")
Back to Journals » International Journal of Nanomedicine » Volume 20
Extracellular Vesicles: A Review of Their Therapeutic Potentials, Sources, Biodistribution, and Administration Routes
Authors Su X , Wang H , Li Q, Chen Z
Received 29 October 2024
Accepted for publication 18 February 2025
Published 13 March 2025 Volume 2025:20 Pages 3175—3199
DOI https://doi.org/10.2147/IJN.S502591
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 4
Editor who approved publication: Dr Kamakhya Misra
Xiaorong Su,1,* Hongxiang Wang,2,* Qiubai Li,3,4 Zhichao Chen1
1Department of Hematology, Union Hospital, Tongji Medical College, Huazhong University of Science and Technology, Wuhan, 430022, People’s Republic of China; 2Department of Hematology, Key Laboratory for Molecular Diagnosis of Hubei Province, the Central Hospital of Wuhan, Tongji Medical College, Huazhong University of Science and Technology, Wuhan, 430014, People’s Republic of China; 3Department of Rheumatology and Immunology, Union Hospital, Tongji Medical College, Huazhong University of Science and Technology, Wuhan, 430022, People’s Republic of China; 4Hubei Engineering Research Center for Application of Extracellular Vesicle, Hubei University of Science and Technology, Xianning, 437100, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Zhichao Chen, Department of Hematology, Union Hospital, Tongji Medical College, Huazhong University of Science and Technology, Wuhan, 430022, People’s Republic of China, Email [email protected] Qiubai Li, Department of Rheumatology and Immunology, Union Hospital, Tongji Medical College, Huazhong University of Science and Technology, Wuhan, 430022, People’s Republic of China, Email [email protected]
Abstract: Extracellular vesicles (EVs) participate in intercellular communication and play an essential role in physiological and pathological processes. In recent years, EVs have garnered significant attention as cell-free therapeutic alternatives, vectors for drug and gene delivery, biomarkers for disease diagnosis and prognosis, vaccine development, and nutraceuticals. The biodistribution of EVs critically influences their efficacy and toxicity. Therefore, this review aims to discuss the main factors influencing the biodistribution of unmodified EVs, highlighting their distribution patterns, advantages, limitations, and applications under different routes of administration. In addition, we provide a comprehensive discussion of the currently available sources of EVs and summarize the current status of the therapeutic potentials of EVs. By optimizing administration routes and selecting appropriate EV sources, we aim to offer valuable insights to enhance the delivery efficiency and therapeutic efficacy of EVs to target tissues.
Keywords: extracellular vesicles, administration routes, biodistribution, drug delivery
Introduction
Extracellular vesicles (EVs) are lipid bilayer vesicles secreted by seemingly all cells. Based on their biogenesis, EVs are primarily classified as exosomes and microvesicles. Moreover, specific cellular processes, such as cell migration and programmed cell death, generate migrasomes and apoptotic bodies, which are also classified as EVs.1,2 According to the MISEV 2023 guidelines, vesicles are recommended to be classified into extracellular vesicles, synthetic vesicles, and artificial cell-derived vesicles. The guidelines further advise against using nomenclature related to biogenesis, including terms like exosomes and microvesicles.2 Therefore, in this review, all types of vesicles will be collectively referred to as EVs.
EVs were first reported in 1946 when researchers observed coagulation-promoting particles derived from platelets in plasma.3 Initially, EVs were thought to be a way for cells to dispose of their “garbage”.4 With the ongoing advancement in research, EVs have been found to carry various proteins, nucleic acids, and lipids derived from their parental cells, playing a crucial role in intercellular communication. Presently, EVs are believed to interact with receptor cells through three mechanisms: direct receptor interaction to trigger downstream signaling cascades, plasma membrane fusion to release their contents into the cytoplasm, and internalization (Figure 1).5,6 Among these, EV internalization is the most extensively reported mechanism, occurring via multiple pathways including clathrin-mediated endocytosis, lipid raft-mediated endocytosis, caveolin-mediated endocytosis, phagocytosis, and micropinocytosis. However, there remains no consensus on the predominant mechanism governing EV uptake and content delivery. Furthermore, some researchers have suggested that the EV uptake pathway may depend more on the type of receptor cell than on the characteristics of the EVs themselves.7 Nonetheless, identifying the precise mechanism of EV-receptor cell interactions has profound implications for advancing therapeutic applications of EVs.
