents the second principal feature in the low-dimensional space after dimensionality reduction. (C) illustrates the expression level distribution of MX1, DDX58, and ISG20 in the five cell subpopulations. |
Subsequently, we employed five immune cell types associated with CKD to construct a transcriptional trajectory to analyze the heterogeneity of different immune cells. In the cell trajectory, we observed that B cells and tissue stem cells were situated in different branches, exhibiting distinct differentiation states. In contrast, lymphatic-associated endothelial cells and B cells were initially in the same branch during differentiation, showing relatively similar differentiation states. In addition, the present study found that most lymphoid-related endothelial cells may develop toward two branches, vascular factor-related endothelial cells, and induced pluripotent stem cells, and eventually differentiate into vascular factor-related endothelial cells (Figure 11).
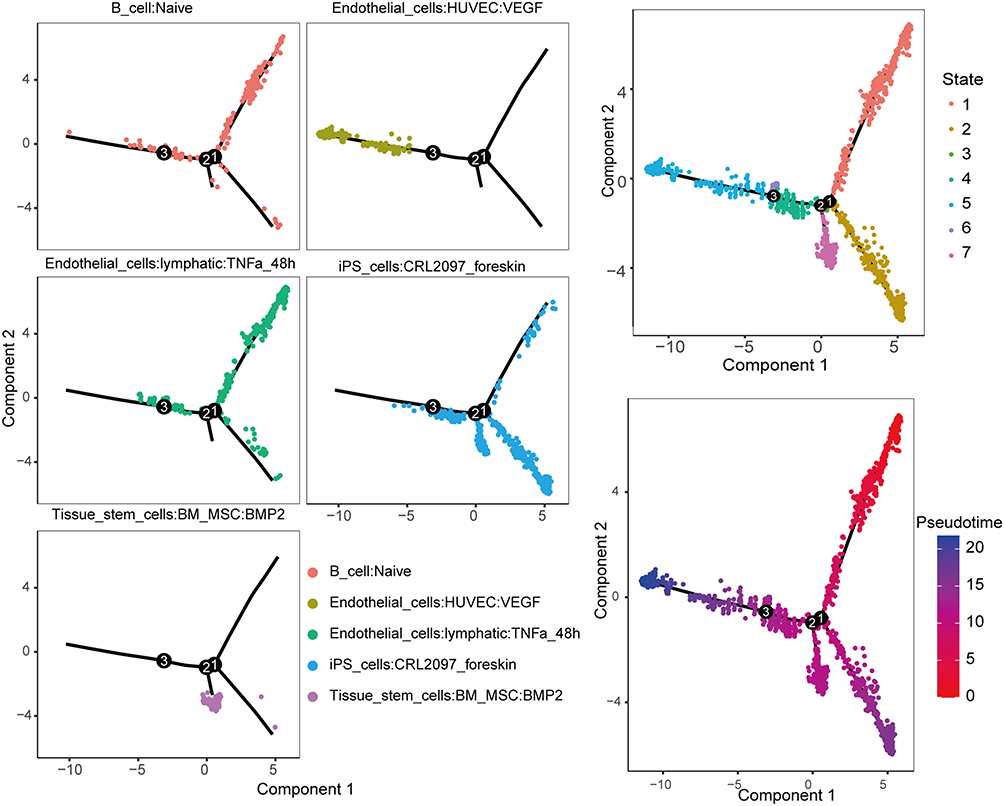 |
Figure 11 Cell trajectory analysis of five chronic kidney disease cell subpopulations with distinct differentiation patterns. The trajectory analysis reveals that the five subpopulations of chronic kidney disease cells exhibit different differentiation patterns. One branch differentiates into a subpopulation primarily composed of endothelial cells associated with blood vessels, while another branch is dominated by induced pluripotent stem cells. The left panel illustrates the differentiation process of different cell subpopulations, with different colors representing various immune cells. The upper right panel displays seven states of cells during the trajectory differentiation process, each represented by a different color. The lower right panel presents a pseudotime plot of cell trajectory differentiation, arranging cells in the order of their differentiation or development based on time information from single-cell data. The color depth in the plot reflects the duration of the differentiation process, indicating the length of time for each differentiation state. |
Discussion
Psoriasis, as a systemic disease, can affect multiple organs and systems, including the kidneys. The condition manifests with diverse symptoms such as proteinuria, hypertension, hematuria, and renal dysfunction.17 However, the mechanisms underlying the association between psoriasis and kidney disease are not fully understood. To further enhance therapeutic and management strategies, our focus was on investigating potential regulatory targets related to the immune system in psoriasis and CKD. In this study, we screened three immunity-related hub genes that are common in psoriasis and CKD and validated the results with single-cell transcriptomic data.
