")
Back to Journals » Neuropsychiatric Disease and Treatment » Volume 21
Auditory N1, N2, and MMN to Pure-Tone and Consonant–Vowel Stimuli in Developmental Dyslexia and Benign Rolandic Epilepsy: A Progressive Insight Into the Underlying Disruptive Neurophysiology
Authors Tomé D , Pereira I, Vieira MC, Soares AP, Barbosa F
Received 31 October 2024
Accepted for publication 21 May 2025
Published 25 June 2025 Volume 2025:21 Pages 1271—1286
DOI https://doi.org/10.2147/NDT.S498193
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Roger Pinder
David Tomé,1,2 Ilídio Pereira,3 Maria Celeste Vieira,4 Ana Paula Soares,5 Fernando Barbosa6
1Department of Audiology, Center for Rehabilitation Research (CiR), School of Health (E2S), Polytechnic Institute of Porto, Porto, Portugal; 2Serviço de Neurologia do Hospital de São João, EPE, Porto, Portugal; 3Department of Biomathematics, Biostatistics and Bioinformatics, School of Health (E2S), Polytechnic Institute of Porto, Porto, Portugal; 4Clínica de Dislexia, Matosinhos, Portugal; 5Department of Basic Psychology, School of Psychology, University of Minho, Braga, Portugal; 6Laboratory of Neuropsychophysiology, Faculty of Psychology and Educational Sciences, University of Porto, Porto, Portugal
Correspondence: David Tomé, School of Health (E2S), Rua Dr. António Bernardino de Almeida, 400, Porto, 4200-072, Portugal, Tel +351 222 061 000, Email [email protected]
Purpose: Benign rolandic epilepsy or benign epilepsy in childhood with centrotemporal spikes (BECTS) and developmental dyslexia (DD) are two of the most studied disorders in childhood. Despite decades of research, the neurophysiological mechanisms underpinning these disorders are largely unknown. Here, we use auditory event-related potentials (AERPs) to shed light on these issues, since several authors have reported the existence of language and learning impairments. AERPs reflect the activation of different neuronal populations and are suggested to contribute to the evaluation of auditory discrimination (N1), attention allocation and phonological categorization (N2), and echoic memory (mismatch negativity (MMN]).
Patients and Methods: This study aims to investigate and document AERP changes in a group of children with BECTS and another group with DD. AERPs were recorded to pure-tones and consonant–vowel (CV) stimuli in an auditory oddball paradigm in eight children with BECTS, seven with DD, and 11 gender- and age-matched controls.
Results: The results revealed perceptual deficits for pure-tone and CV stimuli (pre-attentional and auditory discrimination) in DD, related to N1 reduced amplitude (p< 0.05; Fz: 2.57 μV, Cz: 2.75 μV). The BECTS group revealed no significant results for N1; however, there was an increase in N2b latency.
Conclusion: The findings in the DD group support the anchor-deficit hypothesis as an explanation for neurolinguistic deficits. The increased N2b latency in the BECTS group could be related to a potential lack of inhibitory mechanisms of pyramidal neurons, as justified by the process of epileptogenesis.
Keywords: N1, N2b, auditory processing, developmental dyslexia, BECTS
Introduction
Deficient letter–speech sound associations are considered to be the link between often observed phonological deficits and difficulties with learning to read.1,2 However, direct experimental evidence for this assumption is currently lacking. Thus, auditory processing deficits in rapid temporal processing, formation of stable representations in the brain, and mapping phonemes on to letters are being indicated as possible causal risk factors for specific language impairment (SLI), speech sound disorder (SSD), specific reading disability (SRD), and developmental dyslexia (DD).3
Neurodevelopmental disorders (NDDs) are a group of conditions that manifest during early childhood and affect the “normal” development and functioning of the brain. Common NDDs include autism spectrum disorder, communication/speech disorders, motor disorders, and attention deficit hyperactivity disorder. They affect more than 10% of people and typically manifest through persistent difficulties in acquiring, understanding, and/or using spoken or written language, impacting various aspects of a person’s life, including communication, learning, social interaction, behavior, cognition, and emotional functioning, with important long-term consequences.4 Benign rolandic epilepsy, also called benign epilepsy with centrotemporal spikes (BECTS), is the most common epilepsy syndrome in children. Most of the affected children grow out of this condition by puberty, hence the term “benign”. Although is not considered a learning disorder, no studies have compared children with DD and BECTS children with regard to auditory event-related potentials (AERPs).
Studies on DD have provided mixed findings on AERPs, with varying interpretation of different components. Reduced auditory N1 gating in individuals with dyslexia was linked to their poor word representation and neural adaptation.5–7 Abnormalities of mismatch negativity (MMN) correlated with the degree of phonological impairment,8–10 pointing to a selective processing deficit at an earlier phonetic level as a possible source of the difficulties in learning to read, while at a later lexical level information seems to be processed normally.11 However, MMN findings do to not seem to completely explain the single sound unit processing and discrimination in early stages.
Regarding benign epilepsy with centrotemporal spikes (BECTS), the studies are scarce but with consistent results, indicating neocortical excitability;12,13 however, none has clearly explained how AERPs could be clinically useful.
The ability to detect, discriminate, and analyze auditory stimuli is essential for normal learning and development. AERPs provide objective measures of central auditory processing. These scalp recorded potentials reflect activation of different neuronal populations that are suggested to contribute to auditory discrimination (N1), attention allocation and phonological categorization (N2), automatic preattentive discrimination and perception, and echoic memory (MMN).14–16
A number of electrophysiological studies have provided evidence for basic perceptual deficits in dyslexia, mainly because electrophysiology is a technique with high temporal resolution. Abnormal event-related potentials (ERPs) (amplitude, latency, topography) for auditory processing of non-speech and speech sounds were found in dyslexic children and adults.17 Several studies have been conducted to study auditory processing deficits with respect to discrimination abilities for pitch discrimination, stimulus duration, tone pattern manipulation, frequency modulation, gap detection, and temporal order judgments.
Moreover, AERPs at birth can successfully discriminate 76.5% of children with dyslexia, 100% of poor readers, and 79% of control children at 8 years of age,18,19 and can also correlate N1 amplitude and latency between 1 and 4 years of age, and word reading at 8 years of age.20 AERPs correlated with speech perception can strongly predict reading abilities at a very early age compared to other factors, such as home environment, academic stimulation, socioeconomic status, parental education, preschool language score, and short-term memory.19
In line with the latter studies, an anomalous ERP activity or functional difference (between 540 and 630 ms) in the right hemisphere in at-risk children (defined as having one first-grade relative reporting dyslexia) was found to be correlated with lower word and non-word reading accuracy in the first grade of school and poorer language skills at 2.5 years,21,22 poor verbal memory at 5 years and reduced phonological skills,23 and slower lexical access and less knowledge of letters at 6.5 years,24 compared to controls who presented the largest differences between standard and deviant stimuli in the left hemisphere.