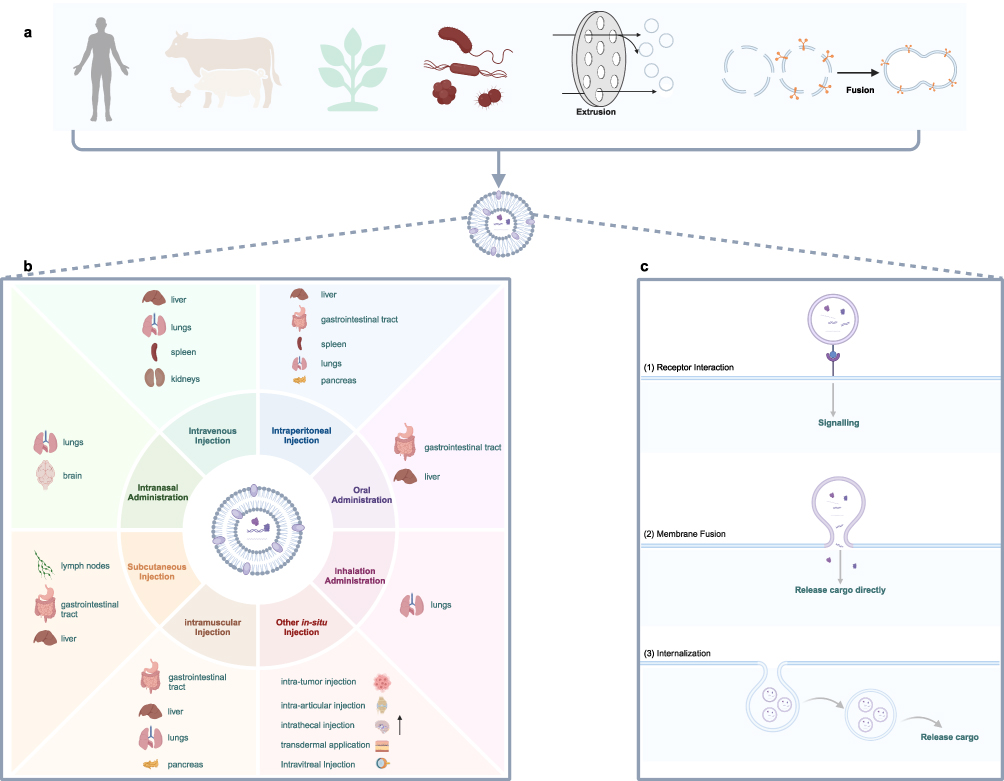 |
Figure 1 Biodistribution and cellular uptake of EVs in organisms (a) EVs originate from diverse sources, including animals, plants, microorganisms, artificial cell-derived vesicles, and synthetic vesicles. (b) The biodistribution profiles of EVs vary depending on the route of administration. (c) Receptor cells employ various mechanisms for the uptake of EVs. Created in BioRender. Su, X. (2025) https://BioRender.com/t38l645. |
Based on their biological activities, EVs play an important role in the physiological and pathological processes of organisms. Furthermore, EVs are distinguished by their intrinsic stability, low immunogenicity, biocompatibility, extended half-life, and superior ability to traverse biological barriers.8,9 Consequently, EVs have recently become a focal point of intensive research as cell-free therapeutic alternatives, drug and gene delivery vectors, biomarkers for disease diagnosis and prognosis, vaccine development platforms, and nutraceutical carriers. The efficacy and toxicity profiles of EVs, whether utilized as drug delivery systems or functional entities, are profoundly influenced by their biodistribution characteristics.10 A significant amount of research has focused on engineering modifications to enhance the circulation stability and tissue-targeting capabilities of EVs.11 However, limited attention has been devoted to investigating how various EV sources, delivery routes, and other conditions influence tissue accumulation and therapeutic efficacy. For instance, intravenously administered EVs derived from mesenchymal stem cells (MSCs) exhibited significant accumulation in the liver, making them a promising option for liver-targeted therapies.12 In contrast, intranasally delivered MSC-derived EVs predominantly accumulated in the lungs and showed greater distribution in the brain compared to intravenous administration.13 Intranasal delivery thus represents a viable route for enhancing the targeting of EVs to central nervous system disorders. This review discusses the main factors influencing the biodistribution of unmodified EVs, emphasizing their distribution and applications across various administration routes, including intravenous, intraperitoneal, oral, intranasal, transdermal, inhalation, intramuscular injection, and in situ administration (Figure 1). This information serves as a valuable reference for researchers selecting appropriate EV delivery routes based on their specific research goals. Furthermore, as the sources of EVs are becoming increasingly diversified, a comprehensive summary of the characteristics of EVs from various sources remains lacking. We provide a comprehensive discussion of current EV sources, highlighting the advantages and limitations of each, including underexplored EVs such as plant exosome-like nanovesicles (PELNVs), bacterial outer membrane vesicles (OMVs), and EV mimetics (EVMs). In addition to targeted modification, the selection of suitable EV sources and optimization of administration routes appear to help enhance the therapeutic potential of EVs.
