")
Back to Journals » Journal of Pain Research » Volume 18
Acupotomy Ameliorates KOA Related Chondrocyte Premature Senescence Through YAP/FOXD1 Pathway
Authors Ma Y , Hu T , Liu N , Guo C, Xing L , Ma W, Cui Y, Chen X
Received 17 September 2024
Accepted for publication 19 March 2025
Published 12 April 2025 Volume 2025:18 Pages 2011—2023
DOI https://doi.org/10.2147/JPR.S475829
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Wendy Imlach
Yunxuan Ma,1,* Tingyao Hu,1,* Naigang Liu,2 Changqing Guo,1 Longfei Xing,3 Weiwei Ma,4 Yongqi Cui,1 Xilin Chen5
1School of Acupuncture-Moxibustion and Tuina, Beijing University of Chinese Medicine, Beijing, People’s Republic of China; 2Department of Acupuncture-moxibustion, China-Japan Friendship Hospital, Beijing, People’s Republic of China; 3Department of Traditional Chinese Medicine, Peking University People’s Hospital, Beijing, People’s Republic of China; 4Department of Traditional Chinese Medicine, Wuzhong People ‘s Hospital, Ningxia, People’s Republic of China; 5Department of Acupuncture and Rehabilitation, The Fifth College of Clinical Medicine, Guangzhou University of Traditional Chinese Medicine, Guangzhou, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Naigang Liu, Department of Acupuncture, China-Japan Friendship Hospital, Beijing, 100010, People’s Republic of China, Tel +86-13488782471, Fax +86-1084205333, Email [email protected]
Purpose: Premature senescence of chondrocytes is a typical lesion of knee osteoarthritis (KOA). Abnormal cartilage stress can inhibit the mechanosensitive Yes-associated protein (YAP) / transcription factor forkhead box D1 (FOXD1) pathway, which is related to premature senescence of chondrocytes, thereby accelerating the progression of the lesion. This study aims to investigate whether acupotomy intervention could inhibit the premature senescence of chondrocytes and protect the cartilage of KOA rabbits.
Methods: 18 male New Zealand rabbits were randomly divided into 3 groups (n = 6 each): control, KOA, and KOA + acupotomy (KOA+Apo). KOA, KOA+Apo rabbits were modeled by modified Videman’s method for 6 weeks. After modeling, the KOA+Apo groups were subjected to acupotomy once a week for 3 weeks on the muscles around the left hind knee. The modified Lequesne MG score and passive range of motion (PROM) were used to evaluate the general condition and exercise ability of rabbits. Cartilage degeneration was detected by safranin O-fast green staining and transmission electron microscope(TEM). Type II collagen (Col-II) and aggrecan by immunohistochemistry (IHC), IL-7 and MMP-13 by Enzyme-Linked Immunosorbent Assay (ELISA), and p53, Rb1, p - YAP, YAP, FOXD1 by IHC, Western blot, or RT - PCR.
Results: Acupotomy effectively curbed cartilage degeneration and chondrocyte premature senescence in KOA rabbits. Mechanistically, it cut IL - 7 and MMP-13 levels, easing the inflammatory milieu and extracellular matrix degradation. It also regulated p53 and Rb1, controlling cell - cycle progression. Crucially, acupotomy upregulated the YAP/FOXD1 pathway, which, by affecting downstream genes, modulated IL - 7, MMP-13, p53, and Rb1 levels, acting as a pivotal molecular link in its regulatory effects.
Conclusion: Acupotomy may protect KOA rabbits’ cartilage by inhibiting chondrocytes premature senescence via the YAP/FOXD1 pathway, offering a new theoretical basis for treating mechanically - induced KOA.
Keywords: acupotomy, knee osteoarthritis, chondrocytes, premature senescence, YAP/FOXD1 pathway
Introduction
KOA is characterized by progressive cartilage degeneration. Joint stiffness, pain, and pathological changes in the knee’s histology, including cartilage inflammation, joint tissue remodeling, osteophyte formation, subchondral bone changes, and joint space narrowing occurs in KOA as well.1,2 Biochemical factors, mechanical stress, and age are the main factors that cause the abnormality of the synthesis and catabolism of the cartilage extracellular matrix (ECM), which mainly includes type II collagen (Col-II) and aggrecan. Mechanical imbalance of the knee joint leads to abnormal cartilage stress, inducing KOA.3 Research indicates that the overloaded mechanical stress on the knee accelerates the initiation of articular cartilage damages and progression of cartilage degeneration, which causes the disturbed stress distribution in cartilage, and leads to mechanical stress concentration. When a vicious circle forms between mechanical loadings and cartilage structure, it promotes the procession of the KOA disease course.4
During the progression of KOA, the abnormal mechanical load on the body exceeds its repair capacity, destroys chondrocyte, downregulate CDK 2 and CDK 4/6 expression through the tumor suppressor p53-p21-Rb pathway and the p16INK4a-Rb pathway, causing stress-induced premature senescence (SIPS), which refers to the cell cycle entering an irreversible state of growth arrest.5,6 Cell cycle arrest is a cellular self-protective strategy in response to the external injury, which inhibits cell proliferation.7 However, instead of protecting cartilage from damage, premature chondrocytes accelerate KOA progression. KOA chondrocytes exhibit changes such as hypertrophy and premature senescence, resulting in articular cartilage loss and degradation.8 Studies have shown that transplantation of premature senescent chondrocytes into the knee joint cavity of wild-type mice leads to KOA-like changes.9,10
In normal situations, chondrocyte remains in a quiescent state, and physiological cyclical loading promotes cartilage homeostasis.11,12 p16 and p21 are known as key senescence biomarkers, their upregulation inhibits cell proliferation and hinders cartilage regeneration in the process of KOA.13 Premature senescence chondrocytes promote cartilage degeneration and KOA aggravation by secreting senescence-associated secretory phenotype (SASP).14 SASP mainly includes proinflammatory factors (e.g., IL-7), reactive oxygen species and ECM metalloproteinases (MMPs, e.g., MMP-13), which aggravates joint inflammation, degradates Col-II and aggrecan in the cartilage matrix,15 induces secondary premature senescence in adjacent normal chondrocytes, and results in a vicious cycle that accelerates cartilage degeneration and KOA progression.16