Regarding the auditory N1 component, there have been reported deficits in gap detection by adults with dyslexia to tone and speech. Gap detection is a temporal processing task which measures the minimum interstimulus interval (ISI) required to perceive an interruption (gap) in a constant train of stimuli. Prolonged N1 and P3 latencies, and increased magnitude of N1, compared to controls, may be related to the general allocation of preattentional resources and/or may reflect compensatory mechanisms serving to increase arousal and readiness.25–28 Furthermore, decreased amplitude of N1–P2 (to tone and speech stimuli) seems to index auditory processing impairments, representing a causal risk factor for both language and reading disorders.29 In one study, aiming to correlate neuronal phonological deficits with auditory sensory processing, decreased N1c amplitude to rise time in the dyslexic group aged 8–10 years old was reported.29
Some studies reported no findings in the visual N1, stating that it depends on experimental paradigm.30
Speech perception involves the mapping of basic auditory information on to phonological units. Therefore, phonemic units alone are abstract but meaningful acoustic representations of speech. Not surprisingly, an auditory speech-processing deficit was determined in dyslexia in a number of AERP studies in children and adults, mainly an attenuated MMN.31–33 In addition, Bishop reviewed several prospective studies on MMN in children with a familial history of dyslexia.34
Benign epilepsy with rolandic or centrotemporal spikes (BECTS) is the most common form of idiopathic epilepsy in children, with an age of seizure onset between 3 and 14 years, representing 8–23% of epilepsies under the age of 16, and is more common in boys than girls.35 It is defined as a truly benign partial idiopathic epilepsy, mainly because spontaneous recovery usually occurs before adolescence in the absence of neurological and cognitive impairment, whether antiepileptic medication is started or not.36 However, several studies have questioned the benign nature of BECTS, reporting neuropsychological deficits in many domains.12,37 This seems to be related to the atypical form of BECTS, characterized by the presence of neurological deficits, continuous spike waves during sleep, atypical absences, speech delay, and intellectual deficits.38
A feasible protocol, diagnosis, and treatment monitoring method is still lacking. Furthermore, very few studies have investigated central auditory processing with AERPs in these disorders, and much so less in children and between disorders. In clinical practice, very similar electroencephalogram (EEG) curves are reported among school-aged children with rolandic epilepsy and children with other developmental disorders, such as dyslexia and autism.39–41 To our knowledge, no studies have established a comparison of passive AERPs between DD and BECTS discussed with a neurocognitive auditory sentence-processing model to interpret and explain the neurophysiological findings.
The overall aim of this research was to investigate (describe and explain) AERPs (N1, N2b, and MMN) in a passive auditory oddball paradigm, and their clinical usefulness (predict) in BECTS and DD, and to hypothesize neurophysiological endophenotypes or biomarkers of auditory processing dysfunction. Furthermore, the paper aims to discuss putative neurophysiological deficits in DD as a possible explanation for the phonological deficit hypothesis and the anchoring-deficit hypothesis, occurring in the very early stages of cortical auditory processing.
Materials and Methods
Participants
Participants were recruited from Serviço de Neurologia do Hospital de São João, EPE, and from Clínica de Dislexia Drª. Celeste Vieira, Matosinhos, Portugal. All participants were enrolled in mainstream academic programs, with successful levels of achievement.
Twenty-four children diagnosed with BECTS, 12 children with DD, and 17 controls were invited to participate, and a total of 26 children who complied with the inclusion criteria accepted to take part in the study.
All participants fulfilled the same inclusion criteria: normal hearing and vision, Portuguese (European) native speakers, no ear malformation, no motor disorders, and no neurological (except for epilepsy, developmental dysphasia, or dyslexia) or cognitive deficits. In addition, all participants from the control group had no ongoing medication that could affect the central or peripheral nervous system. Written signed informed consent was obtained from each child’s parents. Normal hearing was confirmed by audiometric testing according to BIAP clinical guidelines.42 The diagnosis of epilepsy (BECTS) followed the criteria of the Commission on Classification and Terminology of the International League Against Epilepsy, as applied by two neuropediatric clinicians.43 The diagnosis of DD followed the criteria of having specific problems learning to read and spell, despite adequate instruction, intelligence, and sociocultural opportunities.44,45 The diagnosis was made by a multidisciplinary team, composed of a teacher specializing in dyslexia, a psychologist, and a pediatrician. The diagnosis was supported by the evaluation of phonological awareness,46 intelligence quotient (Raven´s Progressive Matrices, Portuguese version),47 reading velocity and reading comprehension,48–50 writing (dysorthography), and Conners’ parents and teacher rating scales for assessing ADHD.51,52 At the time of the study, all participants were sufficiently compensated for the disorder to enable them to achieve normal academic skills in mainstream programs, however still with clinical support.
Exclusion criteria were a history of traumatic brain injury, or otological surgery in the past 12 months. Inclusion and exclusion criteria were verified with a brief anamnesis, parents’ report, and individual clinical process consultation during a team meeting. None of the invited participants met the exclusion criteria. The experimental procedures followed the tenets of the Declaration of Helsinki. The study was approved by the local Ethics Committee from Hospital São João, and parents gave informed consent concerning their children’s participation in the study.
Experimental Design and Data Analysis
Data for two experiments were collected in the same session. Experiment 1 consisted of the presentation of a block of pure-tone stimuli (standard: 1000 Hz; deviant: 1100 Hz; 100 ms duration) created in Audacity® software (v2.0.0), with 300 trials. Experiment 2 consisted of the presentation of a block of consonant–vowel (CV) stimuli digitalized at a rate of 44,100 Hz with 175 ms duration, spoken by a male Portuguese voice; standard: [ba] 118 Hz (F0); deviant: [pa] 127 Hz (F0), recorded with Nuendo® software (v4.3.0), with 300 trials. Before each individual data acquisition, the first experiment to start was randomly selected.
In both experiments, standard stimuli comprised 80% of all trials (240 trials), while deviant stimuli comprised 20% (60 trials), with a stimulus onset asynchrony (SOA) of 800 ms to a proximal sound level of 75 dB SPL confirmed with a sound level meter.
Stimuli were binaurally presented using Presentation® software (Neurobehavioral Systems) via closed headphones, while the participants were comfortably seated in an armchair and instructed to keep alert with their eyes open, actively listen to the auditory stimuli, and focus on a small cross in the center of a computer screen during the recording. Each individual data collection session had an average duration of 40–50 minutes.
The EEG set-up was carried out according to the 10–20 electrode placement international system.53 Recordings were obtained from the following cup-shaped silver chloride (Ag-AgCl) reversible electrodes: Cz, Fz, Fpz, M1, M2, and Oz (ground), referenced to averaged ear lobes (A1 and A2) to reduce the likelihood of artificially inflating activity in one hemisphere.54 We decided on a referential montage as this allows a truer picture of the relative voltage amplitudes at each electrode,55,56 which seemed more appropriate to the research goals. The electrodes used were reversible (non-polarized); these are preferred for common neurophysiological applications because polarization is avoided as the chloride ion is common to both the electrode and the electrolyte.56
An electrode was attached above the right eye to monitor the electrooculogram (EOG). The selected position for the ground electrode was the most appropriate, to obtain a good differential voltage between the active and reference points. Linking reference electrodes from two earlobes reduces the likelihood of artificially inflating activity in one hemisphere; in other words, a non-lateralized reference.54 Prior to this, skin preparation was carried out by cleaning the skin surface with ethyl alcohol and using abrasive paste for slight skin abrasion, to lower the impedance to below 10 kΩ at all sites.
The quality of recordings was monitored during data acquisition, and continuous EEG data were stored on the computer hard disk for off-line analysis and grand averaging. Sweeps with amplitudes exceeding ±70 μV in any channel except for EOG were excluded. Trials during which blinks or saccades, or components that could be attributed to such artfacts based on their topography and time course, were eliminated from the data before averaging. Impedance between the electrodes and skin was kept below 10 kΩ at all sites. The EEG data were recorded with a 32-channel bioamplifier and ASA® software v4.0.6.8 (ANT Neuro, The Netherlands). The sampling rate was 512 Hz for each channel and the frequency recording bandwidth was between 0.05 and 100 Hz.
Preprocessing included band-pass filtering (0.3–30 Hz), entire sweep linear detrending, and a baseline correction of individual epochs (200 ms). The baseline window was also used to inspect the overall noise level of recordings and increase the accuracy for analyzing the different waveforms.57 The first 10 sweeps of each epoch file were excluded from the averaging process to reduce ERP variation associated with the start of the stimulation sequence.58,59
The MMN was determined from the Fpz, Fz, Cz, M1, and M2 electrodes, by computing difference waves, ie, by subtracting the average standard-stimulus response from the average deviant-stimulus response for each electrode and subject.60 N1 and N2b were identified as the two largest negative peaks, occurring between 100 and 300 ms. We analyzed peak latency and peak-to-peak amplitude to increase precision.
