")
Back to Journals » Journal of Experimental Pharmacology » Volume 17
Anti-Inflammatory Activities of Some Plants of Genus Alpinia: Insights from In Vitro, In Vivo, and Human Studies
Authors Yuliawati KM, Febriyanti RM , Sumiwi SA, Levita J
Received 3 October 2024
Accepted for publication 17 December 2024
Published 24 January 2025 Volume 2025:17 Pages 51—91
DOI https://doi.org/10.2147/JEP.S499115
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 4
Editor who approved publication: Prof. Dr. Abdelwahab Omri
Kiki Mulkiya Yuliawati,1,2,* Raden Maya Febriyanti,3,* Sri Adi Sumiwi,4,* Jutti Levita4,*
1Doctoral Program in Pharmacy, Faculty of Pharmacy, Universitas Padjadjaran, Sumedang, West Java, Indonesia; 2Faculty of Mathematics and Natural Sciences, Bandung Islamic University, Bandung, Indonesia; 3Department of Biology Pharmacy, Faculty of Pharmacy, Universitas Padjadjaran, Sumedang, West Java, Indonesia; 4Department of Pharmacology and Clinical Pharmacy, Faculty of Pharmacy, Universitas Padjadjaran, Sumedang, West Java, Indonesia
*These authors contributed equally to this work
Correspondence: Jutti Levita, Department of Pharmacology and Clinical Pharmacy, Faculty of Pharmacy, Universitas Padjadjaran, Jl. Raya Bandung-Sumedang km 21, Sumedang, 45363, Indonesia, Tel +6222-84288888 Ext 3510, Email [email protected]
Abstract: This narrative review intends to provide thorough information on the anti-inflammatory activities of Alpinia plants, the largest genus of the family Zingiberaceae. The articles were searched on the PubMed database using ‘Alpinia AND anti-inflammatory activity’ as the keywords, filtered to articles published from 2020 to 2024 and free full-text. Of the approximately 248 members of the genus Alpinia plants, the most commonly studied for their anti-inflammatory activities are A. galanga, A. officinarum, A. zerumbet, and A. oxyphylla. Only A. galanga, A. officinarum, and A. zerumbet have been studied in humans. Studies in animal models revealed that the plants contributed as exogenous antioxidants, reduced proinflammatory cytokines, inhibited proinflammatory enzymes, improved gastric acid and gastrointestinal motility, and promoted ulcer healing. The terpenoids, flavonoids (such as kaempferol, quercetin, and galangin), and diarylheptanoids obtained from the rhizomes of these plants may crucially play important roles in their anti-inflammatory activities. These plants did not show toxicity toward numerous normal cell lines (RAW 264.7, IEC-6, HepG2, MT-4, NIH-3T3, Vero cells, human peripheral blood mononuclear cells, and HaCaT) but were toxic to cancer cell lines (HT29). In humans, A. galanga was studied for its effects as psychostimulants improving mental health, improving sperm motility, and erectile dysfunction. Similarly, A. officinarum could improve sperm morphology and idiopathic infertility, whereas A. zerumbet worked as a cardio-myorelaxant in patients with cardiovascular diseases.
Keywords: cytokines, flavonoids, interleukins, polyphenols, secondary metabolites, Alpinia genus
Introduction
Alpinia, the largest genus of the Zingiberaceae family, was designated by Prospero Alpino, the Italian botanist. The Alpinia genus covers approximately 248 species distributed in tropical and subtropical regions of Asia and the Pacific Ocean.1–4 Rosemary Margaret Smith, a Scottish botanist who specializes in the taxonomy of Zingiberaceae, proposed that Alpinia genus plants should be subdivided into two subgenera, Alpinia and Dieramalpinia.5 The rhizome of these plants has a spicy flavor and pungent aromatic odor distinct from other Zingiberaceae plants. Alpinia plants, often called galangal (derived from Gao-Liang-Jiang, a Chinese word), are greater galangal or Alpinia galanga (L). Willd. and lesser galangal or Alpinia officinarum Hance.6 Approximately 200 metabolites have been isolated and authenticated from plants of the genus Alpinia, including diarylheptanoids, terpenes, flavonoids, phenylpropanoids, volatile oils, and lignin. We searched for relevant scientific information to understand why these plants can cure disease.
In their phytochemical studies, Zhang and co-workers have confirmed that diterpenoids, flavonoids, and diarylheptanoids, obtained from the rhizomes, are the most prominent metabolites of the Alpinia genus.7 Diarylheptanoids (characterized by a 1.7-diphenyl heptane skeleton) are categorized into linear, cyclic, and dimeric diarylheptanoids, or diarylheptanoids bearing special moieties or in conjugation with monoterpenes or sesquiterpenes.8 Diarylheptanoids were reported for their anti-inflammatory activities by modulating several pathways, for example, by inhibiting the production of nitric oxide (NO) and downregulating tumor necrosis factor (TNF-α) in murine macrophage RAW 264.7 and microglial N-11 cells,9 by decreasing neutrophil and eosinophil counts and malondialdehyde levels, and elevating interferon (IFN)-γ, catalase (CAT), and lymphocyte values in sensitized rats.10 In addition, flavonoids (phenolic compounds with a chemical structure of C6–C3–C6 that consist of two benzene rings (A and B) connected with heterocycle pyrene ring (C), and oxygen) are subcategorized into flavone, flavonol, flavanone, flavanonol, isoflavone, chalcone, and flavonoid glycosides. The anti-inflammatory mechanism of flavonoids was announced, for example, by inhibiting the overproduction of NO, and reducing pro-inflammatory cytokines such as TNF-α, interleukin (IL)-Ιβ, IL-6, and IF-γ in lipopolysaccharide (LPS)-stimulated RAW 264.7.11 Generally, flavonoids are responsible for the antioxidant activity, due to carbonyl, aldehyde, or hydroxyl moieties. The presence of a B-ring catechol group contributes to the donation of hydrogen (electron) to stabilize a radical species. Flavonoids could alter several protein kinase and lipid kinase signaling cascades such as phosphoinositide 3-kinase (PI3-kinase), Akt/PKB, tyrosine kinases, protein kinase C (PKC), and MAP kinases signaling pathways.12
Considering that plants of the genus Alpinia are abundant in diarylheptanoids and flavonoid content, this article discusses in vitro, in vivo, and human studies on the anti-inflammatory activity of these plants. The role of cytokines in the pathogenesis of inflammation and the effects of the Alpinia plants on pro-inflammatory and anti-inflammatory cytokines are also described.
Methods
This narrative review is intended to provide thorough information on the anti-inflammatory activities of Alpinia plants, the largest genus of the family Zingiberaceae. The articles were searched on the PubMed database using the keywords “Alpinia AND anti-inflammatory activity” filtered to free full-text, article language English, and publication period from 2020 to 2024 was used and resulted in 46 articles. Articles were further screened through titles and abstracts. Articles not related to the topic of interest, articles not written in English, reviews, and duplicate articles, were excluded. Additional searches were carried out to comprehend the review.
Botanical and Phytochemical Aspects
The Alpinia genus of the family Zingiberaceae are perennial flowering plants with tuberous rhizomes native to Asia and the Pacific islands.1–4 According to Plants of the World Online, an online database published by the Royal Botanic Gardens, Kew (https://powo.science.kew.org/taxon/urn:lsid:ipni.org:names:328388-2), there are 248 species of the Alpinia genus plants. The botanical names of the Alpinia plants are listed in alphabetical order as follows:
Different parts of the Alpinia genus plants possess significant bioactive metabolites for plant-based drug candidate development, such as diarylheptanoids, terpenes, flavonoids, phenylpropanoids, volatile oil, and lignin, with flavonoids, diterpenoids, and diarylheptanoids, as the most common contributors for their bioactivity.7 It was described that A. galanga is abundant in phenolic compounds and essential oils, with a total phenolic content (TPC) ranging from 26.83 to 252.36 mg gallic acid equivalence (GAE)/g and total flavonoid content (TFC) ranging from 14.12 to 143.13 mg quercetin equivalence (QE)/g extract.13–16 The most abundant phenolic compounds in the extract were, respectively, gallic acid, catechin, quercetin (a flavonoid), catechol, isorhamnetin, trans-cinnamic acid, and protocatechuic acid.13 The variety in TPC and TFC values was caused by the differences in plant sources, with the highest values belonging to A. galanga collected from West Java, Indonesia.16 A. officinarum is rich in flavonoids and diarylheptanoids.8,17 The water extract of A. officinarum rhizome contains protocatechuic acid, epicatechin, and kaempferide.18,19 A. oxyphylla collected in South Korea was reported to include essential oils, sesquiterpenes, flavones, diarylheptanoids, glycosides, and steroids. Among those, nootkatone is the most abundant metabolite.20 The dried fruits of A. oxyphylla collected from Qiongzhong county, Hainan province, China, were assessed under the optimized extraction and UFLC-MS/MS analytical conditions and resulted in nootkatone, diphenylheptanes, and flavones.21 Conversely, little is known about the phytochemistry of A. purpurata,22 opening opportunities for further exploration. The metabolites isolated from the plants of the Alpinia genus are listed in Table 1.