YAP is a downstream protein and effector of the Hippo-YAP signaling pathway, overexpression of YAP protects articular cartilage from degradation during KOA pathogenesis17 It also has been shown to be a mechanosensitive transcription factor, which integrates the cartilage extracellular environment in response to stress, and activates with increased mechanical stress, regulate cell proliferation and premature senescence.18 As a therapeutic target, it promotes cartilage repair and prevents secondary osteoarthritis.19 YAP transcriptionally activates FOXD1 expression whereas YAP deficiency results in down-regulation of FOXD1, which contributes to the early-onset of cellular aging, and FOXD1 blocks cellular senescence and apoptosis through the p21/CDK2/Rb signaling pathway.20 Within the range of repair, joint damage can deactivate the Hippo pathway and activate YAP, allowing it to enter the nucleus and form a complex with TEA domain transcription factor (TEAD), which can jointly activate the transcription of its downstream factor, FOXD1, and YAP attenuates cartilage degradation during KOA progression and preserves cartilage integrity.21 However, under the pathological conditions of KOA, OA - associated cellular environmental alterations activate the Hippo pathway, leading to YAP phosphorylation. Phospho-YAP (p-YAP) is retained in the cytoplasm and degraded by ubiquitination.22 Consequently, YAP and FOXD1 regulation in the nucleus prompts the activation of the p53-p21-Rb and p16INK4a-Rb axis, causing premature senescence of chondrocytes.23 The YAP/FOXD1 pathway, which is well - recognized as a regulator of the cellular stress response, is biologically relevant to KOA caused by trauma or mechanical imbalance. Therefore, the YAP/FOXD1 pathway is crucial for chondrocytes and is a significant target for KOA treatment.24
Currently, there is no single treatment that could completely cure KOA. Non-steroidal anti-inflammatory drugs (NSAIDs) are commonly used, which bring various side effects, including gastrointestinal, cardiovascular, renal, and liver diseases.25,26 Guidelines and consensus recommended acupuncture (including manual acupuncture and acupotomy) as a non-pharmacological treatment for KOA, believing it effectively reduces pain.27,28 TCM diagnosis and treatment of knee osteoarthritis also suggest acupuncture for all stages of KOA.29 The onset of KOA is related to biomechanical structure change, where long-term injury-repair cycle can lead to a decrease of the muscle-tendon function.30 Evidence suggest that biomechanical therapies can promote the mechanical balance of the knee joint, achieve more sustained effects than anti-inflammatory therapies.31 Previous studies have shown that acupotomy, a medical device combining the characteristics of acupuncture needles and surgical knives, plays a significant role in treating KOA caused by abnormal mechanical forces. These abnormal forces, stemming from factors like trauma, poor postures, exercise habits, and aging, can disrupt articular cartilage metabolism, cause synovitis, and lead to KOA. Acupotomy intervenes by improving soft tissue mechanical properties, releasing the tense, contracted, and adhered soft tissues around the knee joint. This helps adjust the mechanical balance of the knee joint, reducing abnormal intra - articular pressure and improving the uneven force distribution and limited joint mobility.32,33 Moreover, it stimulates local blood vessels and nerves, promoting blood circulation, which provides more nutrients to tissues, removes metabolic waste and inflammatory mediators, reduces the inflammatory response and alleviates pain. Additionally, it also helps improve joint function. Previous research by our team has found that acupotomy can reduce the positive expression of HP1 in the chondrocytes of the affected knee joint in KOA model rabbits, and downregulate the protein expression of p16Ink4a and p21Waf1/Cip1.10 This suggests that acupotomy can delay the progression of KOA by inhibiting premature senescence of chondrocytes. How the mechanical effects of acupotomy protect cartilage by alleviating abnormal cartilage stress, and through which mechanically sensitive pathway it inhibits premature senescence of chondrocytes in KOA remain to be further investigated.
Materials and Methods
Animals and Study Design
Eighteen clean grade New Zealand rabbits, weighing 2.1–2.6 kg, 6 months old, were purchased from Fulong Tengfei Experimental Animal Research Institute Ltd., Beijing, China, (Certificate No: SYXK2018–0041). Animals were provided food and water ad libitum and maintained on a 12/12 h light/dark cycle at 20–25°C with 50–60% humidity. Study protocol was approved and reviewed by the Animal Ethics Committee at Beijing University of Chinese Medicine (Beijing, China, Certificate No: BUCM-4-2022010101-1097). Rabbits were randomly divided into 3 groups (n = 6): Control group (Control), KOA group (KOA), KOA + acupotomy group (KOA+Apo). General condition and exercise capacity were evaluated using the modified Lequesne MG score and PROM. Cartilage degeneration was assessed by safranin O-fast green staining and TEM, the expression of cartilage Col-II and aggrecan by IHC, IL-7 and MMP-13 level in synovial fluid by ELISA, and the expression of senescence-related factors p53 and Rb1 in chondrocytes by Western blot, to evaluate the intervention effect of acupotomy on premature senescence of chondrocytes. The protein and gene expression levels of p-YAP, YAP and FOXD1 were detected by Western blot and RT-PCR.
KOA Modeling
Rabbits were anesthetized with 3% pentobarbital sodium (30 mg/kg), placed in a supine position on the operating table, with their left hind legs fully extended. The area from groin to the toe was fixed with a resin bandage. Wound polymer plasters were applied to strengthen the fixation and covered with an anti-bite bandage. During immobilization, the blood supply of the left hind limb was monitored by the swelling and color of the toe, and adjustments to the immobilization mold were made as needed. The control group underwent regular handling without KOA modeling or any intervention. The mold was removed after 6 weeks of fixation.
Acupotomy Interventions
After modeling, rabbits in the KOA+Apo group received acupotomy interventions. After routine skin preparation and disinfection, points on the tendons of the vastus medialis, vastus lateralis, cartilage, biceps femoris, and anserine bursa around the knee were selected. Disposable acupotomy needles (ɸ 0.4×40 mm, Hanzhang acupotome, Beijing Excellence Huayou Medical Instruments Co., Ltd., Beijing, China) were used. The blade tip of the acupotomy needle was kept parallel to the leg and muscle fiber. The needle was inserted perpendicularly to a depth of 4−5 mm and released the tissue 1–2 times, the needle was then taken out, followed by pressure applied for a moment to prevent bleeding. The intervention was administered once a week for 3 weeks.