Statistical analyses were performed using IBM SPSS v.25.61 Mean amplitude and latency values of each component (N1, N2b, MMN) were computed in a two-way repeated-measures analysis of variance (ANOVA) with electrode (Fpz, Fz, Cz) and stimuli (pure-tone and CV) as within-subject factors, and group (control, BECTS, and DD) as a between-subjects factor. Separate tests for group homogeneity were also applied for gender (Fisher’s exact test), handedness (collected prior to ERPs),62,63 and age (Kruskal–Wallis one-way ANOVA).
An alpha level of 0.05 was used. Because we have three levels of a repeated-measures factor, sphericity was evaluated by Mauchly’s test.64 Sphericity was obtained for all measures, and thus we can assume that the variances of the differences between all combinations of the groups are equal. In addition, owing to small sample sizes, Bonferroni correction was performed to control and reduce type I errors when performing multiple hypothesis tests.65
Results
The final sample comprised 26 participants: eight with BECTS (age: mean=9.88, SD=2.10 years), seven with DD (age: mean=10.6, SD=3.31 years), and 11 controls (age: mean=12.1, SD=3.14 years). All were Portuguese native speakers, with normal hearing and vision, no ear malformation, and no neurological or mental deficits. Children from BECTS were seizure free for the past 3 months and had no history of language regression with the onset of seizures. The detailed characterization of participant groups, medication, and other clinical characteristics is shown in Table 1.
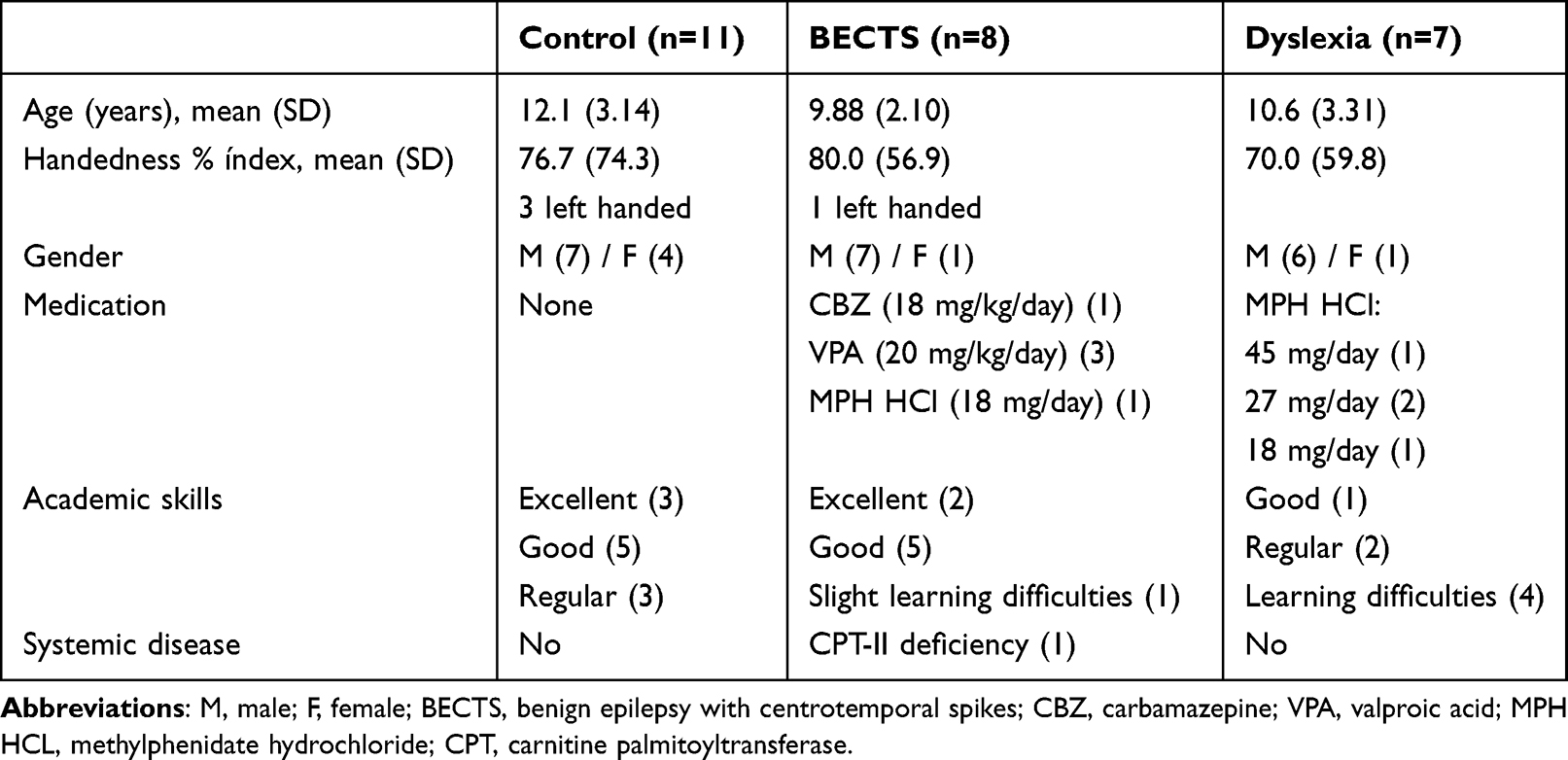 |
Table 1 Clinical Characteristics, Medication, and Outcome of Each Participant Group |
Statistical analysis revealed homogeneity between the groups for gender (Fisher’s exact test bilaterally =0.505, 50 >20%, 3 <5 cells), age (H=0.287, p>0.05), and handedness (H=0.768, p>0.05).
The mean peak-to-peak amplitude and peak latencies of the N1, N2, and MMN wave from Fpz, Fz, and Cz, for the BECTS, dyslexia, and comparison/control groups to pure-tone and speech (CV) paradigms are presented in Table 2.
 |
Table 2 Mean (SD) Peak-to-Peak Amplitude and Peak Latencies of the N1, N2, and MMN Wave in the Fpz, Fz, and Cz Electrodes, for the BECTS, Dyslexia, and Comparison/Control Groups to Pure-Tone and Speech (Consonant–Vowel) Paradigms |
In addition, the grand average waveforms of each of the above-mentioned components for each group (control, BECTS, and DD) and stimuli (pure-tone and CV) are presented in Figure 1. After Mauchly’s test procedure, sphericity was assumed for all measures.
Figure 1 Continued. Figure 1 Grand average deviant waveforms for (a) pure-tones and (b) consonant–vowel (CV) stimuli for the BECTS group (red solid line), the control group (blue solid line), and the dyslexia group (black solid line). Grand average MMN difference waveforms are represented in the right-hand column.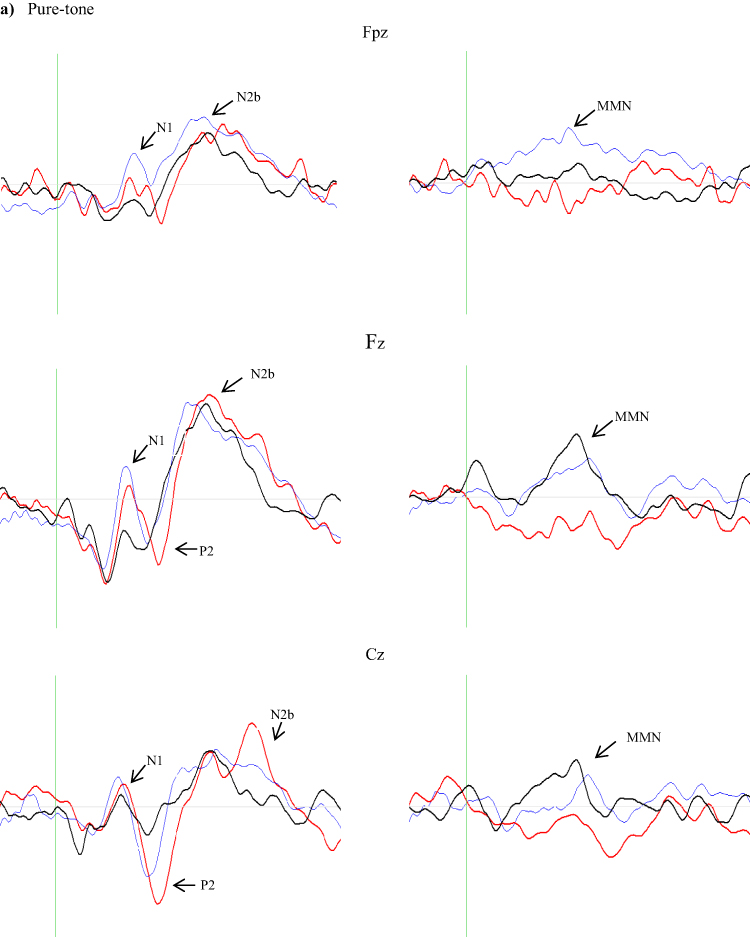

Amplitude
For N1 amplitude, the ANOVA revealed a significant main effect of stimuli [F(1,23)=7.48, p=0.012] and a marginally significant effect of group [F(2,23)=3.34, p=0.053]. A significant main effect of electrode was also found for N1 [F(2,46)=20.4, p=0.001]. Post-hoc comparisons using Bonferroni correction revealed that the N1 amplitude of the DD group was significantly lower than in controls, to both stimuli (Table 2).