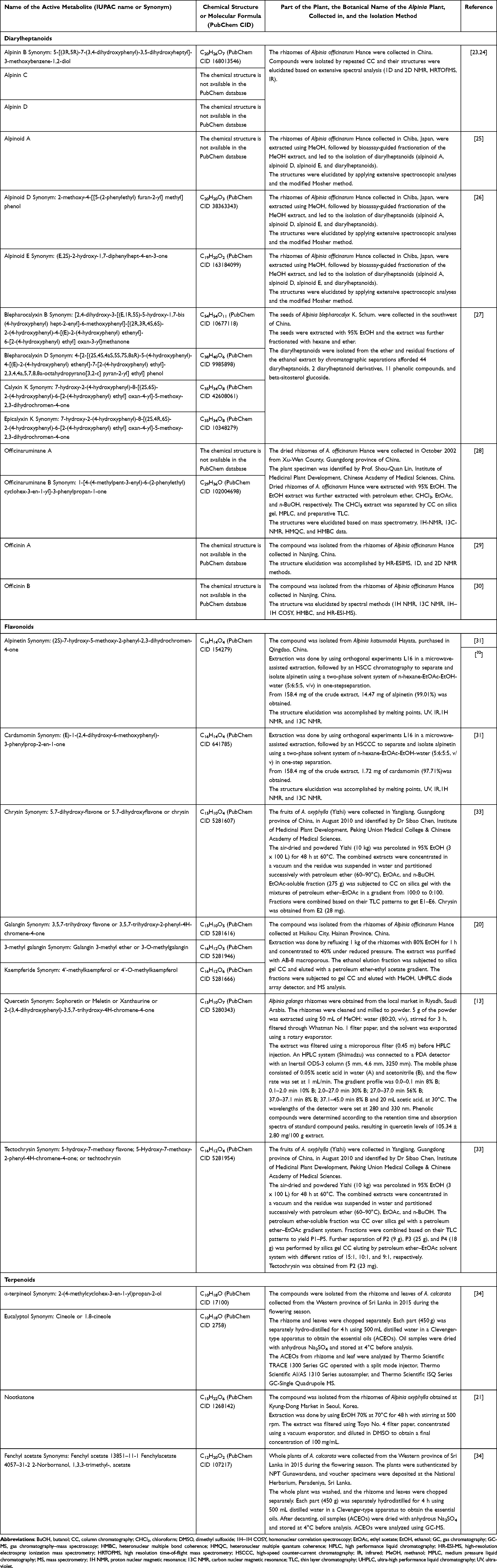 |
Table 1 Metabolites Isolated from Alpinia Genus Plants |
The Role of Cytokines in the Pathogenesis of Inflammation
Inflammation occurs when tissue injury and inflammation-inducing factors, such as histamines and cytokines, lead to blood vessel dilatation and vascular permeability, opening the entry for inflammatory components.35 Inflammatory responses play a crucial role in the host defense mechanism against threats and typically present with signs such as redness, swelling, pain, and fever. When the host encounters external triggers or tissue injury, it responds by producing pro-inflammatory cytokines such as TNF-α, interleukin-1 beta (IL-1β), IL-6, NO, and prostaglandins (PGs).36 TNF-α and IL-1β are important in initiating inflammation following tissue damage from various causes. In addition, NO and PGs contribute to inflammation via the actions of inducible nitric oxide synthase (iNOS) and cyclooxygenase-2 (COX-2), respectively. Arachidonic acid (AA), a substrate of COX, is converted to PGs by both COX-1 and COX-2, leading to the stimulation of inflammation. COX-2 is absent in healthy tissue and is only expressed in response to cellular stress or damage, triggered by the activation of growth factor pathways. Inflammation also impacts AA metabolism through lipoxygenase (LOX) activity, which promotes the synthesis of pro-inflammatory cytokines such as IL-1, IL-6, and IL-8.37 Furthermore, the nuclear factor kappa-light-chain-enhancer of activated B cell (NF-kappaB) has a particular function in inflammation through controlling how inflammatory genes, such as IL-6 and TNF-α, are expressed.38 The NF-kappaB-IkappaB complex exists in an inactive state in the cytoplasm. Upon stimulation by TNF-α, the β-subunit of the IkappaB kinase (IKK) complex, phosphorylates IkappaB proteins, and free NF-kappaB dimers will translocate to the nucleus, and begin the transcription.39 In addition to NF-kappaB, which is an activator of inflammatory genes, this protein also works by limiting the NOD-like receptor protein 3 (NLRP3) inflammasome activation and IL-1β production. The production of inflammatory mediators and cytokines, T-cell differentiation, and cell proliferation, lead to the initiation of the inflammatory pathways, such as the NF-kB pathway and JAK-STAT, and complete the inflammatory responses.40 Therefore, understanding various proteins and cytokines involved in the pathogenesis of inflammation will provide a broader insight into how immunomodulatory activity studies of plant extracts were designed.
In vitro Anti-Inflammatory Activities of Alpinia
The anti-inflammatory activities of Alpinia have been broadly reported in numerous in vitro studies, with the most reported being A. officinarum, A. oxyphylla, A. zerumbet, A. calcarata, A. galanga, and A. katsumadai (Table 2).
 |
Table 2 In Vitro and In Vivo Studies of Anti-Inflammatory Activity of Alpinia Genus Plants |
Alpinia officinarum
Alpinia officinarum, known as lesser galangal, is used to treat a wide range of symptoms including stomach ache, swelling, inflammation, and gastrointestinal ailments. The primary active phytochemical of A. officinarum is a flavonol named galangin (synonym: 3,5,7-trihydroxy flavone or 3,5,7-trihydroxy-2-phenyl-4H-chromene-4-one; molecular formula C15H10O5; PubChem CID 5281616).40,53 Galangin works at multiple target sites such as nitric oxide synthase (NOS), COX-1 and COX-2, androgen, peroxisome proliferator-activated receptor, dipeptidyl peptidase-IV, and serine/threonine-protein kinase,41 and suppresses extracellular signal-regulated kinase (ERK) and NF-kappaB-p65 phosphorylation, which leads to anti-inflammatory activity.53
Bioactive metabolites in A. officinarum, such as galangin, 3-methyl galangin (synonym: 3-O-methyl galangin or galangin 3-methyl ether; molecular formula C16H12O5; PubChem CID 5281946), and kaempferide (synonym: 4’-methyl kaempferol or 4’-O-methyl kaempferol; molecular formula C16H12O6; PubChem CID 5281666), contributed to anti-inflammatory responses.8,17 An in vitro experiment of the ethanol extract of A. officinarum rhizome, collected from Haikou County, Hainan Province, China, in October 2017, was conducted on ethanol-exposed human gastric epithelial cells (GES-1). The apoptosis rate of GES-1 cells induced by ethanol was considerably greater compared with that of the normal group. In this study, the flavonoids significantly lowered the apoptosis rate compared with that of the ethanol model group. Western blotting and quantitative polymerase chain reaction (qPCR) techniques revealed that the flavonoids increased the expression of Bcl-2 protein and gene while lowering that of Bax and caspase-3 proteins and genes. Furthermore, the results showed that the flavonoids of A. officinarum enhanced the number of cells that passed through the polycarbonate membrane and considerably reduced the degree of damage caused by ethanol in GES-1 cells.17
The effects of n-hexane extract of A. officinarum, collected from an Ayurvedic pharmacy in Chennai, India, were studied on the mRNA expression of NF-kappaB and COX-2 in cultured human colon tumor cell line HT29 (obtained from NCCS, Pune). The cells were treated with different concentrations of the extract (0.5, 1, 2.5, 5, 7.5, 10 μg), and the treated cells showed a significant decrease in the mRNA expression levels of NF-kappaB and COX-2 as compared to untreated cells.18
A. officinarum, purchased from KOCBiotech (Daejeon, Korea), was studied on human epidermal keratinocyte (HaCaT) cells. Dried A. officinarum was extracted in distilled water under circumfluence for 3 h at 100 ± 2°C, and lyophilized after filtration to obtain the water extract. The cells were stimulated with interferon-gamma (IFN-γ) and TNF-α (each 10 ng/mL). When treated with nontoxic doses of 50, 100, and 300 μg/mL of A. officinarum water extract, there was a noticeable inhibition of macrophage-derived chemokine (MDC), regulated on activated normal T-cell expressed and secreted (RANTES), interferon-induced protein of 10 kDa (IP-10), and interferon-induced T cell α-chemoattractant (I-TAC) secretion. Furthermore, the IFN-γ/TNF-α exposure on the cells which activated the STAT1 and NF-kappaB subunits (p65 and p50) in the nucleus, was inhibited by A. officinarum extract, thus suggesting that the water extract of A. officinarum exhibited anti-inflammatory effects due to its inhibitory action on MAPK phosphorylation, NF-kappaB, and STAT1.19
Alpinia oxyphylla
A. oxyphylla is abundantly grown in Asia and is widely utilized in oriental medicine. A study reported the plants, purchased from Kyung-Dong Market in Seoul, Korea, that were extracted with ethanol 70% at 70°C for 48 h with stirring at 500 rpm. The resulting A. oxyphylla extract was filtered and vacuum-concentrated, and the concentrate was diluted in dimethyl sulfoxide to obtain a final concentration of 100 mg/mL. The extract was reported could increase the nonsteroidal anti-inflammatory drug-activated gene (NAG-1) expression and suppress cyclin D1 expression levels in various human colon cancer cell lines (HCT-116, SW480, DLD-1, and HT-49).21