Behavioral Test
After modeling and after acupotomy intervention, respectively, the modified Lequesne MG grade (knee pain (0–3 points), gait (0–3 points), joint activity (0–3 points), joint swelling (0–2 points)) was used to evaluate the KOA model. In addition, the PROM of the left hind limb of the rabbits was measured with a manual goniometer (SL1610A0051, Sanliang Instrument Co., Ltd., Shanghai, China) by two independent researchers (Longfei Xing and Weiwei Ma), they were unaware of the grouping. PROM = Maximum extension angle - maximum flexion angle. The average score was used for analysis.34
Safranin O-Fast Green Staining
The cartilage-subchondral bone complex was fixed with 4% paraformaldehyde (Servicebio, Wuhan, China) for 72 hours, then decalcified in EDTA decalcification solution. The solution was changed every 4 days for a total of 8 weeks until the syringe needle could easily penetrate the tissue. Subsequently, the tissue was dehydrated, transparentized, waxed, and embedded in paraffin. Paraffin sections with a thickness of 4 μm were prepared from the sagittal plane using a Leica paraffin microtome (Leica, Wetzlar, Hesse, Germany). The cartilage sections were stained using the modified Safranin O-Fast Green FCF cartilage staining kit (Solarbio, Wuhan, China). Two researchers, blinded to the group assignments, evaluated the stained sections according to the Mankin’s score.
TEM Observation of Rabbits Chondrocyte Ultrastructural Morphology
The TEM fixative (Servicebio, Wuhan, China) was added to the tube and let the precipitation re-suspended in the fixative, and then fixed at 4°C for preservation and transportation. A volume of 1mm × 1mm × 1mm cartilage was removed from the sacrificed rabbit knee with a sharp knife and then fixed at 4°C in TEM fixative for 2–4 hours. After pre-embedding, post-fixation, dehydration, polymerization, ultrathin section and staining, the cartilage was observed under transmission electron microscope (HITACHI, Tokyo, Japan).
IHC Analysis
The cartilage-subchondral bone complex samples were fixed in 4% paraformaldehyde and decalcified in neutral 10% EDTA at room temperature for 4 weeks. After dehydration in ethanol and immersion in paraffin, the samples were sectioned at a thickness of 5 μm from the sagittal plane. To observe the expression of Col-II and aggrecan in cartilage tissue, IHC staining was performed using an IHC kit (ZSGB-BIO Co., Ltd., Beijing, China) according to the instructions. The sections were deparaffinized, rehydrated, and subjected to antigen retrieval. Subsequently, they were treated with 3% H2O2 for 25 min and blocked with 10% normal goat serum (Servicebio, Wuhan, China) for 10 min. The sections were then incubated overnight at 4°C with anti-Col-II (1:200, Proteintech, Chicago, IL, USA) and anti-aggrecan (1:200, Novus, CO, USA). After washing with PBS, the sections were incubated with HRP-conjugated secondary antibody (HRP-conjugated goat anti-mouse IgG, 1:200, Servicebio, Wuhan, China) for 50 min. The DAB IHC staining kit was applied as chromogen, followed by counterstaining with hematoxylin, alcohol gradient dehydration, and mounting with neutral resin. High-quality images were captured in six regions of each sample under microscope (Olympus, Tokyo, Japan), and analyzed with Image-Pro Plus 6.0. Statistical analysis for Col-II and aggrecan was conducted on the ratio of integral optical density to area (IOD/area).
ELISA Analysis
The inflammation cytokine interleukin (IL-7) and matrix metalloproteinase (MMP-13) in the synovial fluid were measured by ELISA assay kits (Nanjing Jiancheng Bioengineering Institute, Nanjing, China). At the time of sacrifice, the synovial fluid was collected and stored at −80°C until assay. The frozen synovial fluid was thawed and centrifuged to obtain supernatant. All analyses were performed according to the manufacturer’s instructions.
Western Blot Analysis
Fresh cartilage from the left knee of rabbits was dissected with bone cutters, ground in a mortar with liquid nitrogen, and collected into centrifuge tubes. RIPA lysis buffer (Servicebio, Wuhan, China) and PMSF (Servicebio, Wuhan, China) were added to the tubes in a 10:1 volume ratio to extract total proteins. Total protein was quantified using the BCA assay kit (Solarbio, Wuhan, China). A 12% SDS-PAGE (Nanjing Aoqing Bioengineering Institute, Nanjing, China) was used to load 3 μL of marker and 10 μL of protein samples at 160 V for 30 min. Proteins were then transferred to a 0.22 μm polyvinylidene difluoride (PVDF) membrane (Millipore, Boston, Mass, USA) at 400 mA for 25 minutes. The PVDF membrane was placed in a rapid blocking solution (Solarbio, Wuhan, China) at room temperature for 20 minutes, followed by overnight incubation at 4°C with primary antibodies against anti-P-YAP (1:1000, Ser127, Affbiotech, Cincinnati, OH, USA), YAP (1:1000, Bioss, Beijing, China), FOXD1 (1:2000, Bioss, Beijing, China), Rb1 (1:2000, Bioss, Beijing, China), p53 (1:1000, Proteintech, Chicago, United States) and GAPDH (1:10,000, Abcam, Cambridge, UK). The membranes were incubated with horseradish peroxidase (HRP)-conjugated secondary antibodies at room temperature for 1 hour, and finally, the ECL chemiluminescence detection system (Servicebio, Wuhan, China) was scanned and analyzed via an image analysis system (Image Lab (TM) Touch Software, version 2.4.0.03, Bio-Rad, CA, US). The blots were quantified by densitometry using the Image J software (Rockville, MD, USA). The detailed experimental procedures are described in Appendix A. Instructions S6.
Real-Time Quantitative Polymerase Chain Reaction (RT-PCR) Analysis
The cartilage of the tibia and femur was quickly collected, frozen in liquid nitrogen, and stored in a −80°C refrigerator. The total RNA of the cartilage of each rabbit was isolated with TRIZOL (Servicebio, Wuhan, China), and reverse-transcribed into cDNA using a RT First Strand cDNA Synthesis Kit. YAP and FOXD1 gene expression levels in cartilage were further quantified by SYBR Green RT-PCR Master Mix (2 ×) (Servicebio, Wuhan, China). The relative expression of the target gene was normalized into the expression of GAPDH using the 2-ΔΔCt method. The primer sequences are presented in Table 1.