Regarding the N2 amplitude, the only effect that reached statistical significance was the main effect of electrode [F(2,46)=39.7, p=0.000]. The same was observed for MMN, where only the main effect for electrode was significant [F(2,46)=26.3, p=0.000].
Latency
For the N1 latency, the ANOVA revealed a significant main effect of stimuli [F(1,23)=4.47, p=0.046, indicating that the processing of speech (CV) stimuli requires more time than pure-tone in all the groups, regardless of the electrode. This may occur in a later stage as a result of overlapping processes since more neural resources are required for speech processing. The results revealed similar phenomena to N1.
For N2 latency, the results revealed an almost significant difference between groups [F(2,23)=3.26, p=0.057] and, importantly, that the two-fold group×electrode interaction reached significance [F (4,46)=5.70, p=0.001. Post-hoc analyses revealed that the interaction was due to the BECTS group revealing a decrease in N2b latency from posterior (Cz) to anterior sites (Fpz), while in the DD group the opposite occurred, with an increase of N2b latency from posterior to anterior (Figure 2).
 |
Figure 2 N2b mean latency for electrodes and participant groups (control, BECTS/epilepsy, and dyslexia). |
This result is complemented by voltage maps (Figures 3 and 4). It can be seen that in the dyslexia group, after an unclear or even absence of frontal N1 negativity, the N2b initiates earlier than in the other groups, and absolute peak-to-peak amplitude values are bigger compared to controls, possibly to compensate for the poorly exogenous (N1) early sensory auditory processing. Lastly, for MMN latency no significant differences were found between groups [F(2,23)=1.44, p=0.257]. However, there is a tendency for an increase in latency across electrodes and between groups for both stimuli.
 |
Figure 3 Topographic voltage map of grand average deviant waveforms to pure-tones in (a) controls, (b) BECTS, and (c) dyslexia, in a 141 ms time window (113–254 ms). |
 |
Figure 4 Topographic voltage map of grand average deviant waveforms to consonant–vowel stimuli in (a) controls, (b) BECTS, and (c) dyslexia, in a 141 ms time window (117–258 ms). |
Finally, topographic MMN voltage maps are presented in Figures 5 and 6. The control group only required four periods of time to onset and fade away, in other words to complete the total curve, traveling between 180 and 227 ms for pure-tone stimuli (Figure 5) and between 148 and 195 ms for CV stimuli (Figure 6), with a clear prefrontal trigger. However, for BECTS, the voltage map is diffuse, and positive for pure-tone stimuli (Figure 5) and highly negative in all presented periods (102–242 ms) for CV stimuli. Despite MMN peak-to-peak amplitude means being decreased compared to the control group, the N2 amplitudes increased and may have contributed to MMN voltage distribution.
 |
Figure 5 Topographic voltage map of MMN to pure-tones in (a) controls, (b) BECTS, and (c) dyslexia, in a 140 ms time window (102–242 ms). |
 |
Figure 6 Topographic voltage map of MMN to consonant–vowel stimuli in (a) controls, (b) BECTS, and (c) dyslexia, in a 140 ms time window (102–242 ms). |
The same happens with the dyslexia group for both stimuli (Figures 5 and 6). This N2 amplitude enhancement may be related to a potential failure of inhibitory mechanisms, mainly at Fz (Table 2). The brain cannot orient a response by inhibiting irrelevant information; thus, attention allocation processes are highly activated to compensate for this. This was expected to happen in the DD group for both stimuli. However, in BECTS group the N2 amplitude is even higher, specially for speech stimuli, but with a latency delay for both stimuli (Table 2). Another explanation is that the enlargement of N2 to speech/CV stimuli seen in the BECTS group shared the same cortical generator as the spontaneous rolandic spikes,13 because none of the children with rolandic epilepsy presented learning or language impairments.
In addition, the absence of a clear frontal negativity of the N1 component for both stimuli in the dyslexia group suggests a deficit in auditory novelty detection and the orienting response. It requires lengthy consideration that in DD neurophysiological processing deficits occur in the very first primary acoustic analysis, thus, in the primary auditory cortices. This results, in later stages, in a neural network deficit processing problem, or in an erroneous “metamorphosic” wave succession.
Discussion
The overall aim of this research was to investigate (describe and explain) auditory event-related potentials (N1, N2b, and MMN) in a passive auditory oddball paradigm, and their clinical usefulness (predict) in neurodevelopmental disorders such as DD and BECTS.
The DD group revealed significantly reduced N1 amplitude to pure-tone and speech CV stimuli, compared to the control group. Furthermore, an interaction effect of stimuli×group was also observed, with N1 amplitude to speech CV stimuli being lower than to pure-tone stimuli in the DD group, for all electrodes. This is consistent with previous studies17,20 suggesting that N1 is the most prominent mesogenous component in response to acoustic input and therefore can reliably correlate and detect neuronal phonological deficits with auditory sensory processing.3,29,66,67 Note that our participants were receiving specialized educational support, despite not being fully considered as compensated for their language deficits. This decrease in N1 amplitude for speech CV stimuli may reflect a residual auditory processing deficit that has not yet been addressed by top–down compensatory strategies. Another explanation might be the weakened capture of auditory attentional discrimination in dyslexia, allowing for a possible impairment in the dynamics that link attention with short-term memory.68,69 Thus, low N1 may represent a deficit in auditory novelty detection and orienting responses.
These results support Ahissar’s opinion that the impaired-anchoring hypothesis could be viewed as a specific type of impaired-attention hypothesis. It is proposed that although top–down attentional mechanisms of people with dyslexia are behaviorally unimpaired, their bottom–up-driven attentional mechanisms are less effective because they cannot use the gradual build-up of predictions around repeated stimuli, which typically reduce attentional load.5,70 Auditory oddball paradigms are particularly suitable to test this. Furthermore, this is consistent with Bonte and Blomert’s findings in ERPs,71 that the early priming-related responses of dyslexics (N1–N2, delays of 100 and 200 ms) were abnormal, whereas their late priming-related response components (N400) were adequate. They interpreted this finding as indicating a deficit at the earlier, perceptual stage, rather than at the later, phonological lexical stage.
The BECTS group presented no differences regarding N1 measures, compared to the control group. Besides, children with BECTS did not reveal any reading or learning impairments, which suggests that reduced auditory N1 amplitude could be a putative endophenotype for DD.
