")
Back to Journals » Infection and Drug Resistance » Volume 18
Construction and Characterization of an lpxM-Deficient Acinetobacter baumannii Strain Using a pyrF/5-FOA Counterselection System
Authors Gao C , Jiang K, Qi J, Zheng C , Liang Y
Received 19 February 2025
Accepted for publication 16 June 2025
Published 27 June 2025 Volume 2025:18 Pages 3175—3185
DOI https://doi.org/10.2147/IDR.S523844
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Sandip Patil
Can Gao,* Kun Jiang,* Jiaxin Qi,* Chang Zheng, Yanli Liang
Department of Clinical Laboratory, Suining Central Hospital, Suining, Sichuan, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Yanli Liang, Email [email protected]
Aim: Multidrug-resistant Acinetobacter baumannii (MDR-AB) is on the rise, making it challenging to achieve the desired therapeutic effects with existing conventional antibiotics. The search for new antibacterial targets has emerged as a significant research focus.
Purpose: The lysophospholipid acyltransferases (LPLATs) proteins encoded by the lpxM gene play a pivotal role in the biosynthesis of lipopolysaccharides (LPS). LPS is a critical component of the outer membrane of the cell wall and is essential for the survival and drug resistance of Gram-negative bacteria. This study aims to investigate the effects of the lpxM gene on the growth and drug susceptibility of MDR-AB.
Methods: The standard strain of Acinetobacter baumannii (A. baumannii, AB) AYE was selected as the target. The lpxM gene was knocked out using the pyrF/5-FOA-based counterselectable method. Subsequently, the growth status and the minimum inhibitory concentration (MIC) of the knockout strain against conventional antibiotics were compared.
Results: The lpxM gene in AB AYE was successfully and fully knocked out. The absorbance value at OD600 for the lpxM knockout strain during the stable period was observed to be as low as 2.5, indicating a significant reduction in growth rate. Furthermore, the MIC of the knockout strain for imipenem decreased from 16 μg/mL to 1 μg/mL, and the MIC for ceftazidime decreased from 32 μg/mL to 16 μg/mL, enhancing antibiotic sensitivity.
Conclusion: This study demonstrates that the deletion of the lpxM gene induces alterations in the growth and drug resistance of AB, providing a crucial foundation for further investigation into the mechanisms underlying LPS-mediated drug resistance and for the screening of effective auxiliary inhibitors targeting lpxM against MDR-AB.
Keywords: Acinetobacter baumannii, lpxM gene, pyrF/5-FOA, nutritional deficiencies, lipopolysaccharide
Introduction
Acinetobacter baumannii is one of the significant nosocomial opportunistic pathogens that primarily causes infections such as ventilator-associated pneumonia, wound infections, urinary tract infections, meningitis, and bacteremia, etc. In severe cases, these infections can be life-threatening. The escalating misuse and mismanagement of antibiotics have led to a rising prevalence of multidrug resistance in AB, with resistance now observed against the majority (95–99%) of antibiotics commonly used in clinical practice.1 Consequently, the current clinical challenges underscore the urgency and necessity of developing and implementing effective antibacterial strategies. LPS is the primary component of the cell wall in gram-negative bacteria. Additionally, LPS functions as an immune-reactive endotoxin capable of inducing inflammation within the host’s body, which, in severe cases, can progress to sepsis.2 Due to the unique structure and virulence of LPS, it has been identified as a potential target for the development of novel antibiotics.
The structure of LPS comprises three primary components: Lipid A, the O-antigen repeating unit, and core oligosaccharides. The O-antigen repeating unit is not essential, but Lipid A and core oligosaccharides are necessary for the survival of bacteria in the host. The modification of Lipid A directly influences the ability of pathogens to endure within the host by altering the permeability of the outer membrane, evading detection by the host immune system, and enhancing resistance to antimicrobial peptides.3 Zeng has identified that inhibitors targeting the fundamental enzymatic steps in Lipid A biosynthesis serve as effective antibacterial agents for the treatment of MDR-AB infections.4 In conclusion, LPS is integral to the virulence and drug resistance of AB.5
Needham’s research on E. coli K-12 demonstrated that LPS synthesis initiates with Kdo2-lipid IVA. The enzyme encoded by LpxL facilitates the transfer of a acyl group (C12:0) from the acyl carrier protein (ACP) to the R-2′-hydroxymyristate acyl chain of Kdo2-lipid IVA. Subsequently, the LPLATs protein encoded by lpxM incorporates a acyl group (C14:0) to the R-3′-hydroxymyristate residue of Kdo2-lipid A, resulting in hexa-acylation. Finally, the activation of PagP leads to the palmitoylation of the R-2′-hydroxymyristate primary acyl chain (C16:0). The lpxM gene is crucial for the synthesis of Kdo2-lipidA, which is the initial step in the LPS biosynthesis pathway. It is considered a potential virulence factor associated with lipid A, significantly influencing the structural and functional properties of LPS.6 One study has shown that mutations in the lpxM gene can inhibit MDR-AB in synergy with polymyxin by affecting the synthesis of Lipid A in LPS. Currently, polymyxin is utilized as the last line of treatment against multidrug-resistant Gram-negative bacteria. However, bacteria such as AB gradually develop resistance to polymyxin when subjected to selection pressure.7 Therefore, it is essential to further explore the impact of lpxM gene deletion on the growth of AB and its sensitivity to other commonly used antibiotics in clinical practice to guide innovative treatment strategies.