Therapeutic Potentials of EVs
EVs as Cell-Free Therapeutic Alternatives
Stem cells, particularly mesenchymal stem cells (MSCs), have emerged as a cornerstone in the field of regenerative medicine. However, certain studies have indicated potential challenges with stem cell applications, including graft rejection after allogeneic transplantation14 and the risk of microvascular occlusion.15 EVs inherit numerous bioactive molecules from their parent cells, thereby exhibiting comparable therapeutic potential.16 For instance, both intra-articular injection of bone marrow-derived MSCs (BM-MSCs) and BM-MSC-derived EVs have been shown to mitigate cartilage destruction and subchondral bone remodeling in a rat model of osteoarthritis.17 Compared to their parent cells, EVs offer distinct advantages such as lower immunogenicity, non-tumorigenicity, and the ability to cross biological barriers.18,19 Additionally, research has demonstrated that the size characteristics of EVs enable secure systemic delivery via various routes without inducing embolism.19
To date, numerous clinical trials have validated the efficacy and safety of EVs. Small EVs derived from human placental MSCs have significantly reduced mortality rates and extended survival times in the intervention groups, suggesting a promising therapeutic strategy for severe COVID-19 patients.20 For chronic kidney disease patients, EVs from human umbilical cord MSCs have been shown to decrease plasma TNF-α levels while increasing TGF-β1 and IL-10 levels, thereby safely mitigating inflammatory immune responses and improving renal function.21 Platelet-derived EVs (pEVs) activate the ERK and Akt signaling pathways in recipient cells, promoting fibroblast proliferation and migration in vitro, as well as angiogenesis. A clinical trial has confirmed the safety of allogeneic pEVs in promoting wound healing in healthy individuals, indicating potential applications for patients with delayed or impaired wound healing.22 Moreover, research on the efficacy and safety of EVs in treating various systemic diseases, including graft-versus-host disease,23 chemotherapy-induced alopecia,24 and Alzheimer’s disease,25 has shown promising results in clinical trials. Despite being in its nascent stages, clinical research on EVs has revealed significant advantages and substantial potential for disease treatment and health promotion.
EVs as Important Carriers for Drug Delivery
Currently, clinically utilized therapeutic agents, including methotrexate, paclitaxel, doxorubicin, and curcumin, face problems such as inadequate water solubility, rapid clearance, and poor bioavailability.26,27 Moreover, non-specific uptake by healthy cells and tissues leads to widespread distribution of these cytotoxic agents throughout the body rather than targeted delivery to tumors.28 The issues of low targeting specificity and high toxicity associated with chemotherapeutic drugs are interrelated and challenging to address. MicroRNAs (miRNAs) play a crucial role in human growth and development, regulating various physiological and pathological processes. However, naked miRNAs and other gene therapy agents exhibit poor stability, short circulation half-life, and inadequate cellular uptake due to their susceptibility to degradation by nucleases in vivo. To overcome these limitations, nanoparticle delivery systems, including polymer nanoparticles, liposomes, and EVs, have been extensively developed to improve the pharmacokinetic and pharmacodynamic properties of therapeutic drugs, proteins, and nucleic acids. Since the FDA approved the first anticancer nano-drug, Doxil®, in 1995, liposomal products have undergone rapid development.29 However, several challenges have limited their clinical application, including poor long-term stability, high toxicity, drug leakage, and rapid clearance from circulation.30 For instance, liposomes can induce acute hypersensitivity reactions known as complement activation-related pseudoallergy (CARPA). Reportedly, 2–45% of patients experience infusion-related hypersensitivity reactions to liposomal treatments.31 Similarly, polymer nanoparticles also face challenges such as insufficient biocompatibility and toxicity.32 Utilizing natural carrier systems represents an effective strategy to address challenges in therapeutic drug delivery.
