")
Back to Journals » Journal of Pain Research » Volume 18
Evaluation of the Therapeutic Efficacy of Entrapment Neuropathy Unties (ENU) Pharmacopuncture in Neuropathic Pain Caused by Sciatic Nerve Ligation in Mice
Authors Kweon B, Kim DU, Park K, Lee S, Youn Y , Park HJ, Shim HS, Yoo J, Lee YK, Bae GS , Choi Y
Received 3 February 2025
Accepted for publication 8 May 2025
Published 12 May 2025 Volume 2025:18 Pages 2393—2405
DOI https://doi.org/10.2147/JPR.S519298
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor E Alfonso Romero-Sandoval
Bitna Kweon,1,2,* Dong-Uk Kim,1,2,* Kyoungsu Park,3,* Sangho Lee,4 Yousuk Youn,3 Hyeok Ju Park,5 Hyun Soo Shim,6 Junsang Yoo,6 Yong Kyu Lee,5 Gi-Sang Bae,1,2,7 Youngjin Choi8
1Department of Pharmacology, School of Korean Medicine, Wonkwang University, Iksan, 54538, Republic of Korea; 2Hanbang Cardio-Renal Syndrome Research Center, Wonkwang University, Iksan, 54538, Republic of Korea; 3Chamjalham Hospital of Korean Medicine, Suwon Kyunggi-do, 16263, Republic of Korea; 4Chamjalham Hospital of Korean Medicine, Seoul, 05316, Republic of Korea; 5Database Laboratory, Department of Computer Science and Engineering, Dongguk University-Seoul, Seoul, 04620, Republic of Korea; 6Laboratory of Regenerative Medicine for Neurodegenerative Disease, Stand up Therapeutics, Seoul, 04418, Republic of Korea; 7Research Center of Traditional Korean Medicine, Wonkwang University, Iksan, 54538, Republic of Korea; 8Kyunghee Dabok Korean Medicine Clinic, Seoul, 05769, Republic of Korea
*These authors contributed equally to this work
Correspondence: Gi-Sang Bae, Department of Pharmacology, School of Korean Medicine, Wonkwang University, 460, Iksan-daero, Iksan, 54538, Republic of Korea, Tel +82 63 850 6842, Email [email protected] Youngjin Choi, Kyunghee Dabok Korean Medicine Clinic, 404, Ogeum-ro, Seoul, 05769, Republic of Korea, Tel +82 02 431 1600, Email [email protected]
Background: Neuropathic pain caused by peripheral nerve injury results from abnormal signaling or processing in the nervous system. Pharmacopuncture with Entrapment Neuropathy Unties (ENUs), a multi-herbal formulation, may offer a complementary therapeutic strategy. However, its efficacy has not been scientifically validated in vivo.
Methods: A mouse model of sciatic nerve ligation (SNL)-induced neuropathic pain was used. Behavioral assessments were performed using Von Frey filaments to measure mechanical allodynia. Immunofluorescence staining was conducted to detect C-FOS and GFAP expression in the spinal dorsal horn. Quantitative PCR (qPCR) was used to evaluate the expression of inflammatory markers, including Gfap, Iba1, Tnf-α, and Il-1β.
Results: Local administration of ENUs at the injury site significantly alleviated mechanical allodynia induced by SNL (*p < 0.05, **p < 0.01, ***p < 0.001). Treatment with ENUs also led to statistically significant reductions in the expression of C-FOS, GFAP, and pro-inflammatory cytokines (*p < 0.05, **p < 0.01, ***p < 0.001). Among the treatment groups, the ENU V2-middle and V2-high dose groups demonstrated the most pronounced therapeutic effects compared to the saline-treated control group.
Conclusion: This study provides the first in vivo evidence supporting the analgesic and anti-inflammatory effects of ENUs in neuropathic pain. ENUs may exert these effects by suppressing glial activation and neuronal sensitization. Further research is warranted to explore its clinical applications and underlying molecular mechanisms.
Keywords: pharmacopuncture, neuropathic pain, inflammation, sciatic nerve ligation, (SNL), Von Frey filaments
Introduction
Neuropathic pain arises from damage or dysfunction of the nervous system, leading to abnormal pain signaling and processing. It can be caused by various factors, including nerve injury, infections, chronic diseases such as diabetes or multiple sclerosis, and certain medications.1–3 This type of pain is often described as burning, shooting, or electric-like, and is frequently accompanied by numbness, tingling, or loss of sensation in the affected areas. Sciatic nerve ligation (SNL) is a widely used experimental model for inducing neuropathic pain in animals. The procedure involves ligating the sciatic nerve, resulting in nerve fiber damage, inflammation, and altered neural activity.4 SNL-induced neuropathy typically presents with allodynia (pain from non-painful stimuli) and hyperalgesia (increased pain from painful stimuli), closely mimicking human neuropathic conditions.5,6 This model is extensively employed to investigate the underlying mechanisms of neuropathic pain and to evaluate the efficacy of novel therapeutic interventions.7
Recent studies highlight the critical role of glial cells—non-neuronal cells within the nervous system—in the development and persistence of neuropathic pain.8 Among these, microglia and astrocytes in the spinal cord are particularly important. After nerve injury, these glial cells become activated and release pro-inflammatory cytokines and chemokines, contributing to neuroinflammation and further neuronal damage.9 Moreover, glial activation is associated with central sensitization—a state where spinal neurons become abnormally sensitive to pain signals.10 Consequently, therapies that target glial cell activation have emerged as promising strategies for the treatment of neuropathic pain.11
Central sensitization is characterized by persistent pain due to altered nociceptive processing in the spinal cord. The immediate early gene C-FOS is widely recognized as a marker of neuronal activity in this process.12 Pain-related stimuli increase the expression of C-FOS in the spinal dorsal horn, enhancing the responsiveness of central neurons.13,14 This sensitization can lead to exaggerated pain perception, even in response to normally non-painful stimuli, and may contribute to the development of chronic pain “memory.” Accordingly, therapeutic strategies that suppress C-FOS expression may help reduce central sensitization and neuropathic pain.
