")
Back to Journals » Drug Design, Development and Therapy » Volume 19
Development and Characterization of LasR Immobilized Monolithic Column for Screening Active Ingredients as Quorum Sensing Inhibitors Against P. aeruginosa in Natural Products
Authors Liu Z, Yang Y, Xie X, Li R, You J , Zhao X, Wang Y, Guo J
Received 18 October 2024
Accepted for publication 1 March 2025
Published 18 March 2025 Volume 2025:19 Pages 2051—2064
DOI https://doi.org/10.2147/DDDT.S501621
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Solomon Tadesse Zeleke
Zheng Liu,1,* Yue Yang,1,* Xiaoyuan Xie,1 Rui Li,1 Jifeng You,1 Xianglong Zhao,1,2 Yuanyuan Wang,1 Jialiang Guo1– 3
1School of Medicine, Foshan University, Foshan, 528000, People’s Republic of China; 2College of Pharmacy, Jinan University, Guangzhou, 510632, People’s Republic of China; 3Guangdong Provincial Engineering Technology Research Center of Whole Process Quality Control and Analysis of Lingnan Traditional Chinese Medicine, Foshan, 528225, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Jialiang Guo; Yuanyuan Wang, Email [email protected]; [email protected]
Background and Aim: The enzyme/protein immobilized monolithic capillary combined with liquid chromatography-mass spectrometry is an efficient screening strategy for the corresponding agonist/antagonist. LasR is the potential therapeutic target since it plays a vital role in the colonization and invasion of Pseudomonas aeruginosa (P. aeruginosa). Therefore, reagents that inhibit LasR may be effective against P. aeruginosa. To screen and find LasR inhibitors rapidly, a LasR-immobilized monolithic capillary column was prepared and characterized.
Methods: Firstly, the recombinant LasR protein was prepared in E. coli. Then, the LasR protein was immobilized to the surface of poly (glycidyl methacrylate-co-poly(ethylene glycol)diacrylate)-ethylenediamine monolithic column. The affinity and stability of prepared column was also evaluated. Furthermore, the prepared column was applied to fishing LasR inhibitor in Scutellaria baicalensis Georgi extract. The interaction of the screening compound to LasR was confirmed through molecular docking.
Results: The recombinant active LasR protein was prepared in E. coli. After purification and validation, a comparative ligand fishing monolithic column was prepared through immobilizing LasR to the surface of the poly (glycidyl methacrylate-co-poly(ethylene glycol)diacrylate)-ethylenediamine through amidation reaction. The LasR was successfully immobilized to the monolithic column characterizing by Fourier transform infrared spectroscopy and scanning electron microscopy. The activity of immobilized LasR was reserved as it has affinity to the nature ligand 3-oxo-C12-HSL and stablied within 24 h at 4 °C. In the Scutellaria baicalensis Georgi extract, baicalein was screened as a potential LasR inhibitor. The molecular docking results and the in vivo evaluation confirmed the activity of baicalein.
Conclusion: The proposed LasR immobilized monolithic column is a viable strategy in screening LasR inhibitors. It can be considered as a possible alternative to traditional methods for screening LasR inhibitors as drug candidates against P. aeruginosa.
Keywords: immobilized LasR monolithic column, quorum sensing, Pseudomonas aeruginosa, screening, natural products
Graphical Abstract:

Introduction
Bacterial biofilm (BF) is a highly structured bacterial community which is a complex aggregate of bacteria encased self-generated matrix of extracellular polymeric substance, metabolic waste and propagating microorganisms.1 The formation of BF is one of the essential strategies for bacteria survival, colonization and reproduction.2 BF enhances the adherence of bacterial cells to surfaces, and protects bacteria against antibiotics through a physical isolation, thus induced drug resistance. It was reported that the dose of some antibacterial medicines must be raised 1000 times against bacteria with BF in order to have the same inhibition effectiveness as against identical strains grown planktonically.3 Pseudomonas aeruginosa (P. aeruginosa), a well-known BF former, has become the major cause of hospital-associated infection and one of the most life-threatening bacteria.4,5 Treatment of P. aeruginosa infections has become a great challenge due to limited efficacy of current clinical antimicrobial agents.6,7 What was worse, with the isolation of the multidrug-resistant bacteria in clinic, eradication of P. aeruginosa has become increasingly difficult. The World Health Organization (WHO) has listed carbapenem-resistant P. aeruginosa as one of three bacterial species in which there is a critical need for the development of new antibiotics to treat infections.8 BF plays a vital role in P. aeruginosa colonization and invasion.9 Therefore, reagents that inhibit the BF formation and promoting degeneration with novel mechanisms, rather than directly sterilizing the bacteria, is a potential solution and one of the hotspots in current study.10,11
Quorum sensing (QS) system, through which bacteria synchronize their behavior by a series of signal molecules in a cell density dependent manner, is the most important regulator of BF formation. 12 P. aeruginosa has three QS systems, las, rhl and pqs, which are primarily involved with LasI/LasR, RhlI/RhlR and PQS (2-heptyl-3-hydroxy-4(1H)-quinolone), respectively (Figure 1). 13 The las system locates at the top of the QS signal cascade since the transcriptional regulator receptor protein LasR acts as the master regulator that controls the expression of Rhl and PQS pathways. 14,15 QS is caused by 10% gene of P. aeruginosa, while LasR regulates the expression of up to 5% of the genome directly or indirectly. 16,17 The signal molecular N-(3-oxododecanoyl)-L-homoserine lactone (3-oxo-C12-HSL) synthesized by LasI binds to LasR and the complex in turn promotes LasI expression. 18,19 Activation of LasR synchronously upregulates rhl gene system and accelerates the RhlI to synthesis N-butanoyl-l-homoserine lactone (C4-HSL), the activator of RhlR. The activated RhlR enhances gene expression and increases production of secondary metabolites. 18 LasR also promotes transcription of pqs and elevates the production of PQS. PQS binds to PqsR, one protein involved in iron acquisition and secretion, subsequentially reinforced the production of PQS through activating pqsA. 20 LasR also regulates the genes along with virulence factor production and BF formation. 21,22 The loss of LasR function leads to the reduction of virulence factors production, while evaluation of lasR mutant promoted the infection of P. aeruginosa. 16,23 What’s more, lots of research has revealed that modulating the LasR reduced the BF formation and mitigated resistant P. aeruginosa infections. 18,24,25 Therefore, impeding LasR binding to 3-oxo-C12-HSL may be an effective therapeutic strategy of P. aeruginosa infection.