Compared with liposomes and synthetic polymer nanoparticles, EVs exhibit superior biocompatibility, stability, circulation half-life, and lower toxicity due to their endogenous nature.28,33 A study comparing liver injury indicators in mice pre-treated with grapefruit-derived nano vectors (GNVs) versus liposomes revealed significantly higher levels of alanine aminotransferase and aspartate aminotransferase in the liposome-treated group compared to those treated with GNVs.28 Furthermore, EVs demonstrate better inherent targeting capabilities compared to liposomes and polymer nanoparticles. Research has shown that when co-incubated with neurons, macrophage-derived EVs accumulated significantly more in neurons compared to polymeric nanoparticles and liposomes.34 Owing to the tight junctions between endothelial cells and the selective permeability of the blood-brain barrier (BBB), 100% of large molecule drugs and over 98% of small molecule drugs are unable to cross the BBB, thereby significantly limiting their therapeutic efficacy for brain diseases.35 Recent studies have demonstrated that EVs can traverse the BBB primarily through endocytosis,9 highlighting their considerable potential in the treatment of brain disorders. For instance, compared to free catalase (CAT), EVs loaded with CAT have been shown to effectively mitigate oxidative stress and enhance neuronal survival in a Parkinson’s disease mouse model.34 Furthermore, as natural nano-carriers, EVs carry bioactive molecules from their parent cells, which are advantageous for therapeutic applications. For instance, intestinal macrophages exhibited a preferential uptake of grapefruit-derived nanovesicles (GDN), leading to the upregulation of heme oxygenase-1 (HO-1) expression and the inhibition of TNF-α and IL-1β production. Compared with free methotrexate (MTX), MTX-loaded GDN significantly reduced MTX toxicity and improved DSS-induced colitis in mice.36
EVs as Biomarkers for Disease Diagnosis and Prognosis
Early diagnosis and prognosis monitoring of diseases are critical for reducing mortality and mitigating disease progression. EVs are present in various biological fluids, carrying bioactive molecules from parent cells and exhibiting distinct expression patterns during physiological and pathological processes. Furthermore, EVs have been identified in all bodily fluids, including blood, urine, saliva, and malignant ascites. They are considered promising biomarkers for disease diagnosis and prognostic monitoring in central nervous system disorders, oncology, cardiovascular diseases, and gastrointestinal conditions.37 Chatterjee et al demonstrated that plasma EVs contain measurable levels of TAR DNA-binding protein 43 (TDP-43) and full-length tau. The quantification and integration of plasma EV TDP-43 levels along with the ratio of 3R/4R tau isoforms could significantly enhance the molecular diagnosis of frontotemporal dementia (FTD) and amyotrophic lateral sclerosis (ALS). This represents a novel biomarker for identifying the molecular pathology associated with FTD and ALS.38 CD63, vimentin, and the epithelial cell adhesion molecule on plasma EVs from osteosarcoma patients can be utilized in a rapid diagnostic classification model for osteosarcoma. This model demonstrates a sensitivity of 100%, specificity of 90%, and overall accuracy of 95%.39 Jiang et al developed an effective urinary EVs-based prostate score (EPS) model for the Chinese population by utilizing urine-derived EVs and integrating the gene expressions of FOXA1, PCA3, and KLK3. This model demonstrates significantly higher diagnostic accuracy for clinically significant prostate cancer compared to prostate-specific antigen.40 The dual biomarker panel, comprising two small RNAs (tRNA-GlyGCC-5 and sRESE) derived from salivary EVs, effectively distinguishes patients with esophageal squamous cell carcinoma from healthy controls with a specificity of 90.5% and an accuracy of 94.2%. Additionally, this biomarker panel can predict overall survival and assess the potential benefit of adjuvant therapy for patients before surgery.41 Furthermore, EVs derived from gut microbiota have emerged as novel biomarkers for the diagnosis of gastrointestinal diseases.42
EVs for Tumor Immunotherapy and Vaccine Development
Cancer continues to pose a significant and lethal health challenge annually, and tumor immunotherapy has emerged as a promising new strategy for cancer treatment. EVs carry diverse components from their parental cells, including antigens, which renders them potential candidates for tumor immunotherapy. Specifically, numerous studies have demonstrated that dendritic cells (DCs)- and tumor-derived EVs exhibit high expression levels of tumor markers and major histocompatibility complex class I molecules (MHC I), thereby participating in antigen presentation and the stimulation of T cells. These mechanisms have been shown to elicit CD8+ T cell-dependent anti-tumor responses both in vitro and in vivo.43 For instance, tumor-derived exosomes and α-galactosylceramide (α-GalCer)-pulsed DCs have been observed to robustly activate and expand tumor-specific cytotoxic T lymphocytes in a glioblastoma rat model, re-establishing immune surveillance and demonstrating more potent anti-tumor immune responses compared to tumor lysates.44 However, concerns regarding the safety of tumor-derived EVs (TEVs) persist. Several studies have indicated that TEVs play a bidirectional role in tumor immune responses.45
Furthermore, EVs can function as adjuvants to potentiate adaptive immune responses elicited by vaccines. A recent investigation demonstrated that ginseng-derived EV-like particles (G-EVLPs), when co-administered with autologous tumor antigens via membrane fusion, served as effective adjuvants. Specifically, G-EVLPs augmented dendritic cell phagocytosis of tumor antigens, activated tumor-specific cytotoxic T lymphocytes (CTLs), and conferred long-term immune protection.46
Pathogenic microorganisms-derived EVs similarly contain a variety of antigens and pathogen-associated molecular patterns (PAMPs), which confer vaccine-like properties.47 For instance, studies have compared the efficacy of Mycobacterium tuberculosis-derived EVs with that of standard BCG vaccination, demonstrating comparable protective effects against Mycobacterium tuberculosis infection.48 Additionally, Li et al utilized a novel lysin-based approach to enhance the production of outer membrane vesicles (OMVs) from Acinetobacter baumannii. In a mouse model of pneumonia and bacteremia, three intramuscular injections of these OMVs resulted in significant protection against A. baumannii infection.49
EVs as Nutraceuticals
Nutraceuticals have garnered significant attention in recent years. Milk-derived EVs (M-EVs), which possess physiological functions including immunomodulation, maintenance of intestinal and bone homeostasis, and promotion of neural development, represent excellent multifunctional components.50 The processing methods used in milk powder production can influence the composition and integrity of these vesicles. Therefore, supplementing M-EVs into infant formula can help prevent necrotizing enterocolitis and promote neonatal brain development.50 Similarly, given their role in maintaining intestinal and bone homeostasis, M-EVs can be incorporated into functional foods to inhibit osteoporosis and alleviate intestinal inflammation.