One must bear in mind that more than half of the dyslexic participants had attention deficit and were under methylphenidate medication. Although it is possible that the N1 results stated above may be due to possible ADHD comorbidity, we suggest that this is not the case. Children with ADHD exhibit increased early automatic attentional orienting (N1), with shorter latencies,72 first failing to inhibit irrelevant stimuli before failing to allocate sufficient attentional resources in further processing stages, ie, decreased N2 and P3.73–75
Children in the BECTS group seemed to have increased N2b amplitude to both stimuli, but our results revealed no statistically significant differences. The literature states that increased amplitudes of N2b may be related to a potential failure of inhibitory mechanisms, as reflected by decreased N1; thus, attention allocation processes are highly activated to compensate (increased N2b).12,76 This is consistent with recent findings showing that the loss of inhibitory properties of pyramidal neurons is related to epileptogenesis.77,78 Based on the Hebbian concept, BECTS may have a failure in the neural self-regulatory inhibitory feedback mechanism necessary to maintain the total activity of the network within certain limits.79 In addition, an unstable positive-feedback loop develops, whereby stronger and stronger responses would follow each new presentation of a given stimulus, leading to the “overgrowth” of one of the cell assemblies, which would rapidly extend and cover most of the network, causing widespread unphysiological states of saturated activation. To confirm this hypothesis, we must wait until further computational models consider in the algorithm the complex dynamics of ionic channels and neurotransmitters that underpin epileptogenesis.
Another hypothesis is that the enlargement of N2b to speech stimuli, as seen in the BECTS group, shares the same cortical generator as the spontaneous rolandic spikes, thus indexing epileptic cortical excitability. This inscreased amplitude is more evident on centro-parietal site (Cz), as revealed by the significant interaction effect of group×electrode, voltage maps, and a statistically significantly prolonged N2b to both stimuli types. As an endogenous component, increased N2b could be due to an antiepileptic drug effect, but the literature confirms that only P300 seems to be affected by carbamazepine.80
On the other hand, increased N2b amplitude in the dyslexia group could be interpreted the other way around. The most probable explanation is that methylphenidate normalizes ERP indices,81,82 with the plausible contribution of specialized educational support. Methylphenidate has no effect on N1 and P2.83 This is complemented by the voltage map: in the dyslexia group, after an unclear or even absence of frontal N1 negativity, the N2b initiates earlier than the other groups, and absolute peak-to-peak amplitude values are bigger than in controls, possibly to compensate for the poorly mesogenous (N1) early sensory auditory processing. Furthermore, this seems worse for speech stimuli. However, this requires further research to confirm whether it is also an effect of methylphenidate or the initial stages of a top–down compensatory mechanism from the right hemisphere.24,84
Regarding AERP latency in dyslexia, the results showed no significant differences between groups, as stated by other authors for N1 and P3,85 and N2,66 possibly owing to sample size. However, the majority of the present studies do not report any significant differences between dyslexia and control groups for N1, N2b, and MMN latencies.
Lastly, we used auditory spectral paradigms with a maximum of 100 Hz difference between standard and deviant stimuli, both speech and pure-tone. Investigations of frequency (pitch) discrimination with MMN suggest a deficit in dyslexia (children and adults) only between stimuli that differ by less than 100 Hz.8,86,87 Studies using large pitch differences (eg, 200 Hz and greater) between tones did not report any abnormalities in adults with dyslexia,88,89 with the exception of a study by Sebastian and Yasin,17 suggesting a deficit for shorter spectral changes only. This deficit may reflect a widened representational width (RW) in sound perception. RW is a concept proposed by Näätänen and Alho to describe an individual’s discrimination accuracy,90 which is dependent on their particular ability to perceive differences in sound. The narrower the width, the better the discrimination ability.89
Statistical analysis did not reveal any significant differences in MMN between groups, for either stimulus effect. This is not a new finding, as several studies report no differences for MMN in dyslexia.91–93 Our results are similar to those of Sebastian and Yasin’s study.17
According to the neurocognitive model of language processing,94 neurophysiological deficits in DD occur during Phase 0 and Phase 1 (N1, MMN, and possibly P2), corresponding to an impaired identification of phonemes, word forms, phonological segmentation, and sequencing. Furthermore, if we apply the notions of the cell assembly model and Garagnani’s computational model of language processing,79 the size of cell assemblies decreases as a linear function of the minimal activation threshold. Therefore, the decreased N1 amplitude in dyslexia may correlate with a non-achieved “ignition threshold” or poorly defined cell assemblies in temporal lobe N1 generators (possibly BA 42 and 44).
One might wonder, if the N2b has normal amplitude, why the deficits? The “normal” N2b amplitude in the dyslexia group (compared to the control group) is probably a medication effect. Methylphenidate normalizes N2 amplitudes. But that does not mean that the cell assemblies are correctly processing the initial erroneous or insufficient processing of auditory information, as indexed by N1. Thus, a complementary hypothesis is to conceive dyslexia as a neurofunctional integration and connectivity neural network problem. Functional integration refers to the interaction of functional specialized systems. The connectivity pattern, in turn, is a function of epigenetic activity and plasticity dependent on experience. Dyslexic brains may have a white matter deficit in the dorsal pathway connecting the posterior part of Brodmann’s area (BA 44) and the posterior STG/STS (BA 42). The latter is crucial for the human language capacity that is characterized by the ability to process complex sentence structures.94 This is corroborated by Brauer,95 according to whom, in children, at an age at which they are still deficient in processing syntactically complex sentences, the dorsal pathway connecting the language areas is not yet fully myelinated. Thus, it seems that the evolution of language is tightly related to the maturation of the dorsal pathway connecting those areas which, in the adult human brain, are involved in the processing of syntactically complex sentences.96
In summary, attenuated N1 to speech CV and pure-tone stimuli may provide a good index of underlying auditory neuroprocessing deficits in dyslexia, though longitudinal AERPs and genetic studies are still required to consider this a feasible biomarker. Diminished MMN to an auditory oddball paradigm may not be the most consistent physiological marker, but the decrease in amplitude from pure-tone to speech may be a stable measure to consider in future research and clinical application.
Limitations and Future Research
Few studies found no speech discrimination deficits to temporal content, but to spectral discrimination instead (right hemisphere).9 Moreover, we still do not know whether the anchoring-deficit hypothesis is specific to dyslexia or whether it can also characterize individuals with a range of other learning disabilities (eg, attention disorder); in other words, whether the anchoring deficit is inherited. The small number of participants in the DD group did not allow us to analyze such a hypothesis.
Keeping the above in mind, further research on auditory P200, N400,97 P600, and visual N17098 is required and may clarify how the development and maturation of phonological processing interfere with learning to read. Furthermore, correlating the AERP results with behavioral measures may demand a clear separation between phonological speech representations and the metacognitive task demands, such as short-term memory or conscious awareness, that may be involved in accessing linguistic representations.99 Future work is needed to analyze the association between phonological awareness and speech rhythm more closely. At present, sensitivity to speech rhythm is increasingly being linked to reading progress.100,101
Furthermore, considering the presented neurocognitive model of auditory sentence processing and the mapping hypothesis,102 research on the integration of auditory and visual modalities may clarify the etiology of dyslexia103 and how orthographic representations map on to phonological ones, and explain letter-processing deficits in phonological dyslexia, as a phonological recoding deficit.104,105 This may also lead to a profound discussion on neuroplasticity, evolving the concept of central auditory processing and its relations and integrations of different sensory modalities, hemispheric dominance, and handedness.
Conclusion
Children with DD revealed decreased N1 amplitude for speech CV stimuli. This finding may reflect a residual auditory processing deficit in auditory novelty detection and the orienting response that has not yet been addressed by top–down compensatory strategies. That is, the bottom–up processes are impaired, thus supporting the anchoring-deficit hypothesis. Although many issues remain to be addressed, this work represents the first step towards a better understanding of how AERP components can be reliably used in clinical practice. Such knowledge would improve diagnosis, prognosis, follow-up, and treatment strategies.