The pyrF/5-FOA-based counterselectable knockout system is a widely utilized method for gene mutation, facilitating succinct, efficient, and trace-free gene editing. This enhances the accuracy of research into bacterial gene function.8 In this study, the lpxM gene of AB was successfully knocked out without detectable traces, employing the pyrF/5-FOA-based counterselectable method established by Wang et al.8 The objective of this investigation was to evaluate the effects of lpxM gene deletion on the growth characteristics and drug susceptibility profiles of MDR-AB. This research aims to provide a foundational understanding for further exploration of resistance mechanisms in AB and to identify novel drug targets. Furthermore, the findings may serve as a critical reference for the development of new antibiotics in clinical settings.
Materials and Methods
Main Reagents
LB medium (purchased from OXOID, UK), MH solid medium (purchased from OXOID, UK), MH broth medium (purchased from Qingdao Haibo Biotechnology Co., Ltd.), 1 kb Plus DNA Ladder (purchased from Chengdu Tiangen Biochemistry & Technology Co., Ltd.), Pst I, BamH I (purchased from Thermo Scientific), C115 recombinase (purchased from Nanjing Novozymes), PCR (polymerase chain reaction) product purification kit (purchased from Shanghai Sangong Biotechnology Engineering Co. Scientific), plasmid extraction kit (purchased from OMEGA), 5-fluoroorotic acid (5-FOA, purchased from Shanghai Sangong Bioengineering Co., Ltd.), Tellurite (purchased from Shanghai Sangong Bioengineering Co., Ltd.), Uracil (purchased from Shanghai Sangong Bioengineering Co., Ltd.), etc. Detailed formulations for the preparation of antibiotics and culture media are provided in the Supplementary Material 1.
Main Instruments and Consumables
PCR Instrument (Eppendorf, Germany), Horizontal Electrophoresis System (Bio-Rad, USA), Benchtop Centrifuge (Eppendorf, Germany), Multifunctional Gel Imager (Bio-Rad, USA), Thermostatic Metal Bath (Hangzhou BORI Science and Technology Co., Ltd.), Growth Curve Analyzer (BMG, Germany), 96-well Plate (Chengdu Ruiyin Bio-tech Co., Ltd.), MIC Multipoint Inoculator (Henao Instruments, Ltd.), etc.
Strain, Plasmid, and Primers
The lpxM gene sequence of Acinetobacter baumannii strain AYE was downloaded from the National Center for Biotechnology Information (NCBI) website (FASTA record: YP_001712836), with a sequence length of 984 bp. The lpxM knockout strain was constructed based on the AB AYE ∆pyrF knockout strain stored in the Key Laboratory of Non-coding RNA and Drugs at Chengdu Medical College. The lpxM knockout plasmid was developed using the pMO130TF plasmid, also stored in this laboratory, as detailed in Table 1. The experimental amplification and validation primers were synthesized by Shanghai Sangong Bioengineering Co., Ltd., as presented in Table 2. The details of the PCR system and procedure can be found in Supplementary Material 2.
 |
Table 1 Strains and Plasmids |
 |
Table 2 Sequence of Primers for PCR Amplification |
Experimental Methods
lpxM Gene Knockout
The orotic acid phosphoribosyl transferase (OPRTase) encoded by the pyrE gene catalyzes the conversion of orotic acid to its normal product, orotic acid-5′-monophosphate (OMP). Additionally, OPRTase mediates the conversion of 5-fluoro orotidine 5′-monophosphate (5-FOMP) from the substrate 5-fluoro-orotic acid (5-FOA). 5-FOMP plays a crucial role in the de novo biosynthesis of pyrimidines through the action of the OMP decarboxylase protein encoded by the pyrF gene. This process is essential for the regulation of body growth and survival. However, the accumulation of 5-FOMP can lead to toxicity, culminating in cell death when the OMP decarboxylase enzyme is inactivated. In the initial phase of this experimental study, a suicide plasmid was constructed by amplifying the upstream and downstream fragments of the lpxM gene. Subsequently, E. coli 47055 was utilized as a vector to facilitate the binding of the suicide plasmid to the AB AYEF strain, which possessed a deletion of the pyrF gene. This marked the first occurrence of homologous recombination. Following this, the bacterial genome underwent a second round of endogenous recombination under conditions of uracil nutritional deficiency and 5-FOA counterselection pressure (Figure 1). Ultimately, PCR screening and verification of knockout clones were conducted.9
 |
Figure 1 Schematic diagram of the knockout of the lpxM gene using the pyrF/5-FOA-based counterselectable method. |
Construction of Knockout Vectors
The DNA from the AB AYE strain served as a template for amplifying the upstream region of the lpxM gene using the lpxM -UF and lpxM -UR primers. Similarly, the downstream region of the lpxM gene was amplified using the lpxM-DF and lpxM-DR primers. The resulting fragments were recovered following electrophoretic verification. The plasmid pMO130TF was digested with BamH I and Pst I restriction enzymes, and subsequent electrophoretic analysis confirmed the successful recovery. The recovered vectors and fragments were then reconstructed. (Figure 2). The resultant recombinant constructs were transferred into E. coli DH5α, which were then plated onto LB agar plates supplemented with kanamycin and cultured overnight at 37°C. Monoclones were isolated and selected for PCR-based identification. Following confirmation, the selected clones were inoculated into LB liquid broth supplemented with kanamycin and cultured overnight. The plasmids were extracted and subsequently submitted to Chengdu Qingke Biotechnology Co., Ltd. for DNA sequencing analysis, utilizing the sequencing primer lpxM-UF. The sequencing analysis was conducted using SnapGene software for pairwise sequence alignments. The sequencing results exhibited a high degree of congruence with the upstream and downstream sequences of lpxM, confirming the successful construction of the pMo130TF-ΔlpxM UD plasmid.