 |
Figure 1 LasI/R-RHLI/R signaling system in P. aeruginosa. |
Traditional Chinese medicine (TCM) is a great treasure of drug discovery. However, it’s a huge challenge to quickly screen and identify the active ingredients from complex TCM extractions. The organic polymer monoliths, with relatively simple preparation processes, abundant monomer availability, and a wide application range of pH, have attracted the attention of chromatographers. 26 Affinity monolithic chromatography (AMC), which gathered the merits of monolithic column and nano HPLC, has been wildly used in active ingredients screening, isolation and recognition from complex mixture. 27 AMC have attracted a great deal of attention as they offer practical advantages, such as more rapid analysis, smaller sample volume requirements and the opportunity for a greater target molecule enrichment, by immobilizing binding agent or affinity ligand to the functional monolithic supports.28 Several enzymes immobilized monolithic columns have been successfully prepared and the effects in ligand fishing from TCM has been proven. 29–31 As the molecular structure of LasR has been reported, it’s possible to prepare LasR and the protein immobilized AMC.32,33
In this study, the LasR protein was firstly prepared in the E. coli system. Afterpurification and validation, LasR was immobilized to the surface of poly(GMA-co-PEGDA)-EDA monolithic column. The prepared poly(GMA-co-PEGDA)-EDA-LasR monolithic column was characterized and the affinity of protein to the ligand was also evaluated. What’s more, the ligand fishing effects of prepared column in TCM systems was also evaluated by the Scutellaria baicalensis Georgi extract. Baicalein, the screened potential LasR inhibitors, showed a good activity in anti P. aeruginosa in vitro.
Materials and Methods
Materials
A prototypic wild-type strain of P. aeruginosa (PAO1, purchased from ATCC) grown in LB medium (Thermo Fisher Scientific Inc., USA). E. coli 2568 was used as cloning hosts. DNA ligase, endonuclease EcoR I, Xho I and DNA marker were purchased from New England Biolabs, USA. Protein marker, fetal bovine serum, horseradish peroxidase (HRP) marker, bacterial genome extraction kit, plasmid extraction kit and PCR Kit were purchased from Gibco, USA. Isopropyl -β-D-thiogalactose glycosides (IPTG), N, N-methylene bisacrylamide, acrylamide, biphenyl diamine (DAB) and coomassie bright blue were purchased from Merck. Co., Germany. Other agents were chemically pure and purchased from Energy Chemical unless otherwise noted.
Preparation of LasR Protein
The preparation of LasR was similar to the published method with little modification.34 The lasR gene was amplified from PAO1 genomic DNA by using PCR with the following primers lasR-P1: 5′- ATA GAA TTC ATG GCC TTG GTT GAC GGT TT −3′, lasR-P2: 5′- CCG CTC GAG GAG AGT AAT AAG ACC CAA ATT AAC GGC −3′. The lasR genes were amplified with Pfu DNA polymerase. PCR reaction mixtures (50 μL total volume) consisted of 20 ng of PAO1 genomic DNA, 2 μM of each primer, 100 μM of each deoxynucleoside triphosphate (dNTP), 4 mm MgSO4, and 1 U of Pfu DNA polymerase in 1× reaction buffer. Mixtures were initially denatured at 94 °C for 5 min followed by 30 cycles of amplification. Each cycle consisted of denaturation at 94 °C for 30s, annealing at 55 °C for 30s, and extension at 72 °C for 1 min. The reaction was concluded with a final extension step at 72 °C for 10 min. The PCR-amplified lasR gene fragments were digested with EcoRI and XhoI restriction enzymes, then ligated into the similarly digested pET32a expression vector. The recombinant plasmids were transformed into E. coli 2568. The transformed bacteria were then plated on LB agar supplemented with (kanamycin/ampicillin, 100 μg/mL) and incubated at 37 °C overnight. A single colony was inoculated into 10 mL of LB medium supplemented with kanamycin and incubated overnight at 37 °C with shaking at 250 rpm. The overnight culture was then diluted 1:100 into 1000 mL of fresh LB medium and incubated at 37 °C with shaking at 250 rpm for an additional 3 h. Protein expression was induced by adding 0.5 mm IPTG, followed by incubation at 20 °C for 12 h. The cells were then harvested by centrifugation at 8,000 g for 10 min. The cell pellet was washed twice with 100 mL of phosphate-buffered saline (PBS, pH 7.4) and resuspended in 100 mL of ultrasonic buffer (50 mm PBS, pH 7.4, 150 mm NaCl). Cell lysis was performed by sonication (300 W, 30 cycles of 10 seconds on/10 seconds off). The lysate was centrifuged at 10,000 × g for 10 min at 4 °C. The pellet (containing inclusion bodies) was washed twice with cold PBS (4 °C) supplemented with 1% TritonX-100, followed by centrifugation at 10,000 × g for 10 minutes at 4 °C. Subsequently, the precipitate was solubilized in 8 M urea.
Protein Purification and Renaturation
The fully solubilized inclusion bodies, resulting in a transparent, gel-like solution, were loaded onto a nitrilotriacetic acid (NTA) column that pre-equilibrated using a gradient of NTAU-X buffer (50 mm PBS, pH 7.4, 150 mm NaCl, 8 M urea) and was then separated and purified by gradient NTAU-X buffer (50 mm PBS, pH 7.4, 150 mm NaCl, 8 M urea, X mM imidazole, where X ranged from 0, 10, 20, 50, 200 and 500 mm). Eluents containing the target protein were collected into a dialysis bag. The dialysis bag containing the denatured proteins was placed into renaturation buffer (50 mm PBS, 150 mm NaCl, 0.5 mm GSSG, 5 mm GSH, 400 mm arginine, pH 7.4) with decreasing urea concentrations (x M urea, where x ranged from 8, 4, 2 to 0 M). Dialysis was carried out at 4°C for 4–12 h at each urea concentration.
SDS-Polyacrylamide Gel Electrophoresis (SDSP-AGE) and Western blotting
LasR proteins were electrophoresed on SDS-polyacrylamide gels (with 10% acrylamide in the running gel). Electrophoretically separated proteins were blotted onto a Nylon Transfer membrane (Millipore) at 100 mA constant ampere for 2 h at 4 °C. Membranes were blocked with skim milk powder. A mouse anti-P. aeruginosa (diluted 1/100, Bio-Rad) and a goat anti-mouse immunoglobulin G (diluted 1/1000, Sigma-Aldrich, A0412) were employed as the primary and secondary antibodies, respectively.
