")
Back to Journals » International Journal of Nanomedicine » Volume 19
Engineering Probiotics for Diabetes Management: Advances, Challenges, and Future Directions in Translational Microbiology
Authors Zhang S, Ma J, Ma Y , Yi J , Wang B , Wang H, Yang Q, Zhang K, Yan X, Sun D , You J
Received 22 August 2024
Accepted for publication 8 October 2024
Published 28 October 2024 Volume 2024:19 Pages 10917—10940
DOI https://doi.org/10.2147/IJN.S492651
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Kamakhya Misra
Shenghao Zhang,1,* Jiahui Ma,1,* Yilei Ma,1,* Jia Yi,1 Beier Wang,2 Hanbing Wang,3 Qinsi Yang,4 Kun Zhang,5 Xiaoqing Yan,6 Da Sun,1,7 Jinfeng You8
1Institute of Life Sciences & Biomedical Collaborative Innovation Center of Zhejiang Province, Wenzhou University, Wenzhou, 325035, People’s Republic of China; 2Department of Hepatobiliary-Pancreatic Surgery, The Second Affiliated Hospital of Wenzhou Medical University, Wenzhou, 325000, People’s Republic of China; 3Department of Biotechnology, The University of Hong Kong, Hong Kong SAR, 999077, People’s Republic of China; 4Wenzhou Institute, University of Chinese Academy of Sciences, Wenzhou, 325000, People’s Republic of China; 5Chongqing Municipality Clinical Research Center for Endocrinology and Metabolic Diseases, Chongqing University Three Gorges Hospital, Chongqing, 404000, People’s Republic of China; 6The Chinese-American Research Institute for Diabetic Complications, School of Pharmaceutical Sciences, Wenzhou Medical University, Wenzhou, 325035, People’s Republic of China; 7Department of Endocrinology, Yiwu Central Hospital, The Affiliated Yiwu hospital of Wenzhou Medical University, Yiwu, 322000, People’s Republic of China; 8Department of Obstetrics, The Quzhou Affiliated Hospital of Wenzhou Medical University, Quzhou People’s Hospital, Quzhou, 324000, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Jinfeng You; Da Sun, Email [email protected]; [email protected]
Background: Diabetes Mellitus (DM) is a substantial health concern worldwide, and its incidence is progressively escalating. Conventional pharmacological interventions frequently entail undesirable side effects, and while probiotics offer benefits, they are hindered by constraints such as diminished stability and effectiveness within the gastrointestinal milieu. Given these complications, the advent of bioengineered probiotics is a promising alternative for DM management.
Aim of Review: The objective of this review is to provide an exhaustive synthesis of the most recent studies on the use of engineered probiotics in the management of DM. This study aimed to clarify the mechanisms through which these probiotics function, evaluate their clinical effectiveness, and enhance public awareness of their prospective advantages in the treatment of DM.
Key Scientific Concepts of Review: Scholarly critiques have explored diverse methodologies of probiotic engineering, including physical alteration, bioenrichment, and genetic manipulation. These techniques augment the therapeutic potency of probiotics by ameliorating gut microbiota, fortifying the intestinal barrier, modulating metabolic pathways, and regulating immune responses. Such advancements have established engineered probiotics as a credible therapeutic strategy for DM, potentially providing enhanced results compared to conventional treatments.
Keywords: engineered probiotics, diabetes mellitus, gut microbiota, genetic engineering, therapeutic efficacy
Graphical Abstract:
Introduction
Diabetes mellitus (DM) is a chronic disease characterized by elevated blood glucose levels owing to inadequate insulin secretion or ineffective insulin action.1 Globally, DM is a major public health concern, with significant impacts on individuals, health systems, and national economies owing to its high prevalence and numerous complications.2,3 According to the International Diabetes Federation, as of 2021, 537 million adults are affected by DM, with three-quarters residing in low - and middle-income countries, leading to an estimated health expenditure of at least US$ 966.4 This burden is projected to increase, with 783.2 million people expected to be affected by 2045.5
There are two primary types of DM: type 1 diabetes mellitus (T1DM) and type 2 diabetes mellitus (T2DM).1 T1DM, which mainly affects children and adolescents, is caused by autoimmune destruction of islet β cells, leading to insufficient insulin secretion.6 Daily insulin injections are the standard treatment. In contrast, T2DM, which can occur at any age, is becoming increasingly prevalent in children and adolescents.7 It is characterized by insulin resistance (IR) and varying degrees of insulin deficiency. T2DM management involves lifestyle interventions and sometimes medications.8 However, the increasing number of DM cases underscores the inadequacies of the current treatments, which often have side effects. For instance, oral insulin is degraded by the gastrointestinal tract, which reduces its efficacy, and subcutaneous injections pose risks. Metformin, a commonly prescribed medication, can cause adverse reactions such as diarrhea.9 New drugs such as liraglutide and semaglutide also have side effects including nausea and cardiovascular issues.10–12 Therefore, there is an urgent need for novel drugs with fewer side effects and more effective delivery systems.
Emerging research has highlighted the relationship between DM and gut microbiota. Disruptions in intestinal flora, particularly a decrease in butyrate-producing species, have been associated with DM development.13–15 Therefore, improving intestinal flora is a potential therapeutic strategy for DM.16,17 Probiotics have historically been beneficial for enhancing the intestinal flora.18 Probiotics, defined as live microorganisms that confer health benefits when administered in adequate amounts, have shown promise in this area.19–21 Probiotics also possess antioxidant,22 anti-inflammatory,23 and metabolic regulatory properties, making them promising candidates for T1DM and T2DM treatments.24,25 However, orally administered probiotics face challenges, such as reduced survival rates in the gastroenteric environment.26
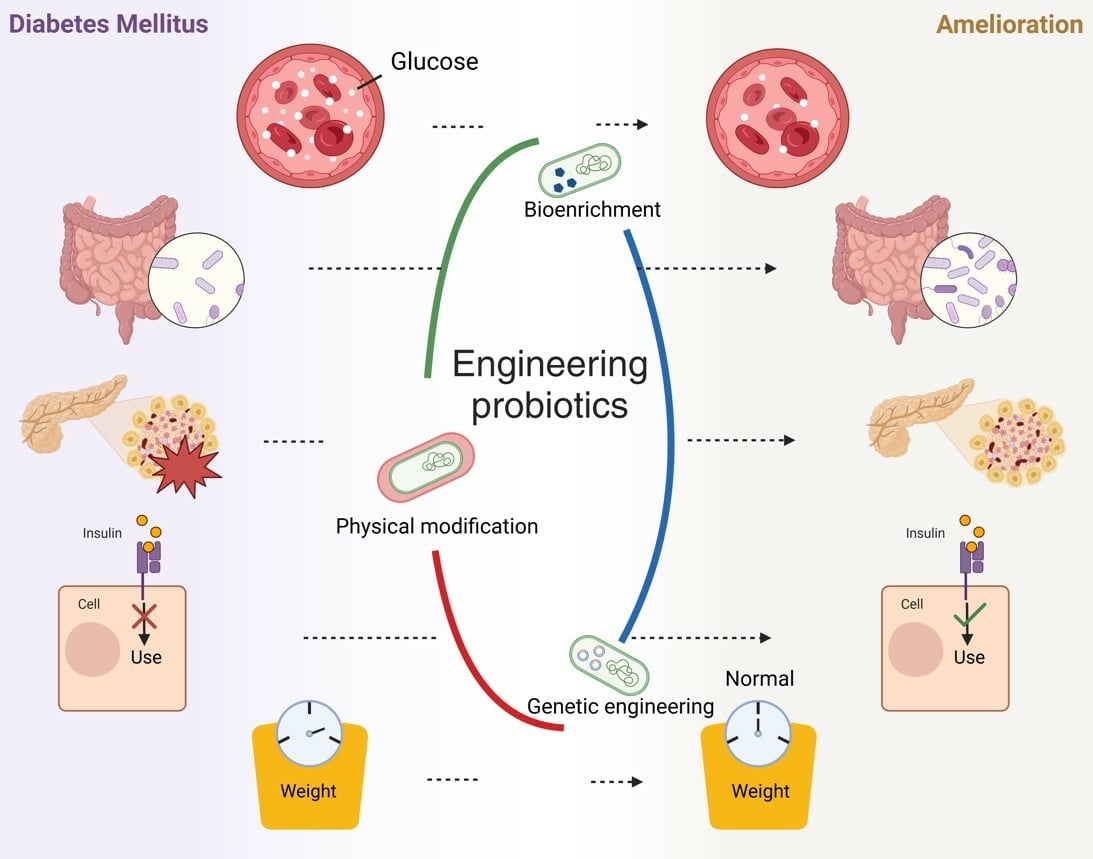
To enhance their efficacy, engineering of probiotics through physical modification, bioenrichment, and genetic engineering is crucial. Physical modifications such as microencapsulation using spray drying and extrusion improve the resilience and survival of probiotics in the digestive tract.27,28 Bioenrichment involves enriching probiotics with trace elements that enhance their metabolic functions and disease-treatment capabilities.29,30 Genetic engineering allows for the incorporation of therapeutic genes into probiotics, enabling targeted drug delivery and improving bioavailability.31
This review discusses the complex interactions between DM and the gut microbiota, design principles and strategies for engineering probiotics, challenges and innovative directions in clinical transformation, and prospects and challenges of engineered probiotics in DM treatment. This study aimed to provide valuable insights for researchers in pharmacy and medicine and to raise public awareness about the potential of engineered probiotics in DM therapy (Figure 1).
 |
Figure 1 Engineered probiotics have higher stability and therapeutic efficacy, but face some challenges needed to be solved. |
Complex Interactions Between DM and Gut Microbiota
The human gut hosts hundreds of different species of microorganisms, predominantly Firmicutes, Bacteroidetes, Proteobacteria and Actinobacteria. Firmicutes and Bacteroidetes are the most abundant, accounting for 60–80% and 20–30% of gut microbiota, respectively.32 These microorganisms maintain mutually beneficial relationships with the host and participate in various metabolic processes. Disturbances in gut microbiota can significantly affect the host.33,34 Alterations in intestinal flora can influence metabolic processes, the intestinal barrier, and the immune response. Probiotics can alleviate DM by improving intestinal flora, enhancing the intestinal barrier, affecting metabolic pathways, and regulating the immune response, thereby contributing to DM alleviation.35 Probiotics can alleviate DM through the same pathway.36
Relationship Between DM and Gut Microbiota
Extensive research has indicated a close relationship between the gut microbiota and the occurrence of DM. Individuals with low microbial diversity in their gut are more likely to exhibit obesity, dyslipidemia, IR, and a pronounced inflammatory phenotype.37 The gut microbiota has been shown to affect the intestinal barrier,38 which in turn can induce T1DM39 and T2DM.40
The intestinal barrier comprises tight junctions and a mucus layer,41 both of which are critical for preventing pathogenic microorganisms from attaching to the intestinal surface and reproducing and invading epithelial cells. Butyrate, a metabolite produced by many intestinal bacteria, promotes the assembly of tight junction,42 enhances mucin-2 expression, and regulates repair of the gut mucus barrier.43 Nevertheless, some bacteria such as Bacteroides species associated with DM development15,44 degrade lactate into substances other than butyrate,45 thereby reducing butyrate production and increasing intestinal permeability. Additionally, bacterial infections and abnormal bacterial colonization can upregulate zonulin, a protein that increases intestinal permeability by affecting the tight junctions.35,46 Increased intestinal permeability allows bacterial lipopolysaccharides (LPS) to enter the bloodstream, a process known as metabolic endotoxemia, which is proposed to contribute to chronic low-grade inflammation, IR, and metabolic syndrome.47 Therefore, a reduction in butyrate-producing bacteria and an increase in harmful bacteria likely contribute to the development of DM.