The ethanol extract of A. oxyphylla fruits, purchased from Ulsan, South Korea, significantly decreased the production of NO (68.2%), PGE2 (92.8%), IL-1β (77.2%), IL-6 (39.9%), and TNF-α (20.7%) and showed the activation of extracellular signal-regulated kinase (ERK), Jun N-terminal kinase (JNK), and p38 mitogen-activated protein kinase (MAPK) in lipopolysaccharide-treated RAW264.7 cells at a dose of 100 µg/mL.41
Alpinia zerumbet
A. zerumbet grows widely from East Asian countries to South America. The leaves, rhizomes, fruits, seeds, and flowers have been exploited to treat various ailments.54
The methanol extract of A. zerumbet leaves, collected from Zoo Garden, Giza, Egypt in May 2014, was reported to be successfully inhibiting COX-1 comparable to that of diclofenac, inhibiting COX-2 with half the potency of celecoxib, and inhibiting LOX with nearly double the potency of zileuton, a LOX inhibitor.42
The essential oil of A. zerumbet fruits, collected in Zhenfeng County, Guizhou province, China, in October 2013, completely prevented LPS-induced human aortic endothelial cells (HAEC) activation and inflammation in vitro, as assessed by the expression of endothelial adhesion molecules, ICAM-1 and VCAM-1. It was demonstrated that TLR4-dependent NF-kappaB signaling may be involved in the process.43
A recent study by Wang et al (2024) reported that the essential oil of A. zerumbet fruits could restrain the formation of macrophage-derived foam cells by increasing cholesterol efflux via the activation of the peroxisome proliferator-activated receptor gamma (PPARγ)-liver X receptor alpha (LXRα)-ATP-binding cassette transporters A1 (ABCA1)/G1 pathway, and decreasing the ubiquitination degradation of PPARγ. This activity was thought to be due to the direct interaction of phytochemicals to the PPARγ protein, thereby increasing the stability of the protein.34
Alpinia calcarata
The anti-inflammatory potential of A. calcarata essential oil, derived from the leaves and rhizomes, was evaluated by examining their impact on LPS-exposed NO generation in murine macrophages. Whole plants of A. calcarata were collected from the Western province of Sri Lanka in 2015 during the flowering season. The plants were washed, and the rhizomes and leaves were cut separately. Each part was separately hydro-distilled for 4 h using 500 mL distilled water to obtain the essential oils. After decanting, water in the oil samples was removed with anhydrous Na2SO4, and the essential oil was evaluated for its inhibitory towards NO production and nitrite scavenging potential in LPS-exposed RAW 264.7 cells. The essential oil demonstrated a concentration-dependent reduction in NO generation. At 50 μg/mL, the rhizome-derived essential oil displayed a maximum inhibition of 85%, whereas the leaf oil displayed 81% inhibition. L-NMMA (N-monomethyl-L-arginine acetate; empirical formula C7H16N4O2.C2H4O2), an NO inhibitor, was utilized as a positive control at a dosage of 250 μM, and after treating LPS-exposed RAW 264.7 cells with L-NMMA for 24 h, 87% NO inhibition was observed.44
NO suppresses Th1 and increases Th2 cytokine response by boosting COX activity and synthesizing proinflammatory eicosanoids. This way, NO works a role as an inflammatory mediator. In addition, NO suppresses the expression of several cytokines in different immune cells, including lymphocytes, eosinophils, and monocytes, including IL-1β, TNF-α, IL-6, and INF-γ. However, excess NO can rapidly be oxidized to reactive nitrogen oxide species and damage numerous proteins and enzymes critical for cell survival and signaling, such as JAK/STAT proteins, NF-kappaB/IkappaB pathway, MAPK, G proteins, and transcription factors. Nitration of cysteines by reactive nitrogen oxide species to these proteins may lead to their activation or inactivation.55,56
Alpinia galanga
A. galanga rhizomes, collected from Tawangmangu in Central Java Indonesia in May 2021, were reported to have strong anti-inflammatory activity in an in vitro study. The rhizomes were extracted in a maceration apparatus with 500 mL of 98% ethanol for 24 h. The ethanol extract of A. galanga rhizomes was assayed on peripheral blood mononuclear cells (PBMCs), isolated from three healthy female participants who did not receive anti-inflammatory, antioxidant, vitamins, and immunosuppressant therapy for a certain period. PBMC acute inflammation cells were stimulated by TNF-α 100 pg/mL and treatment with A. galanga significantly increased the anti-inflammatory cytokine IL-10 and transforming growth-factor-beta (TGF-β) in a concentration-dependent manner. The physiological expression levels of IL-10 might be necessary to control the inflammatory process induced by TNF-α. IL-10 inhibits the expression of pro-inflammatory cytokines including IL-6, IL-12, and TNF-α via the signal transducer and activator of the transcription 3 (STAT3) pathway. TGF-β inhibits inflammatory cytokine-induced iNOS expression in an SMAD3-dependent manner (SMAD3 proteins are mothers against decapentaplegic homolog 3 also known as SMAD family member 3).45
Hydroalcoholic extract of A. galanga downregulated the release of pro-inflammatory mediators (IL-6, TNF-α, NO, and ROS) and stimulated the release of IL-10 in LPS-exposed RAW 264.7 cells. The vital enzymes involved in inflammation (iNOS, COX-2, and MMP-9) were also downregulated by pre-treatment with hydroalcoholic extract of A. galanga. Pretreatment with hydroalcoholic extract of A. galanga significantly inhibited the phosphorylation of JNK and p38 at the protein level. Further, the extract also inhibited the nuclear translocation of NF-kappa B, thus confirming the specific inhibition of the TLR4 and JAK/STAT pathways. It is well known that the inactive form of NF-kappaB is localized in the cytosol bound to IkB, an inhibitor of NF-kappaB activation. In response to an external stimulus like LPS, IκB is phosphorylated, subsequently causing the proteasomal degradation of IκB. This is followed by the translocation of free NF-kappaB-p65 from the cytoplasm to the nucleus.46
Alpinia katsumadai
The anti-inflammatory activity of A. katsumadai was evaluated using the seed extracts. The extracts revealed potential anti-inflammatory activity by significantly decreasing the high levels of PGE2 and COX-2 induced by dental plaque bacteria–fLPS (DPB-LPS) and Porphyromonas gingivalis–LPS (PG-LPS). The dose variations used were 1 µg/mL, 5 µg/mL, and 10 µg/mL, where a higher decreasing ability was indicated by the 10 µg/mL treatment in immortalized gingival fibroblast (IGF), immortalized human oral keratinocyte (IHOK), and RAW264.7 macrophage cells.47
Vivo Anti-Inflammatory Activities of Alpinia
The anti-inflammatory activities of Alpinia plants have been reported in numerous in vivo studies presented in Table 2, with the most common being A. officinarum, A. oxyphylla, and A. zerumbet.