Acupuncture has been explored as an alternative treatment for neuropathic pain. Preclinical studies have shown that acupuncture can attenuate inflammatory responses induced by SNL and reduce pain behaviors in animal models.15,16 These effects may involve modulation of neuroinflammation and gene expression, including microRNAs, involved in pain pathways.17 Building on these concepts, pharmacopuncture—which involves the injection of herbal extracts into acupuncture points or lesion sites—has gained attention as a complementary approach for pain relief.
Entrapment Neuropathy Unties (ENUs) is a pharmacopuncture formulation composed of traditional herbal ingredients, each known for their anti-inflammatory and analgesic properties: Erycibae Caulis has been reported to inhibit COX-2 and inflammatory cytokines, demonstrating potential in reducing neuroinflammation and joint-related pain.18,19 Geranii Herba contains geraniin and other tannins with antioxidant and antinociceptive effects, potentially modulating glial cell activation.20 Fermented Glycine max enhances isoflavone bioavailability, which in turn suppresses pro-inflammatory pathways such as NF-κB and MAPK signaling.21 Polygonati Rhizoma has demonstrated neuroprotective effects via antioxidant mechanisms and has been studied in the context of ischemic brain injury.22 Imperatae Rhizoma is traditionally used for its antipyretic and hemostatic effects and also exhibits anti-inflammatory activity through modulation of oxidative stress.23
Despite their long history of traditional use, the efficacy of this combined herbal formulation has not been scientifically validated in a neuropathic pain model. To our knowledge, this study is the first to evaluate the in vivo therapeutic effects of ENUs in an SNL-induced neuropathic pain model. Moreover, we investigated its mechanisms of action with a focus on glial modulation and suppression of C-FOS expression, which are closely associated with the maintenance of neuropathic pain. Therefore, this study aimed to investigate the therapeutic potential of ENUs in a mouse model of SNL-induced neuropathic pain. We conducted behavioral, histological, and molecular analyses to evaluate its analgesic and anti-inflammatory effects, focusing on glial cell activation and C-FOS expression. This is the first study to demonstrate the in vivo efficacy of ENUs in modulating neuropathic pain. Our findings provide a scientific foundation for the potential use of ENUs as a complementary or alternative treatment strategy for neuropathic pain.
Materials and Methods
Animals
In this study, 6 weeks old male ICR mice (20–25g) were obtained from DBL Co., Ltd. (Eumseong-gun, Chungcheongbuk-do, South Korea). The mice were acclimatized for one week before being used for the animal model creation. During the experimental period, food and water were made available ad libitum. The light/dark cycle was set to 12 h (daylight from 08:00 to 20:00), and the temperature was maintained at 23±2 °C with a humidity of 50±10% to ensure consistent housing conditions until the end of the experiment.
A total of 30 mice were divided into the following groups: sham group (Nor, n=6), control group receiving saline injection after SNL modeling (Control, n=6), treatment group receiving ENU V2-low concentration injection after SNL modeling (ENU V2-low, n=6), treatment group receiving ENU V2-middle concentration injection after SNL modeling (ENU V2-middle, n=6) and treatment group receiving ENU V2-high concentration injection after SNL modeling (ENU V2-high, n=6).
Preparation of ENUs Pharmacopuncture
ENUs pharmacopuncture solution was prepared by NamYi Pharmaceutical Co., Ltd. (South Korea) using standardized protocols. The formulation contained equal parts (100 g each) of Erycibae Caulis cortex/barks, Geranii Herba, fermented Glycine max Merr., Polygonati Rhizoma, and Imperatae Rhizoma. Herbal components were washed, and a 1:10 (w/v) mixture of herbs and purified water underwent low-temperature extraction (65–80°C, 48–60 h), followed by thermal processing (105–115°C, 2 h) to remove volatile compounds. The extract was distilled (105–120°C), filtered sequentially through 0.45 μm and 0.1 μm membranes, and adjusted to physiological salt concentration (0.75–0.98%) and pH (7.25–7.85). After sterilization (125–130°C, 25–30 min), the solution was aseptically dispensed into vials.
Administration Protocol
Mice received 100 μL ENUs solution via perineural injection (31G insulin syringe) around the ligated sciatic nerve every other day for 14 days (total 7 administrations). Sham and control groups received equivalent saline injections.
