")
Back to Journals » Infection and Drug Resistance » Volume 18
Escherichia coli ST542 Co-Harbouring blaNDM-1 and mcr-1 in a Pig Farm, China
Authors Shao PY, Jiang Y, Mei CY, Li W , Wang ZY, Jiao X , Wang J, Sun L
Received 10 December 2024
Accepted for publication 15 February 2025
Published 26 February 2025 Volume 2025:18 Pages 1183—1186
DOI https://doi.org/10.2147/IDR.S508552
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Prof. Dr. Héctor Mora-Montes
Peng-Yun Shao,1,2,* Yue Jiang,1,2,* Cai-Yue Mei,1,2 Wenyi Li,3 Zhen-Yu Wang,1,2 Xinan Jiao,1,2 Jing Wang,1,2 Lin Sun1,2
1Jiangsu Key Laboratory of Zoonosis/Jiangsu Co-Innovation Center for Prevention and Control of Important Animal Infectious Diseases and Zoonoses, Yangzhou University, Yangzhou, 225009, People’s Republic of China; 2Key Laboratory of Prevention and Control of Biological Hazard Factors (Animal Origin) for Agrifood Safety and Quality, Ministry of Agriculture of China, Yangzhou University, Yangzhou, 225009, People’s Republic of China; 3Department of Biochemistry and Chemistry, La Trobe Institute for Molecular Science, La Trobe University, Melbourne, Victoria, 3086, Australia
*These authors contributed equally to this work
Correspondence: Jing Wang; Lin Sun, Jiangsu Key Laboratory of Zoonosis/Jiangsu Co-Innovation Center for Prevention and Control of Important Animal Infectious Diseases and Zoonoses, Yangzhou University, Yangzhou, 225009, People’s Republic of China, Email [email protected]; [email protected]
Carbapenems are potent antibiotics utilized to treat serious infections caused by multidrug-resistant (MDR) Gram-negative bacteria.1 However, the clinical use of carbapenems has led to the emergence and global dissemination of carbapenem-resistant Enterobacteriaceae (CRE), which pose a significant threat to effective clinical therapy.1 The genes encoding carbapenemases, particularly the New Delhi metallo-β-lactamase gene blaNDM, have been widely distributed among humans and detected in animals, food, and the environment.1,2 Colistin is considered as the last therapeutic resort for treating MDR Gram-negative bacteria, especially CRE bacteria.1,3 However, its efficacy has been undermined by the emergence of various antimicrobial resistance mechanisms, such as mutations in the two-component regulatory systems PmrA-PmrB and PhoP-PhoQ.3 Particularly, the global spread of plasmid-mediated colistin resistance gene mcr, first identified in 2015, has further exacerbated this issue.3 Here, we report the recent identification of Escherichia coli ST542 strains co-producing MCR-1 and NDM-1 isolated from a pig farm in Yangzhou, China.
On 17 May 2022, a total of 126 samples were collected from a pig farm in Yangzhou, China, including pig feces (n = 93), feed (n = 3), pig drinking water (n = 5), and floor swabs from pens (n = 25). A single sample was collected per pig, with no more than five samples taken from pigs in the same house. The samples were incubated in buffered peptone water (BPW) broth for 18–24 hours and subsequently inoculated onto MacConkey agar. One suspected E. coli isolate was selected from each sample and confirmed using matrix-assisted laser desorption/ionization time-of-flight mass spectrometry. In total, 107 E. coli isolates were obtained, among which three (2.80%) isolates, namely YZ22PE67, YZ22PE68 and YZ22PE106 from pig feces co-carried the blaNDM-1 and mcr-1 genes, confirmed by PCR and sequencing as previously described.3,4 Minimum inhibitory concentrations (MICs) of 16 antimicrobial agents were determined using the microdilution method (cation-adjusted Mueller–Hinton broth) or the agar dilution method (Mueller–Hinton agar; limited to fosfomycin). E. coli ATCC 25922 served as the quality control strain. Results were interpreted according to the Clinical and Laboratory Standards Institute (CLSI) M100, 32nd edition, or the European Committee on Antimicrobial Susceptibility Testing (EUCAST; www.eucast.org). As shown in Table 1, three NDM- and MCR-producing E. coli isolates were resistant to meropenem and colistin, as well as to ampicillin, cefazolin, cefotaxime, streptomycin, tetracycline, chloramphenicol, florfenicol, sulfamethoxazole/trimethoprim. However, they remained susceptible to gentamicin, amikacin, tigecycline, nalidixic acid, ciprofloxacin, and fosfomycin (Table S1).