Biodistribution of EVs
Elucidating the biodistribution of EVs is critical for their successful clinical application. Extensive research has demonstrated that, due to the activity of the mononuclear phagocyte system, less than 5% of small EVs remain in circulation within five minutes post-intravenous administration.51 These EVs are rapidly distributed to various organs, with predominant accumulation observed in the liver, lungs, spleen, gastrointestinal tract, and kidneys.51,52 Apart from tissue-targeted EVs generated through genetic engineering or chemical modification, the biodistribution of EVs primarily varies based on their size, cellular source, and route of administration. Numerous studies have shown that small EVs predominantly accumulate in the liver, whereas large EVs exhibit a higher propensity for pulmonary accumulation. Small EVs achieve peak concentrations in the liver and kidneys within one hour post-intravenous administration, while peak levels in the lungs and spleen are observed at later time points. Conversely, large EVs attain maximum concentration in the lungs during the first hour post-administration, with increased hepatic accumulation as lung levels decline.51 Furthermore, researchers utilized asymmetric flow field-flow fractionation to isolate two subpopulations of exosomes from B16-F10-derived vesicles: large exosome vesicles (90–120 nm) and small exosome vesicles (60–80 nm). It was found that large exosome vesicles exhibited lymph node tropism following intravenous injection.53
Furthermore, it has been demonstrated that the protein corona, dosage, and isolation methods of EVs also influence their natural biodistribution. Studies have shown that nanoparticles can adhere to proteins in biological fluids to form a “protein corona”, which impacts their in vivo stability, targeting efficacy, cytotoxicity, and immunotoxicity.54 Recent research has identified differences in the protein corona of EVs isolated from 2D versus 3D culture conditions, indicating that EVs derived from 3D cultures enhance hepatocyte uptake and hepatic aggregation.55 Additionally, the dose of EVs has been shown to affect their distribution patterns. For instance, intravenous administration of three different doses of HEK293T EVs revealed a decrease in relative hepatic signal with increasing EV dose, potentially due to saturation of the mononuclear phagocyte system.10 Moreover, isolation methods also affect biodistribution patterns. EVs isolated using ultrafiltration combined with size-exclusion liquid chromatography exhibited significantly reduced lung accumulation compared to those isolated by traditional ultracentrifugation following intravenous administration.56
Among these factors, particular attention will be given to the source and the administration route, which will be elaborated upon in the subsequent sections.
Sources of Vesicles
The Sources of Vesicles Significantly Influence Their Biodistribution
Natural EVs, as bioactive components, exhibit surface receptors that are identical to those of their parent cells. Extensive research has demonstrated that EVs derived from different cellular sources display distinct tropisms. For instance, a study compared the biodistribution of intravenously administered EVs from C2C12 myoblasts, B16F10 melanoma cells, and primary immature bone marrow-derived dendritic cells (DCs). The results indicated that while all EVs showed significant accumulation in the liver, lungs, spleen, and gastrointestinal tract, C2C12-derived EVs exhibited particularly high accumulation in the liver, B16F10-derived EVs accumulated more prominently in the gastrointestinal tract, and DC-derived EVs were preferentially distributed in the spleen.10 Under the same intravenous injection conditions, macrophage-derived EVs exhibited significantly greater accumulation in the liver compared to EVs from mesenchymal stem cells (MSCs) and the melanoma cell line (A375). No significant differences were observed in the distribution of these EVs in the spleen, lung, kidney, muscle, or bone.57 The organ-specific distribution of EVs may be influenced by the membrane components of their parental cells, which partially explains the higher accumulation of macrophage-derived EVs in macrophage-rich tissues such as the liver. Similarly, under identical inhalation administration conditions, human lung spheroid cell (LSC)-derived EVs demonstrated a more pronounced distribution in the lung parenchyma 24 hours post-administration compared to EVs from HEK293T cells. Additionally, LSC-derived EVs appeared later in the liver, spleen, and kidneys and exhibited prolonged retention in the lungs.58 Tumor-derived EVs exhibit a preferential homing tendency to the organs from which their parental cells originate or to sites of tumor metastasis. This phenomenon may be attributed to differential integrin expression profiles.59,60 These findings suggest that in optimizing EVs as drug delivery systems, cell sources can be selectively chosen based on the therapeutic target tissues and the inherent tropism of EVs from different origins for personalized regulation, thereby enhancing accumulation in target tissues. However, the precise mechanisms underlying the intrinsic tropism of EVs derived from endogenous cell sources remain unclear, and the interactions between EVs and receptor cell surfaces warrant further investigation.