Data Sharing Statement
The datasets used and/or analyzed during the current study are available from the corresponding author upon reasonable request.
Ethics Approval and Consent to Participate
This study was conducted with approval from the Ethics Committee of the Hospital of São João and was conducted in accordance with the Declaration of Helsinki. Written informed consent was obtained from all participants (parents or legal representatives).
Acknowledgments
We appreciate the assistance of the Neurology Department and Neuropediatric Service of the Hospital of São João, namely Dr. Sampaio, in authorizing the research.
Funding
This work was supported by grant 2022.05618.PTDC from the Portuguese Foundation for Science and Technology (FCT).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Snowling MJ. The development of grapheme-phoneme correspondence in normal and dyslexic readers. J Exp Child Psychol. 1980;29:294–305. doi:10.1016/0022-0965(80)90021-1
2. Vellutino FR, Fletcher JM, Snowling MJ, Scanlon DM. Specific reading disability (dyslexia): what have we learned in the past four decades? J Child Psychol Psychiatr. 2004;45(1):2–40. doi:10.1046/j.0021-9630.2003.00305.x
3. McArthur G, Atkinson C, Ellis D. Atypical brain responses to sounds in children with specific language and reading impairments. Dev Sci. 2009;12(5):768–783. doi:10.1111/j.1467-7687.2008.00804.x
4. Thapar A, Cooper M, Rutter M. Neurodevelopmental disorders. Lancet Psychiat. 2017;4(4):339–346. doi:10.1016/S2215-0366(16)30376-5
5. Ahissar M. Dyslexia and the anchoring-deficit hypothesis. TiCS. 2007;11(11):458–465. doi:10.1016/j.tics.2007.08.015
6. Panahi R, Akbari M, Jarollahi F, et al. Atypical function of auditory sensory gating in children with developmental dyslexia: investigating its relationship with cognitive abilities. Dyslexia. 2023;29(4):426–440. doi:10.1002/dys.1754
7. Peter B, McCollum H, Daliri A, Panagiotides H. Auditory gating in adults with dyslexia: an ERP account of diminished rapid neural adaptation. Clin Neurophysiol. 2019;130(11):2182–2192. doi:10.1016/j.clinph.2019.07.028
8. Baldeweg T, Richardson A, Watkins S, Foale C, Gruzelier J. Impaired auditory frequency discrimination in dyslexia detected with mismatch evoked potentials. Ann Neurol. 1999;45(4):495–503. doi:10.1002/1531-8249(199904)45:4<495::AID-ANA11>3.0.CO;2-M
9. Kraus N, McGee T, Carrell TD, et al. Auditory neurophysiologic responses and discrimination deficits in children with learning problems. Science. 1996;273(5277):971–973. doi:10.1126/science.273.5277.971
10. Tuomainen OT. Auditory short-term memory trace formation for nonspeech and speech in SLI and dyslexia as indexed by the N100 and mismatch negativity electrophysiological responses. Neuroreport. 2015;26(6):374–379. doi:10.1097/WNR.0000000000000357
11. Bonte ML, Blomert L. Developmental dyslexia: ERP correlates of anomalous phonological processing during spoken word recognition. Brain Res Cogn Brain Res. 2004;21(3):360–376. doi:10.1016/j.cogbrainres.2004.06.010
12. Liasis A, Bamiou DE, Boyd S, Towell A. Evidence for a neurophysiologic auditory deficit in children with benign epilepsy with centro-temporal spikes. J Neural Transm. 2006;113(7):939–949. doi:10.1007/s00702-005-0357-6
13. Tomé D, Sampaio M, Mendes-Ribeiro J, et al. Auditory event-related potentials in children with benign epilepsy with centro-temporal spikes. Epilepsy Res. 2014;108:1945–1949. doi:10.1016/j.eplepsyres.2014.09.021
14. Näätänen R. The mismatch negativity: a powerful tool for cognitive neuroscience. Ear Hear. 1995;16:6–18. doi:10.1097/00003446-199502000-00002
15. Liasis A, Towell A, Boyd S. Intracranial auditory detection and discrimination potentials as substrates of echoic memory in children. Cogn Brain Res. 1999;7:503–506. doi:10.1016/S0926-6410(98)00049-4
16. Henkin Y, Kishon-Rabin L, Gadoth N, Pratt H. Auditory event-related potentials during phonetic and semantic processing in children. Audiol Neurootol. 2002;7:228–239. doi:10.1159/000063739
17. Sebastian C, Yasin I. Speech versus tone processing in compensated dyslexia: discrimination and lateralization with a dichotic mismatch negativity (MMN) paradigm. Int J Psychophysiol. 2008;70:115–126. doi:10.1016/j.ijpsycho.2008.08.004
18. Molfese DL. Predicting dyslexia at 8 years of age using neonatal brain responses. Brain Lang. 2000;72(3):238–245. doi:10.1006/brln.2000.2287
19. Molfese VJ, Molfese DL, Modgline AA. Newborn and preschool predictors of second-grade reading scores: an evaluation of categorical and continuous scores. J Learn Disabil. 2001;34(6):545–554. doi:10.1177/002221940103400607
20. Espy KA, Molfese DL, Molfese VJ, Modglin A. Development of auditory event-related potentials in young children and relations to word-level reading abilities at age 8 years. Ann Dyslexia. 2004;54(1):9–38. doi:10.1007/s11881-004-0002-3
21. Lyytinen P, Eklund K, Lyytinen H. Language development and literacy skills in late talking toddlers with and without familial risk for dyslexia. Ann Dyslexia. 2005;55(2):166–192. doi:10.1007/s11881-005-0010-y
22. Lyytinen H, Guttorm TK, Huttunen T, et al. Psychophysiology of developmental dyslexia: a review of findings including studies of children at risk for dyslexia. J Neurolinguistics. 2005;18(2):167–195. doi:10.1016/j.jneuroling.2004.11.001
23. Guttorm TK, Leppänen PH, Poikkeus AM, et al. Brain event-related potentials (ERPs) measured at birth predict later language development in children with and without familial risk for dyslexia. Cortex. 2005;41(3):291–303. doi:10.1016/S0010-9452(08)70267-3
24. Guttorm TK, Leppänen PH, Hämäläinen JA, et al. Newborn event-related potentials predict poorer pre-reading skills in children at risk for dyslexia. J Learn Disabil. 2009;43(5):391–401. doi:10.1177/0022219409345005
25. Georgiewa P, Rzanny R, Gaser C, et al. Phonological processing in dyslexic children: a study combining functional imaging and event-related potentials. Neurosci Lett. 2002;318(1):5–8. doi:10.1016/S0304-3940(01)02236-4
26. Farmer ME, Klein RM. The evidence for a temporal processing deficit linked to dyslexia. Psychon Bull Rev. 1995;2(4):460–493. doi:10.3758/BF03210983
27. Hämäläinen JA, Fosker T, Szücs D, Goswami U. N1, P2 and T-complex of the auditory brain event-related potentials to tones with varying rise times in adults with and without dyslexia. Int J Psychophysiol. 2011;81(1):51–59. doi:10.1016/j.ijpsycho.2011.04.005
28. Helenius P, Salmelin R, Richardson U, et al. Abnormal auditory cortical activation in dyslexia 100 msec after speech onset. J Cogn Neurosci. 2002;14(4):603–617. doi:10.1162/08989290260045846
29. Stefanics G, Fosker T, Huss M, et al. Auditory sensory deficits in developmental dyslexia: a longitudinal ERP study. Neuroimage. 2011;57(3):723–732. doi:10.1016/j.neuroimage.2011.04.005
30. van Setten EH, Maurits NM, Maassen BM. N1 lateralization and dyslexia: an event-related potential study in children with a familial risk of dyslexia. Dyslexia. 2019;25(1):84–102. doi:10.1002/dys.1604
31. Csépe V. Auditory event-related potentials in studying developmental dyslexia. In: Csépe V, editor. Dyslexia: Different Brain, Different Behaviour. New York, USA: Kluwer Academic/Plenum Publishers; 2003:81–112.