 |
Figure 2 Schematic diagram of pMo130TF-ΔlpxM UD plasmid construction. |
Conversion and Bonding
The pMo130TF-ΔlpxM UD plasmid was co-mixed with the E. coli 47055 strain in a receptive state. Subsequently, the recombinant plasmid was chemically transformed into the receptive E. coli 47055 via a heat shock method, conducted at 42°C for 90 seconds in a metal bath. Following this, LB liquid medium was added to the transformed bacterial suspension, which was then shaked (200 rpm) at 37°C for 45 minutes to facilitate resuscitation. The bacterial solution, precipitated by centrifugation, was uniformly coated onto a Kanamycin LB plate and cultured overnight at 37°C. Both pMo130TF-ΔlpxM UD 47055 and AYEF were inoculated with antibiotics and cultured overnight. The bacterial solutions of 47055 and AYEF were mixed in a 2:1 ratio. Subsequently, the cultures were centrifuged, and the pellets were re-suspended in LB liquid medium, followed by two washes. The resulting precipitation was applied onto sterile NC film placed on the LB plate, which was then allowed to air-dry, inverted, and cultured at 37°C for at least 12 hours. The bacterial solution was scraped, centrifuged, washed, and evenly coated on a resistant plate containing Tellurite and Gentamicin, and cultured overnight at 37°C. The black clones were simultaneously lined on the Tellurite and Gentamicin plates and cultured overnight at 37°C. Clones grown in both Tellurite and Gentamicin plates, and displayed a yellow coloration, were indicative of a successful bonding event. The successful bonding was verified using lpxM-UF and lpxM-DR primers.
Double-Exchange Knockout
The successfully bonded clones were inoculated in LB liquid medium supplemented with 5-FOA and uracil, and cultured at 37°C overnight. Subsequently, the bacterial suspension was passaged 2–3 times. A portion of the bacterial culture was then diluted 106 with sterile normal saline and applied to an LB agar plate containing 5-FOA, which was cultured overnight at 37°C. The white clones were simultaneously lined on Tellurite + Gentamicin plates and cultured at 37°C for at least 12 hours. The strains that exhibited growth on the LB plate but not on the Tellurite plate were preliminarily identified as potential plasmid screening dropout strains. lpxM-UF and lpxM-DR primers were employed to verify the clones via PCR, and final sequence analysis was conducted to determine whether the clones were indeed knockout strains.
Growth Curve Experiment
The bacterial suspension was diluted to a concentration of 0.5 McFarland, followed by a 20-fold dilution. Subsequently, 190 μL of LB liquid medium was combined with 10 μL of the diluted bacterial solution, which was then added to each well of the 96-well plate. The resultant bacterial concentration per well was approximately 5×105 CFU/mL. The OD600 absorbance was measured using a growth curve analyzer, while the culture was subjected to oscillation at 200 rpm and maintained at 37°C. Measurements were taken every 30 minutes over a continuous period of 24 hours. Statistical analysis was conducted using GraphPad Prism 6 software, employing one-way ANOVA to compare multiple groups and a T-test for comparisons between two groups.
MIC Experiment
The Minimum Inhibitory Concentration (MIC) value of AB AYE against conventional laboratory antibiotics was determined using AGAR dilution method. The types of antibiotics used in this experiment and their mechanism of action were shown in Table 3. A stock solution of the antibiotics was prepared at a concentration of 64 mg/mL, which was then serially diluted to achieve a final concentration of 0.0625 mg/mL through continuous double dilution. A drug-sensitive plate was prepared by adding various concentrations of the antibiotic solution into Mueller-Hinton (MH) medium. As in the previous steps of the growth curve experiment, the bacterial solution was diluted to ensure that the bacterial concentration in each well of the sterile 96-well plate was approximately 5×105 CFU/mL. A MIC multipoint inoculator was employed to inoculate the bacterial solution for drug sensitivity testing. After the bacterial solution was absorbed, the incubator was inverted and cultured at 37°C for 16 to 20 hours. The MIC value was determined as the minimum drug concentration that resulted in no bacterial plaque formation at all three replicate points, which was then used for statistical analysis. Strain sensitivity was assessed according to the Clinical and 2023 Laboratory Standards Institute (CLSI) M0210 diffusion method for antimicrobial susceptibility testing. E. coli ATCC® 25922 was utilized as the control strain to verify that the MIC values fell within the acceptable quality control (QC) ranges. If the MIC values exceeded the defined QC ranges, the experimental data were considered invalid.