In-Depth Study of the Immune Regulation Mechanisms in Psoriasis and Chronic Kidney Disease (CKD): Association With Potential Targets and Environmental Factors
Environmental factors may play a crucial role in the onset and progression of psoriasis and CKD, exerting their influence through various aspects, including inflammation and immune dysregulation.18 Studies have indicated that PM2.5 may directly impact the kidneys through multiple pathways, including triggering abnormal activation of the renin-angiotensin system and deposition of immune complexes.19,20 In an inflammatory state, the accumulation of PM2.5 may exacerbate the activation of inflammatory microvesicles, causing damage to endothelial cells and disrupting their function. This endothelial dysfunction may further affect renal microcirculation, leading to kidney function issues.21–24 Simultaneously, PM2.5 can also affect the function of epidermal proteins, triggering skin dysfunction and failure. Specifically, PM2.5 can induce the activation of inflammatory microvesicles, promoting skin damage.25 This aligns with previous research findings, suggesting that the imbalance of cytokines associated with inflammatory microvesicles may lead to excessive infiltration of immune cells, thereby driving the sustained development of skin inflammation.26 Furthermore, the onset and progression of psoriasis are related to changes in the activity of inflammatory microvesicles.27 In this study, differentially expressed genes were significantly enriched in the inflammatory microvesicle complex, further supporting this perspective.
MX1 (myxovirus resistance protein 1) is a type I interferon-induced antiviral protein with multiple functions such as antiviral, modulation of interferon signaling pathway and immune response.28 Studies have shown that MX1 expression is significantly upregulated in the skin of psoriatic lesions, which may be associated with viral infection or immune abnormalities. Although its specific pathway of action is unclear, there is evidence that MX1 is induced by IL-29, which activates the type 1 interferon signaling pathway, which in turn triggers an inflammatory response, including the production of pro-inflammatory factors and chemokines, attracting and activating immune cells to form an inflammatory cascade response.29–31 This suggests that the role of MX1 in psoriasis is closely related to its antiviral capacity and inflammatory regulation. Meanwhile, in chronic kidney disease (CKD), elevated levels of MX1 correlate with INF-λ activation, showing its immunomodulatory role in CKD.32 In addition, MX1 may protect the kidneys from damage brought about by viral infection through antiviral effects32 and influence CKD progression by regulating immune cell activity or participating in renal tissue repair processes. Thus, MX1, as a key crosstalk gene, is involved in inflammatory regulation through multiple pathways in psoriasis and CKD, which is important for the pathomechanism of both diseases.
ISG20 is an RNA nucleic acid exonuclease that plays an important role in antiviral responses, DNA repair, and a variety of physiopathologic processes.33 Specifically, ISG20 expression is closely associated with infiltration of monocyte-derived macrophages and neutrophils, and its overexpression may suppress adaptive immune responses.33 In renal fibrosis studies, ISG20 expression was significantly elevated, and in vitro experiments demonstrated that knockdown of ISG20 inhibited the expression of the fibrotic protein HK-2, thereby slowing the process of renal fibrosis.34 In addition, ISG20 is involved in the antiviral defense of the kidney and may attenuate renal damage by regulating the DNA repair mechanism of renal cells.35 In psoriasis, ISG20 interacts with a variety of genes in keratinocytes and may promote the onset and progression of psoriasis,36 and although its direct role in psoriasis has not been adequately investigated, considering that psoriasis is closely associated with immune-mediated inflammatory responses, ISG20 may indirectly influence psoriasis pathogenesis by regulating immune cell activity or antiviral responses.37 Thus, ISG20 may play diverse functions in different physiologic and pathologic processes, providing important clues to understanding its specific role in psoriasis and CKD.