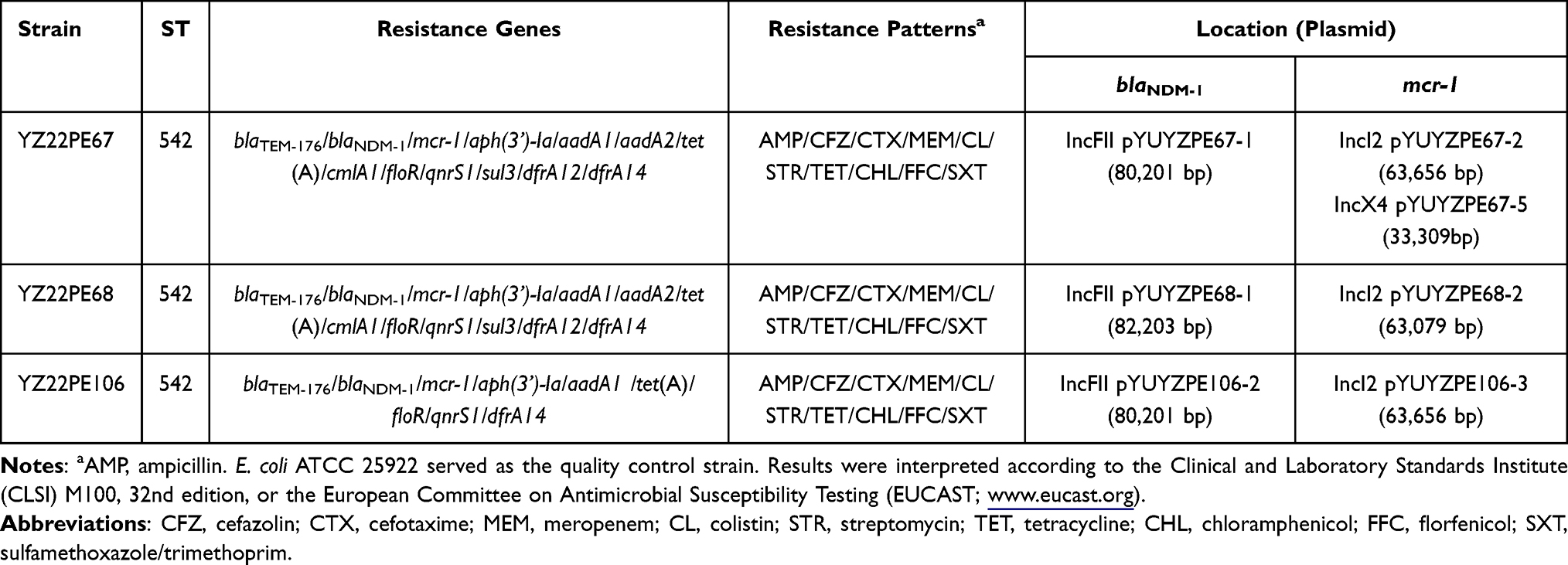 |
Table 1 Characteristics of Mcr-1 and blaNDM-1-Carrying Escherichia coli Isolates in This Study |
To further characterize these E. coli isolates, whole-genome sequencing was performed using Illumina HiSeq technology (Illumina, San Diego, CA, USA). The library was prepared with the NEBNext Ultra II DNA Library Prep Kit for Illumina (New England Biolabs, USA), generating 150 bp paired-end reads. Subsequently, genomic DNA was sequenced on the Oxford Nanopore MinION platform (Oxford Nanopore Technologies, Oxford, UK). The library was constructed using the Ligation Sequencing Kit (SQK-LSK109) and Native Barcoding Expansion (EXP-NBD104/114). Following library preparation, the DNA library was loaded onto the MinION R9.4 flow cells to read the DNA sequence in real-time, generating long-read data. The Illumina short reads and MinION long reads were then subjected to hybrid assembly using Unicycler v0.5.0 to obtain complete genome sequences. The sequences were analyzed using the Center for Genomic Epidemiology (CGE) pipeline (https://www.genomicepidemiology.org/) and BLAST (http://blast.ncbi.nlm.nih.gov/Blast.cgi). All whole-genome sequences have been deposited in GenBank under accession number PRJNA1175595. Three NDM- and MCR-producing E. coli strains belonged to sequence type 542 (ST542). Previously, E. coli ST542 strains carrying carbapenemase gene blaNDM-5 or mcr-1 were reported in patients or animals in China.5,6 In addition to blaNDM-1 and mcr-1, all three isolates harbored the β-lactam resistance genes blaTEM-176, the aminoglycoside resistance genes aph(3’)-Ia and aadA1, the tetracycline resistance gene tet(A), the phenicol exporter gene floR, the quinolone resistance gene qnrS1, and the trimethoprim resistance gene dfrA14. Notably, YZ22PE67 and YZ22PE68 also contained the aminoglycoside resistance genes aadA2, the chloramphenicol-resistant gene cmlA1, the sulfonamide resistance genes sul3 and the trimethoprim resistance gene dfrA12 (Table 1). These findings are consistent with the results of antimicrobial susceptibility testing.