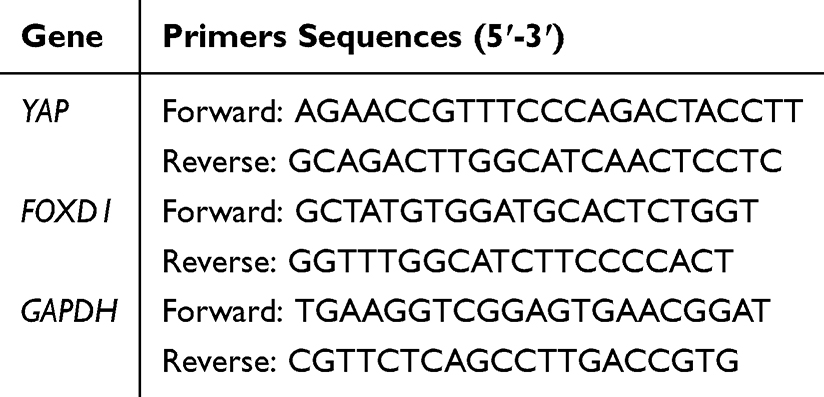 |
Table 1 Primers Used in This Study |
Statistical Analysis
All data were statistically analyzed using SPSS software (version 20.0, SPSS, Inc., Chicago, USA). When conform to a normal distribution, the data were presented as mean ± SD, and variance analyses of the main effect and interaction effect were implemented using a general linear model. Multiple comparisons of differences between groups were achieved using the least significant difference (LSD) test, and among multiple groups by performing one-way analysis of variance (ANOVA). When data do not conform to a normal distribution and variance is not homogeneous, a nonparametric test (Mann–Whitney) was used, and data were expressed as median (interquartile range)/M (QR). A value of P < 0.05 was considered statistically significant, and higher significance levels were established at P < 0.01.
Results
General Condition and Behavior
Before modeling, there were no significant differences observed among the rabbits in terms of general behavior (eye and body movements), luster, feces, and feeding. Following the induction of KOA, the KOA group exhibited varying degrees of lethargy and reduced activity, which intensified over time. In contrast, no significant changes in the general condition of rabbits in the control group were observed throughout the study. After acupotomy intervention, rabbits in the KOA+Apo group showed varying degrees of improvement in their general condition. These findings suggest that acupotomy intervention can effectively ameliorate systemic and behavioral symptoms in the KOA model rabbits.
Acupotomy Improved Lequesne MG and PROM Scores
Stiffness in the knee joint is a characteristic clinical manifestations of KOA. Studies have suggested that acupotomy has the potential to improve joint mobility, gait changes, and pain. This study evaluated the modified Lequesne MG and PROM scores of KOA rabbits to assess the severity of KOA and the mobility of the knee joint. Lower Lequesne MG score and greater PROM indicated better behavior and joint motion. After 6 weeks’ immobilizing the left hindlimb with modified Videman’s method, the KOA rabbits displayed diminished activity, gait changes, muscle atrophy, and palpable fibrous bands in the affected limb. After acupotomy intervention, compared with the KOA group, the KOA+Apo group demonstrated a significant decrease in the Lequesne MG score (P < 0.01; Figure 1A) and a substantial elevation in the PROM score (P < 0.01; Figure 1B). This indicated that by releasing constrained substances within the soft tissues surrounding the knee, acupotomy intervention may alleviate joint stiffness and pain, and improve functional impairment in the KOA rabbit model. The detailed experimental data are available in Supplementary Appendix A. Figure S1A and S1B.
 |
Figure 1 Acupotomy improved Lequesne MG and PROM scores. (A) Lequesne MG scores before and after rabbit knee intervention in each group. (B) PROM scores before and after intervention in rabbit knees.**P < 0.01 vs the Control group. ##P < 0.01 vs the KOA group. |
Acupotomy Alleviates Cartilage Degeneration and Mitigates the Degradation of ECM (Col-II, Aggrecan) in KOA Rabbits
To observe the effect of acupotomy on cartilage degeneration, this study stained the cartilage with Safranin O-Fast Green staining (Figure 2A), and scored the degree with Mankin’s score (Figure 2B). In comparison to the control group, the KOA group displayed a decrease in cartilage thickness, surface defects, reduced chondrocyte necrosis, and a deficiency in Safranin O-Fast Green staining on the cartilage surface. When compared to the KOA group, the KOA+Apo group exhibited less prominent cartilage defects and reduced loss of Safranin O-Fast Green staining. Compared to the control group, the score of KOA group increased significantly (P < 0.01), while KOA+Apo group significantly decreased compared to the KOA group (P < 0.01).These results indicated that acupotomy reduced the cartilage degeneration in KOA rabbits. The detailed experimental data are available in Supplementary Appendix A. Figure S2B.
 |
Figure 2 Acupotomy alleviates cartilage degeneration in KOA rabbits. (A) Representative cartilage tissue from each group with Safranin O-Fast Green staining, Magnification:100 ×, 200 ×. (B) Mankin’s score in each group. **P < 0.01 vs the Control group. ##P < 0.01 vs the KOA group. |
In this experiment, the proteins of Col-II and aggrecan in cartilage ECM were stained brown (Figure 3A). Using Image Pro Plus 6.0 for quantification, the target area was circled, extending from the superficial surface of cartilage to part of the radiate layer, and the IOD value was measured. AOD (IOD/Area) was compared between groups, indicating the relative expression levels of Col-II and aggrecan in each group (Figure 3B). The results showed that the relative expression of Col-II and aggrecan in the KOA group was significantly reduced (P < 0.01), consistent with previous study, and further confirmed that changes in the morphology and microstructure of subchondral bone can seriously affect the homeostasis of cartilage and accelerate the degradation of ECM. After treatment, the relative expression of Col-II and aggrecan was significantly increased in the KOA+Apo group compared with the KOA group (P < 0.01). These results showed that the acupotomy significantly increased the relative expression of Col-II and aggrecan in KOA rabbit cartilage, promoted the repair of Col-II and aggrecan in the cartilage ECM. The detailed experimental data are available in Supplementary Appendix A. Figure S3B.