32. Schulte-Körne G, Deimel W, Bartling J, Remschmidt H. Auditory processing and dyslexia: evidence for a specific speech processing deficit. Neuroreport. 1998;9(2):337–340. doi:10.1097/00001756-199801260-00029
33. Schulte-Kӧrne G, Deimel W, Bartling J, Remschmidt H. Speech perception deficit in dyslexic adults as measured by mismatch negativity (MMN). Int J Psychophysiol. 2001;40:77–87. doi:10.1016/S0167-8760(00)00152-5
34. Bishop DV. Using mismatch negativity to study central auditory processing in developmental language and literacy impairments: where are we, and where should we be going? Psychol Bull. 2007;133(4):651–672. doi:10.1037/0033-2909.133.4.651
35. Bernardina BD, Sgró V, Fejerman N. Epileptic Syndromes in Infancy, Childhood and Adolescence.
36. Nicolai J, van der Linden I, Arends JB, et al. EEG characteristics related to educational impairments in children with benign childhood epilepsy with centrotemporal spikes. Epilepsia. 2007;48(11):2093–2100. doi:10.1111/j.1528-1167.2007.01203.x
37. Myatchin I, Mennes M, Wouters H, et al. Working memory in children with epilepsy: an event-related potentials study. Epilepsy Res. 2009;86(2):183–190. doi:10.1016/j.eplepsyres.2009.06.004
38. Wong PH, Gregory D, Farrell K. Comparison of spike topography in typical and atypical benign Rolandic epilepsy of childhood. Electroencephalogr Clin Neurophysiol. 1985;61:S47. doi:10.1016/0013-4694(85)90207-X
39. Canavese C, Rigardetto R, Viano V, et al. Are dyslexia and dyscalculia associated with Rolandic epilepsy? A short report on ten Italian patients. Epileptic Disord. 2007;9(4):432–436. doi:10.1684/epd.2007.0138
40. Oliveira EP, Neri ML, Capelatto LL, et al. Rolandic epilepsy and dyslexia. Arq Neuropsiquiatr. 2014;72(11):826–831. doi:10.1590/0004-282X20140141
41. Spence SJ, Schneider MT. The role of epilepsy and epileptiform EEGs in autism spectrum disorders. Pediatr Res. 2009;65:599–606. doi:10.1203/PDR.0b013e31819e7168
42. International Bureau for Audiophonology. Audiometric Classification of Hearing Impairment – Recommendation 02/1. BIAP; 2003.
43. Commission on Classification and Terminology of the International League Against Epilepsy. Proposal for revised classification of epilepsies and epileptic syndromes. Epilepsia. 1989;30:389–399. doi:10.1111/j.1528-1157.1989.tb05316.x
44. Critchley M. Developmental dyslexia: a constitutional disorder of symbolic perception. Res Publ Assoc Res Nerv Ment Dis. 1970;48:266–271.
45. McDonough EM, Flanagan DP, Sy M, Alfonso VC. Specific learning disorder. In: Goldstein S, DeVries M, editors. Handbook of DSM-5 Disorders in Children and Adolescents. Springer International Publishing/Springer Nature; 2017:77–104. doi:10.1007/978-3-319-57196-6_4
46. Sim-Sim I. Avaliação da linguagem oral: um contributo para o conhecimento do desenvolvimento linguístico das crianças portuguesas.
47. Simões MR. Investigações no âmbito da aferição nacional do Teste das Matrizes Progressivas Coloridas de Raven (M.P.C.R.). Lisboa, Portugal: Fundação Calouste Gulbenkian; 2000.
48. Sucena A, Castro SL. Aprender a ler e a avaliar a leitura. O TIL Teste de Idade de Leitura.
49. Sucena A, Castro SL. ALEPE: Bateria de Avaliação da Leitura em Português Europeu. Lisboa, Portugal: Edições Cegoc-Tea; 2012.
50. Viana FL, Ribeiro I. PRP – Prova de Reconhecimento de Palavras. Lisboa, Portugal: Edições Cegoc-Tea; 2010.
51. Rodrigues AN. Contributos para a utilização das Escalas de Conners Revistas (1997) no processo de avaliação da PHDA - (Parte I). REER. 2005;12(2):71–95.
52. Rodrigues AN. Contributos para a utilização das Escalas de Conners Revistas (1997) no processo de avalaição da PHDA - Parte II - Diferenças entre professores e pais no processo de referenciação e avaliação. REER. 2006;13:7–20.
53. Jasper HH. The ten-twenty electrode system of the international federation. Electroencephalogr Clin Neurophysiol. 1958;10(2):371–375.
54. Miller GA, Lutzenberger W, Elbert T. The linked-reference issue in EEG and ERP recording. J Psychophysiol. 1991;5(3):273–276.
55. American Electroencephalographic Society. Guidelines in EEG. J Clin Neurophysiol. 1986;3(suppl 1):1–147.
56. Sinclair CM, Gasper MC, Blum AS. Basic electronics in clinical neurophysiology. In: Blum AS, Rutkove SB, editors. The Clinical Neurophysiology. Totowa, New Jersey (USA): Humana Press; 2007:3–18.
57. Luck SJ. An Introduction to the Event-Related Potential Technique. Cambridge (MA): MIT press; 2005:225–294.
58. Pekkonen E, Rinne T, Näätänen R. Variability and replicability of the mismatch negativity. Electroencephalogr Clin Neurophysiol. 1995;96(6):546–554. doi:10.1016/0013-4694(95)00148-R
59. Pettigrew CM, Murdoch BE, Kei J, et al. The mismatch negativity (MMN) response to complex tones and spoken words in individuals with aphasia. Aphasiology. 2005;19(2):131–163. doi:10.1080/02687030444000642
60. Duncan CC, Barry R, Connolly JF, et al. Event-related potentials in clinical research: guidelines for eliciting, recording, and quantifying mismatch negativity, P300, and N400. Clin Neurophysiol. 2009;120:1883–1908. doi:10.1016/j.clinph.2009.07.045
61. IBM. IBM SPSS advanced statistics 25. 2018. Available from: http://library.uvm.edu/services/statistics/SPSS22Manuals/IBM%20SPSS%20Advanced%20Statistics.pdf.
62. Oldfield RC. The assessment and analysis of handedness: the Edinburgh inventory. Neuropsychologia. 1971;9:97–113. doi:10.1016/0028-3932(71)90067-4
63. Veale JF. Edinburgh handedness inventory - short form: a revised version based on confirmatory factor analysis. Laterality. 2014;19:164–177. doi:10.1080/1357650X.2013.783045