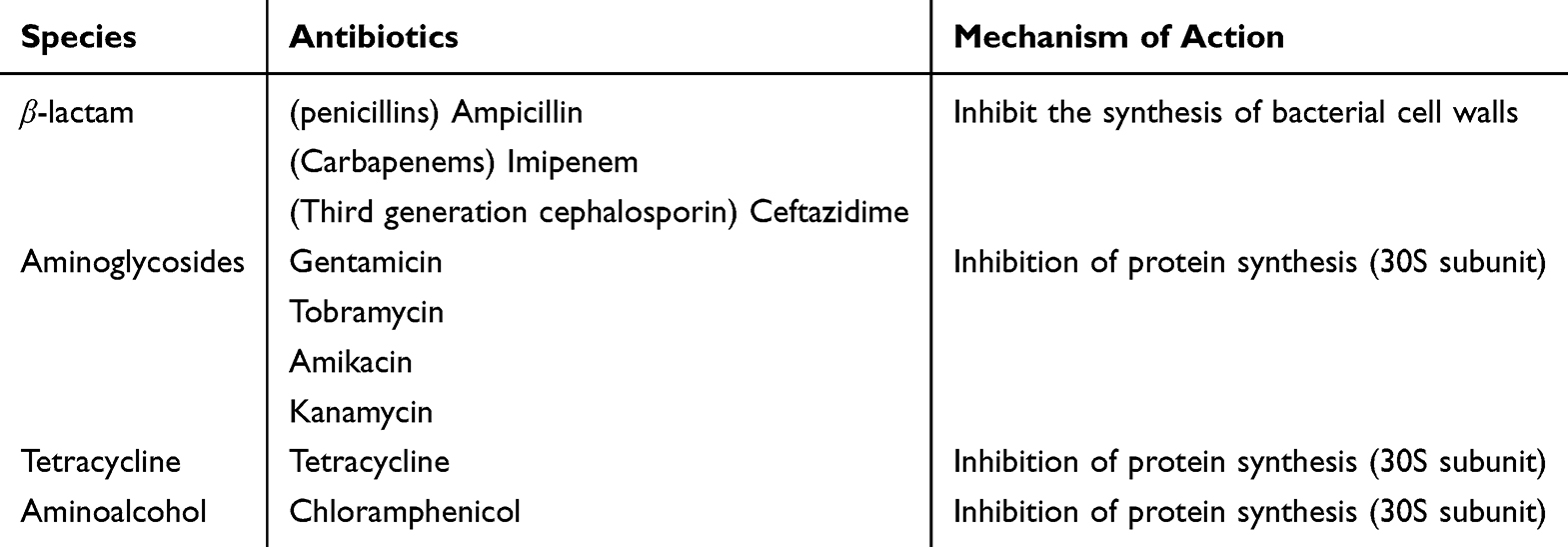 |
Table 3 Routine Antibiotics and Mechanism of action11 |
Results
lpxM Gene Knockout results
In this study, the lpxM gene of AB AYE was successfully knocked out using a double exchange homologous recombination system established in the laboratory. Verification was conducted using lpxM-UF and lpxM-DR primers. The results indicated that the PCR product obtained from the knockout strain measured approximately 1.9 kb, which aligned with the length of the PCR product derived from the knockout vector in the positive control group. Additionally, the target fragment in the knockout strain was approximately 1 kb shorter than that observed in the negative control (Figure 3A). Sequencing results confirmed the absence of the lpxM gene sequence in the strain’s genome (Figure 3B), indicating that the lpxM gene has been successfully and completely knocked out.
 |
Figure 3 lpxM gene knockout validation picture. Notes: (A) lpxM gene knockout verification electrophoresis diagram, lanes 1–4 serve as marker (1 kb Plus DNA Ladder), ΔlpxM knockout strain (AYEF), positive control knockout vector, negative control AYEF strain; (B) DNA sequencing results of ΔlpxM knockout strain. |
The Growth State of the lpxM Gene Knockout Strain Exhibited Poor Conditions
In order to ascertain the importance of the lpxM gene on bacterial growth, comparative growth curves were analyzed for the wild-type AYE and AYEF strains, as well as for the ΔlpxM-AYEF mutant strains. To maintain the experimental principle of a “single variable”, exogenous uracil was incorporated into the culture medium of the ΔlpxM knockout strain. This addition aimed to eliminate bacterial growth and drug sensitivity changes as potential confounding factors resulting from insufficient uracil synthesis following the pyrF gene knockout. As depicted in Figure 4, the AYEF strain exhibited limited growth in the absence of pyrimidine synthesis. In contrast, the AYEF strain with the addition of exogenous uracil (AYEF+Uracil) and the AYE strain demonstrated comparable growth states. After approximately 13 hours, the culture entered a stable growth phase, during which the OD600 absorbance value reached 3.0. However, the ΔlpxM-AYEF knockout strains did not recover to a normal growth state following the addition of uracil (ΔlpxM-AYEF+Uracil), with the OD600 absorbance peaking at a low of 2.5 approximately 15 hours later. The OD600 absorbance values for the AYEF+Uracil and ΔlpxM-AYEF+Uracil strains underwent T-test analysis, revealing a highly significant difference (**** P<0.0001, suggesting a statistically significant distinction between the two groups.