In addition to butyrate and LPS, other gut microbiota metabolites have been linked to DM.25 Short-chain fatty acids (SCFAs), such as acetic acid, propionic acid, and butyric acid, are the primary products of microbial metabolism of dietary fiber. SCFAs can induce the production of glucagon-like peptide (GLP-1) by stimulating fatty acid receptor 2, thereby promoting insulin production, regulating islet β-cell proliferation, improving IR, and inhibiting inflammation.48,49 Normal metabolism of SCFAs inhibits DM, whereas a reduction in SCFA-producing bacteria contributes to T2DM by impairing anti-inflammatory responses and islet cell function, decreasing insulin sensitivity, and promoting IR.40 Additionally, dietary fiber metabolism primarily occurs in the proximal colon, leaving the distal colon flora to metabolize the remaining proteins. While some protein breakdown products are beneficial, others such as branch-chain amino acids, aromatic amino acids, and trimethylamine are associated with T2DM.25,50
The interaction between the gut microbiota and immune system also plays a role in T1DM.51,52 Toll-like receptors (TLRs), which are pattern recognition receptors expressed on immune and non-immune cells, can recognize pathogen-associated molecular patterns in the microbiome and initiate innate immune responses.53 Diverse microorganisms can promote or inhibit autoimmunity in T1DM through TLRs activations.54,55 MyD88, an adaptor protein of multiple TLRs, recognizes microbial stimuli and promotes downstream TLRs signaling pathways.55 Myd88-null non-obese diabetic (NOD) mice are protected from T1DM under conventional conditions because of their beneficial microbial composition, but they have an increased risk of T1DM under germ-free conditions or antibiotic treatment.56 This suggests that certain beneficial bacteria protect against T1DM, while disturbances or decreased abundance of gut microbiota increases T1DM risk. Moreover, autoimmune T cells show high cross-reactivity with insulin peptides and certain peptides from Bacteroides and Clostridium,57 leading to T1DM. In NOD mice, peptides produced by Parabacteroides with homology to the insulin β chain have been identified,58 and microbial mimic peptides from Leptotrichia goodfellowii have been shown to activate autoimmune responses.59
Probiotics and DM
Disturbances in gut microbiota have been implicated in the development of DM. Probiotics, initially proposed as a treatment owing to their capacity to ameliorate the microbiome,18 have demonstrated potential in managing DM in various clinical trials.60,61 These findings suggest that probiotics can enhance intestinal flora, fortify the intestinal barrier, influence metabolic pathways, and regulate the immune response, thereby ameliorating DM (Figure 2).
 |
Figure 2 Mechanism by which probiotics ameliorate DM. A) Probiotics enhance intestinal flora with preventing the invasion of pathogens and secreting bacteriocins or acidic substances.18 B) probiotics secrete SCFAs, thereby strengthen gut barrier, regulating immune system and producing GLP-1 which protecting the pancreas and promoting insulin secretion.45 C) Probiotics regulate the immune system, stimulate immune cells to secrete IL-10 and IgA, while reducing TNF-α, and exert anti-inflammatory effects.62 D) Probiotics possess antioxidant properties, regulating the activities of related factors, superoxide dismutase (SOD), CAT, nitric oxide (NO), and reactive oxygen species (ROS), through metabolic and signaling pathways.22 |
Probiotics enhance gut health by adhering to the intestinal mucosa, preventing colonization by pathogenic microorganisms, and by producing antibacterial substances.63 They secrete specific proteins that facilitate colonization of the intestinal mucosa by health-promoting microflora, thereby creating spatial barriers that inhibit pathogen colonization.64,65 Furthermore, probiotics can alter the surrounding environment by producing lactic acid, acetic acid, and other substances66 that are intolerable to many gram-negative bacteria.67 Some probiotics produce bacteriocins, which are peptides that inhibit other bacteria by pore formation or inhibition of cell wall synthesis, thereby helping probiotics to compete effectively in their niche.62,68 Additionally, probiotics can enhance the host resistance to pathogenic microorganisms by increasing IgA levels in the intestinal mucosa.69,70 All these pathways provide advantages for probiotics to colonize themselves, resist harmful bacteria, and ameliorate intestinal flora.
Probiotics also protect the gut by increasing butyrate levels and promoting secretion of intestinal barrier-associated proteins.71 Butyrate affects the assembly of tight junctions, the synthesis of mucin-2, and the repair of the gut mucus barrier [ref.]. Certain probiotics, such as Lactobacillus plantarum (L. plantarum), modulate gene transcription pathways related to cell-cell adhesion and enhance tight junction and adhesion junction protein synthesis and degradation by affecting TLR signaling.72 Lactobacillus rhamnosus maintains the epithelial barrier and promotes intestinal epithelial cell activation in response to bacterial infection by enhancing Akt phosphorylation and increasing the expression of Zona occludens-1 and occludin proteins.73
Another crucial mechanism by which probiotics ameliorate DM is through their effects on the glucose metabolic pathways.74 Probiotics enhance SCFA production in humans, thereby increasing insulin secretion, improving IR, and promoting glucose metabolism.75 In addition, probiotics lower blood glucose levels by regulating oxidative stress and inhibiting carbohydrate hydrolases. Oxidative stress, a significant factor in the induction of T2DM, leads to tissue and organ damage, including islet β-cells, kidneys, and the liver.76,77 Probiotics possess antioxidant properties; regulate the activities of superoxide dismutase, catalase, and glutathione peroxidase; and inhibit lipid peroxidation and nitric oxide production through metabolic and signaling pathways, thereby reducing oxidative stress markers.22,78 Furthermore, certain probiotics inhibit the activity of α-glucosidase and α-amylase, influence glucose metabolism, alleviate postprandial hyperglycemia, and regulate blood glucose levels.79
The anti-inflammatory effects of probiotics are vital for DM management. There is a strong correlation between inflammation and the T2DM development.80 Pro-inflammatory factors, such as tumor necrosis factor-α (TNF-α), inhibit the phosphorylation of insulin receptor substrates, negatively affect insulin signaling, and cause IR.81 Other factors, including adipokines, chemotactic proteins, and interleukin-6 (IL-6), are also associated with DM development.82 Probiotics impede pathogen invasion and enhance the intestinal flora composition, thereby preventing the onset.82 The immunomodulatory effects of probiotics have been shown to ameliorate inflammation and to alleviate DM.83 For instance, Lactobacillus acidophilus KLDS1.0901 improved intestinal barrier function and suppressed inflammatory responses in the liver and colon in animal models of T2DM.84 Similarly, Lactobacillus casei CCFM419 attenuated IR, pancreatic islet impairment, and proinflammatory markers in diabetic mice.85 Increased expression of TLR2, TLR4, and TLR9, along with improved production and secretion of TNF-α, interferon-γ (IFN-γ), and IL-10 have been observed in mice treated with probiotics.86 Probiotics also induce naive T cells to differentiate into regulatory T cells (Tregs) and produce high levels of IL-10 and transforming growth factor-β (TGF-β) by activating TLR-2 receptors on macrophages and dendritic cells.87,88 Additionally, Lactobacillus reuteri ATCC PTA 6475 suppressed TNF transcription in monocyte-derived macrophages by inhibiting the activation of mitogen-activated protein kinase-regulated c-Jun and transcription factor activation protein-1.89 Certain probiotics improve obesity, enhance insulin signaling sensitivity, and ameliorate IR by reducing monocyte chemoattractant protein-1 and preserving natural killer T cells.90 Probiotics and their metabolites interact with various immune cells to maintain immune homeostasis by balancing the pro-inflammatory and anti-inflammatory responses.91
Moreover, the immunomodulatory effects of probiotics are also significant in T1DM treatment. Autoreactive T cells attack islet β-cells, leading to insulitis and reduced β-cell mass, resulting in insulin deficiency, which is a key factor in T1DM pathogenesis.92 Probiotics alleviate T1DM by reducing insulitis through anti-inflammatory effects.24 After probiotic intervention, T1DM patients exhibit increased TGF-β1 expression and decreased concentrations of IL-8, IL-17, RANTES (a chemotactic protein), and TNF-α.93 Oral probiotic administration induces IL-10 production, reduces islet β-cell destruction, and prevents DM.94 Probiotic metabolites such as primary SCFAs can limit the frequency of autoreactive T cells. Acetate reduces the frequency of autoreactive T cells in lymphoid tissues by influencing islet β cells and expanding the autoreactive T cell population. Butyrate enhances the number and functionality of regulatory T cells.95
Although the precise mechanisms by which probiotics treat DM remain unclear, a growing body of evidence, including the aforementioned studies, suggests that probiotics have demonstrated promising results in the clinical management of diabetes, and may be an effective therapeutic option for DM.61,96 Probiotics have the potential to increase the levels of beneficial substances, enhance intestinal barrier protection, improve intestinal flora, ameliorate IR, reduce blood glucose levels, and exert other potential effects in DM treatment.
Design Principles and Strategies of Engineered Probiotics
Although probiotics have shown potential in ameliorating DM, their supplementation remains minor compared to that of the resident gut microbiota.18 The unclear mechanism by which probiotics treat DM, combined with the significant limitations and limited efficacy of natural probiotics, necessitate the development of advanced strategies. Oral probiotics are one of the easiest ways to quickly supplement probiotics; however, probiotics that enter the body are greatly affected by the environment, such as pH, enzymes, and mechanical agitation in the digestive tract, which reduce their activity and effectiveness upon reaching the gut.28,97 Probiotic engineering is essential to overcome these challenges and enhance therapeutic efficacy. Currently, the most widely used techniques for probiotic engineering include physical modification, bioenrichment, and genetic engineering (Figure 3). Each method has distinct advantages and caters to various requirements. The following sections describe these techniques in detail:
 |
Figure 3 Three strategies of engineering probiotics and their enhancement. A) Physical modification: Probiotics were coated with special materials to improve their stability and colonization.98 B) Bioenrichment: Probiotics are enriched by trace elements to enhance the production of beneficial metabolites and facilitate the secretion of trace elements.29 C) Genetic engineering: Probiotics are transformed by plasmid to secrete medicinal protein.99 |
Physical Modification
Physical modification is one of the most prevalent methods for engineering probiotics, leveraging materials such as alginates to encapsulate and microencapsulate probiotics and to ensure their resistance to the hostile environment of the gastrointestinal tract.28 These encapsulating materials facilitate the safe delivery of probiotics to their target sites, where they can exert beneficial effects in treating diseases, such as DM. This method has been extensively developed and applied to the treatment of inflammatory bowel disease and DM.100,101
A common microencapsulation technique is extrusion, which is straightforward, cost-effective, and allows modified probiotics to retain their optimal cell viability.27 For instance, Arriaga-Morales et al98 suspended Lactobacillus casei subsp. casei NRRL-1922 in 2% sodium alginate solution. Using a peristaltic pump adapted to the syringe hose, droplets of Lactobacillus casei cell suspension in sodium alginate were added to CaCl2 to form calcium alginate gel particles. The survival rate of immobilized probiotics was 51.9%, whereas that of free probiotics was 2.1%. Immobilized probiotics showed good hypoglycemic effects in streptozotocin-nicotinamide-induced T2DM rats. Compared to the disease group, the blood glucose reduction rate of the immobilized probiotic treatment group was 70.3%. Compared to free probiotics, the serum glucose concentration of the immobilized probiotic treatment group was reduced by 67.8%. In addition, the immobilized probiotic treatment had protective effects on the kidneys and liver. The results demonstrated that the immobilized probiotics prepared using this technique were more effective than free probiotics in the treatment of DM (Figure 4A).