Alpinia officinarum
Hexane extract of A. officinarum dried rhizomes, collected from Ayurvedic pharmacy, Chennai, India, exhibited the ability to decrease the expression of inflammatory mediators in dextran sulfate sodium (DSS)-induced acute and chronic ulcerative colitis (UC) male Wistar rats. Both histological and immunological abnormalities observed in animal models of colitis are similar to those observed in human inflammatory bowel disease (IBD). UC is a chronic IBD linked to an immune-mediated gut problem. It arises in genetically susceptible individuals owing to dysregulated immune responses brought on by several environmental factors. The findings demonstrated that DSS-induced rats had higher expressions of pro-inflammatory mediators TNF-α and NF-kappaB, and serum levels of C-reactive proteins (CRP). When the hexane extract of A. officinarum co-treated rats was compared to DSS-induced acute and chronic UC rats, there was a decrease in the expression of these inflammatory mediators, and CRP levels, indicating a beneficial therapeutic effect on UC.18
A. officinarum water extract was tested for anti-inflammatory effects in atopic dermatitis (AD) in NC/Nga mice (an inbred mouse model bred in Japan at Nagoya University in 1957, frequently used as a human AD model for its biological properties). Following Dermatophagoides farina extract (DfE)-induced AD-like symptoms twice per week for 3 weeks, ear thickness was observed to be increased in NC/Nga mice. Ear thickness was evaluated twice per week using a digital caliper. The dermatitis score was determined based on edema, scarring/dryness, erythema/hemorrhage, and excoriation/erosion of the dorsal skin and ear lesions. However, oral treatment with water extract or dexamethasone (the control drug) effectively alleviated the skin complaints. Serum levels of proinflammatory chemokines, such as macrophage-derived chemokine (MDC), regulated on activation normal T-cell generated and secreted (RANTES), and IgE were considerably higher following DfE treatment than in the non-induced group. The water extract of A. officinarum or dexamethasone, when administered orally, significantly reduced MDC, RANTES, and IgE blood levels.19
Moreover, the water extract of A. officinarum rhizomes (Chinese origin) when applied on the dorsal skin of UVB-irradiated hairless SKH-1 mice, was found to dramatically attenuate the wrinkle formation and epidermal thickening in the skin, and to attenuate the expression of metalloproteinase-1a (MMP-1a) and procollagen type-1 (COL1A1), and recovered the reduction of collagen content.48
Alpinia oxyphylla
When A. oxyphylla extract was administered to rats with osteoarthritis induced by monosodium iodoacetate (MIA), serum levels of pro-cytokines LTB4, IL-1β, and IL-6, were considerably reduced. A. oxyphylla extract therapy resulted in a decrease in serum TNF-α levels but was not statistically significant. Furthermore, the control group exhibited significantly higher mRNA levels of IL-6, TNF-α, iNOS, COX-2, MMP-2, and MMP-9 in the cartilage, as well as higher expression levels of all cytokines, whereas the A. oxyphylla extract-treated groups tended to have lower values, thus suggesting that A. oxyphylla extract modulated inflammatory cytokines, lowered inflammation, and preserved cartilage in MIA-induced osteoarthritis models.41
A. oxyphylla Miq, a traditional Chinese herb, is known as Yi Zhi Ren. The extract powder of Yi Zhi Ren, obtained from Chuang Song Zong Pharmaceutical Co., Ltd. (Kaohsiung, Taiwan), was dissolved with 8 mL of double-distilled water. The aqueous extracts at the doses of 0.2 g/kg, 0.4 g/kg, and 0.8 g/kg were investigated on rats’ cerebral infarction at 1 day after 90 min of transient middle cerebral artery occlusion (MCAo) and the molecular mechanisms underlying the regulation of c-Jun N-terminal kinase (JNK)-mediated inflammatory cascades in the penumbral cortex was observed. The extracts of A. oxyphilla doses of 0.4 g and 0.8 g significantly reduced cerebral infarction (both p < 0.05), attenuated neurological deficits, and suppressed the expression of phospho-apoptosis signal-regulating kinase 1 (p-ASK1)/ASK1, tumor necrosis factor receptor-associated factor 3 (TRAF3), TRAF3-interacting JNK-activating modulator (T3JAM), ionized calcium-binding adapter molecule 1 (Iba1), p-JNK/JNK, iNOS, COX-2, TNF-α, TLR4, glial fibrillary acidic protein (GFAP), NF-kappaB, and IL-6 in the penumbral cortex at 1 day after reperfusion.49
The water extract of A. oxyphylla fructus administered orally for 3 weeks to a vascular disorder-related overactive bladder spontaneous hypertensive rats, significantly improved the bladder storage parameters, tightened the detrusor layer, reduced inflammatory infiltration, and decreased collagen proportion in the rats’ bladder. These effects were caused by the reduction of TGFβ1, p-SMAD3, collagen III, Gq, and PLCβ1, involved in collagen synthesis and calcium signaling pathways. It was predicted that sitosterol, chrysin, and nootkatone were responsible for these pharmacological activities.50
Alpinia zerumbet
Some parts of A. zerumbet are also known to have anti-inflammatory properties. The methanol leaf extract elicited an inflammatory response in rats, as evidenced by an increase in paw thickness evaluated hourly for 5 h and 24 h after injection. Rats pretreated with the extract (200 and 400 mg/kg, per oral) 1 h earlier showed a dose-dependent reduction in edema thickness values by 33 and 55% of the control values, respectively. The treatment of the latter dose was stronger than that shown in rats given with the usual anti-inflammatory drug, diclofenac (20 mg/kg, orally), which only reduced edema thickness by 40% compared to control rats. Thirty-seven compounds were characterized in compound profiling using high-performance liquid chromatography in tandem with an electro-spray ionization-double mass spectroscopy/mass spectroscopy (HPLC-ESI-MS/MS). These compounds consist of flavonoids (aglycones and glycosides) and benzoic and cinnamic acid derivatives.42
Moreover, essential oils of A. zerumbet, collected in Zhenfeng County, Guizhou Province, China, in October 2013, reduced the expression of endothelial intercellular adhesion molecules (ICAM-1 and VCAM-1) produced by endothelial cells, and activated NF-kappa B signaling, making it an effective anti-inflammatory drug. Endothelial cells play an important role in inflammatory responses to infection and other stressors by producing adhesion molecules, which activate and recruit circulating leukocytes to the site of tissue inflammation. NF-kappa B signaling controls endothelial cell activation and inflammation. LPS activated NF-kappa B signaling, as evidenced by enhanced phosphorylation and nuclear translocation of p65. Treatment with essential oils of A. zerumbet eliminated these symptoms.43
Essential oil of dry and ripe fruit A. zerumbet showed the ability to reduce inflammatory cytokines TNF-α, IL-6, and IL-1β serum levels in HFD ApoE−/− mice, as an animal model on atherosclerosis, which is characterized by chronic and progressive inflammation. In the reduction of TNF-α and IL-1β expression, EOFAZ showed a dose-dependent manner, whereas, in the reduction of IL-6 expression, a higher dose of EOFAZ exhibited a stronger effect compared to the positive drug atorvastatin.34
Alpinia purpurata
The ethyl acetate extract of A. purpurata leaves, collected from Kanyakumari, Tamil Nadu, India, revealed potential anti-inflammation and anti-cancer against N-methyl N-nitrosourea (MNU) and testosterone-induced prostate cancer in rats. The animals were divided into five groups: (1) normal healthy rats; (2) rats in the negative control, that were intraperitoneally injected with testosterone (T) at a daily dose of 100 mg/kg for 3 days to induce prostate cancer; (3) rats in the positive control group, that were treated as group 2 along with finasteride at a dose of 25 mg/kg, supplemented for 2 months; (4) rats at dose 1 of ethyl acetate extract of A. purpurata, that were treated as group 2 and simultaneously treated with a daily dose of 200 mg/kg of the extract through oral gavage for 2 months; and (5) rats at dose 2 of ethyl acetate extract of A. purpurata, that received a daily dose of 200 mg/kg of the extract through oral gavage for 2 months. Histopathological observation of hepatic tissue in the normal group demonstrated normal morphology of hepatocytes surrounding periportal lobules. Rats in group 2 (prostate cancer-induced) showed periportal inflammatory cell infiltration with sinusoidal congestion and the regeneration of fatty cell changes with hepatic cell necrosis. Rats treated with the standard drug finasteride (group 3) confirmed minimal hemorrhagic necrosis in focal areas of hepatocytes. Rats in group 4 showed minimal congestion in focal areas with minimal inflammatory sites of hepatocytes, thus confirming that treatment with A. purpurata could improve the morphological appearances of the hepatic tissue with minimal hemorrhagic necrosis in focal areas of hepatocytes when compared to the negative control group.51
Alpinia galanga
The anti-inflammatory activity of the ethanolic extract of A. galanga was studied in rats with carrageenan-induced pleurisy. This pleurisy model is considered an excellent acute inflammatory model in which fluid extravasations, leukocyte migration, and the various biochemical parameters involved in inflammatory response can be measured easily in the exudates. The extract of A. galanga rhizome reduced the exudation effect in the inflammatory edema in a dose-dependent manner (100–400 mg/kg) and A. galanga 200 mg/kg and 400 mg/kg significantly inhibited the total leukocyte influx.52
Cytotoxicity Studies of Alpinia
It is known that there are several Alpinia species whose toxicity has been tested both in vitro and in vivo. Cytotoxicity analysis is necessary to identify the safe and bioactive concentrations in mammalian cells for oral and topical curative applications.