Three NDM- and MCR-producing isolates contained multiple plasmids (Table S2). Specifically, three similar blaNDM-1-carrying plasmids (80,201 or 82,203 bp), designated as pYUYZPE67-1, pYUYZPE68-1, and pYUYZPE106-2 were obtained, respectively (Table S2). These plasmids belonged to IncFII plasmids and were highly similar (>99.9%) in organization to some blaNDM-1-bearing IncFII plasmids from China, such as plasmids pDXLP15_1 (E. coli, OR965474.1, pork), plasmid unnamed1 from Salmonella enterica strain sg1722-2 (CP081190.2, child feces), and plasmid unnamed4 from E. coli strain EC1722-1 (CP081195.1, child feces) (Figure S1). Each of them contained a 9067-bp core blaNDM-1 structure (IS26-tat-trpF-bleMBL-blaNDM-1-ISAba125-ΔIS3000-IS26). Furthermore, three blaNDM-1-carrying plasmids pYUYZPE67-1, pYUYZPE68-1, and pYUYZPE106-2 were successfully transferred to E. coli C600 with frequencies of 2.42 × 10−3, 3.11 × 10−2, and 1.32 × 10−4 transconjugants per recipient, respectively.
The mcr-1 gene was located on Incl2 plasmids in three NDM- and MCR-producing E. coli isolates, with an additional mcr-1-carrying IncX4 plasmid identified in isolate YZ22PE67 (Tables 1 and S2). Three IncI2 plasmids pYUYZPE67-2 (63,656 bp), pYUYZPE68-2 (63,079 bp), and pYUYZPE106-3 (63,656 bp) were similar to other mcr-1-carrying IncI2 plasmids from China, such as pYULNMC7-MCR (E. coli, chicken meat, CP126506) and the first reported mcr-1-carrying plasmid pHNSHP45 (E. coli, pig, KP347127) (Figure S2). Notably, the mobile element ISApl1, which is associated with mcr-1 mobilization, was absent in our mcr-1-bearing IncI2 plasmids (Figure S2A). The mcr-1-bearing IncX4 plasmid pYUYZPE67-5 (33,309 bp) was organized similarly to other mcr-1-carrying IncX4 plasmids from animals or food products, including regions for replication, conjugal transfer, maintenance and stability (Figure S2B). Additionally, one copy of IS26 was inserted into the backbone of pYUYZPE67-5 at the same site as observed in plasmids like pHNSHP10 (E. coli, pig, MF774182) and pFS170G (E. coli, pig, KX711707), generating 8-bp direct repeats (5’-CTGTGTGA-3’) (Figure S2). Both mcr-1-carrying plasmids in YZ22PE67 (pYUYZPE67-2 and pYUYZPE67-5) could be successfully transferred to E. coli C600 at frequencies of 2.68 × 10−3 and 3.05 × 10−3 transconjugants per recipient, respectively. The remaining mcr-1-carrying IncI2 plasmids, pYUYZPE68-2 and pYUYZPE106-3, were transferable at a frequency of 2.71 × 10−3 and 3.34 × 10−6 transconjugants per recipient, respectively.