 |
Figure 4 (A) The immobilized Lactobacillus casei, built by extrusion, showed a high survival rate, blood glucose reduction rate and protective effects on the kidney. Reproduced with permission from Arriaga-Morales JJ, Ordaz-Pichardo C, Castro-Muñoz R, Durán-Páramo E. Attenuation of hyperglycemia in diabetic rats assisted by immobilized probiotic in sodium alginate. Probiotics Antimicrob Proteins. 2023.98 Copyright 2023 Springer Nature. (B) Probiotic product, created by spray drying, had great stability. Full factorial analysis of variance showing the main effects and interactions of the variables evaluated. G1 is an index of a sample’s elastic behavior and represents the deformation energy stored in the sample during the shear process. G2 value measures the viscous component of a sample and compares the energy lost during the shear process. Control = milk chocolate formulation, Prob = milk chocolate + probiotics, FO = milk chocolate + fish oil, Prob + FO = milk chocolate + probiotics + fish oil, Sw = isomalt + stevia, Sw + Prob = isomalt + stevia + probiotics, Sw + FO = isomalt + stevia + fish oil, Sw + Prob + FO = isomalt + stevia + probiotics + fish oil. Reproduced with permission from Gómez-Fernández AR, Faccinetto-Beltrán P, Orozco-Sánchez NE, et al Sugar-free milk chocolate as a carrier of Omega-3 polyunsaturated fatty acids and probiotics: a potential functional food for the diabetic population. Foods. 2021;10:1866.102 Copyright 2021 by the authors, licensee MDPI, Basel, Switzerland. (C) The process of constructing SD-S-SNEDDS, the SEM image of SD-S-SNEDDS and cumulative release of drugs. Reprinted from International Journal of Pharmaceutics, 612, Rubiya Khursheed, Sachin Kumar Singh, Bimlesh Kumar, Sheetu Wadhwa, Monica Gulati, Anupriya A, Ankit Awasthi, Sukriti Vishwas, Jaskiran Kaur, Leander Corrie, Arya K.R., Rajan Kumar, Niraj Kumar Jha, Piyush Kumar Gupta, Flavia Zacconi, Kamal Dua, Nitin Chitranshi et al, Self-nanoemulsifying composition containing curcumin, quercetin, Ganoderma lucidum extract powder and probiotics for effective treatment of type 2 diabetes mellitus in streptozotocin inducedrats, 121306, Copyright (2024), with permission from Elsevier103 (D) Schematic illustration of the preparation of encapsulating Escherichia coli Nissle 1917, the TEM image of Escherichia coli (EcN) and encapsulated Escherichia coli (LEG) and the blood glucose level and insulin level of T2DM mice after different treatments. Reproduced with permission from Wang Y, Shi Y, Peng X, et al Biochemotaxis-oriented engineering bacteria expressing GLP-1 enhance diabetes therapy by regulating the balance of immune. Adv Healthcare Mater. 2024;13:2303958. © 2024 Wiley-VCH GmbH104 Asterisks indicate significant difference from a full factorial analysis of variance showing the main effects and interactions of the variables evaluated: *p < 0.05, **p < 0.01, ***p < 0.001. |
Encapsulation can include both probiotics and therapeutic substances to enhance the therapeutic effects. For example, Gómez-Fernández et al102 used a spray-drying method, a more efficient microencapsulation method, spray-dried L. plantarum 299v, and Lactobacillus acidophilus La3 suspended in 750 mL of a microencapsulation mix (10% w/v maltodextrin and w/v 2% food-grade alginate) at 130 °C inlet, 60 °C outlet, and 0.13 MPa. The microcapsules were then packaged with fish oil (as an additional medicine), using sugar-free milk chocolate, to create a stabilized probiotic product. The obtained probiotic products exhibited high probiotic content (7 × 1013 CFU/g of L. plantarum 299v and 1×1014 CFU/g of Lactobacillus acidophilus La3), and the ω-3 polyunsaturated fatty acid content in fish oil was 133.8 ± 8.76 mg per 12 g of probiotic products (Figure 4B).
Lipophilic drugs, such as fish oil, which are effective in DM treatment, face challenges in oral delivery owing to limited dissolution and bioavailability. Self-nanoemulsifying drug delivery systems (SNEDDS) can address this issue. Khursheed et al103 formulated an optimal ratio of four substances, diethylene glycol monoethyl ether, twin 80, oleoyl polyoxyl-6 glycerides, and mono-diglyceride, which are used to dissolve the lipophilic plant drugs curcumin and quercetin, both of which have been reported to have antidiabetic effects. SD-S-SNEDD was prepared by spray drying S-SNEDDS, which was prepared by ganoderma lucidum extract powder and probiotics as solid supports based on L-SNEDDS. To further improve its physical and mechanical stability, SD-S-SNEDDS was transformed into spherical particles via extrusion spheronization. The result found The zeta potential of S-SNEDDS was −38.7 mV, indicating that S-SNEDDS exhibited better absorption and oxidation stability. The drug loading percentages of curcumin and quercetin in SD-S-SNEDDS were 97.8±2.32% and 96.5±3.17%, respectively, and more than 90% of the drug was released within the first 5 min. Furthermore, SD-S-SNEDDS exhibited excellent absorption and bioavailability and enhanced α-glucosidase inhibition. In diabetic rats, SD-S-SNEDDS showed hypoglycemic, lipid-lowering, liver- and kidney-protective effects and a favorable recovery of diabetic parameters (Figure 4C).
Liposomes are popular drug delivery systems in which the phospholipid bilayer provides excellent protection.105 Moreover, certain phospholipids have been shown to exert adjunctive antidiabetic effects. Recently, Wang et al104 encapsulated Escherichia coli Nissle 1917, which is capable of expressing GLP-1, using a film dispersion method with a dilauroyl phosphatidylcholine-based lipid membrane. Lipid membranes protect probiotics from attack by the gastrointestinal environment, effectively inhibiting the production of reactive oxygen species, alleviating oxidative stress, and repairing pancreatic islet β-cell dysfunction with GLP-1. Furthermore, the engineered probiotics regulated insulin secretion, upregulated the expression of key proteins, and enhanced the richness and diversity of intestinal flora (Figure 4D).
Bioenrichment
Trace elements are crucial for the functioning of living organisms including humans and probiotics.106 Bioenrichment involves enriching trace elements with probiotics to enhance therapeutic efficacy. Introducing trace elements into probiotics has been shown to ameliorate DM independently and to enhance the therapeutic effects of probiotics by strengthening metabolic enzymes or pathways. This approach combines the direct and indirect therapeutic effects of trace elements with those of probiotics to exert a greater influence on the body than their individual effects.
In probiotics, some enzymes or other substances produced by metabolism can improve DM. For example, dihydronicotinamide-adenine dinucleotide and glucuronic acid dehydrogenase in Acetobacter aceti (A. aceti) mediate the glycolysis pathway, enabling the breakdown of glucose and the subsequent achievement of a hypoglycemic effect. However, the amount of these substances in normal probiotics does not meet the expectations for disease treatment. Huang et al29 devised a bioenrichment method to enhance the metabolites in probiotics by increasing the quantity of zinc and chromium in probiotics, which in turn enhances the healing capacity of the disease. Notably, zinc, an important component of insulin, can significantly ameliorate glucose tolerance and glucose-stimulated insulin secretion by enhancing the function of islet β-cells, thereby improving and preventing DM.107,108 Similarly, Cr has been shown to exert beneficial effects on blood glucose control.109 They obtained chromium- and zinc-rich A. aceti after being cultured for a period of time (48 h) in liquid medium containing 64 mg/mL chromium trichloride and zinc chloride. After detection, the content of chromium metal, metallic zinc, NADH coenzyme, and glucose dehydrogenase in A. aceti prepared by this method were 28.58–34.34 mg/kg, 5.35–7.52 mg/kg, 5.13–7.26 μM, and 446.812–567.138 U/g, respectively. The levels of these substances were higher than those in uncultured Acetobacter spp. The fasting blood glucose level of diabetic mice treated with the modified probiotics was lower than that of mice treated with metformin tablets. It also protects islet cells, alleviates islet tissue damage, and restores the body weight. It is worth mentioning that mice treated with ten-fold dose of A. aceti rich in chromium and zinc did not show pathological damage to the liver, spleen, kidney, or stomach, which was sufficient to prove the safety of the engineered probiotics (Figure 5A).
 |
Figure 5 (A) The content of related substances in Cr- and Zn-rich A. aceti had been improved, and they had good hypoglycemic effect, and had no damage to many organs. Reproduced with permission from Huang -Y-Y, Qin X-K, Dai -Y-Y, et al Preparation and hypoglycemic effects of chromium- and zinc-rich Acetobacter aceti. World J Diabetes. 2022;13:442–453..29 Copyright 2022, the Author(s). Published by Baishideng Publishing Group Inc. All rights reserved. (B) Se content in the pancreas had been enhanced and the glycated hemoglobin (Hb1Ac), blood glucose level and pancreatic injury had been reduced under the treatment of Se-B. longum DD98. NC, normal control group; T2DM, model group; SS, sodium selenite group; BL, (B) longum DD98 group; SeL, Se-B. longum DD98 low dosage group; SeH, Se-B. longum DD98 high dosage group. a–eMeans with different letters differ significantly for the same indicator after different treatments (p < 0.05). Used with permission of Royal Society of Chemistry, from Antidiabetic effects of selenium-enriched Bifidobacterium longum DD98 in type 2diabetes model of mice, Zhao D, Zhu H, Gao F, et al, 11, 7, copyright 2010; permission conveyed through Copyright Clearance Center, Inc. 110 (C) The content of fasting blood glucose levels, body weights and higher fasting insulin levels, and the expression of insulin signaling pathway proteins had been improved by Nano-Se-B. longum. #p < 0.05, ##p < 0.01, ###p < 0.001 compared with normal group; *p < 0.05, **p < 0.01 compared with the model group. Reproduced with permission from Lin Y, Ren Y, Zhang Y, et al Protective role of nano-selenium-enriched Bifidobacterium longum in delaying the onset of streptozotocin-induced diabetes. R Soc Open Sci. 2018;5:181156.111 Copyright 2018 The Authors. Published by the Royal Society under the terms of the Creative Commons Attribution License. |
Selenium (Se), another essential micronutrient, has demonstrated antidiabetic properties by promoting sustained improvement in glucose homeostasis and regulating key metabolic processes such as glycolysis, gluconeogenesis, and lipid metabolism.112,113 Zhao et al110 used a similar approach to fermenter Bifidobacterium longum DD98 in selenium-rich medium, producing selenium-enriched Bifidobacterium longum DD98 (Se-B. longum DD98). Oral administration of Se-B. longum DD98 in diabetic mice reduced fasting blood glucose levels, glycosylated hemoglobin levels, and pancreatic injury, and improved glucose tolerance, outperforming both untransformed B. longum DD98 and Na2SeO3 treatments (Figure 5B).
Lin et al111 advanced this approach using selenium nanoparticles. They pretreated selenium with a mixture of sodium selenite, reduced glutathione, and bovine serum albumin to produce red nano-selenium. After dialysis with double-distilled water, magnetic stirring, and centrifugation, the purified selenium nanoparticles were obtained. Bifidobacterium longum NQ-1501 was cultured anaerobically overnight in TPY medium containing 25 μg/mL sodium selenite and 5 μg/mL selenium nanoparticles to obtain Nano-Se-B. longum. It is pertinent to note that the selenium in Nano-Se-B. longum exists in the form of selenoproteins, which have been documented to exhibit a high bioavailability and low toxicity.114 Diabetic mice induced with streptozotocin (STZ) treated with Nano-Se-B. longum exhibited significantly lower fasting blood glucose levels, higher body weights, and higher fasting insulin levels than the WT B. longum. Moreover, Nano-Se-B. longum alleviated pathological damage to the liver, pancreas, and kidney and improved the sensitivity of hepatic insulin signaling (Figure 5C).