Preparation of LasR Immobilized Monolithic Column
The preparation of LasR immobilized monolithic column is illustrated in Figure 2. Briefly, 10% of ethylenediamine (EDA) was pumped into a poly(GMA-co-PEGDA) column, which was prepared according to our reported method,29 and aminated for 4 h. The aminated poly(GMA-co-PEGDA)-EDA monolithic column was afforded after the residual EDA was washed with water to neutral. LasR protein was dissolved in 100 mm phosphate buffer (pH 6.0, 150 mm NaCl) to prepare a solution at concentration of 0.5 mg/mL. 2 mg EDC and 3 mg NHS were added into 400 μL of LasR solution and mixed for 10 min. The generated mixture was continuously peristaltic pumped into the poly(GMA-co-PEGDA)-EDA monolithic column. With the catalysis of EDC/NHS, the carboxyl group of LasR protein reacted with amino group of EDA moiety to generate an amide bond, thus the LasR protein was immobilized on the monolithic column surface.
 |
Figure 2 Preparation of poly(GMA-co-PEGDA)-EDA-LasR monolithic column. |
Characterization of Poly(GMA-Co-PEGDA)-EDA-LasR
The generated poly(GMA-co-PEGDA)-EDA-LasR monolithic column was characterized by Fourier transform infrared Spectroscopy (FT-IR) and scanning electron microscope (SEM). For FT-IR analysis, a bulk polymer (poly(GMA-co-PEGDA)) was firstly polymerized within a 1 mL glass vial, and then treated with EDA and LasR to give functionalized polymer, poly(GMA-co-PEGDA) and poly(GMA-co-PEGDA)-EDA-LasR, respectively. Those bulk polymers were cut into small pieces, Soxhlet extracted with methanol for 10 h, and then dried in a 60 °C oven overnight. The obtained polymers were finally crushed into powder, and taken for FT-IR analysis (Jasco FT/IR-480 plus, Jasco, Japan). The poly(GMA-co-PEGDA) column and the prepared poly(GMA-co-PEGDA)-EDA-LasR) column were cut into 1 cm length and dried in a 60 °C oven, and subsequently sputter-coated with gold for SEM analysis (Zeiss Ultra 55, Germany).
Evaluation of Ligand Affinity of Poly(GMA-Co-PEGDA)-EDA-LasR
N-(3-oxododecanoyl) -L-homoserine lactone (3-oxo-C12-HSL) is a natural ligand of LasR protein. It was used as a positive control to investigate the ligand affinity of immobilized LasR protein. Briefly, the 3-oxo-C12-HSL was ultrasonically dissolved in a solution of MeOH/Tris-HCl (20:80, v/v) and filtered. The LasR immobilized poly(GMA-co-PEGDA) monolithic column was installed in the nano-LC system.29 Loaded 20 nL of the filtrate of 3-oxo-C12-HSL and eluted with 25 mm Tris-HCl (pH 8.0) at the flow rate of 1.0 μL/min and detected at the wavelength of 200 nm. The affinity of immobilized LasR was evaluated by the chromatographic retention behavior of 3-oxo-C12-HSL. For the stability evaluation, the poly(GMA-co-PEGDA)-EDA-LasR monolithic column was stored at 4 °C for 24 h after prepared, and then, the affinity to 3-oxo-C12-HSL was evaluated as mentioned above.
Screening LasR Ligand from Scutellaria baicalensis Georgi
The prepared immobilized LasR poly(GMA-co-PEGDA) monolithic column was used for screening LasR inhibitors from TCM Scutellaria baicalensis Georgi. Firstly, 2.0 g of dried powder of Scutellaria baicalensis Georgi roots (purchased from Tong Ren Tang, Beijing) were extracted by ultrasonication with 100 mL ethanol/H2O (70:30, v/v) for 2 h. The ethanol was rotatory evaporated and 100 mL water was added in the residue. The mixture was extracted with ethyl acetate three times. The organic fractions were combined and evaporated to dryness. Finally, the residue was dissolved in water with 20% MeOH at a concentration of 3.0 mg/mL and then filtered through a 0.22 μm filter membrane and stored at 4 °C. The extract solution (200 μL, diluent with PBS to a concentration of 100 μg/mL) was loaded on the column and incubated for 30 min at 25 °C. The non-bound fraction was washed away using 400 μL PBS three times. Subsequently, the column was incubated with the elution buffer for 10 min to dissociate the LasR bonded compounds. The effluents were collected and analyzed by HPLC-ESI-Q-TOF-MS/MS (HPLC: Nexera-XRAD, Shimadzu, Kyoto, Japan; Q-TOF: X500R QTOF, AB SCIEX, Framingham, MA, USA). The chromatographic separation was achieved on a Venusil MP C18 column (250 mm × 4.6 mm, 5 μm, Agela). Gradient elution with solvents A (0.1 formic acid in water) and B (MeOH) was applied as follows: 0–15 min, 10–30% B; 15–50 min, 30–40% B; 50–75 min, 40–55% B; and 75–120 min, 55–75% B. The flow rate and column temperature were 1.0 mL/min and 25 °C, respectively, and the specific LasR ligands in Scutellaria baicalensis Georgi were identified. MS analysis was operated in negative mode. ESI source conditions were as follows: ionspray voltage, −4.5 kV; temperature, 550 oC; the pressure of gas 1, 50 psi, gas 2, 60 psi; pressure of curtain gas, 30 psi; collision energy, −35 (with a collision energy spread of ± 15). The mass spectra were recorded in the range of m/z 200–800.
Evaluation of Screened Compound Against P. aeruginosa in vitro
The evaluation of screened compound against P. aeruginosa in vitro was referred to our published work.35 Briefly, for the minimum inhibition concentration (MIC), P. aeruginosa was diluted with MH broth to 106 CFU/mL. 100 μL of Baicalein at the concentrations of 0.5–128 μg/mL was added into 96-well plates. Then 100 μL of prepared bacterial suspension was added into the 96-well plate. The total mixture of 200 μL was cultured at 37°C overnight, and the baicalein concentration that completely inhibited bacteria growth was recorded as MIC value. Then 100 μL of culture mixture was pipetted into MH AGAR medium and cultured at 37 °C overnight. The lowest concentration without bacterial growth in the culture plate was recorded as the MBC.