SNL Model
The procedure of SNL model was conducted according to the method described by Honda K. and Takano Y. with some modifications.24 The mice were anesthetized by using an intraperitoneal injection of avertin (50 mg/kg), after which one-third to one-half of the left sciatic nerve near the upper thigh was ligated 3 times with a No. 7.0 black silk suture. For the mice that underwent sham surgery, the sciatic nerve was exposed without ligation. All animal procedures were approved by the Institutional Animal Care and Use Committee (IACUC) of Stand Up Therapeutics (22-IACUC-0001) and conducted in accordance with the “IACUC Standard Operating Guidelines for Animal Experimentation” (Animal and Plant Quarantine Agency and Ministry of Food and Drug Safety of Korea, 2020). After SNL or sham surgery, animals were monitored at least twice daily for signs of pain, distress, wound healing, and general health. Analgesics were administered if animals exhibited signs of severe pain or distress, as per IACUC recommendations.
Behavioral Test
Mechanical sensitivity was performed according to the method described by previous study.25 Briefly, it was determined by examining the response rates of paw withdrawals from 10 repeated stimuli applied to the paw using a von Frey filament delivering 0.4 g (Stoelting, Wood Dale, IL). All experiments were conducted by an experimenter blinded to the treatment groups. Mice were placed in a plastic box on a metal grid floor and acclimated for 15 to 20 min prior to testing. The von Frey filament was applied perpendicularly to the skin for 2 to 3 s on the left hindpaw with enough force to bend it slightly. An abrupt withdrawal of the foot during or immediately after stimulation was regarded as a positive response. Response rates were calculated as a percentage of the number of positive responses per 10 stimuli.
Single-Dose Administration Test
On the 14th day after inducing neuropathic pain, mechanical allodynia was assessed at 1, 3, 7, 12, 24, 32, and 48 h following a single administration of acupuncture in order to evaluate the acute analgesic efficacy and determine the duration for repeated administration.
Repeated Administration Test
Based on the effects observed after the single-dose administration, mechanical allodynia was assessed every other day for a total of 7 times, starting from the 14th day after inducing neuropathic pain and continuing until the 28th day.
Immunofluorescence
After completing the behavioral test, the mice were anesthetized with avertin (50 mg/kg, i.p). The mice were perfused with 50 mL of phosphate-buffered saline (PBS) followed by 80 mL of 4% formalin solution prepared in phosphate buffer. The fixed mice were dissected to extract the spinal cord, which were then fixed in the same fixative solution for 24 h. After fixation, the tissues were placed in PBS containing 30% sucrose and stored at 4 °C for one day. The next day, the tissues were rapidly frozen and cut into sections of 8–10 μm thickness. The sectioned tissues were washed three times with Tris-buffered saline with 0.3% Triton-X100 (TBST) for 10 min each, followed by incubation in a solution of 5% bovine serum albumin (BSA) diluted in TBST for 1 hour. Primary antibodies, C-FOS (1:2000, rabbit monoclonal, ImmunSmol) and GFAP (1:1000, Guinea pig monoclonal, Abcam), were prepared by diluting them in TBST containing 5% BSA. The tissues were incubated with the primary antibodies at 4 °C for 24 h. Subsequently, the spinal cord tissues were washed with TBST and incubated with secondary antibodies for 2 h at room temperature. The secondary antibodies used were Alexa anti-rabbit 488 (1:2000, Invitrogen), Alexa anti-mouse 594 (1:2000, Invitrogen), or Alexa anti-guinea pig 594 (1:2000, Invitrogen), diluted in TBST containing 5% BSA. After three washes with TBST, the sections were mounted with mounting medium. The expression levels in different regions of the tissue were observed and captured using a confocal microscope (Olympus Corporation, Tokyo, Japan).
Quantitative Real–Time PCR (qPCR)
Total RNA was extracted from the ipsilateral dorsal horn tissue of the spinal cord using TRIzol reagent (Invitrogen, Carlsbad, CA, USA) according to the manufacturer’s instructions. Quantitative real-time PCR (qRT-PCR) was performed to analyze gene expression levels using the One Step SYBR® Prime Script™ RT-PCR Kit II (Takara, Otsu, Shiga, Japan) in a 20 µL total reaction volume under standard PCR conditions. The ABI Prism 7500 Real-Time PCR System (Life Technologies Corporation, CA, USA) was used for amplification and data collection. Ct values were normalized to the housekeeping gene Gapdh, and relative expression levels were calculated using the 2^−ΔΔCt method, with the control group as the reference. The primer sequences used in this study are shown in Table 1.