Rhizomes of A. officinarum purchased from the Kyungdong oriental medicine market, Seoul, Korea, were extracted with 99.8% methanol for 72 h at room temperature. Extracts were rotary-evaporated and the thick extract was dissolved in dimethylsulfoxide. The extract could inhibit MCF-7 cell proliferation in a dose- and time‑dependent manner by suppressing the expression levels of S-phase cell cycle regulatory proteins, including E2F1, cyclin‑dependent protein kinase 2, and cyclin A.57
A. officinarum rhizomes obtained from Korea Plant Extract Bank (KPEB, Daejeon, Korea) were dried, powdered, and extracted with distilled water (1 L for 2.5 h at 100°C). The supernatant was filtered, concentrated, and lyophilized. The water extracts were cytotoxicity tested on UVB-irradiated NIH-3T3 cells, the fibroblast cell lines that were isolated from a mouse NIH/Swiss embryo. The administration of water extracts of A. officinarum rhizome up to 100 µg/mL did not show cytotoxicity on NIH-3T3 cells.48
A. eremochlamys K. Schum. was collected from Lore Lindu National Park Central Sulawesi, Indonesia, in April 2019. The leaves, pseudostems, and rhizomes were washed, cut, and dried at room temperature with no direct sunlight. Approximately 2.0 kg of each part of the plant was extracted using 5 L of 96% ethanol for 3 × 24 h, filtered, and rotary evaporated. The thick extracts were dissolved in 100% DMSO. The extracts were screened for toxicity in human T lymphocyte (MT-4) cells. It was confirmed that the rhizome extract showed no toxicity in MT-4 cells.58
A. galanga rhizomes, collected from Tawangmangu, Central Java, Indonesia in May 2021, were cleaned, dried, and extracted with 500 mL 98% ethanol for 24 h. A. galanga extract was cytotoxicity assayed on Vero cells, which are normal kidney epithelial cells extracted from an African green monkey. The high IC50 value of A. galanga extract in Vero cells of 1120 µg/mL indicated no cytotoxic effect.45 Fresh rhizomes of A. galanga collected from the medicinal plant garden of Chiang Mai University, Chiang Mai, Thailand in February 2018, were subjected to hydro-distillation for 3 h to obtain the oil. A. galanga oil was investigated for its cytotoxicity on human peripheral blood mononuclear cells (PBMCs). The oil exposure to PBMCs resulted in > 80% survival rates for PBMCs throughout all concentration ranges, according to dose–response curve analysis, indicating no cytotoxicity to human cells.59
A. malaccensis rhizomes were collected from the medicinal garden of Nature Secret (Pvt) Ltd, Millewa, Horana, Sri Lanka. Different concentrations of A. malaccensis hexane extract were cytotoxicity-assayed on A549, HepG2, 3T3, and COS-7 cell lines and revealed that the extract was nontoxic at doses of 2, 1.4, 30, and 1.4 µg/mL for A549, HepG2, 3T3, and COS-7 cells, with no apoptotic/necrotic cell death or DNA damage seen.60
A. oxyphylla petroleum ether fractions were determined in human hepatocellular carcinoma (HCC) cell lines (BEL-7402, HepG2, SMMC-7721, Hep3B), and a human liver cell line, HL-7702. The study found that petroleum ether fraction exhibited cytotoxic effects on HL-7702 cells at a concentration of 100 mg/mL which differed considerably from the survival rate of Hep3B cells.61
Toxicity Studies of Alpinia
Toxicity studies of several Alpinia genus plants in animal models were also reported. A. galanga rhizome water extract, purchased from Chemiloids Pvt. Ltd, Vijayawada, A.P, India, was reported safe as proven by the acute toxicity study using Wistar rats of both males and females. The rats were orally administered a dose of 2000 mg/kg BW and monitored individually over the first 4 h continuously, and then infrequently for another 4 h until 24 h after. After 24 and 72 h, the rats were observed for any lethality or death. Since no animals perished, LD50 of the test drug can be taken in amounts larger than 2000 mg/kg.61
The acute toxicity of the crude n-hexane extract of A. malaccensis rhizome was assessed in three-month-old female Wistar rats. The rats were administered a single oral dose of the extract at 300 or 2000 mg/kg BW, following the modified OECD Test Guidelines. The dose of 2000 mg/kg BW resulted in no notable toxicity or fatality. After 14 days of treatment, no changes were found in behavior, BW, hematological and biochemical markers, or histopathological characteristics as compared to the control group (treated with olive oil), thus confirming the safety of oral dose for A. malaccensis extract at 2000 mg/kg BW.60
An in vivo toxicity investigation of A. oxyphylla petroleum ether fraction in mice was reported by Hui et al (2019). A. oxyphylla fruits were bought from the Yonggang Decoction Pieces Factory Co., Ltd. (Bozhou, China). The fruits were reflux-extracted with 60 L 95% ethanol for 2 h, and the above steps were repeated 3x. The extracts were concentrated at 60°C in a rotary evaporator to obtain 375 g of crude extract. The extracts were suspended in 2.5 L water and further fractionated with the same amount of petroleum ether. The petroleum ether layer was separated and evaporated. The petroleum ether fraction exhibited no evident hepatotoxicity or nephrotoxicity.62
Studies in Humans
A literature search of clinical trials of Alpinia genus plants at the time of writing found only seven articles, with five of them studied on A. galanga and the rest on A. zerumbet and A. officinarum (Table 3).
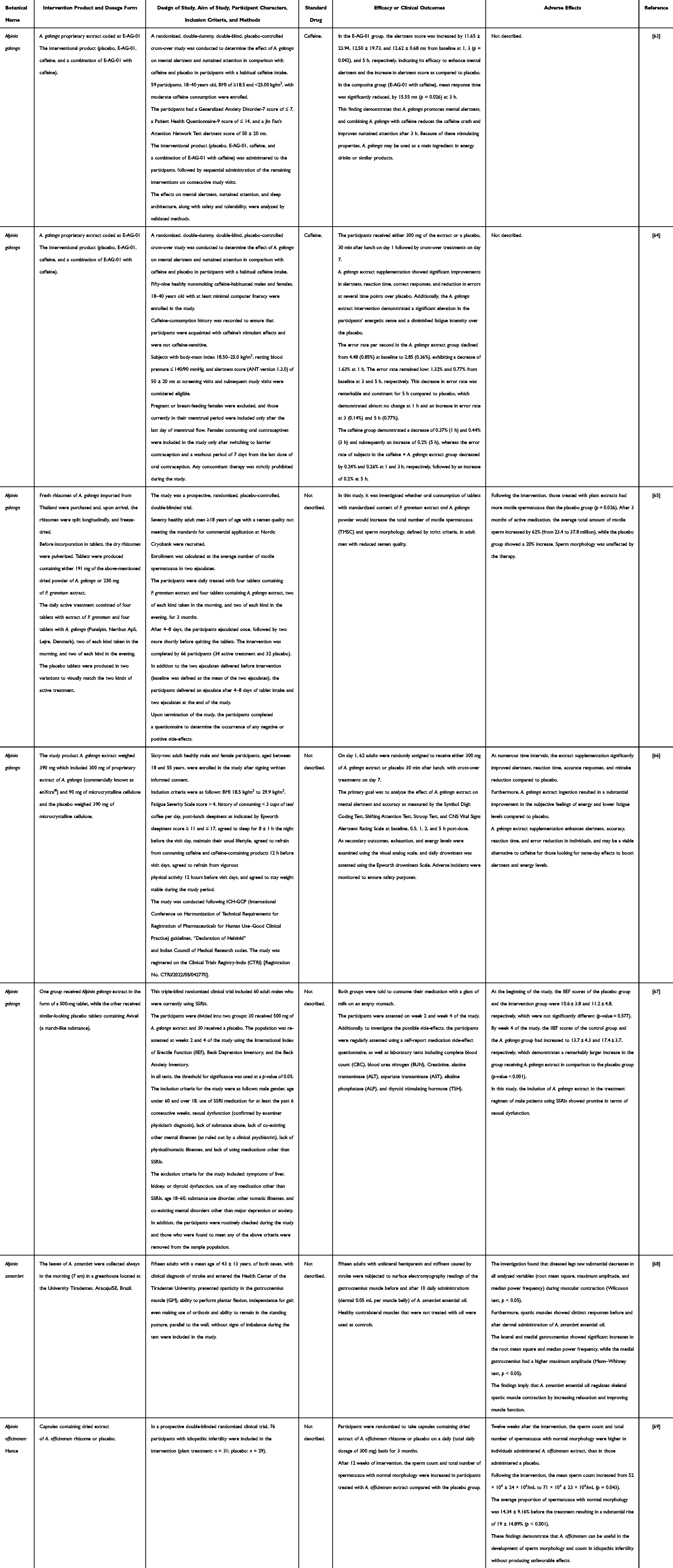 |
Table 3 Human Studies of Alpinia Genus Plants |
Alpinia galanga
A study was conducted to determine the effect of A. galanga proprietary extract on its possible psychostimulant effects in humans. The interventional product (placebo, A. galanga proprietary extract coded as E-AG-01, caffeine, and a combination of E-AG-01 with caffeine), was given to 59 participants aged 18–40 years, with body mass index of ≥ 18.5 and < 25.00 kg/m2, and with moderate caffeine consumption, followed by sequential administration of the remaining interventions on subsequent study visits. The effects on mental alertness, sustained attention, sleep architecture, safety, and tolerability, were assessed using validated methods. The E-AG-01 group enhanced their alertness score by 11.65 ± 23.94, 12.50 ± 19.73, and 12.62 ± 0.68 ms from baseline at 1, 3 (p = 0.042), and 5 h, respectively, showing its efficacy in enhancing mental alertness. At 3 h, the composite group (E-AG-01 plus caffeine) had a significantly lower mean response time of 15.55 ms (p = 0.026). This finding demonstrates that A. galanga promotes mental alertness, and combining A. galanga with caffeine reduces the caffeine crash and improves sustained attention after 3 h. Because of these stimulating properties, A. galanga may be used as a main ingredient in energy drinks or similar products.63
In another study, A. galanga extract (the source and extraction procedure were not described) was studied for its effect on mental alertness, accuracy, and fatigue through a randomized, double-blind, placebo-controlled, cross-over clinical study. Of 124 participants screened for the study, 59 meeting the protocol-defined inclusion criteria were enrolled in the study. The participants received either 300 mg of the extract or a placebo, 30 min after lunch on day 1 followed by cross-over treatments on day 7. A. galanga extract supplementation showed significant improvements in alertness, reaction time, correct responses, and reduction in errors at several time points over placebo. Additionally, the A. galanga extract intervention demonstrated a significant elevation in the participants’ energetic sense and a diminished fatigue intensity over the placebo.64
A. galanga has also been shown to improve sperm motility. This study was planned as a prospective, randomized, controlled, double-blind trial. Fresh rhizomes of A. galanga imported from Thailand were purchased and, upon arrival, the rhizomes were split longitudinally, and freeze-dried. Before incorporation in tablets, the dry rhizomes were pulverized. Tablets were produced containing either 191 mg of the above-mentioned dried powder of A. galanga or 250 mg of P. granatum extract. Seventy healthy adult men ≥ 18 years of age with a semen quality not meeting the standards for commercial application at Nordic Cryobank were recruited. Enrollment was calculated as the average number of motile spermatozoa in two ejaculates. The participants were daily treated with four tablets containing P. granatum extract and four tablets containing A. galanga extract, two of each kind taken in the morning, and two of each kind in the evening, for 3 months. After 4–8 days, the participants ejaculated once, followed by two more shortly before quitting the tablets. The intervention was completed by 66 participants (34 active treatment and 32 placebo). Following the intervention, those treated with plant extracts had more motile spermatozoa than the placebo group (p = 0.026). After 3 months of active medication, the average total amount of motile sperm increased by 62% (from 23.4 to 37.8 million), while the placebo group showed a 20% increase. Sperm morphology was unaffected by the therapy.65