DDX58 (retinoic acid-inducible gene I, also known as RIG-I) acts as a recognition receptor for intracellular viral infections and initiates antiviral signaling pathways. In psoriasis, DDX58 is not only a susceptibility gene, but its upregulation is closely associated with disease pathogenesis, as evidenced by increased expression of RIG-I in damaged skin of patients, which decreases with remission. This may be related to abnormalities in the antiviral signaling pathway in psoriasis susceptibility gene carriers, leading to overexpression of DDX58 and downstream inflammatory factors such as IL-23.38–40 Similarly, although less studied in chronic kidney disease (CKD), activation of RIG-I signaling in peripheral blood mononuclear cells from patients with CKD stage V suggests its possible involvement in CKD development. DDX58 may indirectly influence CKD progression by protecting the kidneys from damage caused by viral infection or by participating in the process of immune regulation and renal tissue repair.41 Single gene GSEA analysis confirmed that the RIG-I-like receptor pathway was associated with high expression of DDX58, highlighting the importance of DDX58 in immunomodulation of psoriasis and CKD. Thus, DDX58 acts as a key crosstalk gene influencing the onset and progression of both diseases through different mechanisms.
Finally, we screened 10 potential compounds through the Connectivity map (CMap) database, among which protein kinase C (PKC) activators showed potential value in the treatment of psoriasis combined with chronic kidney disease (CKD). This compound may play a dual role in ameliorating the pathological processes of psoriasis and CKD by regulating cell proliferation and signaling.42,43 In psoriasis, PKC activators are able to inhibit the abnormal proliferation of keratinocytes, thereby alleviating epidermal thickening;44 whereas in CKD, it may exert positive anti-apoptotic and anti-oxidative stress effects through the selective activation of specific PKC isoforms.45,46 However, the effects of such analogs are dualistic, and non-selective activation of PKC activators may exacerbate inflammation or tissue damage.43 Therefore, future studies should focus on exploring the specific mechanism of action of PKC activators in the pathology of psoriasis and CKD and clarifying the regulatory pathways of their biological effects, so as to provide patients with more precise and effective treatment strategies.
Exploring the Immunological Relationship Between Psoriasis and Chronic Kidney Disease (CKD): Deciphering Gene Expression, Cellular States, and Potential Mechanisms
This study preliminarily explored the potential immune relationships between psoriasis and CKD, revealing significant differences in the immune patterns of the psoriasis group and CKD group compared to the control group, with a particularly pronounced increase in T cells. This finding suggests that in both diseases, IRHGs play a crucial role in regulating the immune microenvironment, enabling the immune system to respond more organizedly to various conditions.
Further research indicated a close association between the development of psoriasis and abnormal activation of T cells, involving multiple crucial aspects. Firstly, activated T cells triggered sustained inflammatory reactions in the lesional areas of psoriasis, releasing various inflammatory mediators, including cytokines and chemokines, promoting the formation of inflammation, and attracting other immune cells to accumulate in that area.47 Secondly, activated T cells, through the release of cell-proliferation-promoting cytokines such as interferon-gamma (IFN-γ) and tumor necrosis factor-alpha (TNF-α), drove the abnormal proliferation of keratinocytes, a major factor leading to the formation of psoriatic plaques in psoriasis-affected skin.48 Simultaneously, the dysregulation of T cell activity in psoriasis disrupted the balance of the immune system, triggering autoimmune responses that not only initiated lesion formation but also sustained its progression.49
Simultaneously, we observed that the abnormal activation of T cells associated with CKD also involved similar mechanisms. In CKD, the aberrant activation of T cells leads to an immune system imbalance and exacerbated inflammatory reactions.50 This process played a critical role in the progression of CKD by inducing inflammation in renal tissues through the production of inflammatory mediators, promoting disease development. Additionally, the balance of T cells was disrupted, triggering immune dysregulation and autoimmune responses, worsening kidney damage.51,52 Furthermore, T cells also participated in regulating the fibrotic process in renal tissues, a typical characteristic of CKD.53 In summary, there are similar immune mechanisms between psoriasis and CKD, including the abnormal activation of T cells, immune dysregulation, inflammatory reactions, and damage and repair of tissues.