 |
Figure 4 Growth curves of AB after different treatments. Notes: AYE is the wild-type strain, AYEF is the ΔpyrF knockout strain, AYEF+Uracil is the addition of Uracil in the AYEF strain medium, ΔlpxM-AYEF+Uracil represents the strain with a knockout of the lpxM gene, which grown in a medium supplemented with uracil. |
The Sensitivity of Bacteria to Certain Antibiotics was Significantly Elevated Subsequent to the Knockout of lpxM Gene
According to the 2023 CLSI M02 drug sensitivity test standard, the MIC test results for the AB AYE strain are presented in Table 4. The strains exhibited resistance to multiple antibiotics, including β-lactam, Aminoglycosides, Tetracycline, and Aminoalcohol, demonstrating a characteristic of multidrug resistance. Notably, following the knockout of the lpxM gene, the MIC value of the strain for the β-lactam carbapenem subclass antibiotic imipenem decreased from 16 μg/mL to 1 μg/mL, indicating a shift from drug resistance to sensitivity. Similarly, the MIC value of the strain for the third-generation cephalosporin subclass antibiotic ceftazidime, also within the β-lactam class, decreased from 32 μg/mL to 16 μg/mL, reflecting a transition from resistance to moderate sensitivity.
 |
Table 4 MIC Results of Wild and Mutant Δlpxm Gene of AB AYE (μg/mL) |
Discussion
AB has emerged as the predominant opportunistic pathogen responsible for nosocomial infections, characterized by its exceptional survivability and aggressiveness.9 The extensive use of antibiotics has exacerbated the issue of resistance, progressively escalating AB into a “superbug” marked by multidrug resistance and enhanced virulence. Current statistics indicate that approximately 1 million individuals worldwide are infected with AB annually, with half of these infections attributed to multidrug-resistant strains.12 The mortality rate can reach as high as 84.3% for patients infected with extensively drug-resistant Acinetobacter baumannii (XDR-AB).13 The threat posed by AB to human health, along with the urgent need for the development of related antibiotics, along with the urgent need for the development of related antibiotics ranked first.12 However, few existing antibiotics are effective in treating multidrug-resistant bacterial infections, and the diversity and prevalence of drug-resistant pathogens continue to rise.14 Currently, there are three main strategies aimed at combating multidrug-resistant (MDR) bacteria: the discovery of novel antibiotics, the development of alternatives to existing antibiotics, and the identification of new antibiotic adjuvants. First, uncovering new antibiotics with unique targets presents a formidable challenge; over the past few decades, the US Food and Drug Administration (FDA) has not approved any new antibiotics specifically targeting Gram-negative bacteria, and relatively few new antibiotics are currently under development.15,16 Second, antibiotic alternatives, such as antibodies, probiotics, and vaccines, are increasingly being considered more suitable for clinical use as prophylactic therapies.17 Therefore, the pursuit of novel targets capable of inhibiting bacterial growth, drug resistance, or virulence factors to develop antibiotic adjuvants dominates contemporary drug research and development.
LPS plays a critical role in bacterial survival and virulence, representing one of the pivotal mechanisms contributing to drug resistance in AB. LPS is essential for transforming the bacterial outer membrane (OM) into an effective osmotic barrier composed of small hydrophobic molecules, thereby protecting the cell from the entry of toxic substances such as antibiotics and bile salts.18 Chui et al demonstrated that structural changes in LPS could significantly enhance the sensitivity of AB to a range of clinically relevant antibiotics, including polymyxin and rifampicin.19,20 Modifying LPS holds promise as a strategy to combat inflammation and bacterial resistance. Blocking LPS synthesis represents an effective approach for discovering new antibiotics. However, LPS deletion strains of AB isolated from clinical specimens are infrequently documented. Most such strains have been induced by colistin in vitro, and the resulting mutations may lead to other unknown genetic alterations, creating a complex and unstable genome.21 In this study, the pyrF/5-FOA-based counterselectable technique was employed to efficiently and selectively isolate lpxM gene knockout strains, providing an effective tool for constructing LPS deletion strains of AB. The growth curve analysis revealed that the growth rate of the lpxM knockout strain was notably slower compared to that of the wild-type strain. The wild-type strain reached a stable growth phase after 13 hours of culture, with an OD600 absorbance value of 3.0, while the knockout strain achieved stability after 15 hours of culture, at which point its OD600 absorbance value was 2.5. The mechanism by which the lpxM gene influences the survival and drug resistance of AB remains unclear. It is hypothesized that the reason for the inhibition of the growth of the knockout strain may be as follows: Lipid A is the most conservative component of LPS. Deletion of lpxM gene results in the absence of myristic acid in lipid A synthesis, leading to a transition from a hexa-acylated to a penta-acylated structure. The alteration of the acylation mode interrupts the synthesis of key fatty acid chains, consequently disrupting the hydrophobic core symmetry of lipid A and reducing the stability and membrane structure of LPS. Knockout strains may struggle to maintain normal morphology, which directly impacts their ability to divide and proliferate.22
Furthermore, the results of drug sensitivity experiments indicated that the deletion of the lpxM gene in AB enhanced its sensitivity to certain antibiotics, notably reversing the organism’s resistance to imipenem and transitioning it from a state of resistance to susceptibility.