 |
Figure 3 Acupotomy mitigates the degradation of extracellular matrix in cartilage (Col-II, aggrecan). (A) IHC staining of Col-II and aggrecan, Bars: 50 μm, magnification: 400 ×. (B) The mean optical density values of Col-II and aggrecan. **P < 0.01 vs the control group. ##P < 0.01 vs the KOA group. |
Acupotomy Inhibits Premature Senescence of Chondrocytes
TEM observation have shown that senescent cells tend to be enlarged and flattened, with irregular shapes, and nucleoli may become more prominent.35 Cytoskeletal reorganization may also lead to distinct changes in cell morphology. In this experiment, the KOA group had the most serious cell damage, with suspected senescent cells, fewer processes around cells, reduced cytoplasmic density, organelle swelling and cavitation, and visible lysosomes. The damage in the KOA+Apo group was mild, with partial protrusion around the cell, uniform cytoplasm, and slightly swollen mitochondria. The control group had the least injury, with protrusion beside the cell membrane, evenly distributed cytoplasm, and acceptable mitochondria and endoplasmic reticulum structures (Figure 4A).
 |
Figure 4 Acupotomy inhibits premature senescence of chondrocytes. (A) TEM observation of rabbits chondrocyte ultrastructural morphology, nucleus (N), nucleoli (Nu), crude surface endoplasmic reticulum (RER), mitochondria (The green arrow), lysosome (The red arrow), autolysosome (The blue arrow). Bars: 5 µm in the first row; 1 µm in the second row, magnification: 2500 ×, 8000 ×. (B) IL-7 and MMP-13 content in the synovial fluid of the rabbit joint cavity in each group. (C) Western blots for each group. (D) Protein level comparison of p53 and Rb1 between groups. **P < 0.01 vs the Control group. ##P < 0.01 vs the KOA group. |
In this study, the content of IL-7 and MMP-13 in synovial fluid was detected by ELISA, which showed that the expression of IL-7 and MMP-13 was significantly increased in the KOA group (P < 0.01) compared with the control group, and it was significantly decreased (P < 0.01) in the KOA+Apo group compared with KOA group (Figure 4B). These results suggested that acupotomy inhibited the increased levels of IL-7 and MMP-13 in synovial fluid in KOA rabbits, improving the environment of chondrocytes in KOA rabbits by reducing the expression of IL-7 and MMP-13, the premature senescence related factors. Western blot was used to detect the protein expression of p53 and Rb1 in the cartilage tissue of rabbit knee joints (Figure 4C). The analysis of the results showed that compared with the control group, p53 and Rb1 were significantly increased in the KOA group (P < 0.01). Compared with the KOA group, chondrocytes expressing p53 and Rb1 were significantly reduced in the KOA+Apo group (P < 0.01, Figure 4D). These results suggest that acupotomy can release the muscles and tendons around the knee joint of KOA rabbits, improve the mechanical environment of articular cartilage, regulate the normal cycle of cells, and delay the degeneration of cartilage. The detailed experimental data are available in Supplementary Appendix A. Figure S1A and S1B.
Acupotomy Up-Regulated the Expression of YAP-FOXD1 Pathway
The YAP/FOXD1 pathway plays a vital role in premature senescence. This study detected the effect of acupotomy in the process of premature senescence through YAP/FOXD1 pathway and the expression of YAP and FOXD1 (Figure 5). Western blot (Figure 5A) showed that compared with the control group, the expression of YAP and FOXD1 in KOA group was significantly up-regulated (P < 0.01). Compared with KOA group, the expression of YAP and FOXD1 in KOA+Apo group was significantly down-regulated (P < 0.01) (Figure 5B).
 |
Figure 5 Acupotomy up-regulated the expression of YAP-FOXD1 pathway. (A) Western blots for each group. (B) Protein level comparison of p-YAP and FOXD1 between groups. (C) mRNA expression levels comparison of p-YAP and FOXD1 between groups. **P < 0.01 vs the control group. #P < 0.05 vs the KOA group, ##P < 0.01 vs the KOA group. |
Similar results were shown in RT-PCR when detecting the mRNA expression of YAP, FOXD1 in cartilage (Figure 5C). Compared with the control group, the mRNA expression of YAP and FOXD1 in KOA group was up-regulated (P < 0.05, P < 0.01). Compared with KOA group, the mRNA expression of YAP and FOXD1 in KOA+Apo group was significantly down-regulated (P < 0.01). The results suggested that acupotomy may help regulate premature senescence through the YAP-FOXD1 pathway. The detailed experimental data are available in Supplementary Appendix Figure S5A–S5C.
Discussion
According to this study, after 6 weeks of modeling by the modified Videman’s method, factors associated with the YAP/FOXD1 pathway as well as premature in chondrocytes were inhibited, the ECM was damaged, the mitochondria were accumulated, and the distribution of Col-II and aggrecan in cartilage was reduced. Acupotomy improved mitochondrial morphology, and enhanced the distribution of Col-II and aggrecan in cartilage, and these effects were may be mediated through the activation of the YAP/FOXD1 pathway.
Acupotomy is a treatment method combining acupoint-stimulation of TCM acupuncture and soft tissue releasing of scalp, which is effective in restoring the biomechanical characteristics of joints in KOA.35,36 Joint immobilization is one of the commonly used methods for the preparation of KOA experimental models, which can diminished activity of the knee joint and its surrounding muscles, lead to abnormal morphology, muscle function and varying degrees of joint stiffness and swelling, muscle atrophy, adhesion, cartilage degeneration, all of which can result in abnormal mechanical stress in KOA.37 Acupotomy can improve the state of abnormal structure and function of the tendons, the key to cause or aggravate pain and other symptoms. In this study, modeling KOA using a modified Videman’s method was found to be more suitable for exploring the relevant mechanisms of acupotomy treatment for KOA. The modified Lequesne MG score of KOA rabbits decreased, pain relieved, knee dysfunction improved and the range of motion of the rabbit’s knee joint increased, which indicated that by releasing the muscles and tendons around the rabbit knee joint, acupotomy intervention corrected the abnormal mechanical environment of articular cartilage, promoted the repair of the knee joint microcirculation, and reduced cartilage degeneration, thereby restored the biomechanical characteristics of the joint.