Genetic Engineering
Genetic engineering is a widely used method for modifying probiotics for the treatment of DM. This approach involves the introduction of genes encoding therapeutic proteins into the probiotics. Engineered probiotics can produce and deliver these proteins directly to specific locations in the gastrointestinal tract, thereby enhancing their therapeutic efficacy by protecting them from degradation. Probiotics can also serve as expression vectors to synthesize therapeutic substances, making treatment more accessible and cost-effective.31 Key therapeutic proteins used in this context include insulin and its analogs, GLP-1, GLP-1 receptor agonists (GLP-1RAs), antigens, ILs, and other proteins known to play roles in DM treatment (Table 1).
 |
Table 1 The Summary of Major Studies on the Use of Engineering Probiotics in the Management of DM |
Insulin and its Analogues
In 1978, Goeddel et al pioneered the synthesis of the first recombinant human insulin DNA by combining single strands of A and B expressed in Escherichia coli.115 Most insulin produced in this way is suitable for injection rather than oral administration. Owing to the advantages of oral insulin,116 researchers have focused on expressing insulin in probiotics to simulate oral insulin delivery. Lactococcus lactis (L. lactis), a safe and well-characterized probiotic, is commonly used for this purpose.117,118 However, synthesizing insulin, a heterodimeric protein with three disulfide bonds, using L. lactis presents specific challenges.
To overcome this challenge, Ng et al119 expressed the single-chain insulin analog SCI-57 in L. lactis ZN9000. SCI-57, designed for oral administration, simplifies synthesis while maintaining biological activity and stability comparable to those of wild-type insulin. They constructed the plasmid pNZPnisA:uspSCI-57his, which was transformed into L. lactis strain ZN9000. Recombinant L. lactis successfully secreted active SCI-57 under the induction of nisin at certain pH conditions.
Mao et al120 developed another single-chain insulin, SCI-59, linked by an eight-residue linker between insulin B and A chains. SCI-59 was optimized and introduced into L. lactis NZ3900. Rather than using the recombinant bacteria directly, SCI-59-3LysM was combined with non-viable LAB to avoid the use of antibiotic resistance genes and to enhance stability. This engineered probiotic reduces DM incidence, improves glucose tolerance, promotes C-peptide secretion, ameliorates insulitis, and improves the regulatory immune response in NOD mice.121
Glp-1
GLP-1 is a 30-amino acid peptide hormone secreted by intestinal cells that plays a crucial role in glycemic control in patients with T2DM by interacting with the GLP-1 receptor.122 GLP-1 is rapidly degraded by dipeptidyl peptidase-4 (DPP-4) and trypsin, leading to a short half-life. To address this issue, researchers have explored various strategies to enhance the stability and efficacy of GLP-1 delivered through probiotics.
Hu et al123 synthesized the plasmid pUC-SPglnPH2-GLP-1 by incorporating the codon-optimized GLP-1 gene with glnPH2 signal peptide sequences (SPglnPH2) in the pUC57 vector. This DNA fragment was PCR-amplified, digested, and ligated into the pMG36e plasmid, which was then electrotransformed into L. plantarum to produce L. plantarum-pMG36e-GLP-1. In T2DM mice, these engineered bacteria effectively reduced blood glucose levels, improved glucose tolerance, and inhibited weight gain by reducing pancreatic inflammation, improving the intestinal flora, and regulating the expression of genes related to fat metabolism.
To further enhance GLP-1 stability, specific amino acid modifications have been made. The Ala at position 8 and Lys at positions 26 and 34 in the DNA sequence of mature active human GLP-1 (7–36) are sensitive to DPP-4 and trypsin. The alteration of these amino acids renders these enzymes incapable of recognizing GLP-1 effectively, thereby increasing half-life of GLP-1.124 For example, Wang et al125 mutated Ala8 to Gly8 in human GLP-1(7–36) to prevent DPP-4 recognition, and Lys26 and 34 to Gln26 and Asp34 to inhibit trypsin digestion. These modified GLP-1 genes, which are suitable for expression in L. plantarum WCFS1, were cloned into the plasmid pSIP403. Recombinant L. plantarum WCFS1 demonstrated improved glucose control, including lower fasting blood glucose levels, extended blood glucose maintenance, inhibited glucagon secretion, promoted β-cell proliferation, and reduced renal damage.
In addition, researchers have explored the use of tandem repeats of GLP-1 to enhance its stability and resistance to enzymatic degradation. Hou et al126 constructed plasmids with multiple GLP-1 repeats (2×, 4×, and 8× GLP-1) via sequential ligation and transformation in E. coli BL21(DE3) cells. The resulting strains successfully secreted GLP-1 analogs, which reduced serum glucose levels in diabetic rats. Subsequently, 10×GLP-1 was successfully generated by a similar method.127 It is worth mentioning that Fangfang Xu et al128 further improved stability by adding cysteine residues to form a disulfide bond between GLP-1 dimers. This approach has been successfully applied to probiotics and has demonstrated enhanced stability and efficacy.129
Enhancing the expression and efficiency of GLP-1 in probiotics can improve their therapeutic outcomes. Wu et al130 constructed an efficient expression vector, pNK1-PGK-10×GLP-1, and transformed it into Saccharomyces cerevisiae INVSc1. The recombinant yeast, named lLHY168, expressed GLP-1 at 1.56 mg/g cell wet weight with stable gene retention. The engineered yeast showed significant benefits in treatingT2DM mice.
GLP-1RAs
GLP-1RAs represent a novel class of glucose-control drugs that have gained widespread use in the treatment of T2MD in recent years. These agonists mimic GLP-1 by binding to its receptor and promoting insulin secretion, thus aiding in the regulation of blood sugar levels. One key advantage of GLP-1RAs is their structural homology with GLP-1, which makes them resistant to degradation by DPP-4, thereby extending their half-life and enhancing their therapeutic potential.131
Exendin-4 (Exd4) is a crucial component of GLP-1RAs. It is a 39-amino-acid peptide with a 53% sequence homology to human GLP-1.132 Exenatide, the first GLP-1RA drug, was approved by the US FDA in 2005 as an adjunctive therapy to improve blood sugar control in T2DM patients.133 Zeng et al134 demonstrated the feasibility of introducing Exd4 into probiotics, enabling engineered probiotics to produce and deliver Exd4 directly to the gut. They optimized the codon sequence of Exd4 for lactic acid bacteria and inserted it into the plasmid vector pMG76e, which also included the signal peptides Usp45 and LEISSTCDA, Exd4, a 6×His tag, and restriction sites XbaI and XhoI. The transformation of L. paracasei L14 with this recombinant plasmid resulted in the successful production of active Exe-4, which was able to cross the Caco-2 cell monolayer, promote insulin secretion, enhance β-cell proliferation, and reduce apoptosis. Subsequently, the same research group135 refined their methodology by synthesizing an oligonucleotide that included the propeptide sequences of Usp45 and LEISSTCDA, codon-optimized Exe-4, and restriction sites SacI and Hin dIII. This oligonucleotide was inserted into the NICE vector pNZ8048, which was then used to transform L. lactis NZ9000. The maximum concentration of Exd4 secreted by recombinant Lactococcus lactis was 249 nmol/L at a bacterial concentration of 6×108 cfu/mL. This secretion significantly stimulated insulin production in rat pancreatic β cell line INS-1, reaching 69.96±0.74 ng/mL, which outperformed the previous preparation methods.
Antigens and ILs
T1DM is an autoimmune disease in which insulin or other hypoglycemic drugs can alleviate symptoms, but do not address the underlying cause. Currently, antigenic vaccines are promising immunotherapies for treating T1DM.136 Robert et al99 have highlighted the potential of recombinant L. lactis as an effective tool for inducing antigen-specific oral tolerance. Liang et al137 successfully expressed a fusion protein (Hsp65-6×P277) in E. coli BL21 (DE3) using the plasmid pET28-Hsp65-6×P277. This protein elicits an anti-inflammatory immune response, thus providing a potential vaccine against T1DM. However, the instability of HSP65 requires further refinement. Ma et al138 constructed pHJ: HSP65-6P277 by replacing the nisin-inducible promoter PnisA with the constitutive promoter P32, thereby enhancing stability and expression in L. lactis NZ9000. This modified strain showed lower T1DM incidence and reduced islet inflammation in treated mice than in controls.
Furthermore, Kun-Feng Liu et al139 took a different approach, integrating the PCR product 6IA2P2 with plasmids pCYT: HSP65-P277 and pHJ: HSP65-6P277 to generate pCYT: HSP65-6IA2P2 and pHJ: HSP65-6IA2P2. Transformed L. lactis NZ9000 exhibited prolonged expression in the intestinal mucosa and long-lasting immunoregulatory effects, effectively regulating blood sugar levels and reducing insulitis in NOD mice.
Another promising antigen is the colonization factor antigen I (CFA/I), which has shown efficacy in evoking bystander immunity and accelerating the development of antigen-specific Tregs. Massimo Maddaloni et al140 synthesized pBzMM153 containing the cfaI operon, which was transformed into lactis IL1403. The recombinant strain regulates the immune system, ameliorates intestinal flora, and provides long-term protective effects.141,142
ILs play an important role in the immune response. The strategy of expressing ILs genes in probiotics has shown the potential for enhancing antigen-induced immune responses. L. lactis expressing IL-10 has been established to alleviate murine colitis,143 a technique adapted for DM treatment.144 The main strategy is to insert ILs gene in the developed expression system in lactic acid bacteria, which make the lactic acid bacteria has the ability of expressing and secreting ILs.145 Recombinant L. lactis expressing both proinsulin and IL-10 have been successfully used to prevent T1DM.146–148 For instance, Robert et al149 further demonstrated the use of recombinant L. lactis expressing glutamic acid decarboxylase 65 and IL-10 to effectively prevent T1DM by improving immune regulation and reducing inflammation.
Other Proteins
Genetic engineering has enabled the introduction of various proteins into probiotics for the treatment of DM. These engineered probiotics can produce specific proteins with therapeutic effects on DM. In addition to the categories mentioned in the previous section, notable examples of this approach exist.
Staphylococcal Nucleases: Chinese researchers developed a strain of L. lactis that expresses staphylococcal nucleases (CN Patent 201610353343.5), targeting DM treatment.150 This genetically modified probiotic can disrupt neutrophil extracellular traps, reduce inflammation, regulate blood glucose levels, and delay the onset of diabetes with lower mortality and morbidity.
L-Arabinose Isomerase: Moez Rhimi et al151 engineered L. lactis to express L-arabinose isomerase, which can convert galactose into tagatose, a sugar with a low glycemic index, thereby reducing hyperglycemia. This modification utilized an expression system developed for L. lactis, enhancing its potential for diabetes management.
Challenges and Innovations in Clinical Translation of Engineered Probiotics
Clinical Innovative Applications and Prospects of Engineered Probiotics
Engineered probiotics have garnered attention for their multifaceted roles in managing diabetes and associated complications. This section discusses their distinct advantages, synergistic effects, and their broader applications across different diseases, particularly in addressing diabetic complications.
Advantages of Engineered Probiotics in Diabetes Treatment
Engineered probiotics offer several unique advantages that distinguish them from traditional treatments for diabetes. These advantages include:
- Targeted metabolic modulation: Engineered probiotics can produce bioactive compounds such as GLP-1, which directly enhances insulin secretion and improves glucose metabolism.123 This allows for a more precise regulation of blood glucose levels.
- Restoration of gut microbiota: Engineered probiotics contribute to a healthier gut microbiota, which is vital for enhancing insulin sensitivity and reducing systemic inflammation.152 Balanced gut flora is increasingly recognized as a crucial factor in managing diabetes and preventing complications.