In single-cell RNA sequencing (scRNA-seq) profiling of psoriasis, we identified three immune cell types, including keratinocytes, dendritic cells, and T cells. In further investigations, we delved the expression of IRHGs in these three immune cell types. The results indicated that ISG20 was expressed to varying degrees in all three cell types, particularly significantly in keratinocytes. A previous study analyzed the gene expression pattern of keratinocytes in psoriasis samples and found that ISG20 was highly expressed in keratinocytes,35 which is consistent with our research. In the scRNA-seq profile of CKD, we observed that endothelial cells exhibited the highest expression in CKD samples. Previous studies have noted variable reductions in brachial artery diameter in CKD patients, indicating endothelial dysfunction.54 Endothelial dysfunction may be associated with increased oxidative stress, leukocyte adhesion, and chronic inflammation, playing a crucial role in the pathogenesis of CKD.55
Additionally, we analyzed the differentiation trajectories of cell subpopulations in psoriasis and CKD, revealing that various cell types were not confined to a single state. Instead, each state encompassed diverse cell types. This diversity was also reflected in gene expression, with cells showing various gene combinations across different states. During cellular state transitions, some genes may be suppressed, while others may be reactivated to fulfill their functions. Our study found that MX1, DDX58, and ISG20 exhibited varying degrees of expression in the trajectory differentiation processes of psoriasis and CKD (Supplementary Figure 2). Therefore, MX1, DDX58, and ISG20 are likely closely related to the cellular composition and states in the immune microenvironment of psoriasis and CKD. In this study, we explored the immunity-related hub genes and potential connections between psoriasis and CKD, providing additional insights into unraveling the underlying mechanisms of these two diseases. However, our study has some limitations, including the use of computational methods and potential sample biases. Furthermore, in the ROC curve analysis of the validation sets for psoriasis and CKD, we observed AUC values of 0.753 and 0.732 for DDX58, indicating certain limitations as a potential immune biomarker for these two diseases.
Specifically, insufficient sample size or sample heterogeneity may have affected the performance of DDX58 in the validation set. Also, other biological and environmental factors that were not considered may have interfered with the performance of DDX58, resulting in a decrease in its sensitivity to differentiate between diseased and non-diseased individuals. In addition, data on age, gender, medication use, and comorbidities of the patients in the sample were not considered in this study, and these data may have influenced the current results. To improve the reliability of our findings, future studies will focus on expanding the sample size and selecting more representative sample sources.
Conclusions
In this study, we successfully identified a new set of immune marker genes and their associated immune infiltration patterns based on transcriptomic and single-cell data of psoriasis and CKD using multiple bioinformatics algorithms. These findings provide potential predictors for the treatment of psoriasis combined with CKD and provide new ideas for the future development of targeted therapies for psoriasis combined with CKD. Meanwhile, our study reveals the important role of T cell-mediated immune responses in the pathophysiologic process of psoriasis and CKD, providing new insights into understanding the mechanisms of co-morbidity between these two diseases.
Data Sharing Statement
The data underlying this article are available in the article and its online Supplementary Material.
Ethical Approval and Informed Consent
The datasets used in this study (GSE30999, GSE109248, GSE201827, GSE66494, GSE15072, GSE70528, GSE151177, GSE199711) were obtained directly from the GEO public data platform (https://www.ncbi.nlm.nih.gov/). GEO is a public database storing ethically approved patient data, open to researchers to support research and publication of results. According to Article 32 (1) of the Measures for Ethical Review of Life Science and Medical Research Involving Human Beings, promulgated in China on February 18, 2023, “research that uses legally obtained public data or data generated through observation and does not interfere with the behavior of the public” is exempt from ethical review. This study followed this principle, as all data were obtained legally and the research process did not interfere with public behavior, thus meeting the requirements for exemption from ethical review.
Acknowledgments
We thank the anonymous editor and reviewers for helpful suggestions on the manuscript and all the contributors to the data sets. We also thank numerous members of our lab for assistance. This paper has been uploaded to Research Square as a preprint: https://www.researchsquare.com/article/rs-3299710/v1.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
The National Natural Science Foundation of China (32060300 32060145 and 31860308), the China Scholarship Council, and the Science Foundation of Shihezi University (RCZK201953) supported the research. The funders had no role in the study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Disclosure
The authors declare no conflicts of interest in this work.
References
1. Parisi R, Symmons DP, Griffiths CE, Ashcroft DM. Global epidemiology of psoriasis: a systematic review of incidence and prevalence. J Invest Dermatol. 2013;133(2):377–385. doi:10.1038/jid.2012.339
2. Grän F, Kerstan A, Serfling E, Goebeler M, Muhammad K. Current developments in the immunology of psoriasis. Yale J Biol Med. 2020;93(1):97–110.