64. Shaffer JP. Multiple hypothesis testing. Annu Rev Psychol. 1995;46:561–584. doi:10.1146/annurev.ps.46.020195.003021
65. Maroco J. Análise Estatística: com utilização do SPSS.
66. Abad EV, García MB. The locus of deficits in dysphonemic dyslexia: an ERP analysis. Psicothema. 2000;12(1):79–85.
67. Xue L, Zhao J, Weng X. Selective impairments in fine neural tuning for print in Chinese children with developmental dyslexia. Brain Sci. 2023;13(3):379. doi:10.3390/brainsci13030379
68. Papageorgiou C, Giannakakis GA, Nikita KS, et al. Abnormal auditory ERP N100 in children with dyslexia: comparison with their control siblings. Behav Brain Funct. 2009;5:26. doi:10.1186/1744-9081-5-26
69. Menashe S. Audiovisual processing and selective attention in adult dyslexic readers: an event-related potential study. Dyslexia. 2021;27(2):245–264. doi:10.1002/dys.1674
70. Banai K, Ahissar M. Poor frequency discrimination probes dyslexics with particularly impaired working memory. Audiol Neurootol. 2004;9(6):328–340. doi:10.1159/000081282
71. Bonte ML, Blomert L. Developmental changes in ERP correlates of spoken word recognition during early school years: a phonological priming study. Clin Neurophysiol. 2004b;115(2):409–423. doi:10.1016/S1388-2457(03)00361-4
72. Oades RD. Frontal, temporal and lateralized brain function in children with attention-deficit hyperactivity disorder: a psychophysiological and neuropsychological viewpoint on development. Behav Brain Res. 1998;94(1):83–95. doi:10.1016/S0166-4328(97)00172-1
73. Banaschewski T, Brandeis D. Annotation: what electrical brain activity tells us about brain function that other techniques cannot tell us – a child psychiatric perspective. JCPP. 2007;48(5):415–435. doi:10.1111/j.1469-7610.2006.01681.x
74. Brandeis D, Banaschewski T, Baving L, et al. Multicenter P300 brain mapping of impaired attention to cues in hyperkinetic children. JAACAP. 2002;41(8):990–998. doi:10.1097/00004583-200208000-00018
75. Trujillo-Orrego N. El N200: una señal electrofisiológica asociada con el control inhibitorio como candidato a endofenotipo del trastorno por déficit de atención/hiperactividad familiar. Rev Neurol. 2010;53(1):35–43. doi:10.33588/rn.5301.2010607
76. Rosburg T, Trautner P, Ludowig E, et al. Sensory gating in epilepsy – effects of the lateralization of hippocampal sclerosis. Clin Neurophysiol. 2008;119(6):1310–1319. doi:10.1016/j.clinph.2008.02.007
77. Dravet C. How Dravet syndrome became a model for studying childhood genetic epilepsies. Brain. 2012;135:2309–2311. doi:10.1093/brain/aws198
78. Yu FH, Mantegazza M, Westenbroek RE, et al. Reduced sodium current in GABAergic interneurons in a mouse model of severe myoclonic epilepsy in infancy. Nat Neurosci. 2006;9(9):1142–1149. doi:10.1038/nn1754
79. Garagnani M. Understanding Language and Attention: Brain-Based Model and Neurophysiological Experiments [Ph.D. thesis]. Cambridge: Wolfson College, UK; 2008.
80. Enoki H, Sanada S, Oka E, Ohtahara S. Effects of high-dose antiepileptic drugs on event-related potentials in epileptic children. Epilepsy Res. 1996;25(1):59–64. doi:10.1016/0920-1211(96)00025-3
81. Ozdag MF, Yorbik O, Ulas UH, et al. Effect of methylphenidate on auditory event related potential in boys with attention deficit hyperactivity disorder. Int J Pediatr Otorhinolaryngol. 2004;68(10):1267–1272. doi:10.1016/j.ijporl.2004.04.023
82. Sunohara GA, Malone MA, Rovet J, et al. Effect of methylphenidate on attention in children with attention deficit hyperactivity disorder (ADHD): ERP evidence. Neuropsychopharmacol. 1999;21(2):218–228. doi:10.1016/S0893-133X(99)00023-8
83. Verbaten MN, Overtoom CE, Koelega HS, et al. Methylphenidate influences on both early and late ERP waves of ADHD children in a continuous performance test. J Abnorm Child Psychol. 1994;22(5):561–578. doi:10.1007/BF02168938
84. Habib M. The neurological basis of developmental dyslexia: an overview and working hypothesis. Brain. 2000;123(12):2373–2399. doi:10.1093/brain/123.12.2373
85. Moisescu-Yiflach T, Pratt H. Auditory event related potentials and source current density estimation in phonologic/auditory dyslexics. Clin Neurophysiol. 2005;116(11):2632–2647. doi:10.1016/j.clinph.2005.08.006
86. Maurer U, Bucher K, Brem S, Brandeis D. Altered responses to tone and phoneme mismatch in kindergartners at familial dyslexia risk. Neuroreport. 2003;14(17):2245–2250. doi:10.1097/01.wnr.0000096518.69073.a7
87. Shankarnarayan VC, Maruthy S. Mismatch negativity in children with dyslexia speaking Indian languages. Behav Brain Funct. 2007;3:36. doi:10.1186/1744-9081-3-36
88. Kujala T, Belitz S, Tervaniemi M, Näätänen R. Auditory sensory memory disorder in dyslexic adults as indexed by the mismatch negativity. Eur J Neurosci. 2003;17(6):1323–1327. doi:10.1046/j.1460-9568.2003.02559.x
89. Schulte-Kӧrne G, Bruder J. Clinical neurophysiology of visual and auditory processing in dyslexia: a review. Clin Neurophysiol. 2010;121(11):1794–1809. doi:10.1016/j.clinph.2010.04.028
90. Näätänen R, Alho K. Mismatch negativity – the measure for central sound representation accuracy. Audiol Neurootol. 1997;2:341–353. doi:10.1159/000259255
91. Alonso-Búa B, Díaz F, Ferraces MJ. The contribution of AERPs (MMN and LDN) to studying temporal vs. linguistic processing deficits in children with reading difficulties. Int J Psychophysiol. 2006;59(2):159–167. doi:10.1016/j.ijpsycho.2005.03.020
92. Heim S, Eulitz C, Kaufmann J, et al. Atypical organization of the auditory cortex in dyslexia as revealed by MEG. Neuropsychologia. 2000;38(13):1749–1759. doi:10.1016/S0028-3932(00)00075-0
93. Paul I, Bott C, Heim S, et al. Phonological but not auditory discrimination is impaired in dyslexia. Eur J Neurosci. 2006;24(10):2945–2953. doi:10.1111/j.1460-9568.2006.05153.x
94. Friederici AD. Towards a neural basis of auditory sentence processing. TiCS. 2002;6(2):78–84.
95. Brauer J. Functional Development and Structural Maturation in the Brain’s Neural Network Underlying Language Comprehension [Ph.D. thesis in Cognitive Neuroscience (Dr. rer. nat.)]. Germany: University Leipzig; 2008.
96. Friederici AD. Pathways to language: fiber tracts in the human brain. TiCS. 2009;13(4):175–181.
97. Noordenbos MW, Segers E, Wagensveld B, Verhoeven L. Aberrant N400 responses to phonological overlap during rhyme judgements in children at risk for dyslexia. Brain Res. 2013;1537:233–243. doi:10.1016/j.brainres.2013.09.018
98. Parviainen T, Helenius P, Poskiparta E, et al. Cortical sequence of word perception in beginning readers. J Neurosci. 2006;26(22):6052–6061. doi:10.1523/JNEUROSCI.0673-06.2006
99. Duncan LG, Castro SL, Defior S, et al. Phonological development in relation to native language and literacy: variations on a theme in six alphabetic orthographies. Cognition. 2013;127:398–419. doi:10.1016/j.cognition.2013.02.009
100. Goswami U, Thomson J, Richardson U, et al. Amplitude envelope onsets and developmental dyslexia: a new hypothesis. PNAS. 2002;99(16):10911–10916. doi:10.1073/pnas.122368599
101. Holliman AJ, Wood C, Sheehy K. Does speech rhythm sensitivity predict children’s reading ability 1 year later? J Educ Psychol. 2010;102:356–366. doi:10.1037/a0018049
102. Harm MW, McCandliss BD, Seidenberg MS. Modeling the successes and failures of interventions for disabled readers. SSR. 2003;7(2):155–182.
103. Oana S, Inagaki M, Suzuki S, et al. Developmental changes of N400 event related potential of a semantic category decision task and modality specific findings in patients with developmental dyslexia. No to Hattatsu. 2006;38(6):431–438.
104. Fernandes T, Vale AP, Martins B, et al. The deficit of letter processing in developmental dyslexia: combining evidence from dyslexics, typical readers and illiterate adults. Dev Sci. 2014;17(1):125–141. doi:10.1111/desc.12102
105. Ahissar M, Lubin Y, Putter-Katz H, Karen K. Dyslexia and the failure to form a perceptual anchor. Nat Neurosci. 2006;9:1558–1564. doi:10.1038/nn1800
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms.php
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms.php
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.