- Non-invasive and patient-friendly: Probiotics, available in oral formulations, offer a non-invasive alternative to insulin injections, improving patient adherence.153
- Personalized treatment potential: Engineering probiotics allows for customization based on an individual’s microbiome profile, facilitating personalized therapeutic interventions that are tailored to the specific needs of each patient.154
Multiple Probiotics or Synergies Between Probiotics and Drugs
While single-strain probiotics are beneficial, multi-strain probiotics offer enhanced biological activity due to the additive and synergistic effects among individual strains. Research has shown that multi-strain probiotics can significantly improve glucose metabolism, modulate immune responses, and restore gut microbiota more effectively than single-strain formulations.155
Besides, the co-administration of probiotics with herbal therapies demonstrates significant synergistic effects. Recent studies have shown that combining probiotics with specific herbal extracts (such as garlic, fenugreek, and berberine) enhances therapeutic outcomes. These herbs, when co-administered with probiotics, optimize glucose metabolism, reduce insulin resistance, and modulate gut microbiota.156,157 Similarly, when engineered probiotics are co-administered with active pharmaceutical ingredients (APIs) like metformin, they can improve drug bioavailability, enhance gut health, and mitigate gastrointestinal side effects, such as those commonly associated with API treatments.158
Engineered Probiotics and Diabetic Complications
Engineered probiotics play a crucial role in mitigating diabetic complications, which are often driven by chronic inflammation, oxidative stress, and endothelial dysfunction. Diabetic complications, such as neuropathy, retinopathy, and cardiovascular diseases, can be alleviated through the use of probiotics that modulate gut microbiota, reduce systemic inflammation, and improve lipid profiles.159,160
Furthermore, while our focus remains on diabetes, the potential applications of engineered probiotics extend to other conditions, such as inflammatory bowel disease and metabolic syndrome, or even Alzheimer’s disease.143,161–163 Their ability to restore gut microbiota balance and enhance immune responses makes them valuable candidates for a variety of therapeutic contexts.
Clinical Translation Challenge of Engineered Probiotics
While engineered probiotics hold great promise for diabetes management, several challenges must be addressed for successful clinical application.164 These include ensuring the stability and viability of probiotics during storage and delivery, navigating complex regulatory landscapes, and generating robust clinical trial data to validate efficacy and safety. Additionally, understanding patient-specific factors, such as individual microbiome profiles, is crucial for optimizing therapeutic outcomes (Figure 6). Addressing these issues will be essential for translating engineered probiotics from research into routine clinical practice.
 |
Figure 6 The challenges and corresponding innovation in clinical translation of engineered probiotics. |
Safety Concerns
Most engineered probiotics are genetically modified organisms (GMOs). These probiotics express the desired substances by accepting foreign recombinant plasmids, which are under strict regulatory scrutiny.165 Such GMOs are prone to mutations or plasmid loss, leading to the loss of therapeutic substance expression in subsequent generations and the potential production of harmful substances, posing risks of genetic instability. Furthermore, the inclusion of antibiotic-resistance genes in plasmids for screening during production poses additional safety concerns. The release of probiotics with these resistance genes into the environment can lead to the transfer of resistance genes to other microorganisms, thereby exacerbating the antibiotic resistance crisis. Not all orally administered probiotics can colonize the gut, and some are excreted and proliferate in nature, potentially contaminating the environment.
To address these issues, it is crucial to develop more efficient and user-friendly genetic engineering methods to reduce operational complexity and costs, while minimizing potential risks. Innovation of recombinant protein surface display technologies to link therapeutic proteins to the surfaces of unmodified probiotics can help avoid the use of GMOs and ensure drug targeting and efficacy. Moreover, accelerating the transition from traditional microencapsulation to nanoencapsulation technologies can enhance the survival rate of probiotics and enable their large-scale application.
Operational Challenges
The probiotics used in the gut are predominantly anaerobes, which are highly sensitive to oxygen, posing significant challenges in their isolation, cultivation, industrial production, and formulation. Genetic engineering techniques require high operational standards and many microorganisms lack manipulable genetic tools. Current research focuses on a limited subset of the vast microbial family, restricting modifications to a few strains.31 Moreover, the stringent safety requirements for engineered probiotics have increased technical demands, such as prohibiting the use of antibiotic resistance genes in probiotics, complicating the screening process, or designing probiotics that do not proliferate extensively in the environment, necessitating the creation of nutritionally deficient strains.
Enhancing scientific and technological advancements to develop more efficient genetic engineering methods can reduce operational complexity and costs, while ensuring the safety of production strains. Exploring and developing surface display technologies and nano-encapsulation methods can enhance probiotic viability and facilitate large-scale probiotic modifications and application.100
Regulatory and Public Acceptance
Regulatory hurdles and public skepticism towards GMOs significantly impede the clinical translation of engineered probiotics. Strict regulations govern the use of GMOs owing to their potential environmental and health risks. Additionally, public acceptance of GMOs remains low, further complicating their clinical adoption.
Extensive research and public outreach to demonstrate the safety and efficacy of engineered probiotics, engaging with regulatory bodies to streamline the approval processes for probiotics with well-documented safety profiles and promoting transparency in the development and testing of engineered probiotics can build public trust and acceptance.
Conclusion
The intricate relationship between the gut microbiota and diabetes highlights the potential of engineered probiotics to manage the disease. Although conventional probiotics have demonstrated efficacy, their limitations necessitate advancements in engineering techniques. This review details the physical, bioenrichment, and genetic modifications that enhance the probiotic stability, functionality, and therapeutic efficacy. Future studies should focus on elucidating the mechanisms of action, improving the genetic engineering methods, and developing novel therapeutic proteins. Engineered probiotics offer a promising alternative to conventional therapies, potentially reducing side effects and expanding treatment options for diabetes and other diseases. The successful application of these innovations could mark a significant milestone in diabetes management and in broader medical applications.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This work was supported by the National Natural Science Foundation of China (51901160), Natural Science Foundation of Zhejiang Province (LY22H020005), and Graduate Scientific Research Foundation of Wenzhou University (3162024003057, 3162024003056, and 3162024004102).
Disclosure
The authors report no conflicts of interest in this work.
References
1. American Diabetes Association Professional Practice Committee. 2. Diagnosis and classification of diabetes: standards of care in diabetes—2024. Diabetes Care. 2023;47:S20–42. doi:10.2337/dc24-S002
2. Zhou B, Lu Y, Hajifathalian K, et al. Worldwide trends in diabetes since 1980: a pooled analysis of 751 population-based studies with 4·4 million participants. Lancet. 2016;387:1513–1530. doi:10.1016/S0140-6736(16)00618-8
3. Ceriello A, Prattichizzo F. Variability of risk factors and diabetes complications. Cardiovasc Diabetol. 2021;20:101. doi:10.1186/s12933-021-01289-4
4. Magliano DJ, Boyko EJ. IDF diabetes Atlas 10th edition scientific committee. In: IDF Diabetes Atlas.
5. Sun H, Saeedi P, Karuranga S, et al. IDF diabetes Atlas: global, regional and country-level diabetes prevalence estimates for 2021 and projections for 2045. Diabetes Res Clin Pract. 2022;183:109119. doi:10.1016/j.diabres.2021.109119
6. Xia Y, Xie Z, Huang G, Zhou Z. Incidence and trend of type 1 diabetes and the underlying environmental determinants. Diabetes/Metab Res Rev. 2019;35:e3075. doi:10.1002/dmrr.3075
7. Bjornstad P, Chao LC, Cree-Green M, et al. Youth-onset type 2 diabetes mellitus: an urgent challenge. Nat Rev Nephrol. 2023;19:168–184. doi:10.1038/s41581-022-00645-1
8. Jia W, Weng J, Zhu D, et al. Standards of medical care for type 2 diabetes in China 2019. Diabetes Metab Res Rev. 2019;35:e3158. doi:10.1002/dmrr.3158
9. Sanchez-Rangel E, Inzucchi SE. Metformin: clinical use in type 2 diabetes. Diabetologia. 2017;60:1586–1593. doi:10.1007/s00125-017-4336-x
10. Pratley R, Amod A, Hoff ST, et al. Oral semaglutide versus subcutaneous liraglutide and placebo in type 2 diabetes (PIONEER 4): a randomised, double-blind, phase 3a trial. Lancet. 2019;394:39–50. doi:10.1016/S0140-6736(19)31271-1
11. Huang L, Yu H, Fang Y. A diabetic patient increased premature ventricular contractions after using liraglutide: a case report. Front Cardiovasc Med. 2024;11:1332754. doi:10.3389/fcvm.2024.1332754
12. Smits MM, Van Raalte DH. Safety of semaglutide. Front Endocrinol. 2021;12:645563. doi:10.3389/fendo.2021.645563
13. Crudele L, Gadaleta RM, Cariello M, Moschetta A. Gut microbiota in the pathogenesis and therapeutic approaches of diabetes. eBioMedicine. 2023;97. doi:10.1016/j.ebiom.2023.104821
14. de Goffau MC, Fuentes S, van den Bogert B, et al. Aberrant gut microbiota composition at the onset of type 1 diabetes in young children. Diabetologia. 2014;57:1569–1577. doi:10.1007/s00125-014-3274-0
15. Zhou Z, Sun B, Yu D, Zhu C. Gut microbiota: an important player in type 2 diabetes mellitus. Front Cell Infect Microbiol. 2022;12:834485. doi:10.3389/fcimb.2022.834485
16. Yang G, Wei J, Liu P, et al. Role of the gut microbiota in type 2 diabetes and related diseases. Metab Clin Exp. 2021:117. doi:10.1016/j.metabol.2021.154712
17. de Groot P, Nikolic T, Pellegrini S, et al. Faecal microbiota transplantation halts progression of human new-onset type 1 diabetes in a randomised controlled trial. Gut. 2021;70:92–105. doi:10.1136/gutjnl-2020-322630
18. Wieërs G, Belkhir L, Enaud R, et al. How probiotics affect the microbiota. Front Cell Infect Microbiol. 2020;9:454. doi:10.3389/fcimb.2019.00454
19. Li G, Feng H, Mao X-L, et al. The effects of probiotics supplementation on glycaemic control among adults with type 2 diabetes mellitus: a systematic review and meta-analysis of randomised clinical trials. J Transl Med. 2023;21:442. doi:10.1186/s12967-023-04306-0