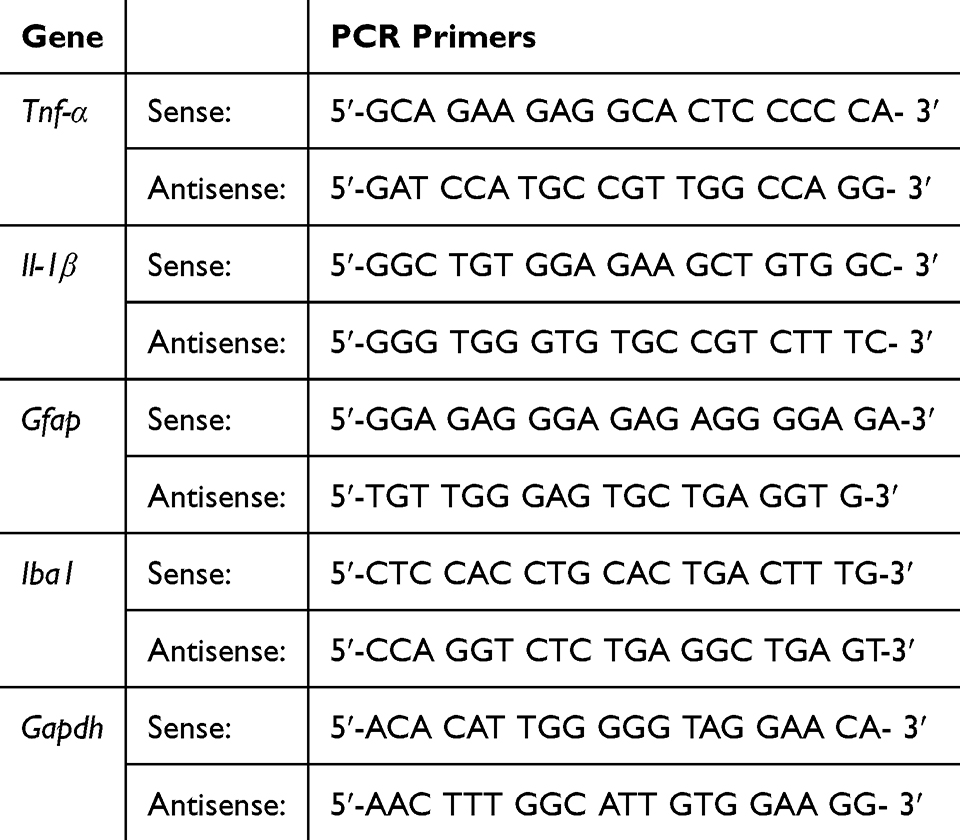 |
Table 1 Primers for RT-PCR Analysis |
Statistics
Data are represented as the mean ± standard error of the mean (SEM). For single comparisons, significance was assessed using Student’s t-test. For multiple comparisons, data were analyzed by one-way ANOVA followed by Bonferroni post-hoc correction to control the family-wise error rate (GraphPad Prism 8, USA).
Results
Experimental Design and Baseline Evaluation of Mechanical Allodynia
To establish a neuropathic pain model, we performed unilateral ligation of the left sciatic nerve in mice. Mechanical allodynia was assessed using the Von Frey filament test on postoperative days 1, 3, 7, 10, and 14 to monitor pain hypersensitivity over time. As shown in Figure 1A and B, mechanical allodynia significantly increased in all surgery groups compared to the sham group from day 3 onward (p < 0.001), indicating successful induction of neuropathic pain.
 |
Figure 1 Experimental scheme and baseline evaluation of mechanical allodynia in the SNL mouse model. Notes: (A) Schematic timeline of the experimental procedures, including neuropathic pain induction by sciatic nerve ligation (SNL), behavioral assessment, and ENU treatment. (B) Mechanical allodynia was assessed using the Von Frey test on days 1, 3, 7, 10, and 14 post-SNL to confirm the development of neuropathic pain. Data are expressed as mean ± SE (n = 6 per group). Statistical analysis was performed using one-way ANOVA followed by Bonferroni post hoc correction. ####p < 0.001 vs control group. |
On day 14, mice exhibiting sustained mechanical hypersensitivity were randomly assigned to treatment groups for further pharmacological testing. To evaluate the acute analgesic effects of ENU compounds, a single dose was administered, and mechanical thresholds were measured at 1, 3, 7, 12, 24, 32, and 48 hours post-treatment. Based on the observed time course of analgesia, repeated dosing was subsequently performed every 2 days between days 14 and 28. Mechanical allodynia was measured prior to each administration to evaluate the cumulative or sustained effects of repeated ENU treatment. These results confirm the successful induction of neuropathic pain and provide a defined baseline for assessing the therapeutic potential of ENUs in subsequent experiments.
Acute and Repeated Analgesic Effects of ENUs Administration in Neuropathic Pain-Induced Mice
To assess the acute analgesic effect of ENUs, the reduction in mechanical allodynia was measured using Von Frey filaments. Administration of ENU V1 in neuropathic pain-induced mice showed a statistically significant reduction in pain compared to the control group at 7, 12, and 24 h (p<0.05, p<0.01). Administration of ENU V2-low exhibited a trend of pain reduction, while ENU V2-middle and ENU V2-high showed statistically significant analgesic effects compared to the control group (p<0.05, p<0.01, p<0.001) at 7, 12, and 24 h. The acute pain reduction effects of ENUs administration are shown in Figure 2A.
 |
Figure 2 Analgesic effects of acute and repeated administration of ENUs in mice with SNL-induced neuropathic pain. Notes: (A) To assess the acute effects of ENUs, mechanical allodynia was measured at 1, 3, 7, 12, 24, 32, and 48 hours after a single administration. (B) Repeated ENU treatment was conducted once every two days from day 14 to day 28 post-SNL, and mechanical allodynia was measured prior to each treatment. Data are expressed as mean ± SE (n = 6 per group). Statistical analysis was performed using one-way ANOVA followed by Bonferroni post hoc correction. *p < 0.05, **p < 0.01, ***p < 0.001 vs control group. |
To evaluate the analgesic effects of repeated ENUs administration, mechanical allodynia was measured using Von Frey filaments once every 2 days for a duration of 14 days. Administration of ENU V2-low showed a trend of pain reduction compared to the control group, but there was no statistically significant difference. The ENU V2-middle group exhibited statistically significant reduction in pain compared to the control group at 6, 8, 10, and 14 days, showing the best effect on the last day (p<0.05, p<0.01, p<0.001). Administration of ENU V2-high demonstrated analgesic effects starting from day 4 (p<0.05) and showed statistically significant analgesic effects compared to the control group treated with saline at 6, 8, 10, and 14 days (p<0.05, p<0.01, p<0.001). The analgesic effects of repeated ENUs administration on mechanical allodynia are shown in Figure 2B.