A. galanga extract has been used in clinical studies to improve mental health. A randomized, double-blind, placebo-controlled, cross-over clinical trial involving healthy human participants was conducted to assess the acute effects of A. galanga extract on mental alertness, accuracy, and weariness. On day 1, 62 adults were randomly assigned to receive either 300 mg of A. galanga extract or placebo 30 min after lunch, with cross-over treatments on day 7. The primary goal was to analyze the effect of A. galanga extract on mental alertness and accuracy as measured by the Symbol Digit Coding Test, Shifting Attention Test, Stroop Test, and CNS Vital Signs Alertness Rating Scale at baseline, 0.5, 1, 2, and 5 h post-dose. As secondary outcomes, exhaustion, and energy levels were examined using the visual analog scale, and daily drowsiness was assessed using the Epworth drowsiness Scale. Adverse incidents were monitored to ensure safety purposes. At numerous time intervals, the extract supplementation significantly improved alertness, reaction time, accurate responses, and mistake reduction compared to placebo. Furthermore, A. galanga extract ingestion resulted in a substantial improvement in the subjective feelings of energy and lower fatigue levels compared to placebo. A. galanga extract supplementation enhances alertness, accuracy, reaction time, and error reduction in individuals, and may be a viable alternative to caffeine for those looking for same-day effects to boost alertness and energy levels.66
Another study aimed to determine whether adding A. galanga extract to the treatment regimen of adult males with selective serotonin reuptake inhibitors (SSRIs) could improve SSRI-induced erectile dysfunction. This triple-blind randomized clinical trial included 60 adult males who were currently using SSRIs. The participants were divided into two groups: 30 received 500 mg of A. galanga extract and 30 received a placebo. The population was re-assessed at weeks 2 and 4 of the study using the International Index of Erectile Function (IIEF), Beck Depression Inventory, and the Beck Anxiety Inventory. In all tests, the threshold for significance was used at a p-value of 0.05. The IIEF scores were significantly higher in the group receiving A. galanga extract than in the placebo group (p < 0.001). In this study, the inclusion of A. galanga extract in the treatment regimen of male patients using SSRIs showed promise in terms of sexual dysfunction. If validated, similar findings can help both patients and therapists to develop and implement better treatment programs with more positive outcomes.67
Alpinia zerumbet
A. zerumbet essential oil has myorelaxant and antispasmodic properties for cardiac and smooth muscle. A previous study examines the effect of A. zerumbet essential oil on skeletal muscle contractions in patients with post-stroke spasticity. Fifteen adults with unilateral hemiparesis and stiffness caused by stroke were subjected to surface electromyography readings of the gastrocnemius muscle before and after 10 daily administrations (dermal 0.05 mL per muscle belly) of A. zerumbet essential oil. Healthy contralateral muscles that were not treated with oil were used as controls. The investigation found that diseased legs saw substantial decreases in all analyzed variables (root mean square, maximum amplitude, and median power frequency) during muscular contraction (Wilcoxon test, p < 0.05). Furthermore, spastic muscles showed distinct responses before and after dermal administration of A. zerumbet essential oil. The lateral and medial gastrocnemius showed significant increases in the root mean square and median power frequency, while the medial gastrocnemius had a higher maximum amplitude (Mann–Whitney test, p < 0.05). The findings imply that A. zerumbet essential oil regulates skeletal spastic muscle contraction by increasing relaxation and improving muscle function. Thus, the A. zerumbet essential oil may be effective in the clinical management of secondary effects in individuals with cerebral vascular disease.68
Alpinia officinarum
This study aimed to determine the effect of A. officinarum on semen analysis in men suffering from idiopathic infertility. In this clinical research, 76 people with idiopathic infertility were enrolled in the intervention (plant treatment: 31 and placebo: 29). The participants were randomly assigned to receive capsules containing dried extract of A. officinarum rhizome or placebo daily (total daily dosage of 300 mg) for 3 months. Twelve weeks after the intervention, the sperm count and total number of spermatozoa with normal morphology were higher in individuals administered A. officinarum extract than in those administered a placebo. Following the intervention, the mean sperm count increased from 52 × 106 ± 24 × 106/mL to 71 × 106 ± 23 × 106/mL (p = 0.043). The average proportion of spermatozoa with normal morphology was 14.34 ± 9.16% before the treatment resulting in a substantial rise of 19 ± 14.89% (p < 0.001). These findings demonstrate that A. officinarum can be useful in the development of sperm morphology and count in idiopathic infertility without producing unfavorable effects.69
Herbal Drug–Drug Interaction
The administration of herbal drugs in combination with therapeutic drugs may raise the potential for pharmacokinetic or pharmacodynamic interactions. Pharmacokinetic herbal–drug interactions occur due to altered absorption, metabolism, distribution, and excretion of the therapeutic drugs.70 It was described that the oral administration of A. officinarum significantly altered the pharmacokinetic parameters of indomethacin by decreasing the systemic exposure of indomethacin and increasing its elimination.23 Another species, A. galanga, was reported for its possible interaction with warfarin by increasing the warfarin effect.24
Alpinia plants may increase the release of hydrochloric acid in the stomach, thus decreasing the effectiveness of some medications such as histamine H2 receptor antagonists, including cimetidine (Tagamet), ranitidine (Zantac), nizatidine (Axid), and famotidine (Pepcid). (https://www.rxlist.com/supplements/alpinia).
Conclusion
Our study unveiled that only A. galanga, A. officinarum, A. zerumbet, and A. oxyphylla have been the most reported for their anti-inflammatory activity in numerous in vitro and in vivo studies. Alpinia plants have the potential as exogenous antioxidants, can reduce proinflammatory cytokines such as tumor necrosis-alpha and interleukins, inhibit proinflammatory enzymes such as cyclooxygenases and inducible nitric oxide synthase, improve gastric acid and gastrointestinal motility, and promote ulcer healing, by which their diterpenoids, flavonoids, and diarylheptanoids contents may crucially contribute. These plants did not show toxicity toward numerous normal cells or animal models. During the study period, we found only A. galanga, A. officinarum, and A. zerumbet have been used to treat humans. Of the three plants, A. galanga was the most studied for psychostimulant effects, sperm motility, and erectile dysfunction, with the fewest adverse effects. However, the use of supplements containing Alpinia extract should be monitored for patients treated with warfarin, NSAIDs, and histamine H2 receptor antagonists, to avert the herbal–drug interactions.
Acknowledgments
The authors thank the Rector of Universitas Padjadjaran, West Java, Indonesia for facilitating the APC of this article. The present study was conducted in the framework of the dissertation project of the first author at the Doctoral Program in Pharmacy, Faculty of Pharmacy, Universitas Padjadjaran, West Java, Indonesia.
Disclosure
The authors state that they have no conflicts of interest in this work.
References
1. Kress WJ, Liu AZ, Newman M, Li QJ. The molecular phylogeny of Alpinia (Zingiberaceae): a complex and polyphyletic genus of gingers. Am J Bot. 2005;92(1):167–178. doi:10.3732/ajb.92.1.167
2. Tushar, Basak S, Sarma GC, Rangan L, Rangan L. Ethnomedical uses of Zingiberaceous plants of Northeast India. J Ethnopharmacol. 2010;132(1):286–296. doi:10.1016/j.jep.2010.08.032
3. Ghosh S, Rangan L. Alpinia: the gold mine of future therapeutics. Biotech. 2013;3(3):173–185. doi:10.1007/s13205-012-0089-x
4. Roxb A. Plants of the World Online. Aavilable from:: https://powo.science.kew.org/taxon/urn:lsid:ipni.org:names:328388-2.