20. Kocsis T, Molnár B, Németh D, et al. Probiotics have beneficial metabolic effects in patients with type 2 diabetes mellitus: a meta-analysis of randomized clinical trials. Sci Rep. 2020;10:11787. doi:10.1038/s41598-020-68440-1
21. Hill C, Guarner F, Reid G, et al. Expert consensus document. The international scientific association for probiotics and prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat Rev Gastroenterol Hepatol. 2014;11:506–514. doi:10.1038/nrgastro.2014.66
22. Wang Y, Wu Y, Wang Y, et al. Antioxidant properties of probiotic bacteria. Nutrients. 2017;9:521. doi:10.3390/nu9050521
23. Virk MS, Virk MA, He Y, et al. The anti-inflammatory and curative exponent of probiotics: a comprehensive and authentic ingredient for the sustained functioning of major human organs. Nutrients. 2024;16:546. doi:10.3390/nu16040546
24. Dovi KS, Bajinka O, Conteh I. Evidence and possible mechanisms of probiotics in the management of type 1 diabetes mellitus. J Diabetes Metab Disord. 2022;21:1081–1094. doi:10.1007/s40200-022-01006-2
25. Zhai L, Wu J, Lam YY, Kwan HY, Bian Z-X, Wong HLX. Gut-microbial metabolites, probiotics and their roles in type 2 diabetes. Int J Mol Sci. 2021;22:12846. doi:10.3390/ijms222312846
26. Dodoo CC, Wang J, Basit AW, Stapleton P, Gaisford S. Targeted delivery of probiotics to enhance gastrointestinal stability and intestinal colonisation. Int J Pharm. 2017;530:224–229. doi:10.1016/j.ijpharm.2017.07.068
27. Yoha KS, Nida S, Dutta S, Moses JA, Anandharamakrishnan C. Targeted delivery of probiotics: perspectives on research and commercialization. Probiotics Antimicrob Proteins. 2022;14:15–48. doi:10.1007/s12602-021-09791-7
28. Yao M, Xie J, Du H, McClements DJ, Xiao H, Li L. Progress in microencapsulation of probiotics: a review. Compr Rev Food Sci Food Saf. 2020;19:857–874. doi:10.1111/1541-4337.12532
29. Huang -Y-Y, Qin X-K, Dai -Y-Y, et al. Preparation and hypoglycemic effects of chromium- and zinc-rich Acetobacter aceti. World J Diabetes. 2022;13:442–453. doi:10.4239/wjd.v13.i6.442
30. Yang J, Yang H. Recent development in Se-enriched yeast, lactic acid bacteria and bifidobacteria. Crit Rev Food Sci Nutr. 2023;63:411–425. doi:10.1080/10408398.2021.1948818
31. Ma J, Lyu Y, Liu X, et al. Engineered probiotics. Microb Cell Fact. 2022;21:72. doi:10.1186/s12934-022-01799-0
32. Stephens RW, Arhire L, Covasa M. Gut microbiota: from microorganisms to metabolic organ influencing obesity. Obesity. 2018;26:801–809. doi:10.1002/oby.22179
33. Pascale A, Marchesi N, Marelli C, et al. Microbiota and metabolic diseases. Endocrine. 2018;61:357–371. doi:10.1007/s12020-018-1605-5
34. Fan Y, Pedersen O. Gut microbiota in human metabolic health and disease. Nat Rev Microbiol. 2021;19:55–71. doi:10.1038/s41579-020-0433-9
35. Del Chierico F, Rapini N, Deodati A, Matteoli MC, Cianfarani S, Putignani L. Pathophysiology of type 1 diabetes and gut microbiota role. Int J Mol Sci. 2022;23:14650. doi:10.3390/ijms232314650
36. Iatcu CO, Steen A, Covasa M. Gut microbiota and complications of type-2 diabetes. Nutrients. 2021;14:166. doi:10.3390/nu14010166
37. Le Chatelier E, Nielsen T, Qin J, et al. Richness of human gut microbiome correlates with metabolic markers. Nature. 2013;500:541–546. doi:10.1038/nature12506
38. Takiishi T, Fenero CIM, Câmara NOS. Intestinal barrier and gut microbiota: shaping our immune responses throughout life. Tissue Barriers. 2017;5. doi:10.1080/21688370.2017.1373208
39. Zhou H, Sun L, Zhang S, Zhao X, Gang X, Wang G. Evaluating the causal role of gut microbiota in type 1 diabetes and its possible pathogenic mechanisms. Front Endocrinol. 2020;11:125. doi:10.3389/fendo.2020.00125
40. Ma Q, Li Y, Li P, et al. Research progress in the relationship between type 2 diabetes mellitus and intestinal flora. Biomed Pharmacother. 2019;117:109138. doi:10.1016/j.biopha.2019.109138
41. Thoo L, Noti M, Krebs P. Keep calm: the intestinal barrier at the interface of peace and war. Cell Death Dis. 2019;10:849. doi:10.1038/s41419-019-2086-z
42. Xu W, Ishii Y, Rini DM, Yamamoto Y, Suzuki T. Microbial metabolite n-butyrate upregulates intestinal claudin-23 expression through SP1 and AMPK pathways in mouse colon and human intestinal Caco-2 cells. Life Sci. 2023;329:121952. doi:10.1016/j.lfs.2023.121952
43. Liang L, Liu L, Zhou W, et al. Gut microbiota-derived butyrate regulates gut mucus barrier repair by activating the macrophage/WNT/ERK signaling pathway. Clin Sci. 2022;136:291–307. doi:10.1042/CS20210778
44. Mokhtari P, Metos J, Anandh Babu PV. Impact of type 1 diabetes on the composition and functional potential of gut microbiome in children and adolescents: possible mechanisms, current knowledge, and challenges. Gut Microbes. 2021;13:1–18. doi:10.1080/19490976.2021.1926841
45. Louis P, Flint HJ. Formation of propionate and butyrate by the human colonic microbiota. Environ Microbiol. 2017;19:29–41. doi:10.1111/1462-2920.13589
46. Wood Heickman LK, DeBoer MD, Fasano A. Zonulin as a potential putative biomarker of risk for shared type 1 diabetes and celiac disease autoimmunity. Diabetes Metab Res Rev. 2020;36:e3309. doi:10.1002/dmrr.3309
47. Crudele L, Piccinin E, Moschetta A. Visceral adiposity and cancer: role in pathogenesis and prognosis. Nutrients. 2021;13. doi:10.3390/nu13062101
48. Bolognini D, Dedeo D, Milligan G. Metabolic and inflammatory functions of short-chain fatty acid receptors. Curr Opin Endocr Metab Res. 2021;16:1–9. doi:10.1016/j.coemr.2020.06.005
49. Morrison DJ, Preston T. Formation of short chain fatty acids by the gut microbiota and their impact on human metabolism. Gut Microbes. 2016;7:189–200. doi:10.1080/19490976.2015.1134082
50. Canfora EE, Meex RCR, Venema K, Blaak EE. Gut microbial metabolites in obesity, NAFLD and T2DM. Nat Rev Endocrinol. 2019;15:261–273. doi:10.1038/s41574-019-0156-z
51. Rosell-Mases E, Santiago A, Corral-Pujol M, et al. Mutual modulation of gut microbiota and the immune system in type 1 diabetes models. Nat Commun. 2023:14. doi:10.1038/s41467-023-43652-x
52. Simon M-C, Reinbeck AL, Wessel C, et al. Distinct alterations of gut morphology and microbiota characterize accelerated diabetes onset in nonobese diabetic mice. J Biol Chem. 2020;295:969–980. doi:10.1074/jbc.RA119.010816
53. Fitzgerald KA, Kagan JC. Toll-like receptors and the control of immunity. Cell. 2020;180. doi:10.1016/j.cell.2020.02.041
54. Alkanani AK, Hara N, Lien E, et al. Induction of diabetes in the RIP-B7.1 mouse model is critically dependent on TLR3 and MyD88 pathways and is associated with alterations in the intestinal microbiome. Diabetes. 2014;63:619–631. doi:10.2337/db13-1007
55. Gianchecchi E, Fierabracci A. Gene/environment interactions in the pathogenesis of autoimmunity: new insights on the role of Toll-like receptors. Autoimmunity Rev. 2015;14:971–983. doi:10.1016/j.autrev.2015.07.006
56. Burrows MP, Volchkov P, Kobayashi KS, Chervonsky AV. Microbiota regulates type 1 diabetes through toll-like receptors. Proc Natl Acad Sci USA. 2015;112:9973–9977. doi:10.1073/pnas.1508740112
57. Cole DK, Bulek AM, Dolton G, et al. Hotspot autoimmune T cell receptor binding underlies pathogen and insulin peptide cross-reactivity. J Clin Invest. 2016;126:2191–2204. doi:10.1172/JCI85679
58. Girdhar K, Huang Q, Chow I-T, et al. A gut microbial peptide and molecular mimicry in the pathogenesis of type 1 diabetes. Proc Natl Acad Sci U S A. 2022;119:e2120028119. doi:10.1073/pnas.2120028119
59. Tai N, Peng J, Liu F, et al. Microbial antigen mimics activate diabetogenic CD8 T cells in NOD mice. J Exp Med. 2016;213. doi:10.1084/jem.20160526
60. Moravejolahkami AR, Shakibaei M, Fairley AM, Sharma M. Probiotics, prebiotics, and synbiotics in type 1 diabetes mellitus: a systematic review and meta-analysis of clinical trials. Diabetes Metab Res Rev. 2024;40:e3655. doi:10.1002/dmrr.3655
61. AkbariRad M, Shariatmaghani SS, Razavi BM, et al. Probiotics for glycemic and lipid profile control of the pre-diabetic patients: a randomized, double-blinded, placebo-controlled clinical trial study. Diabetol Metab Syndr. 2023;15:71. doi:10.1186/s13098-023-01050-9
62. Heilbronner S, Krismer B, Brötz-Oesterhelt H, Peschel A. The microbiome-shaping roles of bacteriocins. Nat Rev Microbiol. 2021;19:726–739. doi:10.1038/s41579-021-00569-w
63. van Zyl WF, Deane SM, Dicks LMT. Molecular insights into probiotic mechanisms of action employed against intestinal pathogenic bacteria. Gut Microbes. 2020;12:1831339. doi:10.1080/19490976.2020.1831339
64. Yu Y, Zong M, Lao L, Wen J, Pan D, Wu Z. Adhesion properties of cell surface proteins in Lactobacillus strains in the GIT environment. Food Funct. 2022;13:3098–3109. doi:10.1039/D1FO04328E
65. van Zyl WF, Deane SM, Dicks LMT. Bacteriocin production and adhesion properties as mechanisms for the anti-listerial activity of Lactobacillus plantarum 423 and Enterococcus mundtii ST4SA. Benef Microbes. 2019;10:329–349. doi:10.3920/BM2018.0141
66. Wan MLY, Forsythe SJ, El-Nezami H. Probiotics interaction with foodborne pathogens: a potential alternative to antibiotics and future challenges. Crit Rev Food Sci Nutr. 2019;59:3320–3333. doi:10.1080/10408398.2018.1490885
67. Ji Q-Y, Wang W, Yan H, et al. The effect of different organic acids and their combination on the cell barrier and biofilm of Escherichia coli. Foods. 2023;12:3011. doi:10.3390/foods12163011
68. Prudêncio CV, Dos Santos MT, Vanetti MCD. Strategies for the use of bacteriocins in Gram-negative bacteria: relevance in food microbiology. J Food Sci Technol. 2015;52:5408–5417. doi:10.1007/s13197-014-1666-2
69. Ashraf R, Shah NP. Immune system stimulation by probiotic microorganisms. Crit Rev Food Sci Nutr. 2014;54:938–956. doi:10.1080/10408398.2011.619671
70. Wan LYM, Chen ZJ, Shah NP, El-Nezami H. Modulation of intestinal epithelial defense responses by probiotic bacteria. Crit Rev Food Sci Nutr. 2016;56:2628–2641. doi:10.1080/10408398.2014.905450
71. McMurdie PJ, Stoeva MK, Justice N, et al. Increased circulating butyrate and ursodeoxycholate during probiotic intervention in humans with type 2 diabetes. BMC Microbiol. 2022;22:19. doi:10.1186/s12866-021-02415-8
72. Mujagic Z, de Vos P, Boekschoten MV, et al. The effects of Lactobacillus plantarum on small intestinal barrier function and mucosal gene transcription; a randomized double-blind placebo controlled trial. Sci Rep. 2017;7:40128. doi:10.1038/srep40128
73. Zhang W, Zhu Y-H, Yang J-C, Yang G-Y, Zhou D, Wang J-F. A selected Lactobacillus rhamnosus strain promotes EGFR-independent Akt activation in an Enterotoxigenic Escherichia coli K88-infected IPEC-J2 cell model. PLoS One. 2015;10:e0125717. doi:10.1371/journal.pone.0125717
74. Yao K, Zeng L, He Q, Wang W, Lei J, Zou X. Effect of probiotics on glucose and lipid metabolism in type 2 diabetes mellitus: a meta-analysis of 12 randomized controlled trials. Med Sci Monit. 2017;23:3044–3053. doi:10.12659/MSM.902600
75. Markowiak-Kopeć P, Śliżewska K. The effect of probiotics on the production of short-chain fatty acids by human intestinal microbiome. Nutrients. 2020;12:1107. doi:10.3390/nu12041107
76. Tonucci LB, Santos KMOD, Ferreira CLDLF, Ribeiro SMR, Oliveira LLD, Martino HSD. Gut microbiota and probiotics: focus on diabetes mellitus. Crit Rev Food Sci Nutr. 2017;57: 2296–2309.