These findings align with the conclusions reported by Sugawara, who observed that the deletion of the lpxM gene in E. coli resulted in a growth delay and increased sensitivity to multiple antibiotics.23 The knockout strains exhibited increased sensitivity to certain β-lactam antibiotics, but no significant changes were observed for aminoglycosides, tetracyclines, and aminoalcohols. This differential sensitivity can be attributed to the distinct mechanisms of action of these antibiotic classes. Aminoglycosides utilize active transport systems for cellular entry, tetracyclines employ both passive diffusion and active transport mechanisms, and aminoalcohols exert their effects through inhibition of protein synthesis by binding to 50S ribosomal subunits. These uptake mechanisms exhibit minimal dependence on outer membrane permeability, explaining the limited impact of lpxM knockout on their efficacy. The mechanism of action of β-lactam antibiotics is primarily associated with the synthesis of bacterial cell walls (Table 3). The absence of lpxM can result in the lack of secondary acyl chains, such as C14:0, in lipid A. Incomplete acylation of lipid A may expose additional negatively charged sites, thereby rendering the outer membrane more susceptible to penetration by hydrophilic antibiotics, such as β-lactam antibiotics, which can then enter bacterial cells and exert their effects, ultimately reducing bacterial drug resistance.24 Notably, the findings of this study indicate that knockout strains exhibit altered sensitivity to certain β-lactam drugs, including imipenem and ceftazidime, while showing no effect on ampicillin. Possible explanations for this observation include: imipenem and ceftazidime possess strong activity against Gram-negative bacteria and primarily enter the bacteria by directly penetrating the outer membrane.25 Furthermore, structural changes in LPS may influence the expression or conformation of penicillin-binding proteins (PBPs). Imipenem and ceftazidime demonstrate higher affinities for specific PBPs, such as PBP2 and PBP3, and their activities are directly influenced by the state of PBPs.26 The deletion of lpxM may alter the permeability of the outer membrane and affect the synthesis of PBPs, thereby impacting the uptake efficiency of these two drugs. However, the ability of ampicillin to penetrate the outer membrane of Gram-negative bacteria is weaker than that of imipenem and ceftazidime. The resistance mechanism of ampicillin primarily relies on specific pore protein channels, such as OmpF and OmpC, and more significantly on the hydrolysis of the β-lactam ring by β-lactamases, including TEM-1 and SHV-1, rather than solely depending on the permeability of the outer membrane. Even in the absence of lpxM, bacteria can remain resistant if they possess the β-lactamase gene.27 The impact of lpxM deletion on pore protein channels and β-lactamases is limited and insufficient to significantly enhance the uptake efficiency of ampicillin. The specific influence mechanism should be considered a potential avenue for future research.
Targeting lpxM as an antimicrobial target is currently a prominent area of research, particularly for Gram-negative bacteria. Andrey et al demonstrated that the knockout of the lpxM gene in Yersinia pestis significantly reduces the virulence of the mutant strain by 2.5 to 16-fold, while concomitantly enhancing immunoprotective efficacy in mice.28 Rossi et al found that the lpxM gene of Shigella flexneri is pivotal in mediating its inflammatory response, and the inactivation of this gene could diminish the secretion of TNF-α from human monocytes and weaken the inflammatory destruction of the intestinal epithelial barrier.29 Pan et al found that the deletion of lpxM may enhance the immune recognition of AB by host cells. The five-acylated lipid A is recognized more readily by the host TLR4/MD-2 complex, resulting in a three- to five-fold increase in the secretion of the pro-inflammatory factor IL-6.30,31 Boll elucidated that Lipid A, which is acetylated by lpxM -synthesizing proteins, plays a critical role in the desiccation tolerance of AB and constitutes a vital resistance mechanism that enables its persistence within the hospital environment. Inhibition of lpxMab expression can synergistically enhance the efficacy of polymyxin B, presenting an innovative strategy for controlling the spread and prevention of MDR-AB infections.7 Both domestic and international research have shown a correlation between the lpxM gene and bacterial virulence, inflammatory responses, and various biological activities. The results of this experiment also prove that the knockout of the target gene lpxM significantly affects bacterial growth and their resistance to certain antibiotics. This finding suggests that the lpxM gene may serve as a target for screening lead compounds of imipenem and ceftazidime in the development of novel antibiotics.