3. Perera GK, Meglio PD, Nestle FO. Psoriasis. Annu Rev Pathol. 2012.
4. Ammirati AL. Chronic kidney disease. Rev Assoc Med Bras. 2020;66(Suppl 1):s03–s9. doi:10.1590/1806-9282.66.s1.3
5. Verhelst D. Characteristics and epidemiology of chronic kidney disease. Soins. 2018;63(826):14–16. doi:10.1016/j.soin.2018.04.004
6. Ungprasert P, Raksasuk S. Psoriasis and risk of incident chronic kidney disease and end-stage renal disease: a systematic review and meta-analysis. Int Urol Nephrol. 2018;50(7):1277–1283. doi:10.1007/s11255-018-1868-z
7. Kaftan O, Kaftan B, Toppare MF, Ekşioğlu M. Renal involvement in psoriasis. Dermatology. 1996;192(2):189–190. doi:10.1159/000246358
8. Zi J, Deng D. Pathogenesis and manifestations of psoriasis complicated with kidney damage. China J Lep Skin Dis. 2022;38(9):659–663.
9. Ren F, Zhang M, Zhang C, Sang H. Psoriasis-like inflammation induced renal dysfunction through the TLR/NF- κ B signal pathway. BioMed Res Int. 2020;2020(3–4):1–11. doi:10.1155/2020/3535264
10. Wencke W, Fátima S-C, Mercedes R. GOplot: an R package for visually combining expression data with functional analysis. Bioinformatics. 2015;31(17):2912–2914. doi:10.1093/bioinformatics/btv300
11. Ideker T. Cytoscape 2.8: new features for data integration and network visualization. Bioinformatics. 2011;27(3):431–432. doi:10.1093/bioinformatics/btq675
12. Chen B, Khodadoust MS, Liu CL, Newman AM, Alizadeh AA. Profiling tumor infiltrating immune cells with CIBERSORT. Methods Mol Biol. 2018;1711:243–259.
13. Subramanian A, Tamayo P, Mootha VK, et al. Gene set enrichment analysis: a knowledge-based approach for interpreting genome-wide expression profiles. Proc Natl Acad Sci U S A. 2005;102(43):15545–15550. doi:10.1073/pnas.0506580102
14. Gribov A, Sill M, Lück S, et al. SEURAT: visual analytics for the integrated analysis of microarray data. BMC Med Genomics. 2010;3. doi:10.1186/1755-8794-3-21
15. Becht E, Mcinnes L, Healy J, et al. Dimensionality reduction for visualizing single-cell data using UMAP. Nat Biotechnol. 2019;37(1):38–44. doi:10.1038/nbt.4314
16. Aran D, Looney AP, Liu L, et al. Reference-based analysis of lung single-cell sequencing reveals a transitional profibrotic macrophage. Nat Immunol. 2019;20(2):163–172. doi:10.1038/s41590-018-0276-y
17. Jie TU, Zhiqiang YIN. Update of complications of psoriasis. China J Lep Skin Dis. 2019;35(2):120–123.
18. Zeng J, Luo S, Huang Y, Lu Q. Critical role of environmental factors in the pathogenesis of psoriasis. J Dermatol. 2017;44(8):863–872. doi:10.1111/1346-8138.13806
19. Xiao X, Yao T, Du S, Zhang J, Cao Y. Age differences in the pulmonary and vascular pathophysiologic processes after long-term real-time exposure to particulate matter in rats. Chemosphere. 2020;261:127710. doi:10.1016/j.chemosphere.2020.127710
20. Zeng Y, Li M, Zou T, Chen X, Xu H. The impact of particulate matter (PM2.5) on human retinal development in hESC-derived retinal organoids. Front Cell Dev Biol. 2021;9. doi:10.3389/fcell.2021.607341
21. Neophytou AM, Hart JE, Cavallari JM, et al. Traffic-related exposures and biomarkers of systemic inflammation, endothelial activation and oxidative stress: a panel study in the US trucking industry. Environ Health. 1966;12.
22. Chuang KJ, Pan CH, Su CL, et al. Urinary neutrophil gelatinase-associated lipocalin is associated with heavy metal exposure in welding workers. Sci Rep. 2015;5:18048. doi:10.1038/srep18048
23. Kim YJ. Exposure to welding fumes is associated with acute systemic inflammatory responses. Occup Environ Med. 2005;62(3):157–163. doi:10.1136/oem.2004.014795
24. Tan C, Wang Y, Lin M, et al. Long-term high air pollution exposure induced metabolic adaptations in traffic policemen. Environ Toxicol Pharmacol. 2018;58:156–162. doi:10.1016/j.etap.2018.01.002
25. Contassot E, French LE, Beer H-D. The inflammasomes in autoinflammatory diseases with skin involvement. J Invest Dermatol. 2014.