77. Yang S, Lian G. ROS and diseases: role in metabolism and energy supply. Mol Cell Biochem. 2020;467:1–12. doi:10.1007/s11010-019-03667-9
78. Mishra V, Shah C, Mokashe N, Chavan R, Yadav H, Prajapati J. Probiotics as potential antioxidants: a systematic review. J Agric Food Chem. 2015;63:3615–3626. doi:10.1021/jf506326t
79. Kumari VBC, Huligere SS, Alotaibi G, et al. Antidiabetic activity of potential probiotics Limosilactobacillus spp. Levilactobacillus spp. and Lacticaseibacillus spp. isolated from fermented sugarcane juice: a comprehensive in vitro and in silico study. Nutrients. 2023;15:1882. doi:10.3390/nu15081882
80. Nie Y, Zhou H, Wang J, Kan H. Association between systemic immune-inflammation index and diabetes: a population-based study from the NHANES. Front Endocrinol. 2023;14:1245199. doi:10.3389/fendo.2023.1245199
81. Hotamisligil GS. Foundations of immunometabolism and implications for metabolic health and disease. Immunity. 2017;47:406. doi:10.1016/j.immuni.2017.08.009
82. Kaur H, Ali SA. Probiotics and gut microbiota: mechanistic insights into gut immune homeostasis through TLR pathway regulation. Food Funct. 2022;13:7423–7447. doi:10.1039/d2fo00911k
83. Cristofori F, Dargenio VN, Dargenio C, Miniello VL, Barone M, Francavilla R. Anti-inflammatory and immunomodulatory effects of probiotics in gut inflammation: a door to the body. Front Immunol. 2021;12. doi:10.3389/fimmu.2021.578386
84. Yan F, Li N, Shi J, et al. Lactobacillus acidophilus alleviates type 2 diabetes by regulating hepatic glucose, lipid metabolism and gut microbiota in mice. Food Funct. 2019;10:5804–5815. doi:10.1039/C9FO01062A
85. Wang G, Li X, Zhao J, Zhang H, Chen W. Lactobacillus casei CCFM419 attenuates type 2 diabetes via a gut microbiota dependent mechanism. Food Funct. 2017;8:3155–3164. doi:10.1039/C7FO00593H
86. Castillo NA, Perdigón G, de Moreno de Leblanc A. Oral administration of a probiotic Lactobacillus modulates cytokine production and TLR expression improving the immune response against Salmonella enterica serovar Typhimurium infection in mice. BMC Microbiol. 2011;11:177. doi:10.1186/1471-2180-11-177
87. Thakur BK, Saha P, Banik G, et al. Live and heat-killed probiotic Lactobacillus casei Lbs2 protects from experimental colitis through toll-like receptor 2-dependent induction of T-regulatory response. Int Immunopharmacol. 2016;36:39–50. doi:10.1016/j.intimp.2016.03.033
88. Lee C, Kim SW, Verma R, et al. Probiotic consortium confers synergistic anti-inflammatory effects in inflammatory disorders. Nutrients. 2024;16:790. doi:10.3390/nu16060790
89. Peng Y, Ma Y, Luo Z, Jiang Y, Xu Z, Yu R. Lactobacillus reuteri in digestive system diseases: focus on clinical trials and mechanisms. Front Cell Infect Microbiol. 2023;13:1254198. doi:10.3389/fcimb.2023.1254198
90. Nemati M, Ebrahimi B, Montazeri-Najafabady N. Probiotics ameliorate endocrine disorders via modulating inflammatory pathways: a systematic review. Genes Nutr. 2024;19. doi:10.1186/s12263-024-00743-8
91. Kang H-J, Im S-H. Probiotics as an immune modulator. J Nutr Sci Vitaminol. 2015;61:S103–5. doi:10.3177/jnsv.61.S103
92. Mishra S, Wang S, Nagpal R, et al. Probiotics and prebiotics for the amelioration of type 1 diabetes: present and future perspectives. Microorganisms. 2019;7:67. doi:10.3390/microorganisms7030067
93. Wang C-H, Yen H-R, Lu W-L, et al. Adjuvant probiotics of Lactobacillus salivarius subsp. salicinius AP-32, L. johnsonii MH-68, and Bifidobacterium animalis subsp. lactis CP-9 Attenuate glycemic levels and inflammatory cytokines in patients With type 1 diabetes mellitus. Front Endocrinol. 2022;13:754401. doi:10.3389/fendo.2022.754401
94. Wei S-H, Chen Y-P, Chen M-J. Selecting probiotics with the abilities of enhancing GLP-1 to mitigate the progression of type 1 diabetes in vitro and in vivo. J Funct Foods. 2015;18:473–486. doi:10.1016/j.jff.2015.08.016
95. Mariño E, Richards JL, McLeod KH, et al. Gut microbial metabolites limit the frequency of autoimmune T cells and protect against type 1 diabetes. Nat Immunol. 2017;18:552–562. doi:10.1038/ni.3713
96. Chen Y, Shen X, Ma T, et al. Adjunctive Probio-X treatment enhances the therapeutic effect of a conventional drug in managing type 2 diabetes mellitus by promoting short-chain fatty acid-producing bacteria and bile acid pathways. mSystems. 2023;8:e0130022. doi:10.1128/msystems.01300-22
97. Han S, Lu Y, Xie J, et al. Probiotic gastrointestinal transit and colonization after oral administration: a long journey. Front Cell Infect Microbiol. 2021;11:609722. doi:10.3389/fcimb.2021.609722
98. Arriaga-Morales JJ, Ordaz-Pichardo C, Castro-Muñoz R, Durán-Páramo E. Attenuation of hyperglycemia in diabetic rats assisted by immobilized probiotic in sodium alginate. Probiotics Antimicrob Proteins. 2023. doi:10.1007/s12602-023-10166-3
99. Robert S, Steidler L. Recombinant Lactococcus lactis can make the difference in antigen-specific immune tolerance induction, the type 1 diabetes case. Microb Cell Fact. 2014;
100. Xu C, Ban Q, Wang W, Hou J, Jiang Z. Novel nano-encapsulated probiotic agents: encapsulate materials, delivery, and encapsulation systems. J Control Release. 2022;349:184–205. doi:10.1016/j.jconrel.2022.06.061
101. Han M, Lei W, Liang J, Li H, Hou M, Gao Z. The single-cell modification strategies for probiotics delivery in inflammatory bowel disease: a review. Carbohydr Polym. 2024;324:121472. doi:10.1016/j.carbpol.2023.121472
102. Gómez-Fernández AR, Faccinetto-Beltrán P, Orozco-Sánchez NE, et al. Sugar-free milk chocolate as a carrier of Omega-3 polyunsaturated fatty acids and probiotics: a potential functional food for the diabetic population. Foods. 2021;10:1866. doi:10.3390/foods10081866
103. Khursheed R, Singh SK, Kumar B, et al. Self-nanoemulsifying composition containing curcumin, quercetin, Ganoderma lucidum extract powder and probiotics for effective treatment of type 2 diabetes mellitus in streptozotocin induced rats. Int J Pharm. 2022;612:121306. doi:10.1016/j.ijpharm.2021.121306
104. Wang Y, Shi Y, Peng X, et al. Biochemotaxis-oriented engineering bacteria expressing GLP-1 enhance diabetes therapy by regulating the balance of immune. Adv Healthcare Mater. 2024;13:2303958. doi:10.1002/adhm.202303958
105. Guimarães D, Cavaco-Paulo A, Nogueira E. Design of liposomes as drug delivery system for therapeutic applications. Int J Pharm. 2021;601:120571. doi:10.1016/j.ijpharm.2021.120571
106. Ma Y, Fei Y, Ding S, Jiang H, Fang J, Liu G. Trace metal elements: a bridge between host and intestinal microorganisms. Sci China Life Sci. 2023;66:1976–1993. doi:10.1007/s11427-022-2359-4
107. Cooper-Capetini V, de Vasconcelos DAA, Martins AR, et al. Zinc supplementation improves glucose homeostasis in high fat-fed mice by enhancing pancreatic β-cell function. Nutrients. 2017;9:1150. doi:10.3390/nu9101150
108. Fernández-Cao JC, Warthon-Medina M, Moran VH, et al. Zinc intake and status and risk of type 2 diabetes mellitus: a systematic review and meta-analysis. Nutrients. 2019:11. doi:10.3390/nu11051027
109. Suksomboon N, Poolsup N, Yuwanakorn A. Systematic review and meta-analysis of the efficacy and safety of chromium supplementation in diabetes. J Clin Pharm Ther. 2014;39:292–306. doi:10.1111/jcpt.12147
110. Zhao D, Zhu H, Gao F, et al. Antidiabetic effects of selenium-enriched Bifidobacterium longum DD98 in type 2 diabetes model of mice. Food Funct. 2020;11:6528–6541. doi:10.1039/D0FO00180E
111. Lin Y, Ren Y, Zhang Y, et al. Protective role of nano-selenium-enriched Bifidobacterium longum in delaying the onset of streptozotocin-induced diabetes. R Soc Open Sci. 2018;5:181156. doi:10.1098/rsos.181156
112. Zhang F, Li X, Wei Y. Selenium and selenoproteins in health. Biomolecules. 2023;13:799. doi:10.3390/biom13050799
113. Prabhu KS, Lei XG. Selenium12. Adv Nutr. 2016;7:415–417. doi:10.3945/an.115.010785
114. Qi Z, Duan A, Ng K. Selenoproteins in Health. Molecules. 2023;29:136. doi:10.3390/molecules29010136
115. Falcetta P, Aragona M, Bertolotto A, et al. Insulin discovery: a pivotal point in medical history. Metabolism. 2022;127:154941. doi:10.1016/j.metabol.2021.154941
116. Akbari V, Hendijani F, Feizi A, et al. Efficacy and safety of oral insulin compared to subcutaneous insulin: a systematic review and meta-analysis. J Endocrinol Invest. 2016;39:215–225. doi:10.1007/s40618-015-0326-3
117. Miranda C, Contente D, Igrejas G, Câmara SPA, Dapkevicius MDLE, Poeta P. Role of exposure to lactic acid bacteria from foods of animal origin in human health. Foods. 2021;10:2092. doi:10.3390/foods10092092
118. Kok J, van Gijtenbeek LA, de Jong A, van der Meulen SB, Solopova A, Kuipers OP. The Evolution of gene regulation research in Lactococcus lactis. FEMS Microbiol Rev. 2017;41:S220–43. doi:10.1093/femsre/fux028
119. Ng DTW, Sarkar CA. Nisin-inducible secretion of a biologically active single-chain insulin analog by Lactococcus lactis NZ9000. Biotechnol Bioeng. 2011;108:1987–1996. doi:10.1002/bit.23130
120. Mao R, Wu D, Hu S, Zhou K, Wang M, Wang Y. Secretory expression and surface display of a new and biologically active single-chain insulin (SCI-59) analog by lactic acid bacteria. Appl Microbiol Biotechnol. 2017;101:3259–3271. doi:10.1007/s00253-017-8125-8
121. Mao R, Chen Y, Wu Q, et al. Oral delivery of single-chain insulin (SCI-59) analog by bacterium-like particles (BLPs) induces oral tolerance and prevents autoimmune diabetes in NOD mice. Immunol Lett. 2019;214:37–44. doi:10.1016/j.imlet.2019.08.008
122. Drucker DJ. Mechanisms of action and therapeutic application of glucagon-like peptide-1. Cell Metab. 2018;27:740–756. doi:10.1016/j.cmet.2018.03.001
123. Hu H, Luo J, Liu Y, et al. Improvement effect of a next-generation probiotic L. plantarum-pMG36e-GLP-1 on type 2 diabetes mellitus via the gut–pancreas–liver axis. Food Funct. 2023;14:3179–3195. doi:10.1039/D3FO00044C