Currently Available Vesicle Sources
EVs originate from a diverse array of sources, with virtually any cell type in nature serving as a potential source. These sources encompass animal cells (such as stem cells, tumor cells, immune cells, etc)., plant cells, and microorganisms. However, EVs naturally secreted by cells often exhibit limitations, particularly in terms of low productivity. To address these challenges, researchers have developed extracellular vesicle mimetics (EVMs), including artificial cell-derived vesicles and synthetic vesicles, which play crucial roles in drug delivery applications. Vesicles from different origins possess distinct advantages and disadvantages (Table 1), allowing researchers to select the most suitable cell sources based on their specific research objectives and requirements.
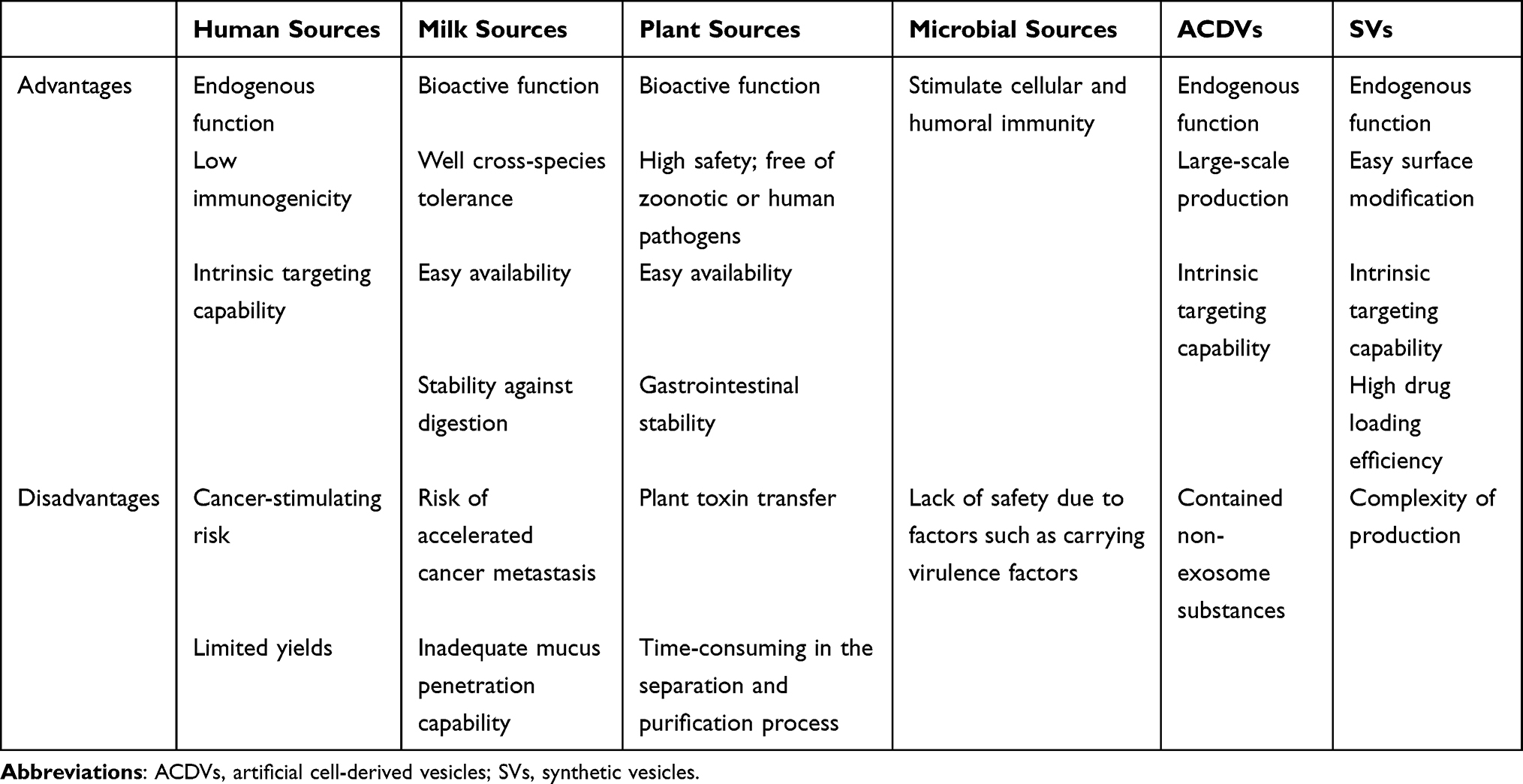 |
Table 1 The Advantages and Disadvantages of Different Sources of EVs |
Animal Sources
Human cell-derived EVs, including those from macrophages, dendritic cells (DCs), neutrophils, B cells, T cells, neurons, oligodendrocytes, tumor cells, and stem cells, have garnered significant attention in clinical applications.61 For instance, EVs derived from monocytes and macrophages inherit the surface components of their parent cells, enabling them to target inflammation sites while evading rapid clearance by the mononuclear phagocyte system, thereby extending their circulation half-life and making them ideal drug carriers for inflammatory diseases.62 Additionally, EVs from dendritic cells play a crucial role in antigen presentation and T-cell stimulation, with research demonstrating their capacity to transfer peptide-MHC complexes to other DCs, thereby enhancing tumor rejection.63 Consequently, these EVs have been extensively investigated for vaccine development and immunotherapy. Neutrophil-derived EVs exhibit chemotactic properties similar to those of neutrophils themselves. In a glioma mouse model, neutrophil-derived EVs loaded with doxorubicin effectively targeted inflamed brain tumors and inhibited tumor growth.64
Given that tumor-derived EVs (TEVs) exhibit a tropism for their parental cells,65 they represent promising candidates as vectors for targeted drug delivery. Compared to EVs derived from HeLa cells, those isolated from HT1080 cells demonstrated a preferential fusion with their parental cancer cells in vitro and exhibited greater accumulation in tumor tissues within a subcutaneous HT1080 tumor-bearing nude mouse model.59 In a murine model involving multiple tumor cell lines inoculated into the pleural cavity, TEVs induced by UVB and loaded with methotrexate significantly reduced pleural tumor burden, the number of pleural lesions, and effusion volume compared to empty TEVs and free methotrexate. These TEVs demonstrated potential efficacy and safety in a clinical trial involving 11 patients with malignant pleural effusion secondary to advanced lung cancer.66 Moreover, TEVs carry a diverse array of proteins and nucleic acids, including antigens from parental tumor cells, which are essential for disease diagnosis and tumor immunotherapy. However, the high heterogeneity of TEVs and the bidirectional tumor immune response raise significant safety concerns, thereby limiting their broader application.