26. Pasparakis M, Haase I, Nestle FO, et al. Mechanisms regulating skin immunity and inflammation. Nat Rev Immunol. 2014;14:289–301. doi:10.1038/nri3646
27. Carlström M, Ekman AK, Petersson S, Söderkvist P, Enerbäck C. Genetic support for the role of the NLRP3 inflammasome in psoriasis susceptibility. Exp Dermatol. 2012;21(12):926–931. doi:10.1111/exd.12049
28. Raposo RA, Gupta R, Abdel‐Mohsen M. Antiviral gene expression in psoriasis. J Eur Acad Dermatol Venereol. 2015;29(10):1951–1957. doi:10.1111/jdv.13091
29. Fäh J, Pavlovic J, Burg G. Expression of MxA protein in inflammatory dermatoses. J Histochem Cytochem. 1995;43:47–52. doi:10.1177/43.1.7822763
30. Melero JL, Andrades S, Arola L, Romeu A. Deciphering psoriasis. A bioinformatic approach. J Dermatol Sci. 2017;89(2):120–126. doi:10.1016/j.jdermsci.2017.11.010
31. van der Fits L, van der Wel LI, Laman JD, et al. Psoriatic lesional skin exhibits an aberrant expression pattern of interferon regulatory factor-2 (IRF-2). J Pathol. 2003;199:107–114. doi:10.1002/path.1263
32. Xia Y, Yang Q, Wu SY, Wu Z, Li Q, Du J. Interferon lambda modulates proinflammatory cytokines production in PBMCs from patients with chronic kidney disease. Hum Immunol. 2023;84(9):464–470. doi:10.1016/j.humimm.2023.06.001
33. Gao M, Lin Y, Liu X, et al. ISG20 promotes local tumor immunity and contributes to poor survival in human glioma. OncoImmunology. 2018;8:1–10.
34. Sun YC, Qiu ZZ, Wen FL, Yin JQ, Zhou H. Revealing potential diagnostic gene biomarkers associated with immune infiltration in patients with renal fibrosis based on machine learning analysis. J Immunol Res. 2022;2022:3027200. doi:10.1155/2022/3027200
35. Jia M, Li L, Chen R, et al. Targeting RNA oxidation by ISG20-mediated degradation is a potential therapeutic strategy for acute kidney injury. Mol Ther. 2023;31(10):3034–3051. doi:10.1016/j.ymthe.2023.07.008
36. Merleev A, Ji-Xu A, Toussi A, et al. Proprotein convertase subtilisin/kexin type 9 is a psoriasis-susceptibility locus that is negatively related to IL36G. JCI Insight. 2022;7(16). doi:10.1172/jci.insight.141193
37. Zhang YJ, Sun YZ, Gao XH, Qi RQ. Integrated bioinformatic analysis of differentially expressed genes and signaling pathways in plaque psoriasis. Mol Med Rep. 2019;20(1):225–235. doi:10.3892/mmr.2019.10241
38. Vabret N, Blander JM. Sensing microbial RNA in the cytosol. Front Immunol. 2013;4:468. doi:10.3389/fimmu.2013.00468
39. Reikine S, Nguyen JB, Modis YE. Pattern recognition and signaling mechanisms of RIG-I and MDA5. Front Immunol. 2014;5. doi:10.3389/fimmu.2014.00342
40. Prens EP, Kant M, Van DG, et al. IFN-alpha enhances poly-IC responses in human keratinocytes by inducing expression of cytosolic innate RNA receptors: relevance for psoriasis. J Invest Dermatol. 2008;128(4):932–938. doi:10.1038/sj.jid.5701087
41. He T, Xiong J, Huang Y, et al. Klotho restrain RIG-1/NF-κB signaling activation and monocyte inflammatory factor release under uremic condition. Life Sci. 2019;231:116570. doi:10.1016/j.lfs.2019.116570
42. Nakashima S. Protein kinase C alpha (PKC alpha): regulation and biological function. J Biochem. 2002;132(5):669–675. doi:10.1093/oxfordjournals.jbchem.a003272
43. Diaz-Meco MT, Moscat J. The atypical PKCs in inflammation: NF-κB and beyond. Immunol Rev. 2012;246(1):154–167. doi:10.1111/j.1600-065X.2012.01093.x