Molecular Docking of Baicalein and LasR
SurflexDock in SYBYL 8.1 was employed to study the molecular docking. The crystal structure of LasR was retrieved from the RCSB Protein Data Bank (PDB entry: 3IX3). All ligands and water molecules were removed. The polar hydrogen atoms and Gasteiger charges were added. The active site was defined with a radius of 2.5 Å around the ligand present in the crystal structure and a grid was generated at the center of gravity of the active site for docking. Studied compound was docked into the catalytic pocket of the protein.
The molecular dynamics (MD) simulations were conducted in GROMACS 2022.2. The protein structure is obtained from the Protein Data Bank (PDB code: 3IX3). The protein is prepared by removing crystallographic water molecules, adding missing hydrogens, and parameterizing with the AMBER14SB force field. The ligands are optimized using quantum mechanics at HF/6-31G* level to calculate RESP charges, and the General Amber Force Field (GAFF) is assigned with ACPYPE script. The ligand-protein complex is assembled and solvated in a TIP3P periodic box with 1.2 nm buffer space. Counter ions are added to neutralize the system and mimic physiological ionic strength (0.15 M NaCl). After solvation, energy minimization is performed using the steepest descent algorithm to relieve steric clashes. Equilibration is conducted in NVT (constant volume and temperature) and NPT (constant pressure and temperature) to stabilize temperature and pressure, with positional restraints on the protein and ligand heavy atoms. For each system the production run is carried out without any restraints for 100 ns, with a 2 fs time step, employing the LINCS algorithm for bond constraints, PME for long-range electrostatics.
Results and Discussion
The Expression and Purification of LasR in Recombinational E. coli 2568
Firstly, to determine whether the recombinant pET32a was successfully transfected into E. coli 2568, the total proteins of the bacterium pro- and post-induction were firstly analyzed with SDS-PAGE. The result are shown in Figure 3A. After being induced with 0.5 mm IPTG at 20 °C for 12 h, there is an obviously inducible protein band at 44 kD, which was not observed in the uninduced bacterium (lane 1). Since the molecular weight of pET32a was 44 kD, the LasR was successfully introduced and expressed into E. coli 2568. After being sonicated and centrifuged, the total proteins in pellet and supernatant were analyzed. As shown in Figure 3B, LasR protein was mainly expressed in the pellet of lysate centrifugation. Then, the generated pellet was dissolved in 8 M urea solution and purified on an NTA column. As illustrated in Figure 3C, the gradient NTAU elution with imidazole ranged from 10 to 500 mm was effective in purifying LasR protein, and the purity was increased with the concentration of imidazole. The purified LasR protein was dialyzed in the gradient renaturation solution and the soluble LasR was afforded. LasR is known as a master positive QS regulator in P. aeruginosa.36 Mutations in lasR gene, have long been detected and are common in clinical P. aeruginosa isolates, and mostly associated with worse disease progressions.37 The antibacterial agents targeted in LasR are urgent. Genetic engineering is recognized as a favored approach for the large-scale synthesis of peptides/proteins,38 which will facilitate the discovery of LasR inhibitors.
 |
Figure 3 The production and purification of LasR proteins. (A) Whole-cell protein profiles of E. coli 2568 with pET32a-LasR. Lane 1: E. coli 2568 with pET32a-LasR before IPTG induction; Lane 2: E. coli 2568 with pET32a-LasR inducted with 0.5 mm IPTG; M: marker protein. (B) The total proteins in the pellet and supernatant after sonication. Lane 1: E. coli 2568 with pET32a-LasR before IPTG induction; Lane 2: E. coli 2568 with pET32a-LasR induced with 0.5mM IPTG; Lane 3: the supernatant of centrifuged lysates solution; Lane 4: the pellet of centrifuged lysates solution; M: marker protein. (C) The LasR protein purified by NTA column through gradient NTAU elution. M: marker protein; Lane 1: the solution of pellet dissolved in 8 M urea; Lane 2: the flow-through solution; Lane 3: NTAU-10 (10 mm imidazole) elution; Lane 4: NTAU-20 (20 mm imidazole) elution; Lane 5: NTAU-50 (50 mm imidazole) elution; Lane 6: NTAU-200 (200 mm imidazole) elution; Lane 7: NTAU-500 (500 mm imidazole) elution. |
Characterization of Poly(GMA-Co-PEGDA)-EDA-LasR
The FT-IR was widely used in monitoring the reaction of polymeric compounds since the absorption of the characteristic infrared group changes after reacted, therefore, the immobilization of LasR to the poly(GMA-co-PEGDA)-EDA was firstly characterized by FT-IR. As illustrated in Figure 4A, a strong absorption peak was obtained at 1734 cm−1, which was attributed to the stretching of carbonyl group of PEGDA, indicates that the poly(GMA-co-PEGDA) monolithic column was successfully prepared.29 Characteristic bands for LasR include those of amide I and amide II, which originate from the amide bonds linking the amino acids.29 There is a strong absorption peak at 1658 cm−1 in Figure 4C, which was associated with amide I band, resulting from the stretching vibrations of the C=O bonds of the amide groups in LasR. After LasR immobilization, the bands resulting from stretching vibrations of the C=O bonds appeared (1663 cm−1, Figure 4B), indicating that LasR was successfully immobilized on the poly(GMA-co-PEGDA)-EDA monolith.
 |
Figure 4 The IR spectrum of poly(GMA-co-PEGDA)-EDA, LasR and poly(GMA-co-PEGDA)-EDA-LasR. |
The internal morphology of poly(GMA-co-PEGDA) monolithic column before and after LasR immobilization was observed by Ultra-55 field emission SEM and the results are shown in Figure 5. The poly(GMA-co-PEGDA) monolithic column (Figure 5A) was polymerized well internal, and the polymers were tightly attached to the inner wall of the capillary tube with uniform distribution of large pores. The inner surface of poly(GMA-co-PEGDA)-EDA-LasR monolithic column prepared (Figure 5B) became rough and the pore size was significantly smaller. However, the LasR immobilized columns still retained the morphological structure of alternating distribution of large and small holes. The changes of polymer morphology and pore size indicate that the immobilization of LasR induced the physical and chemical changes of the column.