Among all human-derived cells, mesenchymal stem cells (MSCs) are considered the most promising source of EVs. This is attributed to their diverse origins (including umbilical cord, bone marrow, and adipose tissue) and their immunomodulatory and regenerative capacities in various diseases.65 Consequently, MSC-derived EVs represent a viable alternative to cell-based therapies for tissue repair and regeneration. During in vitro passage culture of mesenchymal stem cells (MSCs), both the proliferative and differentiation capacities of MSCs diminish, leading to reduced expansion potential.67,68 Research has demonstrated that EVs from senescent MSCs and those from young MSCs exhibit distinct effects. For instance, EVs from senescent MSCs can elevate senescence markers in young cells, whereas EVs from young MSCs have the opposite effect.69 Numerous studies have explored methods to reverse MSC senescence during expansion, including intrinsic factors such as targeting signaling pathways, small molecule therapies, epigenetic regulation, and extrinsic factors like using paracrine factors to modulate the external environment and altering culture conditions to rejuvenate senescent MSCs.70 Notably, induced pluripotent stem cells (iPSCs) have garnered significant interest due to their unlimited proliferation capacity. Unlike UC-MSCs, which lose their MSC characteristics by the 15th passage, MSCs derived from iPSCs (iMSCs) maintain their MSC properties without chromosomal abnormalities. Furthermore, somatic cells obtained from the patient can be utilized to generate induced pluripotent stem cells (iPSCs), thereby circumventing ethical concerns and mitigating the risk of immune rejection.71 In a mouse model of ischemic stroke, human urine-derived iPSC-derived small EVs (sEVs) specifically targeted the peri-infarct region, leading to the upregulation of neuroprotective genes and the downregulation of inflammation-related genes. Notably, BDNF-loaded sEVs derived from human urine-induced iPSCs demonstrated more pronounced effects on enhancing functional recovery, reducing infarct volume, promoting neurogenesis and angiogenesis, and controlling inflammation.72 However, EVs derived from iMSCs still face the conventional challenges associated with the clinical translation of EVs, and there is an urgent need to enhance their yield. Yuan et al demonstrated a significant increase in the yield of EVs derived from mesenchymal stem cells (MSCs) through three-dimensional dynamic culture, which not only boosted their immunomodulatory potential but also presented a promising approach for enhancing the yield of EVs from MSCs.73 It remains to be explored whether this method can similarly improve both the yield and functionality of EVs derived from iMSCs, making it a worthwhile area for further investigation.