5. Smith RM. Alpinia (Zingiberaceae): a proposed new infrageneric classification. Edinb J Bot. 1990;47(1):1–75. doi:10.1017/S0960428600003140
6. Zhou Y-Q, Liu H, He M-X, et al. Chapter 11 - A Review of the Botany, Phytochemical, and Pharmacological Properties of Galangal. In: Natural and Artificial Flavoring Agents and Food Dyes, Handbook of Food Bioengineering. Academic Press; 2018:351–396. doi:10.1016/B978-0-12-811518-3.00011-9
7. Zhang WJ, Luo JG, Kong LY. The genus Alpinia: a review of its phytochemistry and pharmacology. World J Tradit Chin Med. 2016;2(1):26–41. doi:10.15806/j.issn.2311-8571.2015.0026
8. Sun Y, Kurokawa M, Miura M, Kakegawa T, Motohashi S, Yasukawa K. Bioactivity and synthesis of diarylheptanoids from Alpinia officinarum. Stud Nat Prod Chem. 2016;49:157–187.
9. Raju R, Singh A, Gunawardena D, Reddell P, Münch G. Diarylheptanoids with anti-inflammatory activity from the rhizomes of Pleuranthodium racemigerum (Zingiberaceae). Phytochem Lett. 2019;30:10–13. doi:10.1016/j.phytol.2019.01.004
10. Shakeri F, Boskabady MH. Anti-inflammatory, antioxidant, and immunomodulatory effects of curcumin in ovalbumin-sensitized rat. Biofactors. 2017;43(4):567–576. doi:10.1002/biof.1364
11. Guo Z, Xu HY, Xu L, Wang SS, Zhang XM, Zhang X-M. In vivo and in vitro immunomodulatory and anti-inflammatory effects of total flavonoids of Astragalus. Afr J Tradit Complement Altern Med. 2016;13(4):60–73. doi:10.21010/ajtcam.v13i4.10
12. Williams RJ, Spencer JP, Rice-Evans C. Flavonoids: antioxidants or signaling molecules? Free Radic Biol Med. 2004;36(7):838–849. doi:10.1016/j.freeradbiomed.2004.01.001
13. Aljobair MO. Chemical composition, antimicrobial properties, and antioxidant activity of galangal rhizome. Food Sci Technol. 2022;
14. Guneidy RA, Zaki ER, Gad AAM, Saleh NSE, Shokeer A. Evaluation of phenolic content diversity along with antioxidant/pro-oxidant, glutathione transferase inhibition, and cytotoxic potential of selected commonly used plants. Prev Nutr Food Sci. 2022;27(3):282–298. doi:10.3746/pnf.2022.27.3.282
15. Aziz IM, Alfuraydi AA, Almarfadi OM, et al. Phytochemical analysis, antioxidant, anticancer, and antibacterial potential of Alpinia galanga (L.) rhizome. Heliyon. 2024;10(17):e37196. doi:10.1016/j.heliyon.2024.e37196
16. Mutakin, Saptarini NM, Amalia R, Sumiwi SA, Megantara S, Saputri FA, Levita J. Molecular docking simulation of phenolics towards tyrosinase, phenolic content, and radical scavenging activity of some Zingiberaceae plant extracts. Cosmetics. 2023;10(6):149. doi:10.3390/cosmetics10060149
17. Lin K, Wang Y, Gong J, Tan Y, Deng T, Wei N. Protective effects of total flavonoids from Alpinia officinarum rhizoma against ethanol-induced gastric ulcer in vivo and in vitro. Pharm Biol. 2020;58(1):854–862. doi:10.1080/13880209.2020.1803370
18. Rajendiran V, Natarajan V, Devaraj SN. Anti-inflammatory activity of Alpinia officinarum Hance on rat colon inflammation and tissue damage in DSS induced acute and chronic colitis models. Food Sci Hum Wellness. 2018;7(4):273–281.doi:10.1016/j.fshw.2018.10.004.
19. Song HK, Park SH, Kim HJ, Jang S, Kim T. Alpinia officinarum water extract inhibits the atopic dermatitis-like responses in NC/Nga mice by regulation of inflammatory chemokine production. Biomed Pharmacother. 2021;144:112322. doi:10.1016/j.biopha.2021.112322
20. Lin K, Deng T, Qu H, et al. Gastric protective effect of Alpinia officinarum flavonoids: mediating TLR4/NF-κB and TRPV1 signaling pathways and gastric mucosal healing. Pharm Biol. 2023;61(1):50–60. doi:10.1080/13880209.2022.2152058
21. Yoo E, Lee J, Lertpatipanpong P, et al. Anti-proliferative activity of A. oxyphylla and its bioactive constituent nootkatone in colorectal cancer cells. BMC Cancer. 2020;20(1):881. doi:10.1186/s12885-020-07379-y
22. Chan EWC, Wong SK. Phytochemistry and pharmacology of ornamental gingers, Hedychium coronarium, and Alpinia purpurata: a review. J Integr Med. 2015;13(6):368–379. doi:10.1016/S2095-4964(15)60208-4
23. Zhang X, Xie Z, Chen X, et al. Herb-drug interaction in the protective effect of Alpinia officinarum against gastric injury induced by indomethacin based on pharmacokinetic, tissue distribution and excretion studies in rats. J Pharm Anal. 2021;11(2):200–209. doi:10.1016/j.jpha.2020.05.009
24. Leite PM, Martins MAP, Das Graças Carvalho M, Castilho RO. Mechanisms and interactions in concomitant use of herbs and warfarin therapy: an updated review. Biomed Pharmacother. 2021;143:112103. doi:10.1016/j.biopha.2021.112103
25. Liu D, Qu W, Zhao L, Guan FQ, Liang JY. A new dimeric diarylheptanoid from the rhizomes of Alpinia officinarum. Chin J Nat Med. 2014;12(2):139–141. doi:10.1016/S1875-5364(14)60022-4
26. Liu D, Liu YW, Guan FQ, Liang JY. New cytotoxic diarylheptanoids from the rhizomes of Alpinia officinarum Hance. Fitoterapia. 2014;96:76–80. doi:10.1016/j.fitote.2014.04.008
27. Sun Y, Matsubara H, Kitanaka S, Yasukawa K. Diarylheptanoids from the Rhizomes of Alpinia officinarum. Helv Chim Acta. 2008;911(1):118–123. doi:10.1002/hlca.200890001
28. Sun Y, Tabata K, Matsubara H, Kitanaka S, Suzuki T, Yasukawa K. New cytotoxic diarylheptanoids from the rhizomes of Alpinia officinarum. Planta Med. 2008;74(4):427–431. doi:10.1055/s-2008-1034345
29. Ali MS, Banskota AH, Tezuka Y, Saiki I, Kadota S. Antiproliferative activity of diarylheptanoids from the seeds of Alpinia blepharocalyx. Biol Pharm Bull. 2001;24(5):525–528. doi:10.1248/bpb.24.525
30. An N, Zhang H-W, Xu L-Z, Yang S-L, Zou Z-M. New diarylheptanoids from the rhizome of Alpinia officinarum Hance. Food Chemistry. 2010;119(2):513–517. doi:10.1016/j.foodchem.2009.06.046
31. Zhao L, Liang JY, Zhang JY, Chen Y. A novel diarylheptanoid bearing flavonol moiety from the rhizomes of Alpinia officinarum Hance. Chin Chem Lett. 2010;21(2):194–196. doi:10.1016/j.cclet.2009.09.011
32. Zhao L, Liang JY, Qu W. A novel dimeric diarylheptanoid from the rhizomes of Alpinia officinarum. Chin Nat Comp. 2012;48(5):836–838. doi:10.1016/j.cclet.2011.11.013
33. Lü HT, Zou YL, Deng R, Shan R. Extraction, purification and antiradical activities of alpinetin and cardamomin from Alpinia katsumadai Hayata. Asian J Chem. 2013;25(17):9503–9507. doi:10.14233/ajchem.2013.15046
34. Wang SQ, Xiang J, Zhang GQ, et al. Essential oil from fructus Alpinia zerumbet ameliorates atherosclerosis by activating PPARγ-LXRα-ABCA1/G1 signaling pathway. Phytomedicine. 2024;123:155227. doi:10.1016/j.phymed.2023.155227
35. Alharbi KS, Alenezi SK, Gupta G. Chapter 1-Pathophysiology and pathogenesis of inflammation. In: Recent Developments in Anti-Inflammatory Therapy. Academic Press; 2023:1–9. doi:10.1016/B978-0-323-99988-5.00006-1
36. Goldring MB, Otero M. Inflammation in osteoarthritis. Curr Opin Rheumatol. 2011;23(5):471–478. doi:10.1097/bor.0b013e328349c2b1
37. Yu SH, Kim HJ, Jeon SY, et al. Anti-inflammatory and anti-nociceptive activities of Alpinia oxyphylla Miquel extracts in animal models. J Ethnopharmacol. 2020;260:112985. doi:10.1016/j.jep.2020.112985
38. Hayden MS, Ghosh S. Signaling to NF-kappaB. Genes Dev. 2004;18(18):2195–2224. doi:10.1101/gad.1228704
39. Albensi BC. What is nuclear factor kappa B (NF-κB) doing in and to the mitochondrion? Front Cell Dev Biol. 2019;7:154. doi:10.3389/fcell.2019.00154