 |
Figure 5 SEM images of the cross-sections of poly(GMA-co-PEGDA) monolith (A) and poly(GMA-co-PEGDA)-EDA-LasR monolith (B). |
Evaluation of Ligand Affinity for LasR Immobilized Monolithic Column
3-oxo-C12-HSL is the nature ligand of LasR, therefore, it was used to explore the ligand affinity of immobilized LasR. Poly(GMA-co-PEGDA) monolithic column has no retention effect on 3-oxo-C12-HSL (Figure 6A), and the modification with ethylenediamine in the column did not change the retention characteristics significantly (Figure 6B). However, after being immobilized with LasR protein, the retention time of 3-oxo-C12-HSL in the monolithic column was significantly lengthened (Figure 6C). These data indicated that the LasR protein was successfully immobilized to the monolithic column, and more importantly, the immobilized LasR reserved the ligand affinity and enzymatic activity.
 |
Figure 6 The LasR immobilized monolithic column reacted with the natural ligand. Chromatograms of 3-oxo-C12-HSL acted on poly(GMA-co-PEGDA) monolith (A), poly(GMA-co-PEGDA)-EDA monolith (B) and poly(GMA-co-PEGDA)-EDA-LasR monolith (C). |
LasR protein contains two ligand binding domains, the N-terminal ligand binding domain and the C-terminal DNA binding domain, in which the N-terminal ligand binding domain is the binding site of LasR protein and 3-oxo-C12-HSL.39,40 Our unpublished data also confirmed that the immobilization of LasR at the residual amino group eliminated the affinity to 3-oxo-C12-HSL. The results showed that the carboxyl immobilized LasR protein had certain ligand affinity. It can be seen that the ligand binding function of the N-terminal ligand domain of LasR protein is not affected when the carboxyl group of LasR protein is used as the immobilization site.
The structure stability of the protein is essential for the activity. Thus, the stability of immobilized LasR monolithic column was also investigated. As shown in Figure 7B, the retention time of 3-oxo-C12-HSL was significantly shortened to 2 min after stored at 4 °C for 24 h compared with 10 min of newly immobilized column (Figure 7A). The results indicated that LasR protein monolith had good activity only when newly prepared, so, only the newly prepared LasR immobilized columns were used for the further studies.
 |
Figure 7 Stability of poly(GMA-co-PEGDA)-EDA-LasR monolith column. The retention time of 3-oxo-C12-HSL on newly prepared column (A) and column stored at 4 °C for 24 h (B). |
Screening LasR Ligands from Scutellaria baicalensis Georgi Extract
P. aeruginosa infections are an important cause of nosocomial and ventilator-associated pneumonia characterized by high prevalence and fatality rates.41,42 Scutellaria baicalensis Georgi is a medicinal herb with a rich history of applications in traditional Chinese medicine with lots of pharmacological activities.43 It was widely used as an important ingredient of prescription in TCMs to treat lung inflammation related diseases. As an antibiotic alternative, Scutellaria baicalensis Georgi also showed a great potential against Salmonella, Streptococcus mutans and other bacteria.44,45 The most important ingredients, baicalein and baicalin, were also widely reported effective in against BF of P. aeruginosa with unclear mechanisms.46,47 Thus, the prepared poly(GMA-co-PEGDA)-EDA-LasR monolithic column was applied in the screening of LasR from the extraction of Scutellaria baicalensis. Comparing the chromatograms of compounds bound to poly(GMA-co-PEGDA)-EDA-LasR (Figure 8B) and poly(GMA-co-PEGDA) (Figure 8A), the peak (baicalein) was found to be strong affinity to LasR. Its structure was confirmed by comparing with the MS data of standard compound. The binding degree of baicalein was measured as 0.98% after eliminating nonspecific binding. The anti P. aeruginosa activity of baicalein was further evaluated in vitro, and the MIC of baicalein is 64 µg/mL while the MBC is 128 µg/mL, respectively. These data demonstrated the LasR inhibitor baicalein, which was fished by poly(GMA-co-PEGDA)-EDA-LasR monolithic column, was potential in controlling P. aeruginosa.
 |
Figure 8 Specificity evaluation of the LasR immobilized column for bioactivity screening of Scutellaria baicalensis Georgi extract. (A) HPLC chromatogram of Scutellaria baicalensis extract. (B) HPLC chromatogram of compounds bound to poly(GMA-co-PEGDA)-EDA-LasR (the blue line) and contrast poly(GMA-co-PEGDA) (the black line). |
Molecular Docking of Baicalein to LasR
In order to understand the interaction between the compound and protein, molecular docking of baicalein into the active site of LasR (PDB code: 3IX3), was constructed using Sybyl 8.1.1. The docking grid was generated according to the centroid of in-bound ligand of 3IX3, coordinates of the center of the grid are (10.65, 4.5, 20.96). As illustrated in Figure 9A, baicalein clearly showed the good complementarity within the LasR pocket. The aromatic group extensively interacted with the pocket through hydrogen bond interactions with the backbone of Trp 60, Arg 61, and Asp 73. And the side chain of baicalein formed hydrogen bonds with Thr 75 in the active pocket of LasR. And the 3-oxo-C12-HSL binds with LasR through hydrogen bonds with Tyr 56, Tpr 60, Asp 73 and Ser 129 (Figure 9B). In addition, the docking score/binding energy of baicalein and 3-oxo-C12-HSL were also evaluated (Table 1).
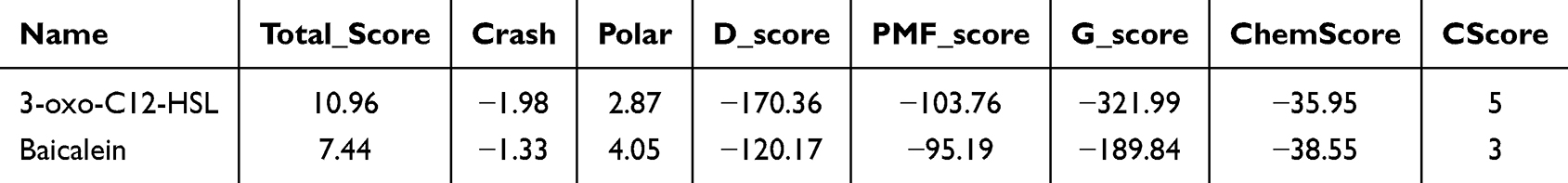 |
Table 1 The Docking Scores of Compounds to LasR |
 |
Figure 9 Docking conformation of baicalein and 3-oxo-C12-HSL in LasR. (A) The docked binding modes of baicalein within the LasR pocket. (B) The docked binding modes of 3-oxo-C12-HSL within the LasR pocket (PDB code: 3IX3). |
We further validated the docking results of 3-oxo-C12-HSL and baicalein with LasR using molecular dynamic simulations. Both complex structures underwent 100 ns of conventional simulations. The RMSD results showed that both docking results reached stability around 20 ns with the maximum RMSD of ~1.8 Å (Figure 10). Compared to the 3-oxo-C12-HSL molecule (which mainly interacts with Asp 73 and Trp 60, Figure 10A), the interaction between baicalein and LasR primarily occurs through hydrogen bonds and water bridges with Arg 61 and Thr 75, with the contact percentage exceeding 100% in the simulations (Figure 10B). This indicates that these sites exhibit more than a single interaction in the simulations. This finding could not be observed in the docking study, as the solvent water was not included in the docking process. Additionally, from the molecular Rg (Radius of Gyration) and RMSF in the simulation, it is evident that the torsion profile of baicalein’s relatively rigid structure favors more to that of the flexible 3-oxo-C12-HSL molecule.