Cartilage is mainly composed of chondrocytes and ECM. Chondrocytes are the only cell type, representing about 5% of cartilage volume, while the ECM constitutes about 95%, including Col-II and aggrecan. The biomechanical properties of ECM and its effects on cell homeostasis have been identified as drivers of premature senescence. They provide tensile and compressive strength to cartilage and confer biomechanical properties to articular cartilage.38 It is recognized that the main pathological change in KOA is articular cartilage degeneration, with an imbalance in the production and degradation of ECM being a key factor in the occurrence and development of articular cartilage degeneration.39 The loss of balance between synthesis and degradation of the cartilage matrix shifts to catabolism, promoting pathological tissue remodeling and eventually leading to cartilage degradation.40 Pathological results after model creation showed obvious defects on the cartilage surface, significant loss of Col-II and aggrecan staining, and a significant reduction in Col-II distribution and content, consistent with our previous model creation effects. This suggests that the established model of KOA is similar to clinical practice. Mankin’s score indicated that compare with the KOA group, the surface defects of the cartilage were significantly reduced, and the contents of Col-II and aggrecan were increased in the ECM, indicating that the acupotomy can inhibit the degradation of cartilage ECM, slow down the progression of KOA, and effectively protect the cartilage.
SIPS of chondrocytes is a major factor leading to the imbalance of cartilage homeostasis and functional degradation, and is a critical link in the pathogenesis of KOA.15 Abnormal stress can induce premature decay of chondrocytes and accelerate cartilage degeneration, further accelerating KOA progression.3 When cells enter a premature senescence state their protein expression profile changes significantly, with secreting numerous cytokines and chemokines. They also present senescence-associated secretory phenotype (SASP) which can induce a senescence phenotype in cells around, causing premature senescence of other cellular components and establishing a pro-senescence cycle.41,42 In the pathological development process of KOA, the accumulation of SASP (such as the inflammatory cytokines IL-7) leads to an increase in intracellular MMPs, which damages the ECM and causes tissue senescence, and then causing joint structural damage.43,44 MMP13, a key enzyme in cartilage pathology, is regulated by growth factors, hormones, cytokines and transcription factors. Significantly elevated levels of MMP-13 expression can lead to OA cartilage matrix degradation and disease progression.45,46 In summary, the pathological development process of premature senescence is driven by an inflammatory microenvironment and cellular damage.47 The inflammatory environment induced by SASP factors is associated with cartilage degradation and subchondral bone remodeling, ultimately leading to cartilage loss and KOA progression.48 Inhibiting chondrocyte premature senescence is particularly important to delay KOA progression. Our study chose the senescence-associated secretory phenotypes IL-7 and MMP-13, and the cellular senescence-related factors p53 and Rb1, to suggest premature senescence of chondrocytes. In this study, IHC and Western blot showed that in the KOA group, the secretion of the above premature senescence factors increased, and the expression decreased after acupotomy, suggesting an improvement of the inflammatory microenvironment. The increased expression of Rb1 and p53 after KOA modeling, indicating premature senescence of KOA chondrocytes (Figure 4). Acupotomy intervention can decrease the level of Rb1 and p53 and inhibit premature chondrocytes, which may explain the promotion of Col-II and aggrecan production and metabolism in the ECM. In general, it can inhibit premature senescence of chondrocytes by suppressing the paracrine of SASP.
YAP/FOXD1 pathway is a key to initiating premature senescence of chondrocytes and is an important target for the treatment of KOA. Slowing the premature senescence of local osteoarticular cells mediated by YAP or FOXD1 helps create a pro-chondrogenic environment and alleviate disease symptoms. Studies have shown that intra-articular injection of lentivirus expressing YAP or reduces the number of senescent cells, suppresses joint inflammation and cartilage erosion, and ameliorates pathological symptoms.22 Up-regulating the YAP/FOXD1 pathway can inhibit the premature senescence of chondrocytes and effectively protect cartilage. This study observed the YAP was significantly down-regulated in KOA cartilage, and lower YAP expression indicated more severe articular cartilage degeneration.49 When the mechanical stress on the knee joint is increased, YAP is activated in chondrocytes, which then phosphorylates YAP and activates downstream FOXD1. Therefore, up-regulation of YAP and FOXD1 expression and restoring the normal function of this pathway are key to inhibiting the premature senescence of chondrocytes. Additionally, proper mechanical stress can activate the YAP/FOXD1 pathway.50 In this study, acupotomy may contribute to upregulating repressed YAP and FOXD1 in KOA model rabbit chondrocytes.
Conclusions
In conclusion, this study suggests that acupotomy intervention may inhibit premature senescence of chondrocytes in rabbit KOA models by reducing chondrocyte matrix degradation and paracrine secretion of SASP related to premature senescence. The mechanism might involve inhibiting the YAP/FOXD1 pathway, thus alleviating cartilage degeneration and protecting cartilage. These findings could provide a new theoretical basis for acupotomy’s potential in delaying mechanical KOA progression.
Abbreviations
KOA, knee osteoarthritis; YAP, Yes-associated protein; FOXD1, transcription factor forkhead box D1; PROM, passive range of motion; Col-II, type II collagen; ELISA, Enzyme-Linked Immunosorbent Assay; RS, Replicative senescence; Rb, retinoblastoma; SIPS, stress-induced premature senescence; SASP, senescence associated secretory phenotype; MMPs, Matrix metalloproteinases; ROS, reactive oxygen species.
Ethics Approval
All animal experiments were performed at Beijing University of Chinese Medicine (Beijing China) according to the guidelines of the China Council on Animal Care and Use. This study was approved by the Committee of Experimental Animals of Beijing University of Chinese Medicine (approval number: BUCM-4-2022-010101-1097). Every effort was made to minimize pain and discomfort to the animals.