40. Thapa R, Afzal O, Altamimi ASA, et al. Galangin as an inflammatory response modulator: an updated overview and therapeutic potential. Chem Biol Inter. 2023;378:110482. doi:10.1016/j.cbi.2023.110482
41. Lee YM, Son E, Kim SH, Kim DS. Effect of Alpinia oxyphylla extract in vitro and in a monosodium iodoacetate-induced osteoarthritis rat model. Phytomedicine. 2019;65:153095. doi:10.1016/j.phymed.2019.153095
42. Ghareeb MA, Sobeh M, Rezq S, El-Shazly AM, Mahmoud MF, Wink M. HPLC-ESI-MS/MS profiling of polyphenolics of a leaf extract from Alpinia zerumbet (Zingiberaceae) and its anti-inflammatory, anti-nociceptive, and antipyretic activities in vivo. Molecules. 2018;23(12):3238. doi:10.3390/molecules23123238
43. Ji YP, Shi TY, Zhang YY, et al. Essential oil from fructus Alpinia zerumbet (fruit of Alpinia zerumbet (Pers.) Burtt. et Smith) protected against aortic endothelial cell injury and inflammation in vitro and in vivo. J Ethnopharmacol. 2019;237:149–158. doi:10.1016/j.jep.2019.03.011
44. Chandrakanthan M, Handunnetti SM, Premakumara GSA, Kathirgamanathar S. Topical Anti-inflammatory activity of essential oils of Alpinia calcarata Rosc. its main constituents, and possible mechanism of action. Evid Based Complement Alternat Med. 2020;2020(1):2035671. doi:10.1155/2020/2035671
45. Cahyono B, Suzery M, Amalina ND. Anti-inflammatory effect of Alpinia galanga extract on acute inflammatory cell model of peripheral blood mononuclear cells stimulated with TNF-α. Med Gla. 2023;20(2). doi:10.17392/1561-23
46. George G, Shyni GL, Abraham B, Nisha P, Raghu KG. Downregulation of TLR4/MyD88/p38MAPK and JAK/STAT pathway in RAW 264.7 cells by Alpinia galanga. Reveals Its Beneficial Effects in Inflammation: Immunomodulatory Effect of Alpinia Galanga J Ethnopharmacol. 2021;275. doi:10.1016/j.jep.2021.114132
47. Shin SW, Hwang YS. Anti-periodontitis effect of ethanol extracts of Alpinia katsumadai seeds. Nutrients. 2021;14(1):136. doi:10.3390/nu14010136
48. Jung JM, Kwon OY, Choi JK, Lee SH. Alpinia officinarum rhizome ameliorates the UVB-induced photoaging through attenuating the phosphorylation of AKT and ERK. BMC Complement Med Ther. 2022;22(1):232. doi:10.1186/s12906-022-03707-w
49. Cheng CY, Chiang SY, Kao ST, Huang SC. Alpinia oxyphylla Miq extract reduces cerebral infarction by downregulating JNK-mediated TLR4/T3JAM- and ASK1-related inflammatory signaling in the acute phase of transient focal cerebral ischemia in rats. Chin Med. 2021;16(1):82. doi:10.1186/s13020-021-00495-2
50. Tie Y, Sun Z, Tong X, et al. Multi-omic analysis revealed the therapeutic mechanisms of Alpinia oxyphylla fructus water extract against bladder overactivity in spontaneously hypertensive rats. Phytomedicine. 2024;123:155154. doi:10.1016/j.phymed.2023.155154
51. Palanirajan A, Kannappan P, Kanniappan GK. Anticancer activity of Alpinia purpurata (Vieill) K. Schum. against MNU and testosterone-induced prostate cancer in male Wistar albino rats. Pharmacol Res Mod Chin Med. 2022;3. doi:10.1016/j.prmcm.2022.100105
52. Subash KR, Bhanu G, Vijaya K, Manjunath K, Umamaheswara K. Anti-inflammatory activity of ethanolic extract of Alpinia galanga in carrageenan induced pleurisy rats. Nat J Phys Pharm Pharmacol. 2016;6(5):468–470. doi:10.5455/njppp.2016.6.0719013072016
53. Patila S, Ujalambkara V, Rathoreb A, Rojatkarc S, Pokharkar V. Galangin loaded galactosylated pluronic F68 polymeric micelles for liver targeting. Biomed Pharmacother. 2019;112:108691. doi:10.1016/j.biopha.2019.108691
54. Nishidono Y, Tanaka K. Phytochemicals of Alpinia zerumbet: a review. Molecules. 2024;29(12):2845. doi:10.3390/molecules29122845
55. Shreshtha S, Sharma P, Kumar P, Sharma R, Singh SP. Nitric oxide: its role in immunity. J Clin Diagn Res. 2018;12(7):BE01–BE05. doi:10.7860/JCDR/2018/31817.11764
56. Coleman JW. Nitric oxide in immunity and inflammation. Int Immunopharmacol. 2001;1(8):1397–1406. doi:10.1016/s1567-5769(01)00086-8
57. Ghil S. Antiproliferative activity of Alpinia officinarum extract in the human breast cancer cell line MCF-7. Mol Med Rep. 2013;7(4):1288–1292. doi:10.3892/mmr.2013.1305
58. Zubair MS, Khairunisa SQ, Widodo A, Nasronudin, Pitopang R, Pitopang R. Antiviral screening on Alpinia eremochlamys, Etlingera flexuosa, and Etlingera acanthoides extracts against HIV-infected MT-4 cells. Heliyon. 2021;7(4):e06710. doi:10.1016/j.heliyon.2021.e06710
59. Khumpirapang N, Suknuntha K, Wongrattanakamon P, et al. The binding of Alpinia galanga oil and its nanoemulsion to mammal GABAA receptors using rat cortical membranes and an in silico modeling platform. Pharmaceutics. 2022;14(3):650. doi:10.3390/pharmaceutics14030650
60. Somarathna T, Thammitiyagodage MG, Ranaweera KKDS, et al. In vivo and in vitro toxicity profiles of hexane extract of Alpinia malaccensis rhizome in rat and cell line models. J Toxicol. 2021;2021:1–12. doi:10.1155/2021/9578474
61. Johnley IIR, Somasundaram G, Salwe KJ, Manimekalai K. Anti-ulcerogenic and anti-oxidant activity of Alpinia galanga rhizomes aqueous extract in indomethacin-induced gastric mucosal damage in Wistar albino rats. Biomed Pharmacol J. 2020;13(1). doi:10.13005/bpj/1860
62. Hui F, Qin X, Zhang Q, et al. Alpinia oxyphylla oil induces apoptosis of hepatocellular carcinoma cells via PI3K/Akt pathway in vitro and in vivo. Biomed Pharmacother. 2019;109:2365–2374. doi:10.1016/j.biopha.2018.11.124
63. Srivastava S, Mennemeier M, Pimple S. Effect of Alpinia galanga on mental alertness and sustained attention with or without caffeine: a randomized placebo-controlled study. J Am Coll Nutr. 2017;36(8):631–639. doi:10.1080/07315724.2017.1342576
64. Srivastava S. Selective enhancement of focused attention by Alpinia galanga in subjects with moderate caffeine consumption. Open Access J Clin Trials. 2018;10:43–49. doi:10.2147/OAJCT.S164450
65. Fedder MD, Jakobsen HB, Giversen I, Christensen LP, Parner ET, Fedder J. An extract of pomegranate fruit and galangal rhizome increases the numbers of motile sperm: a prospective, randomized, controlled, double-blinded trial. PLoS One. 2014;9(9):e108532. doi:10.1371/journal.pone.0108532
66. Eraiah MM, Kundapur M, Joshua L, Thomas JV. Acute effects of Alpinia galanga extract on mental alertness, accuracy and fatigue in human subjects: a randomized, double blind, placebo-controlled, cross-over study. Adv Complement Alt Med. 2023;7(4). doi:10.31031/ACAM.2023.07.000669
67. Akbarzadeh F, Eslamzadeh M, Behravan G, et al. Assessing the effect of Alpinia galanga extract on the treatment of SSRI-induced erectile dysfunction: a randomized triple-blind clinical trial. Front Psychiatry. 2023;14:1105828. doi:10.3389/fpsyt.2023.1105828
68. Maia MO, Dantas CG, Xavier Filho L, Cândido EA, Gomes MZ. The effect of Alpinia zerumbet essential oil on post-stroke muscle spasticity. Basic Clin Pharmacol Toxicol. 2016;118(1):58–62. doi:10.1111/bcpt.12439
69. Kolangi F, Shafi H, Memariani Z, et al. Effect of Alpinia officinarum Hance rhizome extract on spermatogram factors in men with idiopathic infertility: a prospective double-blinded randomised clinical trial. Andrologia. 2019;51(1):e13172. doi:10.1111/and.13172
70. Segal MS, Yu X. Herbal and Over-The-Counter Medicines and the Kidney. In: Floege J, Johnson RJ, Feehally J, editors. Comprehensive Clinical Nephrology.
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms.php
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms.php
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.