The coexistence of carbapenemase genes (blaNDM/blaKPC) and mcr has been reported in some Enterobacteriaceae species, such as E. coli and Klebsiella pneumoniae from humans, animals, and food.7–9 The concurrent presence of carbapenemase genes and mcr in Enterobacteriaceae significantly limits treatment options and complicates infection management. Furthermore, the dissemination of such organisms can occur through multiple transmission routes, including the food chain, environmental contamination, and direct contact with animals, thereby posing a serious threat to public health. Consequently, it is essential to implement control measures to reduce and prevent the transmission of such organisms in animal husbandry. Furthermore, exploring alternative strategies, such as bacteriophages and antimicrobial peptides, is critical for preventing and treating infections in both animals and humans, without relying solely on antibiotics.10,11
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This work was supported by grants from the Key Research and Development Program (Modern Agriculture) project of Jiangsu Province (no. BE2021331) and Priority Academic Program Development of Jiangsu Higher Education Institutions (PAPD).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Wu W, Feng Y, Tang G, et al. NDM metallo-β-Lactamases and their bacterial producers in health care settings. Clin Microbiol Rev. 2019;32(2):e00115–18. doi:10.1128/CMR.00115-18
2. Taggar G, Attiq Rheman M, Boerlin P, et al. Molecular epidemiology of carbapenemases in Enterobacteriales from humans, animals, food and the environment. Antibiotics. 2020;9(10):693. doi:10.3390/antibiotics9100693
3. Liu Y-Y, Wang Y, Walsh TR, et al. Emergence of plasmid-mediated colistin resistance mechanism MCR-1 in animals and human beings in China: a microbiological and molecular biological study. Lancet Infect Dis. 2016;16(2):161–168. doi:10.1016/S1473-3099(15)00424-7
4. Poirel L, Walsh TR, Cuvillier V, et al. Multiplex PCR for detection of acquired carbapenemase genes. Diagn Microbiol Infect Dis. 2011;70(1):119–123. doi:10.1016/j.diagmicrobio.2010.12.002
5. Sekizuka T, Inamine Y, Segawa T, et al. Characterization of NDM-5- and CTX-M-55-coproducing Escherichia coli GSH8M-2 isolated from the effluent of a wastewater treatment plant in Tokyo Bay. Infect Drug Resist. 2019;12:2243–2249. doi:10.2147/IDR.S215273
6. Li F, Cheng P, Li X, Liu R, Zhang X, Zhang X. Molecular epidemiology and colistin-resistant mechanism of mcr-positive and mcr-negative Escherichia coli isolated from animal in Sichuan Province, China. Front Microbiol. 2022;13:818548. doi:10.3389/fmicb.2022.818548
7. Guo C-H, Chu M-J, Liu T, et al. High prevalence and transmission of blaNDM-positive Escherichia coli between farmed ducks and slaughtered meats: an increasing threat to food safety. Int J Food Microbiol. 2024;424:110850. doi:10.1016/j.ijfoodmicro.2024.110850
8. Liu R, Chen Y, Xu H, et al. Fusion event mediated by IS903B between chromosome and plasmid in two MCR-9- and KPC-2-co-producing Klebsiella pneumoniae isolates. Drug Resist Updat. 2024;77:101139. doi:10.1016/j.drup.2024.101139
9. Wang X, Wang Y, Zhou Y, et al. Emergence of a novel mobile colistin resistance gene, mcr-8, in NDM-producing Klebsiella pneumoniae. Emerg Microbes Infect. 2018;7(1):122. doi:10.1038/s41426-018-0124-z
10. Gómez-Ochoa SA, Pitton M, Valente LG, et al. Efficacy of phage therapy in preclinical models of bacterial infection: a systematic review and meta-analysis. Lancet Microbe. 2022;3(12):e956–e968. doi:10.1016/S2666-5247(22)00288-9
11. Li W, Separovic F, O’Brien-Simpson NM, et al. Chemically modified and conjugated antimicrobial peptides against superbugs. Chem Soc Rev. 2021;50(8):4932–4973. doi:10.1039/d0cs01026j
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms.php
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms.php
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.