Funding
This work was supported by National Natural Science Foundation of China: To investigate the mechanism of acupotomy in the treatment of KOA by “regulating the tendon and treating the bone” based on hypoxic cartilage vascularization mediated by HIF-1α/VEGF pathway (No. 82074523); National Natural Youth Science Foundation of China: To investigate the mechanism of acupotomy in the cartilage protective of KOA by “regulating the tendon and treating the bone” based on premature senescence of chondrocytes regulated by YAP-FOXD1 pathway (No. 8210151833).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Yao Q, Wu X, Tao C, et al. Osteoarthritis: pathogenic signaling pathways and therapeutic targets. Signal Transduct Target Ther. 2023;8(1):56. doi:10.1038/s41392-023-01330-w
2. Hochberg MC, Guermazi A, Guehring H, et al. Effect of intra-articular sprifermin vs placebo on femorotibial joint cartilage thickness in patients with osteoarthritis: the forward randomized clinical trial. JAMA. 2019;322(14):1360–1370. doi:10.1001/jama.2019.14735
3. Xu BY, Jin Y, Ma XH, et al. The potential role of mechanically sensitive ion channels in the physiology, injury, and repair of articular cartilage. J Orthop Surg. 2020;28(3):2309499020950262. doi:10.1177/2309499020950262
4. Feng C, Yang M, Zhang Y, et al. Cyclic mechanical tension reinforces DNA damage and activates the p53-p21-Rb pathway to induce premature senescence of nucleus pulposus cells. Int J mol Med. 2018;41(6):3316–3326.
5. Huang Z, Gan S, Zhuang X, et al. Artesunate inhibits the cell growth in colorectal cancer by promoting ros-dependent cell senescence and autophagy. Cells. 2022;11(16):2472. doi:10.3390/cells11162472
6. Jiang W, Chen H, Lin Y, et al. Mechanical stress abnormalities promote chondrocyte senescence-The pathogenesis of knee osteoarthritis. Biomed Pharmacother. 2023;167:115552. doi:10.1016/j.biopha.2023.115552
7. Kung ML, Lin PY, Huang ST, et al. Zingerone nanotetramer strengthened the polypharmacological efficacy of zingerone on human hepatoma cell lines. ACS Appl Mater Interfaces. 2019;11(1):137–150. doi:10.1021/acsami.8b14559
8. Coryell PR, Diekman BO, Loeser RF. Mechanisms and therapeutic implications of cellular senescence in osteoarthritis. Nat Rev Rheumatol. 2021;17(1):47–57. doi:10.1038/s41584-020-00533-7
9. Xu M, Bradley EW, Weivoda MM, et al. Transplanted senescent cells induce an osteoarthritis-like condition in mice. J Gerontol. 2017;72(6):780–785. doi:10.1093/gerona/glw154
10. Pan JK, Zhang Y, Chen XL, et al. Effect of acupotomy on expressions of cellular senescence markers p16Ink4a and p21Waf1/Cip1 in cartilage tissue of rabbits with knee osteoarthritis. Acupuncture Res. 2023;48(7):658–665. doi:10.13702/j.1000-0607.20221091
11. Gong Z, Wang K, Chen J, et al. CircZSWIM6 mediates dysregulation of ECM and energy homeostasis in ageing chondrocytes through RPS14 post-translational modification. Clin Transl Med. 2023;13(1):e1158. doi:10.1002/ctm2.1158
12. He S, Sharpless NE. Senescence in health and disease. Cell. 2017;169(6):1000–1011. doi:10.1016/j.cell.2017.05.015
13. Tian M, Huang Y, Song Y, et al. MYSM1 suppresses cellular senescence and the aging process to prolong lifespan. Adv Sci. 2020;7(22):2001950.
14. Diekman BO, Sessions GA, Collins JA, et al. Expression of p16INK4a is a biomarker of chondrocyte aging but does not cause osteoarthritis. Aging Cell. 2018;17(4):e12771. doi:10.1111/acel.12771
15. Jeon OH, David N, Campisi J, et al. Senescent cells and osteoarthritis: a painful connection. J Clin Invest. 2018;128(4):1229–1237. doi:10.1172/JCI95147
16. Xie J, Lin J, Wei M, et al. Sustained Akt signaling in articular chondrocytes causes osteoarthritis via oxidative stress-induced senescence in mice. Bone Res. 2019;7(1):1–9. doi:10.1038/s41413-018-0036-5
17. Deng Y, Lu J, Li W, et al. Reciprocal inhibition of YAP/TAZ and NF-κB regulates osteoarthritic cartilage degradation. Nat Commun. 2018;9(1):4564. doi:10.1038/s41467-018-07022-2
18. Si H-B, Yang T-M, Li L. miR-140 attenuates the progression of early-stage osteoarthritis by retarding chondrocyte senescence. mol Ther Nucleic Acids. 2020;19:15–30. doi:10.1016/j.omtn.2019.10.032
19. Sun K, Guo J, Guo Z, et al. The roles of the Hippo-YAP signalling pathway in cartilage and osteoarthritis. Ageing Res Rev. 2023;90:102015. doi:10.1016/j.arr.2023.102015
20. Wu T, Yang Z, Chen W, et al. miR-30e-5p-mediated FOXD1 promotes cell proliferation by blocking cellular senescence and apoptosis through p21/CDK2/Rb signaling in head and neck carcinoma. Cell Death Discov. 2023;9(1):295. doi:10.1038/s41420-023-01571-2
21. Deng Y, Lu J, Li W, et al. Reciprocal inhibition of YAP/TAZ and NF-κB regulates osteoarthritic cartilage degradation. Nat Commun. 2018;9(1):1–14.
22. Fu L, Hu Y, Song M, et al. Up-regulation of FOXD1 by YAP alleviates senescence and osteoarthritis. PLoS Biol. 2019;17(4):e3000201. doi:10.1371/journal.pbio.3000201
23. Zhang Y, Li K, Ying Y, et al. C21 steroid-enriched fraction refined from marsdenia tenacissima inhibits hepatocellular carcinoma through the coordination of hippo-yap and PTEN-PI3K/AKT signaling pathways. Oncotarget. 2017;8(66):110576–110591. doi:10.18632/oncotarget.22833
24. Jing X, Yang X, Zhang W, et al. Mechanical loading induces HIF-1α expression in chondrocytes via YAP. Biotechnol Lett. 2020;42(9):1645–1654. doi:10.1007/s10529-020-02910-4
25. Fraenkel L, Buta E, Suter L, et al. Nonsteroidal anti-inflammatory drugs vs cognitive behavioral therapy for arthritis pain: a randomized withdrawal trial. JAMA Intern Med. 2020;180(9):1194–1202. doi:10.1001/jamainternmed.2020.2821
26. Jung SH, Lee W, Park SH, et al. Diclofenac impairs autophagic flux via oxidative stress and lysosomal dysfunction: implications for hepatotoxicity. Redox Biol. 2020;37:101751. doi:10.1016/j.redox.2020.101751
27. Kolasinski SL, Neogi T, Hochberg MC, et al. 2019 American college of rheumatology/arthritis foundation guideline for the management of osteoarthritis of the hand, hip, and knee. Arthritis Rheumatol. 2020;72(2):220–233. doi:10.1002/art.41142
28. Ruhdorfer A, Wirth W, Eckstein F. Association of knee pain with a reduction in thigh muscle strength–a cross-sectional analysis including 4553 osteoarthritis initiative participants. Osteoarthritis Cartilage. 2017;25(5):658–666. doi:10.1016/j.joca.2016.10.026
29. Wang Z, Xu H, Wang Z, et al. Effects of externally-applied, non-pharmacological Interventions on short- and long-term symptoms and inflammatory cytokine levels in patients with knee osteoarthritis: a systematic review and network meta-analysis. Front Immunol. 2023;14:1309751. doi:10.3389/fimmu.2023.1309751
30. Chen WH. Guidelines for TCM diagnosis and treatment of knee osteoarthritis(2020 edition). J Traditional Chine Orthopedics Traumatol. 2020;32(10):1–14.