Effects of ENUs on Spinal Dorsal Horn Neuronal Activation in Neuropathic Pain-Induced Mice
To examine the abnormal activation of neurons in the spinal cord induced by SNL, the expression of C-FOS was assessed through immunohistochemical staining in the ipsilateral spinal dorsal horn of the damaged sciatic nerve. SNL significantly increased the expression of C-FOS in the spinal dorsal horn compared to the control group (p<0.01). Administration of ENU V2-low, V2-middle, and ENU V2-high in neuropathic pain-induced mice resulted in a statistically significant decrease in the expression of C-FOS compared to the control group (p<0.01). The administration of ENU V2 at different concentrations showed a dose-dependent trend in reducing the expression of C-FOS (Figure 3).
 |
Figure 3 Effects of ENUs on C-FOS expression in the spinal dorsal horn of SNL mice. Notes: (A) Representative immunofluorescence images showing C-FOS expression (green) in the spinal dorsal horn of SNL-induced neuropathic pain mice across treatment groups. (B) Quantification of C-FOS-positive cells per section. Fluorescence-positive cells were counted from three randomly selected regions of interest (ROIs) in the ipsilateral dorsal horn of each animal. Data are expressed as mean ± SEM (n = 6 per group). Statistical analysis was conducted using one-way ANOVA followed by Bonferroni post hoc correction. **p < 0.01 vs control group. |
Effects of ENUs on Spinal Dorsal Horn Astrocytes Activation in Neuropathic Pain-Induced Mice
To examine the activation of inflammatory cells in the spinal cord induced by SNL, the expression of the astrocyte marker GFAP was assessed through immunofluorescent staining in the ipsilateral spinal dorsal horn tissue section of the damaged sciatic nerve. In the results shown in Figure 4B, SNL significantly increased the expression intensity and area of GFAP protein in the spinal dorsal horn compared to the control group (p<0.001). Administration of ENU V2-low, V2-middle, and ENU V2-high in neuropathic pain-induced mice resulted in a statistically significant suppression of GFAP expression compared to the control group (p<0.001). The administration of ENU V2 at different concentrations showed a dose-dependent trend in reducing the expression of GFAP. The size of glial cells was also increased due to SNL, but it was statistically significantly decreased by the administration of ENU V2-low, V2-middle, and ENU V2-high (p<0.05, p<0.001) (Figure 4).
 |
Figure 4 Effects of ENUs on GFAP expression in the spinal dorsal horn of SNL mice. Notes: (A) Representative immunofluorescence images showing GFAP expression (red) in the spinal dorsal horn of SNL mice across treatment groups. (B) Quantitative analysis of GFAP-positive area and fluorescence intensity. Three ROIs were analyzed per section from the ipsilateral side of the spinal cord. Data are expressed as mean ± SEM (n = 6 per group). Statistical analysis was conducted using one-way ANOVA followed by Bonferroni post hoc correction. *p < 0.05, **p < 0.01 vs control group. |
Effects of ENUs on the Expression of Inflammatory Markers in Neuropathic Pain-Induced Mice
To examine the activation of inflammatory cells and the expression of inflammatory cytokines in the spinal cord induced by SNL, the expression of the astrocyte marker Gfap and the microglia marker Iba1 were assessed through qPCR in the ipsilateral spinal dorsal horn tissue of the damaged sciatic nerve. The expression of the inflammatory markers Gfap and Iba1 was significantly increased in the control group compared to the normal group (p<0.001). The expression of Gfap showed a significant decrease in all groups administered with ENUs (Figure 5A left, p<0.01, p<0.001). On the other hand, the expression of Iba1 showed a significant decrease only in the groups administered with V2-middle and ENU V2-high (Figure 5A right, p<0.05).
 |
Figure 5 Effects of ENUs on mRNA expression of inflammatory markers in the spinal dorsal horn and peripheral tissue. Notes: (A) ENUs significantly reduced the mRNA expression of Gfap and Iba1 in the spinal dorsal horn of SNL mice. (B) ENUs also decreased the mRNA levels of Tnf-α and Il-1β in the edema-induced paw tissue. Quantification was performed by qPCR, normalized to housekeeping genes. Data are expressed as mean ± SE (n = 6 per group). Statistical analysis was conducted using one-way ANOVA followed by Bonferroni post hoc correction. *p < 0.05, **p < 0.01 vs control group. |
To examine the expression of inflammatory cytokines in the spinal cord induced by SNL, the expression of the inflammatory cytokines Tnf-α and Il-1β were assessed through qPCR in the ipsilateral spinal dorsal horn tissue of the damaged sciatic nerve. The expression of the inflammatory cytokine marker Il-1β was significantly increased in the control group compared to the normal group (p<0.001). However, this increase showed a significant decrease in all groups administered with ENUs (Figure 5B left, p<0.01, p<0.001). On the other hand, the expression of Tnf-α showed a significant decrease in the groups administered with V2-middle and ENU V2-high (Figure 5B right, p<0.001).