Conclusion
The full lpxM knockout strain of AB AYE was constructed using the pyrF/5-FOA-based counterselectable system. The growth characteristics and drug susceptibility profiles of the knockout strains were compared with those of the wild-type strains. The results indicated that the ΔlpxM knockout strain exhibited a slower growth rate and increased sensitivity to certain antibiotics. The lpxM gene is proposed as a potential target for the development of novel therapeutics against MDR-AB. However, the present study only assessed the growth and drug sensitivity of the knockout strains, which limits the scope of the investigation. This focus did not extend to the detection of biofilm formation. Additionally, homologous modeling of the LPLATs protein was not conducted, thereby precluding the screening of potential small molecule ligands. These limitations may have hindered a comprehensive understanding of the strain’s characteristics and potential therapeutic applications. Nonetheless, the experimental results suggest that the lpxM gene plays a pivotal role in the growth and drug resistance of AB. This finding presents an effective knockout strategy for subsequent investigations into the alterations in compensatory pathways that occur following the deletion of LPS in AB, and establishing a solid foundation for demonstrating that the lpxM gene may serve as a potential target for the development of novel drugs against MDR-AB. Furthermore, it underscores a broader direction for the future development of preventive and therapeutic strategies against MDR-AB infections.
Acknowledgments
Non-coding RNA and Drug Discovery Key Laboratory of Sichuan Province, Chengdu Medical College: Provide experimental platform. Can Gao: Methodology, Data curation, Formal analysis, Writing original draft. Chang Zheng: Methodology, Data curation. Jiaxin Qi: Methodology, Resources. Kun Jiang: Language help. Yanli Liang: Conceptualization, Supervision. All authors have read and approved the final article.
Funding
This work was supported by Grants from the National Natural Science Foundation of China (grant 31701078), 2024 Suining Health Science and Technology Plan Project (Guiding Project) (grant 24GGWS04).
Disclosure
The authors declare that there is no conflict of interest.
References
1. Ibrahim S, Al-Saryi N, Al-Kadmy IMS, Aziz SN. Multidrug-resistant Acinetobacter baumannii as an emerging concern in hospitals. Mol Biol Rep. 2021;48(10):6987–6998. doi:10.1007/s11033-021-06690-6
2. Skrzypczak-Wiercioch A, Sałat K. Lipopolysaccharide-induced model of neuroinflammation: mechanisms of action, research application and future directions for its use. Molecules. 2022;27(17):5481. doi:10.3390/molecules27175481
3. Whitfield C, Trent MS. Biosynthesis and export of bacterial lipopolysaccharides. Annu Rev Biochem. 2014;83:99–128. doi:10.1146/annurev-biochem-060713-035600
4. Zeng D, Zhao J, Chung HS, Guan Z, Raetz CR, Zhou P. Mutants resistant to LpxC inhibitors by rebalancing cellular homeostasis. J Biol Chem. 2013;288(8):5475–5486. doi:10.1074/jbc.M112.447607
5. Kamoshida G, Akaji T, Takemoto N, et al. Lipopolysaccharide-deficient Acinetobacter baumannii due to colistin resistance is killed by neutrophil-produced lysozyme. Front Microbiol. 2020;11:573. doi:10.3389/fmicb.2020.00573
6. Yang J, Chu P, Song S, et al. Construction and biological characteristics analysis of lpxM gene deletion strains of Haemophilus parasuis. Chin J Agric Sci. 2020;53(16):3394–3403.
7. Boll JM, Tucker AT, Klein DR, et al. Reinforcing lipid A acylation on the cell surface of Acinetobacter baumannii promotes cationic antimicrobial peptide resistance and desiccation survival. mBio. 2015;6(3):e00478. doi:10.1128/mBio.00478-15
8. Wu S, Xu R, Su M, et al. A pyrF-based efficient genetic manipulation platform in Acinetobacter baumannii to explore the vital DNA components of adaptive immunity for I-F CRISPR-Cas. Microbiol Spectr. 2022;10(5):e0195722. doi:10.1128/spectrum.01957-22
9. Yang Y, Sun Q, Liu Y, et al. Development of a pyrF-based counterselectable system for targeted gene deletion in Streptomyces rimosus. J Zhejiang Univ Sci B. 2021;22(5):383–396. doi:10.1631/jzus.B2000606