 |
Figure 10 Molecular dynamic simulation results of baicalein and 3-oxo-C12-HSL with LasR. |
Baicalein was widely reported to inhibit QS and therefore suppress the formation of BF in different bacterium.48–50 However, recently studies revealed the combination of low concentration of baicalein synergized the effects of antibiotics against the BF formation of P. aeruginosa PAO1.46,51 Sub-MIC levels of baicalein also effective in suppressing the production of virulence factors, detaching the biofilm and reducing adhesion of P. aeruginosa PAO1 through downregulating lasR by 80.8%.52 The residues of Tyr 47, Ala 50, Tyr 56, Trp 60, Tyr 64, Asp 73, Val 76 and Ser 129 constituted the catalytic site of LasR. In a published work, baicalein binds to LasR by forming 4 hydrogen bonds with the residues Trp 60, Thr 75, Thr 115 and Ser 129.53 A recent study revealed that baicalein mostly reacted with Arg 61 through hydrogen bond.25 The interactions of baicalein with Trp 60, Arg 61, Asp 73 and Thr 75, in this study, was in accordance with the reported results. The molecular dynamic result of LasR-Baicalein showed that the docking completed stability after 15 ns with the maximum RMSD of ~2.2 Å, which was also corresponded to our result.54 The experimental confirmation is important for validating the baicalein binding to LasR. The effects of baicalein on the virulence factors production and BF formation was reported and validated by different groups, although we did not evaluate in this study.25,52,54,55
Conclusions
In this research, a recombinant LasR was prepared in the E. coli system. Through a chemical reaction with the amino group of the poly(GMA-co-PEGDA)-EDA, LasR was immobilized to the surface of the monolithic column. The generated monolithic column was characterized by FT-IR and SEM and the results indicated the successful immobilization of LasR. The prepared column reversed the activity of LasR for 24 h at 4 °C. It was used for fishing the LasR inhibitors from Scutellaria baicalensis Georgi extract. Baicalein was screened out as a LasR inhibitor. The proposed LasR immobilized monolithic column is a viable strategy and showed potential in screening LasR inhibitors. It can be considered as a possible alternative to traditional methods for screening reagents against P. aeruginosa. However, the rapid denaturation and inactivation (< 24 h) of immobilization LasR requires timely application. The durability and repeatability of the column also needs further investigation.
Funding
This work was supported by the National Natural Science Foundation of China (Grant Nos.: 82373835, 82173781, 82404579, and 82304313), Regional Joint Fund Project of Guangdong Basic and Applied Basic Research Fund (Grant Nos.: 2023A1515110417 and 2023A1515140131), Medical Scientific Research Foundation of Guangdong Province of China (A2022061), Scientific Research Platforms and Projects of Guangdong Universities (Grant Nos.: 2022ZDZX2056 and 2024KCXTD045), the Engineering Technological Center of Guangdong Province (Grant No.: 204266924217), and the Engineering Technological Center of Foshan City (Grant No.: 2017GA00025).
Disclosure
The authors confirm that there is no conflict of interest related to the manuscript.
References
1. Sara ER, Badr E, Matencio A, et al. Plant-derived bioactive compounds for the inhibition of biofilm formation: a comprehensive review. Environ Sci Pollut Res Int. 2024;31(24):34859–34880. doi:10.1007/s11356-024-33532-2
2. Thi MT, Wibowo D, Rehm HA. Pseudomonas aeruginosa biofilms. Int J mol Sci. 2020;21(22):8671. doi:10.3390/ijms21228671
3. Lewis K. Riddle of biofilm resistance. Antimicrob Agents Chemother. 2001;45(4):999–1007. doi:10.1128/AAC.45.4.999-1007.2001
4. Lee K, Yoon SS. Pseudomonas aeruginosa biofilm, a programmed bacterial life for fitness. J Microbiol Biotechnol. 2017;27(6):1053–1064. doi:10.4014/jmb.1611.11056
5. Pang Z, Raudonis R, Glick BR, Lin TJ, Cheng Z. Antibiotic resistance in Pseudomonas aeruginosa: mechanisms and alternative therapeutic strategies. Biotechnol Adv. 2019;37(1):177–192. doi:10.1016/j.biotechadv.2018.11.013
6. Ye J, Kan CH, Yang X, Ma C. Inhibition of bacterial RNA polymerase function and protein-protein interactions: a promising approach for next-generation antibacterial therapeutics. RSC Med Chem. 2024;15(5):1471–1487. doi:10.1039/D3MD00690E
7. Qin S, Xiao W, Zhou C, et al. Pseudomonas aeruginosa: pathogenesis, virulence factors, antibiotic resistance, interaction with host, technology advances and emerging therapeutics. Signal Transduct Target Ther. 2022;7(1):199. doi:10.1038/s41392-022-01056-1
8. World Health Organization. Global priority list of antibiotic-resistant bacteria to guide research, discovery, and development of new antibiotics. World Health Organization. 2017;1–7.
9. Irene JM, Maite SM, Siobhán M. Pseudomonas aeruginosa: an audacious pathogen with an adaptable arsenal of virulence factors. Int J mol Sci. 2021;22(6):3128. doi:10.3390/ijms22063128
10. Roy R, Tiwari M, Donelli G, Tiwari V. Strategies for combating bacterial biofilms: a focus on anti-biofilm agents and their mechanisms of action. Virulence. 2018;9(1):522–554. doi:10.1080/21505594.2017.1313372
11. Yu Q, Wang C, Zhang X, Chen H, Wu MX, Lu M. Photochemical strategies toward precision targeting against multidrug-resistant bacterial infections. ACS nano. 2024;18(22):14085–14122. doi:10.1021/acsnano.3c12714
12. Liu Z, Zhang LH, Wang JC, et al. Virtual screening and biological evaluation of anti-biofilm agents targeting LuxS in the quorum sensing system. Nat Prod Commun. 2021;16(6):1–10.