31. Katz JN, Arant KR, Loeser RF. Diagnosis and treatment of hip and knee osteoarthritis: a review. JAMA. 2021;325(6):568–578.
32. Cho K, Kim Y, Yoon S-H. The effect of miniscalpel acupuncture and splint treatment for joint deformity of hand osteoarthritis: a case report. Integr Med Res. 2019;8(2):120–122.
33. Ma SN, Xie ZG, Guo Y, et al. Effect of acupotomy on FAK-PI3K signaling pathways in KOA rabbit articular cartilages. Evid Based Complement Alternat Med. 2017;2017:4535326. doi:10.1155/2017/4535326
34. Xing L, Chen X, Guo C, et al. Electroacupuncture exerts chondroprotective effect in knee osteoarthritis of rabbits through the mitophagy pathway. J Pain Res. 2023;16:2871–2882.
35. Liu ZF, Zhang Y, Liu J, et al. Effect of traditional Chinese non-pharmacological therapies on knee osteoarthritis: a narrative review of clinical application and mechanism. Orthop Res Rev. 2024;16:21–33. doi:10.2147/ORR.S442025
36. Lee CJ, Luo WT, Tam KW, et al. Comparison of the effects of acupotomy and acupuncture on knee osteoarthritis: a systematic review and meta-analysis. Complement Ther Clin Pract. 2023;50:101712.
37. Guo Y, Xu Y, He M, et al. Acupotomy improves synovial hypoxia, synovitis and angiogenesis in KOA rabbits. J Pain Res. 2023;16:749–760. doi:10.2147/JPR.S396955
38. Statzer C, Park JYC, Ewald CY. Extracellular matrix dynamics as an emerging yet understudied hallmark of aging and longevity. Aging Dis. 2023;14(3):670–693.
39. Chen D, Shen J, Zhao W, et al. Osteoarthritis: toward a comprehensive understanding of pathological mechanism. Bone Res. 2017;5:16044. doi:10.1038/boneres.2016.44
40. Ben-Sahra I, Manning BD. mTORC1signaling and the metabolic control of cell growth. Curr Opin Cell Biol. 2017;45:72–82.
41. Kureel SK, Blair B, Sheetz MP. Recent advancement in elimination strategies and potential rejuvenation targets of senescence. Adv Biol. 2024;8(1):e2300461.
42. Khosla S, Farr JN, Tchkonia T, et al. The role of cellular senescence in ageing and endocrine disease. Nat Rev Endocrinol. 2020;16(5):263–275. doi:10.1038/s41574-020-0335-y
43. Stojanović SD, Fuchs M, Kunz M, et al. Inflammatory drivers of cardiovascular disease: molecular characterization of senescent coronary vascular smooth muscle cells. Front Physiol. 2020;11:520. doi:10.3389/fphys.2020.00520
44. Mehana EE, Khafaga AF, El-Blehi SS. The role of matrix metalloproteinases in osteoarthritis pathogenesis: an updated review. Life Sci. 2019;234:116786. doi:10.1016/j.lfs.2019.116786
45. Hu Q, Ecker M. Overview of MMP-13 as a promising target for the treatment of osteoarthritis. Int J mol Sci. 2021;22(4):1742. doi:10.3390/ijms22041742
46. Slovacek H, Khanna R, Poredos P, et al. Interrelationship of osteopontin, MMP-9 and ADAMTS4 in patients with osteoarthritis undergoing total joint arthroplasty. Clin Appl Thromb Hemost. 2020;26:1076029620964864. doi:10.1177/1076029620964864
47. Vinatier C, Domínguez E, Guicheux J, et al. Role of the inflammation-autophagy-senescence integrative network in osteoarthritis. Front Physiol. 2018;9:706. doi:10.3389/fphys.2018.00706
48. Liu Y, Zhang Z, Li T, et al. Senescence in osteoarthritis: from mechanism to potential treatment. Arthritis Res Ther. 2022;24(1):174. doi:10.1186/s13075-022-02859-x
49. JingX YX, Zhang W, Zhang W, et al. Mechanical loading induces HIF-1α expression in chondrocytes via YAP. Biotechnol Lett. 2020;42(9):1645–1654.
50. Yao W, Ma A, Zhang Z, et al. Intermittent hydrostatic pressure promotes cartilage repair in an inflammatory environment through Hippo-YAP signaling in vitro and in vivo. Biomed Res Int. 2022;2022:3215461. doi:10.1155/2022/3215461
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms.php
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms.php
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Therapeutic Effect of Acupotomy at Sanheyang for Cartilage Collagen Damage in Moderate Knee Osteoarthritis: A Rabbit Model
Li Y, Hou Y, Sun J, Wei J, Chai Y, Guo M, Wang R
Journal of Inflammation Research 2023, 16:2241-2254
Published Date: 24 May 2023
Effect of Traditional Chinese Non-Pharmacological Therapies on Knee Osteoarthritis: A Narrative Review of Clinical Application and Mechanism
Liu ZF, Zhang Y, Liu J, Wang YY, Chen M, Liu EY, Guo JM, Wang YH, Weng ZW, Liu CX, Yu CH, Wang XY
Orthopedic Research and Reviews 2024, 16:21-33
Published Date: 26 January 2024