Discussion
This study investigated the therapeutic effects of ENUs on neuropathic pain using behavioral, histological, and molecular analyses to examine whether ENUs promote functional recovery through the inhibition of C-FOS expression and anti-inflammatory effects. Local administration of ENUs near the affected area significantly reduced mechanical allodynia induced by SNL. Additionally, ENUs regulated the expression of the neuronal activation marker C-FOS in the ipsilateral spinal dorsal horn. The increased presence of inflammatory cells and pro-inflammatory cytokines in the spinal dorsal horn due to SNL was reduced by local administration of ENUs.
The pharmacological activity of ENUs appears to be attributed to the synergistic effects of its herbal components. Erycibae Caulis contains compounds that exhibit anti-inflammatory effects, which may mitigate inflammation-driven pain responses.19 Similarly, Geranii Herba is rich in geraniin, an antioxidant with significant anti-inflammatory activity, potentially benefiting neuropathic pain management.26 Fermented Glycine max has enhanced bioavailability of isoflavones through fermentation, which increases its antioxidant and anti-inflammatory effects, thus aiding in the reduction of oxidative stress associated with nerve pain.27 Polygonati Rhizoma has been recognized for its neuroprotective properties, promoting nerve regeneration and reducing inflammation in neuropathic conditions.28 Finally, Imperatae Rhizoma extract is known for its ability to reduce inflammation and oxidative stress, which are key factors in neuropathic pain pathways.29 Collectively, these herbal extracts offer a promising complementary approach to managing neuropathic pain through their multifaceted bioactive mechanisms. Compared to conventional treatments such as gabapentin, NSAIDs, and opioids, which primarily target neuronal excitability or pain perception, ENUs demonstrate multi-target efficacy involving glial modulation, neuronal sensitization, and peripheral immune regulation. These properties suggest that ENUs may serve as a promising complementary therapy, especially for patients who are unresponsive to conventional drugs or experience side effects.
The SNL model, a well-established preclinical tool, was used to induce neuropathic pain characterized by allodynia and hyperalgesia.30 Consistent with previous studies,6,7 mechanical allodynia developed rapidly after surgery and persisted throughout the experimental period. (Figure 1). Subsequently, neuropathic pain persisted until day 28, as seen in the control group results shown in Figure 2B. The acute analgesic effect of ENUs became statistically significant from 7 hours post-administration and lasted up to 48 hours (Figure 2A). Repeated administration of ENUs significantly reduced pain from day 4 (Figure 2B). Notably, ENU V2-middle and ENU V2-high groups showed statistically significant pain reduction starting from 7 hours post-injection, and more than 50% reduction was sustained compared to pre-administration levels.
C-FOS is an immediate early gene that reflects neuronal activity. In the context of pain, the expression of C-FOS signifies abnormal neuronal activation due to tactile stimuli.31 Pain stimuli originating from the central nervous system increase the expression of C-FOS in the spinal dorsal horn.13 Therefore, the C-FOS protein plays a crucial role in understanding and treating pain originating from the central nervous system.14,32 SNL significantly increased the expression of C-FOS in the spinal dorsal horn compared to the control group (Figure 4). Administration of ENU V2-low, V2-middle, and ENU V2-high significantly reduced the expression of C-FOS compared to the control. The administration of ENU V2 in different concentrations appeared to dose-dependently decrease the expression of C-FOS.
Activated microglia and astrocytes contribute to the pathogenesis of chronic neuropathic pain by releasing pro-inflammatory cytokines such as TNF-α and IL-1β.8,9,11 In this study, SNL-induced neuropathic pain resulted in a marked increase in GFAP expression and astrocyte hypertrophy in the spinal dorsal horn, indicating glial activation (Figure 4). Local administration of ENU V2 at all tested concentrations (low, middle, and high) significantly reduced GFAP expression compared to the control group, with a clear dose-dependent effect observed. In addition, ENUs suppressed the mRNA expression of inflammatory markers in the spinal cord (Figure 5). Specifically, Gfap expression was significantly decreased in all ENU-treated groups, while Iba1 expression was significantly reduced only in the ENU V2-middle and ENU V2-high groups, suggesting a concentration-dependent inhibition of microglial activation. Furthermore, the mRNA levels of the pro-inflammatory cytokines Tnf-α and Il-1β were significantly elevated in the SNL control group compared to the sham group (Figure 5B). Treatment with ENUs significantly attenuated these elevations across all treatment groups. Notably, Tnf-α expression showed a significant reduction only in the ENU V2-middle and ENU V2-high groups (p < 0.001), indicating a partial dose-dependent effect. These findings collectively suggest that ENUs modulate neuroinflammation by suppressing both glial activation and cytokine expression, contributing to their overall analgesic effect in neuropathic pain.
Despite the promising results, this study has several limitations. First, we did not perform protein-level validation such as Western blotting or histological staining beyond immunofluorescence. Second, the study was limited to short-term outcomes, and the long-term safety and efficacy of ENUs remain unknown. Third, variability in herbal composition and lack of clinical pharmacokinetic data may affect reproducibility and translational application. Lastly, interspecies differences between mice and humans limit direct extrapolation to clinical settings. Future studies should address these limitations by evaluating the long-term efficacy and safety of ENUs, standardizing herbal extract quality, and investigating its mechanisms using protein-level validation techniques. In addition, comparative trials with standard drugs such as gabapentin or duloxetine will be needed to determine the clinical relevance of ENUs.