10. Clinical and Laboratory Standards Association, CLSI M02. Drug susceptibility testing standard 2023 edition. 2023.
11. Baquero F, Levin BR. Proximate and ultimate causes of the bactericidal action of antibiotics. Nat Rev Microbiol. 2021;19(2):123–132. doi:10.1038/s41579-020-00443-1
12. Willyard C. The drug-resistant bacteria that pose the greatest health threats. Nature. 2017;543(7643):15. doi:10.1038/nature.2017.21550
13. Uwingabiye J, Frikh M, Lemnouer A, et al. Acinetobacter infections prevalence and frequency of the antibiotics resistance: comparative study of intensive care units versus other hospital units. Pan Afr Med J. 2016;23:191. doi:10.11604/pamj.2016.23.191.7915
14. Shatalin K, Nuthanakanti A, Kaushik A, et al. Inhibitors of bacterial HS biogenesis targeting antibiotic resistance and tolerance. Science. 2021;372(6547):1169–1175. doi:10.1126/science.abd8377
15. Brown ED. Gram-negative resistance. ACS Infect Dis. 2015;1(11):507. doi:10.1021/acsinfecdis.5b00123
16. Escolà-Vergé L, Los-Arcos I, Almirante B. New antibiotics for the treatment of infections by multidrug-resistant microorganisms. Med Clin. 2020;154(9):351–357. doi:10.1016/j.medcli.2019.11.002
17. Gupta R, Sharma S. Role of alternatives to antibiotics in mitigating the antimicrobial resistance crisis. Indian J Med Res. 2022;156(3):464–477. doi:10.4103/ijmr.IJMR_3514_20
18. Guest RL, Rutherford ST, Silhavy TJ. Border control: regulating LPS biogenesis. Trends Microbiol. 2021;29(4):334–345. doi:10.1016/j.tim.2020.09.008
19. Moffatt JH, Marina H, Paul H, et al. Colistin resistance in Acinetobacter baumannii is mediated by complete loss of lipopolysaccharide production. Antimicrob Agents Chemother. 2010;54(12):4971–4977. doi:10.1128/AAC.00834-10
20. Chin CY, Gregg KA, Napier BA, Ernst RK, Weiss DS. A PmrB-regulated deacetylase required for lipid A modification and polymyxin resistance in Acinetobacter baumannii. Antimicrob Agents Chemother. 2015;59(12):7911–7914. doi:10.1128/AAC.00515-15
21. Eisenreich W, Rudel T, Heesemann J, Goebel W. Link between antibiotic persistence and antibiotic resistance in bacterial pathogens. Front Cell Infect Microbiol. 2022;12:900848. doi:10.3389/fcimb.2022.900848
22. Needham BD, Trent MS. Fortifying the barrier: the impact of lipid A remodelling on bacterial pathogenesis. Nat Rev Microbiol. 2013;11(7):467–481. doi:10.1038/nrmicro3047
23. Sugawara T, Onoue S, Takimoto H, Kawahara K. Modification of lipid A structure and activity by the introduction of palmitoyltransferase gene to the acyltransferase-knockout mutant of Escherichia coli. Microbiol Immunol. 2018;62(8):497–506. doi:10.1111/1348-0421.12631
24. Nikaido H. Molecular basis of bacterial outer membrane permeability revisited. Microbiol Mol Biol Rev. 2003;67(4):593–656. doi:10.1128/MMBR.67.4.593-656.2003
25. Freed S, Hanson ND. AmpC induction by imipenem in Pseudomonas aeruginosa occurs in the absence of OprD and impacts imipenem/relebactam susceptibility. Microbiol Spectr. 2024;12(11):e0014224. doi:10.1128/spectrum.00142-24
26. van Langevelde P, Kwappenberg KM, Groeneveld PH, Mattie H, van Dissel JT. Antibiotic-induced lipopolysaccharide (LPS) release from Salmonella typhi: delay between killing by ceftazidime and imipenem and release of LPS. Antimicrob Agents Chemother. 1998;42(4):739–743. doi:10.1128/AAC.42.4.739
27. Bush K, Jacoby GA. Updated functional classification of beta-lactamases. Antimicrob Agents Chemother. 2010;54(3):969–976. doi:10.1128/AAC.01009-09
28. Anisimov AP, Shaikhutdinova RZ, Pan’kina LN, et al. Effect of deletion of the lpxM gene on virulence and vaccine potential of Yersinia pestis in mice. J Med Microbiol. 2007;56(Pt 4):443–453. doi:10.1099/jmm.0.46880-0
29. Rossi O, Pesce I, Giannelli C, et al. Modulation of endotoxicity of Shigella generalized modules for membrane antigens (GMMA) by genetic lipid A modifications: relative activation of TLR4 and TLR2 pathways in different mutants. J Biol Chem. 2014;289(36):24922–24935. doi:10.1074/jbc.M114.566570
30. Pan S, Peng L, Yi Q, et al. Ginsenoside Rh(2) Alleviates LPS-induced inflammatory responses by binding to TLR(4)/MD-2 and Blocking TLR(4) Dimerization. Int J Mol Sci. 2024;25(17):9546. doi:10.3390/ijms25179546
31. Park BS, Song DH, Kim HM, Choi BS, Lee H, Lee JO. The structural basis of lipopolysaccharide recognition by the TLR4-MD-2 complex. Nature. 2009;458(7242):1191–1195. doi:10.1038/nature07830
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms.php
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms.php
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.