124. Johnson LM, Barrick S, Hager MV, et al. A potent α/β-peptide analogue of GLP-1 with prolonged action in vivo. J Am Chem Soc. 2014;136:12848. doi:10.1021/ja507168t
125. Wang Q, Guo H, Mao W, Qian X, Liu Y. The oral delivery system of modified GLP-1 by probiotics for T2DM. Pharmaceutics. 2023;15:1202. doi:10.3390/pharmaceutics15041202
126. Hou J, Yan R, Ding D, et al. Oral administration of a fusion protein containing eight GLP-1 analogues produced in Escherichia coli BL21(DE3) in streptozotocin-induced diabetic rats. Biotechnol Lett. 2007;29:1439–1446. doi:10.1007/s10529-007-9427-1
127. Ma B, Tu P, Zhao X, et al. Expression and purification of optimized rolGLP-1, a novel GLP-1 analog, in Escherichia coli BL21(DE3) and its good glucoregulatory effect on type 2 diabetic mice. Curr Pharm Biotechnol. 2013;14:985–994. doi:10.2174/1389201014666131226155553
128. Xu F, Wang KY, Wang N, Li G, Liu D. Modified human glucagon-like peptide-1 (GLP-1) produced in E. coli has a long-acting therapeutic effect in type 2 diabetic mice. PLoS One. 2017;12:e0181939. doi:10.1371/journal.pone.0181939
129. Lin Y, Krogh-Andersen K, Pelletier J, Marcotte H, Östenson C-G, Hammarström L. Oral delivery of pentameric glucagon-like peptide-1 by recombinant Lactobacillus in diabetic rats. PLoS One. 2016;11:e0162733. doi:10.1371/journal.pone.0162733
130. Wu R, Ma C, Li X-D, et al. Construction of yeast strains expressing long-acting glucagon-like peptide-1 (GLP-1) and their therapeutic effects on type 2 diabetes mellitus mouse model. Yi Chuan. 2015;37:183–191. doi:10.16288/j.yczz.14-206
131. Zhao X, Wang M, Wen Z, et al. GLP-1 receptor agonists: beyond their pancreatic effects. Front Endocrinol. 2021;12:721135. doi:10.3389/fendo.2021.721135
132. Gentilella R, Pechtner V, Corcos A, Consoli A. Glucagon-like peptide-1 receptor agonists in type 2 diabetes treatment: are they all the same? Diabetes/Metab Res Rev. 2019;35:e3070. doi:10.1002/dmrr.3070
133. Heimbürger SM, Brønden A, Johansen NJ, Dejgaard TF, Vilsbøll T, Knop FK. The efficacy and safety of exenatide once weekly in patients with type 2 diabetes. Expert Opin Pharmacother. 2019;20:501–510. doi:10.1080/14656566.2019.1571040
134. Zeng Z, Yu R, Zuo F, et al. Heterologous expression and delivery of biologically active Exendin-4 by Lactobacillus paracasei L14. PLoS One. 2016;11:e0165130. doi:10.1371/journal.pone.0165130
135. Zeng Z, Yu R, Zuo F, Zhang B, Ma H, Chen S. Recombinant Lactococcus lactis expressing bioactive exendin-4 to promote insulin secretion and beta-cell proliferation in vitro. Appl Microbiol Biotechnol. 2017;101:7177–7186. doi:10.1007/s00253-017-8410-6
136. Ni Q, Pham NB, Meng WS, Zhu G, Chen X. Advances in immunotherapy of type I diabetes. Adv Drug Delivery Rev. 2019;139:83–91. doi:10.1016/j.addr.2018.12.003
137. Liang J, Aihua Z, Yu W, Yong L, Jingjing L. HSP65 serves as an immunogenic carrier for a diabetogenic peptide P277 inducing anti-inflammatory immune response in NOD mice by nasal administration. Vaccine. 2010;28:3312–3317. doi:10.1016/j.vaccine.2010.02.100
138. Ma Y, Liu J, Hou J, et al. Oral administration of recombinant Lactococcus lactis expressing HSP65 and tandemly repeated P277 reduces the incidence of type I diabetes in non-obese diabetic mice. PLoS One. 2014;9:e105701. doi:10.1371/journal.pone.0105701
139. Liu K-F, Liu X-R, Li G-L, Lu S-P, Jin L, Wu J. Oral administration of Lactococcus lactis-expressing heat shock protein 65 and tandemly repeated IA2P2 prevents type 1 diabetes in NOD mice. Immunol Lett. 2016;174:28–36. doi:10.1016/j.imlet.2016.04.008
140. Maddaloni M, Kochetkova I, Jun S, Callis G, Thornburg T, Pascual DW. Milk-based nutraceutical for treating autoimmune arthritis via the stimulation of IL-10- and TGF-β-producing CD39+ regulatory T cells. PLoS One. 2015;10:e0117825. doi:10.1371/journal.pone.0117825
141. Nelson AS, Maddaloni M, Abbott JR, et al. Oral therapy with colonization factor antigen I prevents development of type 1 diabetes in non-obese diabetic mice. Sci Rep. 2020;10:6156. doi:10.1038/s41598-020-62881-4
142. Nelson AS, Akgul A, Maddaloni M, Bhagyaraj E, Hoffman C, Pascual DW. Oral probiotic promotes indoleamine 2,3-dioxygenase- and TGF-β–producing plasmacytoid dendritic cells to initiate protection against type 1 diabetes. Immunol Lett. 2021;239:12–19. doi:10.1016/j.imlet.2021.07.009
143. Zhang T, Zhang J, Duan L. The role of genetically engineered probiotics for treatment of inflammatory bowel disease: a systematic review. Nutrients. 2023;15:1566. doi:10.3390/nu15071566
144. Preisser TM, da Cunha VP, Santana MP, et al. Recombinant Lactococcus lactis carrying IL-4 and IL-10 coding vectors protects against type 1 diabetes in NOD mice and attenuates insulitis in the STZ-induced model. J Diabetes Res. 2021;2021:6697319. doi:10.1155/2021/6697319
145. Campos GM, Américo MF, Dos Santos Freitas A, et al. Lactococcus lactis as an interleukin delivery system for prophylaxis and treatment of inflammatory and autoimmune diseases. Probiotics Antimicrob Proteins. 2023;16:1–15. doi:10.1007/s12602-023-10041-1
146. Takiishi T, Korf H, Van Belle TL, et al. Reversal of autoimmune diabetes by restoration of antigen-specific tolerance using genetically modified Lactococcus lactis in mice. J Clin Invest. 2012;122:1717–1725. doi:10.1172/JCI60530
147. Cook DP, Cunha JPMCM, Martens P-J, et al. Intestinal delivery of proinsulin and IL-10 via Lactococcus lactis combined with low-dose anti-CD3 restores tolerance outside the window of acute type 1 diabetes diagnosis. Front Immunol. 2020;11:1103. doi:10.3389/fimmu.2020.01103
148. Takiishi T, Cook DP, Korf H, et al. Reversal of diabetes in NOD mice by clinical-grade proinsulin and IL-10–secreting Lactococcus lactis in combination with low-dose anti-CD3 depends on the induction of Foxp3-Positive T cells. Diabetes. 2016;66:448–459. doi:10.2337/db15-1625
149. Robert S, Gysemans C, Takiishi T, et al. Oral delivery of glutamic acid decarboxylase (GAD)-65 and IL10 by Lactococcus lactis reverses diabetes in recent-onset NOD mice. Diabetes. 2014;63:2876–2887. doi:10.2337/db13-1236
150. Lang J, Wang X, Liu K, et al. Oral delivery of staphylococcal nuclease by Lactococcus lactis prevents type 1 diabetes mellitus in NOD mice. Appl Microbiol Biotechnol. 2017;101:7653–7662. doi:10.1007/s00253-017-8480-5
151. Rhimi M, Bermudez-Humaran LG, Huang Y, et al. The secreted l-arabinose isomerase displays anti-hyperglycemic effects in mice. Microb Cell Fact. 2015;14:204. doi:10.1186/s12934-015-0391-5
152. Shen H, Zhao Z, Zhao Z, Chen Y, Zhang L. Native and engineered probiotics: promising agents against related systemic and intestinal diseases. IJMS. 2022;23:594. doi:10.3390/ijms23020594
153. Zepeda-Hernández A, Garcia-Amezquita LE, Requena T, García-Cayuela T. Probiotics, prebiotics, and synbiotics added to dairy products: uses and applications to manage type 2 diabetes. Food Res Int. 2021;142:110208. doi:10.1016/j.foodres.2021.110208
154. Meng J, Liu S, Wu X. Engineered probiotics as live biotherapeutics for diagnosis and treatment of human diseases. Crit Rev Microbiol. 2024;50:300–314. doi:10.1080/1040841X.2023.2190392
155. Kwoji ID, Aiyegoro OA, Okpeku M, Adeleke MA. Multi-strain probiotics: synergy among isolates enhances biological activities. Biology. 2021;10:322. doi:10.3390/biology10040322
156. Bouhenni H. Study of combined effect of some medicinal plants and probiotics against Helicobacter pylori responsible for gastroduodenal diseases [Thesis]. Faculty of Nature and Life Sciences; 2023.
157. Ali HA, Mohamed SH, Alharbi HF, Algheshairy RM. Synergism between probiotics and herbs to manage type 2 diabetes in rats. Int J Pharm Pharm Sci. 2020;12:26–35. doi:10.22159/ijpps.2020v12i5.37008
158. Paquette S, Thomas SC, Venkataraman K, Appanna VD, Tharmalingam S. the effects of oral probiotics on type 2 diabetes mellitus (T2DM): a clinical trial systematic literature review. Nutrients. 2023;15:4690. doi:10.3390/nu15214690
159. Verma A, Xu K, Du T, et al. Expression of human ACE2 in lactobacillus and beneficial effects in diabetic retinopathy in mice. Mol Ther Methods Clin Dev. 2019;14:161–170. doi:10.1016/j.omtm.2019.06.007
160. Barati M, Jabbari M, Abdi Ghavidel A, et al. The engineered probiotics for the treatment of chronic diseases: a systematic review. J Food Biochem. 2022:46. doi:10.1111/jfbc.14343
161. Chen H, Lei P, Ji H, et al. Advances in Escherichia coli Nissle 1917 as A customizable drug delivery system for disease treatment and diagnosis strategies. Mater Today Bio. 2023;18:100543. doi:10.1016/j.mtbio.2023.100543
162. Gupta R, Raghuwanshi S. Designer probiotics in metabolic disorders. In: Chopra K, Bishnoi M, Kondepudi KK, editors. Probiotic Research in Therapeutics: Volume 5: Metabolic Diseases and Gut Bacteria. Singapore: Springer Nature Singapore; 2022:241–260.https://doi.org/10.1007/978-981-16-8444-9_12.
163. Qu L, Liu F, Fang Y, et al. Improvement in Zebrafish with diabetes and alzheimer’s disease treated with pasteurized Akkermansia muciniphila. Microbiol Spectr. 2023;11:e00849–23. doi:10.1128/spectrum.00849-23
164. Baroni I, Fabrizi D, Luciani M, et al. Probiotics and synbiotics for glycemic control in diabetes: a systematic review and meta-analysis of randomized controlled trials. Clin Nutr. 2024;43:1041–1061. doi:10.1016/j.clnu.2024.03.006
165. Bruetschy C. The EU regulatory framework on genetically modified organisms (GMOs). Transgenic Res. 2019;28:169–174. doi:10.1007/s11248-019-00149-y
 © 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms.php
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms.php
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Effects of Different Carbohydrate Content Diet on Gut Microbiota and Aortic Calcification in Diabetic Mice

Shen X, Guo G, Feng G, Wang Z
Diabetes, Metabolic Syndrome and Obesity 2024, 17:2327-2346
Published Date: 10 June 2024