These findings are supported by recent studies reporting the effects of herbal agents and acupuncture-like therapies on neuroinflammation and glial cell modulation in neuropathic models.33–35 Together, this work provides the first in vivo evidence of the therapeutic potential of ENUs for neuropathic pain, laying the foundation for future translational research.
Conclusion
This study investigated the therapeutic effects of ENUs on neuropathic pain through behavioral, histological, and molecular biology analyses. ENUs demonstrated pain-relieving effects by alleviating mechanical allodynia, suppressing C-FOS expression, and exerting anti-inflammatory effects to restore spinal cord function. Based on the overall research results, ENU V2-middle and ENU V2-high exhibited the highest analgesic effects. This study provides the first evidence of the in vivo effects of ENUs in a neuropathic mouse model, scientifically validating the therapeutic effects of ENUs. ENUs can be beneficial for the treatment of neuropathic pain and may be utilized in the development of new pain therapies.
Data Sharing Statement
All the datasets in manuscript or additional file are available from the corresponding authors (Gi-Sang Bae and Youngjin Choi) on reasonable request.
Ethics Approval and Consent to Participate
All experiments were approved by the Institutional Animal Care and Use Committee of Stand Up Therapeutics for ethical considerations.
Funding
This paper was sponsored by Soongsan Fellowship in Wonkwang University in 2023.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Baron R, Maier C, Attal N, et al. Peripheral neuropathic pain: a mechanism-related organizing principle based on sensory profiles. Pain. 2017;158(2):261–272. doi:10.1097/j.pain.0000000000000753
2. Finnerup NB, Attal N, Haroutounian S, et al. Pharmacotherapy for neuropathic pain in adults: a systematic review and meta-analysis. Lancet Neurol. 2015;14(2):162–173. doi:10.1016/S1474-4422(14)70251-0
3. Gilron I, Baron R, Jensen T. Neuropathic pain: principles of diagnosis and treatment. Mayo Clin Proc. 2015;90(4):532–545. doi:10.1016/j.mayocp.2015.01.018
4. Jaggi AS, Jain V, Singh N. Animal models of neuropathic pain. Fundament Clin Pharmacol. 2011;25(1):1–28. doi:10.1111/j.1472-8206.2009.00801.x
5. Decosterd I, Woolf CJ. Spared nerve injury: an animal model of persistent peripheral neuropathic pain. Pain. 2000;87(2):149–158. doi:10.1016/S0304-3959(00)00276-1
6. Kim SH, Chung JM. An experimental model for peripheral neuropathy produced by segmental spinal nerve ligation in the rat. Pain. 1992;50(3):355–363. doi:10.1016/0304-3959(92)90041-9
7. Devor M. Pathophysiology of damaged nerves in relation to chronic pain. 1999.
8. Ikeda H, Tsuda M, Inoue K, Murase K. Long‐term potentiation of neuronal excitation by neuron–glia interactions in the rat spinal dorsal horn. Eur J Neurosci. 2007;25(5):1297–1306. doi:10.1111/j.1460-9568.2007.05386.x
9. Milligan ED, Watkins LR. Pathological and protective roles of glia in chronic pain. Nat Rev Neurosci. 2009;10(1):23–36. doi:10.1038/nrn2533
10. Ren K, Dubner R. Interactions between the immune and nervous systems in pain. Nat Med. 2010;16(11):1267–1276. doi:10.1038/nm.2234
11. Raghavendra V, Tanga F, DeLeo JA. Inhibition of microglial activation attenuates the development but not existing hypersensitivity in a rat model of neuropathy. J Pharmacol Exp Ther. 2003;306(2):624–630. doi:10.1124/jpet.103.052407
12. Ji -R-R, Nackley A, Huh Y, Terrando N, Maixner W. Neuroinflammation and central sensitization in chronic and widespread pain. Anesthesiology. 2018;129(2):343–366. doi:10.1097/ALN.0000000000002130
13. Gao Y-J, Ji -R-R. c-Fos and pERK, which is a better marker for neuronal activation and central sensitization after noxious stimulation and tissue injury? Open Pain J. 2009;2:11. doi:10.2174/1876386300902010011
14. Peirs C, Williams S-PG, Zhao X, et al. Mechanical allodynia circuitry in the dorsal horn is defined by the nature of the injury. Neuron. 2021;109(1):73–90.e77. doi:10.1016/j.neuron.2020.10.027
15. Baek YH, Huh JE, Lee JD, Park DS. Antinociceptive effect and the mechanism of bee venom acupuncture (Apipuncture) on inflammatory pain in the rat model of collagen-induced arthritis: mediation by α2-adrenoceptors. Brain Res. 2006;1073:305–310. doi:10.1016/j.brainres.2005.12.086
16. Yoon Y-J, Kim S, Cho J-H, Kim K-W, Song M-Y. Effect of acupuncture on sciatica in rat models: systematic review and meta-analysis. J Korean Med Rehabil. 2020;30(1):79–93. doi:10.18325/jkmr.2020.30.1.79
17. Ou S, Zhao Y-D, Xiao Z, Wen H-Z, Cui J, Ruan H-Z. Effect of lappaconitine on neuropathic pain mediated by P2X3 receptor in rat dorsal root ganglion. Neurochem Int. 2011;58(5):564–573. doi:10.1016/j.neuint.2011.01.016
18. Pan R, Gao XH, Li Y, Xia YF, Dai Y. Anti‐arthritic effect of scopoletin, a coumarin compound occurring in Erycibe obtusifolia Benth stems, is associated with decreased angiogenesis in synovium. Fundament Clin Pharmacol. 2010;24(4):477–490. doi:10.1111/j.1472-8206.2009.00784.x
19. Chen Z, Liao L, Zhang Z, Wu L, Wang Z. Comparison of active constituents, acute toxicity, anti-nociceptive and anti-inflammatory activities of Porana sinensis Hemsl. Erycibe obtusifolia Benth. and Erycibe schmidtii Craib. J Ethnopharmacol. 2013;150(2):501–506. doi:10.1016/j.jep.2013.08.059
20. Kim B-M, Kim E-S, Lee Y-H, Yu B-S, Chai K-Y. Antiinflammatory effect of bioactive compounds from geranium nepalense. Herbal Formula Sci. 2010;18(2):241–249.