44. Adhikary G, Chew YC, Reece EA, Eckert RL. PKC-δ and-η, MEKK-1, MEK-6, MEK-3, and p38-δ are essential mediators of the response of normal human epidermal keratinocytes to differentiating agents. J Invest Dermatol. 2010;130(8):2017–2030. doi:10.1038/jid.2010.108
45. Ruiz S, Pergola PE, Zager RA, Vaziri ND. Targeting the transcription factor Nrf2 to ameliorate oxidative stress and inflammation in chronic kidney disease. Kidney Int. 2013;83(6):1029–1041. doi:10.1038/ki.2012.439
46. Steinberg SF. Structural basis of protein kinase C isoform function. Physiol Rev. 2008;88(4):1341–1378. doi:10.1152/physrev.00034.2007
47. Ovigne J-M, Baker BS, Brown DW, et al. Epidermal CD8+ T cells in chronic plaque psoriasis are Tc1 cells producing heterogeneous levels of interferon-gamma. Exp Dermatol. 2001;10(3):168–174. doi:10.1034/j.1600-0625.2001.010003168.x
48. Di Meglio P, Duarte JOH. CD8 T cells and IFN-γ emerge as critical players for psoriasis in a novel model of mouse psoriasiform skin inflammation. J Invest Dermatol. 2013;133(4):871–874. doi:10.1038/jid.2012.426
49. Ellis CN, Fradin MS, Messana JM, Brown MD, Parish TG. Cyclosporine for plaque-type psoriasis. Results of a multidose, double-blind trial. New Eng J Med. 1991;324(5):277–284. doi:10.1056/NEJM199101313240501
50. Kaszubowska L, Foerster J, Kwiatkowski P, Schetz D. NKT-like cells reveal higher than T lymphocytes expression of cellular protective proteins HSP70 and SOD2 and comparably increased expression of SIRT1 in the oldest seniors. Folia Histochem Cytobiol. 2019;56(4):231–240. doi:10.5603/FHC.a2018.0025
51. Atanasiu V, Manolescu B, Stoian I. Hepcidin the link between inflammation and anemia in chronic renal failure. Rom J Intern Med. 2006;44(1):25–33.
52. Krause A, Neitz S, Mgert H-J, et al. LEAP-1, a novel highly disulfide-bonded human peptide, exhibits antimicrobial activity. FEBS Lett. 2000;480(2–3):147–150. doi:10.1016/S0014-5793(00)01920-7
53. Xiong J, Qiao Y, Yu Z, et al. T-lymphocyte subsets alteration, infection and renal outcome in advanced chronic kidney disease. Front Med. 2021;8:742419. doi:10.3389/fmed.2021.742419
54. Shanahan CM. Mechanisms of vascular calcification in CKD-evidence for premature ageing? Nat Rev Nephrol. 2013;9(11):661–670. doi:10.1038/nrneph.2013.176
55. Sarakpi T, Mesic A, Speer T. Leukocyte-endothelial interaction in CKD. Clin Kidney J. 2023;16(11):1845–1860. doi:10.1093/ckj/sfad135
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms.php
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms.php
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Identification of an Immune-Related Gene Diagnostic Model and Potential Drugs in Sepsis Using Bioinformatics and Pharmacogenomics Approaches
Chen P, Chen J, Ye J, Yang L
Infection and Drug Resistance 2023, 16:5665-5680
Published Date: 28 August 2023
A Case Report of Adalimumab Successfully Treated a Severe Plaque Psoriasis Patient with Psoriatic End-Stage Renal Disease
Xu J, Chen A, Peng Y, Wen Z, Huang K, Wang P
Clinical, Cosmetic and Investigational Dermatology 2023, 16:2841-2846
Published Date: 12 October 2023
Identification of Immune-Related Genes as Biomarkers for Uremia

Lyu D, He G, Zhou K, Xu J, Zeng H, Li T, Tang N
International Journal of General Medicine 2023, 16:5633-5649
Published Date: 29 November 2023
Based on Weighted Gene Co-Expression Network Analysis Reveals the Hub Immune Infiltration-Related Genes Associated with Ulcerative Colitis
Tan Z, Liu C, He P, Wu Y, Li J, Zhang J, Dong W
Journal of Inflammation Research 2024, 17:357-370
Published Date: 17 January 2024