13. Fan H, Dong Y, Wu D, Bowler MW, Zhang L, Song H. QsIA disrupts LasR dimerization in antiactivation of bacterial quorum sensing. Proc Natl Acad Sci U S A. 2013;110(51):20765–20770. doi:10.1073/pnas.1314415110
14. Xuan G, Lv C, Xu H, et al. Sulfane sulfur regulates LasR-mediated quorum sensing and virulence in Pseudomonas aeruginosa PAO1. Antioxidants. 2021;10(9):1498. doi:10.3390/antiox10091498
15. Waters CM, Goldberg JB. Pseudomonas aeruginosa in cystic fibrosis: a chronic cheater. Proc Natl Acad Sci U S A. 2019;116(14):6525–6527. doi:10.1073/pnas.1902734116
16. Hennemann LC, Nguyen D. LasR-regulated proteases in acute vs. chronic lung infection: a double-edged sword. Microb Cell. 2021;8(7):161–163. doi:10.15698/mic2021.07.755
17. Williams P, Cámara M. Quorum sensing and environmental adaptation in Pseudomonas aeruginosa: a tale of regulatory networks and multifunctional signal molecules. Curr Opin Microbiol. 2009;12(2):182–191. doi:10.1016/j.mib.2009.01.005
18. Datta S, Singh V, Nag S, Roy DN. Carvacrol, a monoterpenoid, binds quorum sensing proteins (LasI and LasR) and swarming motility protein BswR of Pseudomonas aeruginosa, resulting in loss of pathogenicity: an in silico approach. Can J Microbiol. 2025;71:1–15. doi:10.1139/cjm-2024-0155
19. Wang J, Yang JY, Durairaj P, et al. Discovery and evaluation of 3-(2-isocyanobenzyl)-1H-indole derivatives as potential quorum sensing inhibitors for the control of Pseudomonas aeruginosa infections in vitro. RSC Med Chem. 2024;15(9):3256–3271. doi:10.1039/D4MD00354C
20. Simanek KA, Paczkowski JE. Resistance is not futile: the role of quorum sensing plasticity in Pseudomonas aeruginosa infections and its link to intrinsic mechanisms of antibiotic resistance. Microorganisms. 2022;10(6):1247. doi:10.3390/microorganisms10061247
21. Thakur M, Khushboo, Kumar Y, Yadav V, Pramanik A, Dubey KK. Understanding resistance acquisition by Pseudomonas aeruginosa and possible pharmacological approaches in palliating its pathogenesis. Biochem Pharmacol. 2023;215:115689. doi:10.1016/j.bcp.2023.115689
22. Suneby EG, Herndon LR, Schneider TL. Pseudomonas aeruginosa LasR·DNA binding is directly inhibited by quorum sensing antagonists. ACS Infect Dis. 2017;3(3):183–189. doi:10.1021/acsinfecdis.6b00163
23. Zhao K, Yang X, Zeng Q, et al. Evolution of lasR mutants in polymorphic Pseudomonas aeruginosa populations facilitates chronic infection of the lung. Nat Commun. 2023;14(1):5976. doi:10.1038/s41467-023-41704-w
24. Xu Y, Tong X, Sun P, Bi L, Lin K. Virtual screening and biological evaluation of biofilm inhibitors on dual targets in quorum sensing system. Future Med Chem. 2017;9(17):1983–1994. doi:10.4155/fmc-2017-0127
25. Lima EMF, de Almeida FA, Pinto UM. Exploring the antivirulence potential of phenolic compounds to inhibit quorum sensing in Pseudomonas aeruginosa. World J Microbiol Biotechnol. 2025;41(2):32. doi:10.1007/s11274-025-04255-4
26. Ding X, Yang J, Dong Y. Advancements in the preparation of high-performance liquid chromatographic organic polymer monoliths for the separation of small-molecule drugs. J Pharm Anal. 2018;8(2):75–85. doi:10.1016/j.jpha.2018.02.001
27. Yang B, Yang H, Liang J, et al. A review on the screening methods for the discovery of natural antimicrobial peptides. J Pharm Anal. 2024;15:101046. doi:10.1016/j.jpha.2024.101046
28. Poddar S, Sharmeen S, Hage DS. Affinity monolith chromatography: a review of general principles and recent developments. Electrophoresis. 2021;42(24):2577–2598. doi:10.1002/elps.202100163
29. Lin H, Zhang C, Lin Y, et al. A strategy for screening trypsin inhibitors from traditional Chinese medicine based on a monolithic capillary immobilized enzyme reactor coupled with offline liquid chromatography and mass spectrometry. J Sep Sci. 2019;42(11):1980–1989. doi:10.1002/jssc.201900169
30. Wang L, Zhao Y, Zhang Y, et al. Online screening of acetylcholinesterase inhibitors in natural products using monolith-based immobilized capillary enzyme reactors combined with liquid chromatography-mass spectrometry. J Chromatogr A. 2018;1563:135–143. doi:10.1016/j.chroma.2018.05.069
31. Wang J, Huang X, Mei J, et al. Screening of trypsin inhibitors in Cotinus coggygria Scop. extract using at-line nanofractionation coupled with semi-preparative reverse-phase liquid chromatography. J Chromatogr A. 2023;1691:463817. doi:10.1016/j.chroma.2023.463817
32. Kafle P, Amoh AN, Reaves JM, et al. Molecular insights into the impact of oxidative stress on the quorum-sensing regulator protein LasR. J Biol Chem. 2016;291(22):11776–11786. doi:10.1074/jbc.M116.719351
33. Dalal A, Kushwaha T, Choudhir G, et al. Computational investigations on the potential role of hygrophorones as quorum sensing inhibitors against LasR protein of Pseudomonas aeruginosa. J Biomol Struct Dyn. 2023;41(6):2249–2259. doi:10.1080/07391102.2022.2029570
34. Ou XK, Li W. Construction, identification, and expression of a recombinant Ef-LasR vaccine against Pseudomonas aeruginosa. J Pathogen Biol. 2021;16(06):630–633+638.