21. Kim HY, Cha HY, Ha K, Choi JY, Chen JH, Kim K. Pilot study on anti-wrinkle effect of herbal cosmetic containing the extracts of Douchi (fermented Glycine max Merr.). J Physiol Pathol Korean Med. 2017;31(6):380–384. doi:10.15188/kjopp.2017.12.31.6.380
22. Kang M-W, Chang J-P, Doh E-S, Kil K-J, Yoo J-H. Antioxidant activities of water extracts from steamed Polygonati rhizoma. Korea J Herbol. 2017;32(1):33–40. doi:10.6116/kjh.2017.32.1.33
23. Kim B-K, Lim J-S, Kil K-J. Effects of imperatae Rhizoma extract on T helper 2 cell differentiation. Korea J Herbol. 2014;29(6):27–33. doi:10.6116/kjh.2014.29.6.27
24. Honda K, Takano Y. Experimental methods in pain using neuropathic and inflammatory animal models. Nihon yakurigaku zasshi Folia pharmacologica Japonica. 2007;130(1):39–44. doi:10.1254/fpj.130.39
25. Shim HS, Bae C, Wang J, et al. Peripheral and central oxidative stress in chemotherapy-induced neuropathic pain. Molecular Pain. 2019;15:1744806919840098. doi:10.1177/1744806919840098
26. Greenway FL, Frome BM, Engels TM, McLellan A. Temporary relief of postherpetic neuralgia pain with topical geranium oil. Am J Med. 2003;115(7):586–587. doi:10.1016/S0002-9343(03)00434-0
27. Moussa L, Bezirard V, Salvador-Cartier C, et al. A low dose of fermented soy germ alleviates gut barrier injury, hyperalgesia and faecal protease activity in a rat model of inflammatory bowel disease. PLoS One. 2012;7(11):e49547. doi:10.1371/journal.pone.0049547
28. Wang S, He F, Wu H, et al. Health-promoting activities and associated mechanisms of Polygonati rhizoma polysaccharides. Molecules. 2023;28(3):1350. doi:10.3390/molecules28031350
29. Zou W, Dong Y, Yang S, et al. Imperatae rhizoma-Hedyotis diffusa Willd. herbal pair alleviates nephrotic syndrome by integrating anti-inflammatory and hypolipidaemic effects. Phytomedicine. 2021;90:153644. doi:10.1016/j.phymed.2021.153644
30. Malmberg AB, Basbaum AIJP. Partial sciatic nerve injury in the mouse as a model of neuropathic pain: behavioral and neuroanatomical correlates. Pain. 1998;76(1–2):215–222. doi:10.1016/S0304-3959(98)00045-1
31. Latremoliere A, Woolf C. Central sensitization: a generator of pain hypersensitivity by central neural plasticity. J Pain. 2009;10(9):895–926. doi:10.1016/j.jpain.2009.06.012
32. Neumann S, Braz JM, Skinner K, Llewellyn-Smith IJ, Basbaum AI. Innocuous, not noxious, input activates PKCγ interneurons of the spinal dorsal horn via myelinated afferent fibers. J Neurosci. 2008;28(32):7936–7944. doi:10.1523/JNEUROSCI.1259-08.2008
33. Zhang D, Jing B, Chen ZN, et al. Ferulic acid alleviates sciatica by inhibiting neuroinflammation and promoting nerve repair via the TLR4/NF-κB pathway. CNS Neurosci Ther. 2023;29(4):1000–1011. doi:10.1111/cns.14060
34. Di Z, Zhenni C, Zifeng Z, et al. Danggui Sini Decoction normalizes the intestinal microbiota and serum metabolite levels to treat sciatica. Phytomedicine. 2024;132:155740. doi:10.1016/j.phymed.2024.155740
35. Chang S, Li X, Zheng Y, et al. Kaempferol exerts a neuroprotective effect to reduce neuropathic pain through TLR4/NF-ĸB signaling pathway. Phytother Res. 2022;36(4):1678–1691. doi:10.1002/ptr.7396
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms.php
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms.php
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Ligusticum chuanxiong Hort. Ameliorates Neuropathic Pain by Regulating Microglial M1 Polarization: A Study Based on Network Pharmacology
Cui S, Feng X, Xia Z
Journal of Pain Research 2024, 17:1881-1901
Published Date: 23 May 2024