35. Yang H, Ma R, Chen J, et al. Discovery of melittin as triple-action agent: broad-spectrum antibacterial, anti-biofilm, and potential anti-quorum sensing activities. Molecules. 2024;29(3):558. doi:10.3390/molecules29030558
36. Wang Y, Gao L, Rao X, et al. Characterization of lasR-deficient clinical isolates of Pseudomonas aeruginosa. Sci Rep. 2018;8(1):13344. doi:10.1038/s41598-018-30813-y
37. Qiu HF, Li YH, Dai WJ. Codon-usage frequency mediated SNPs selection in lasR gene of cystic fibrosis Pseudomonas aeruginosa isolates. Microbiol Res. 2019;223-225:137–143. doi:10.1016/j.micres.2019.05.002
38. Saroha P, Patil RS, Rathore AS. Recent advancements in soluble expression of recombinant antibody fragments in microbial host systems. Prep Biochem Biotechnol. 2024;1–10.
39. Chowdhury N, Bagchi A. Molecular insight into the activity of LasR protein from Pseudomonas aeruginosa in the regulation of virulence gene expression by this organism. Gene. 2016;580(1):80–87. doi:10.1016/j.gene.2015.12.067
40. Grabski H, Hunanyan L, Tiratsuyan S, Vardapetyan H. Interaction of N-3-oxododecanoyl homoserine lactone with transcriptional regulator LasR of Pseudomonas aeruginosa: insights from molecular docking and dynamics simulations. bioRxiv. 2017;121681.
41. Maurice NM, Bedi B, Sadikot RT. Pseudomonas aeruginosa biofilms: host response and clinical implications in lung infections. Am J Respir Cell mol Biol. 2018;58(4):428–439. doi:10.1165/rcmb.2017-0321TR
42. Hermann DR, Timsit JF. Management strategies for severe Pseudomonas aeruginosa infections. Curr Opin Infect Dis. 2023;36(6):585–595. doi:10.1097/QCO.0000000000000981
43. Guo F, Li C, Dou J, et al. Research progress on pharmacological properties and application of probiotics in the fermentation of Scutellaria baicalensis Georgi. Front Nutr. 2024;11:1407182. doi:10.3389/fnut.2024.1407182
44. Jiang H, Bai Z, Xu Z, et al. Antimicrobial mechanism of semi-bionic extracts of three traditional medicinal plants-Rheum palmatum L. Scutellaria baicalensis Georgi, and Houttuynia cordata Thunb-That can be used as antibiotic alternatives. Front Vet Sci. 2022;9:1083223. doi:10.3389/fvets.2022.1083223
45. Zhao T, Tang H, Xie L, et al. Scutellaria baicalensis Georgi. (Lamiaceae): a review of its traditional uses, botany, phytochemistry, pharmacology and toxicology. J Pharm Pharmacol. 2019;71(9):1353–1369. doi:10.1111/jphp.13129
46. Kart D, Reçber T, Nemutlu E, Sagiroglu M. Sub-inhibitory concentrations of ciprofloxacin alone and combinations with plant-derived compounds against P. aeruginosa biofilms and their effects on the metabolomic profile of P. aeruginosa biofilms. Antibiotics. 2021;10(4):414. doi:10.3390/antibiotics10040414
47. Luo J, Dong B, Wang K, et al. Baicalin inhibits biofilm formation, attenuates the quorum sensing-controlled virulence and enhances Pseudomonas aeruginosa clearance in a mouse peritoneal implant infection model. PLoS One. 2017;12(4):e0176883. doi:10.1371/journal.pone.0176883
48. Qin X, Wu Y, Zhao Y, et al. Revealing active constituents within traditional Chinese Medicine used for treating bacterial pneumonia, with emphasis on the mechanism of baicalein against multi-drug resistant Klebsiella pneumoniae. J Ethnopharmacol. 2024;321:117488. doi:10.1016/j.jep.2023.117488
49. Mao Y, Liu P, Chen H, Wang Y, Li C, Wang Q. Baicalein inhibits the Staphylococcus aureus biofilm and the LuxS/AI-2 system in vitro. Infect Drug Resist. 2023;16:2861–2882. doi:10.2147/IDR.S406243
50. Vijayakumar A, Sarveswari HB, Vasudevan S, Shanmugam K, Solomon AP, Neelakantan P. Baicalein inhibits Streptococcus mutans biofilms and dental caries-related virulence phenotypes. Antibiotics. 2021;10(2):215. doi:10.3390/antibiotics10020215
51. Lima EMF, Bueris V, Germano LG, Sircili MP, Pinto UM. Synergistic effect of the combination of phenolic compounds and tobramycin on the inhibition of Pseudomonas aeruginosa biofilm. Microb Pathog. 2024;197:107079. doi:10.1016/j.micpath.2024.107079
52. Luo J, Kong JL, Dong BY, et al. Baicalein attenuates the quorum sensing-controlled virulence factors of Pseudomonas aeruginosa and relieves the inflammatory response in P. aeruginosa-infected macrophages by downregulating the MAPK and NFκB signal-transduction pathways. Drug Des Devel Ther. 2016;10:183–203. doi:10.2147/DDDT.S97221
53. Rajkumari J, Borkotoky S, Murali A, Suchiang K, Mohanty SK, Busi S. Attenuation of quorum sensing controlled virulence factors and biofilm formation in Pseudomonas aeruginosa by pentacyclic triterpenes, betulin and betulinic acid. Microb Pathog. 2018;118:48–60. doi:10.1016/j.micpath.2018.03.012
54. Rajkumari J, Borkotoky S, Murali A, Suchiang K, Mohanty SK, Busi S. Cinnamic acid attenuates quorum sensing associated virulence factors and biofilm formation in Pseudomonas aeruginosa PAO1. Biotechnol Lett. 2018;40(7):1087–1100. doi:10.1007/s10529-018-2557-9
55. Kolouchová I, Maťátková O, Paldrychová M, et al. Resveratrol, pterostilbene, and baicalein: plant-derived anti-biofilm agents. Folia Microbiol. 2018;63(3):261–272. doi:10.1007/s12223-017-0549-0
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms.php
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms.php
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Prevalence of Quorum Sensing and Virulence Factor Genes Among Pseudomonas aeruginosa Isolated from Patients Suffering from Different Infections and Their Association with Antimicrobial Resistance
Ghanem SM, Abd El-Baky RM, Abourehab MAS, Fadl GFM, Gamil NGFM
Infection and Drug Resistance 2023, 16:2371-2385
Published Date: 21 April 2023