In addition to human-derived EVs, EVs from other animal sources, particularly milk-derived EVs (M-EVs), have garnered significant attention from researchers. This interest is driven by their ease of availability, safety, stability against digestion, and compatibility with tissue absorption.50,74 Moreover, M-EVs exhibit immunomodulatory properties, contribute to the maintenance of intestinal and bone homeostasis, support neurodevelopment, and possess antifibrotic and anticancer activities, making them suitable for functional products and oral delivery systems.50 For example, orally administered paclitaxel-loaded M-EVs exhibited significantly enhanced tumor growth suppression and demonstrated substantially reduced systemic toxicity and immunotoxicity compared to intraperitoneal administration of paclitaxel.75 Notably, Samuel et al observed that oral administration of M-EVs reduced the primary tumor burden in mouse models of breast and pancreatic cancer while simultaneously accelerating tumor metastasis.76 Nevertheless, several studies have demonstrated that natural M-EVs lose approximately 85% of their protein content in simulated gastric environments and encounter significant challenges posed by the mucus-epithelial barrier.77 To address these issues, researchers have employed surface modifications using hydrophilic and zwitterionic peptides to enhance the stability and intestinal epithelial absorption of M-EVs.77 Further investigation is required to elucidate the potential and mechanisms of M-EVs in crossing gastrointestinal barriers. Engineering EVs to improve their gastrointestinal stability is essential for the realization of oral EV-based drug delivery systems. Furthermore, EVs derived from various animal sources have been extensively investigated. Notably, EVs isolated from oyster mantle have demonstrated the ability to promote osteoblast differentiation while simultaneously inhibiting osteoclast differentiation. This dual effect has been shown to effectively alleviate osteoporosis in ovariectomized mice following oral administration.78
Plant Sources
Plant Exosome-like Nanovesicles (PELNVs) were initially identified in plant cells in 1967 but have received limited attention until recently.79 Over the past decade, significant advancements have been made in understanding their diverse biological effects, including anti-tumor, immunomodulatory, and regenerative properties, positioning PELNVs as promising therapeutic agents. For instance, ginger-derived PELNVs have demonstrated anticancer potential by reducing cyclin D1 mRNA levels in a mouse model of colorectal cancer.80 Turmeric-derived PELNVs have alleviated colitis through the regulation of antioxidant gene HO-1 and pro-inflammatory cytokines such as IL-6, IL-1β, and TNF-α, showcasing their anti-inflammatory and antioxidant capabilities.81 Wheat-derived PELNVs have promoted skin regeneration and angiogenesis.82 Compared to animal-derived EVs, PELNVs offer several advantages: they are readily available from a wide range of sources (eg, carrot, lemon, citrus, ginger, grapefruit, wheat, and broccoli), enabling large-scale production at lower economic costs. Moreover, PELNVs exhibit lower toxicity and higher safety profiles, avoiding potential risks associated with animal-derived EVs, such as tumor stimulation and zoonotic or human pathogens.33 Studies have also shown that PELNVs can withstand harsh gastrointestinal environments, resisting digestive enzymes (eg, pepsin, trypsin, and bile extract solution) and strong gastric acid conditions, highlighting their potential for oral drug delivery.30 For example, turmeric-derived EVs remained stable in simulated gastric (pH ~2.0) and intestinal (pH ~6.5) solutions, accelerating colitis remission when administered orally.81 However, it is important to address potential risks, such as the transmission of plant toxins like trichosanthin and allergens, which could pose challenges for clinical translation.33 As emerging therapeutic agents, limited information is available regarding the specific mechanisms of biogenesis, release, and function of PELNVs, which poses significant challenges to their clinical translation. Moreover, the isolation and purification processes for PELNVs, which are considerably more time-consuming compared to those of EVs derived from cell cultures, represent a critical area requiring substantial improvement for large-scale production.33
Bacterial and Other Microbial Sources
The bacterial outer membrane vesicles (OMVs) play a crucial role in mediating both bacteria-bacteria and bacteria-host interactions. These interactions encompass modulating host immune responses, enhancing antibiotic resistance, and influencing bacterial pathogenicity.83 The cargo transported by OMVs, which includes proteins and nucleic acids, can reflect the physiological or pathological states of the host. Consequently, OMVs serve as potential sources of specific biomarkers for various diseases, including both tumor and non-tumor conditions such as psychiatric disorders, abdominal obesity, allergic rhinitis, periodontitis, and atopic dermatitis.84 Furthermore, OMVs carry a diverse array of pathogen-associated antigens, effectively eliciting robust cellular and humoral immune responses, which positions them as promising candidates for vaccine development. For instance, Jiang et al engineered a novel SARS-CoV-2 vaccine by conjugating the mammalian cell culture-derived Spike receptor-binding domain (RBD) to EVs derived from Salmonella typhimurium. This intranasally administered vaccine induced neutralizing antibodies against both the wild-type and Delta variants, providing effective protection against SARS-CoV-2.85 The immunosuppressive tumor microenvironment has posed a significant challenge to tumor immunotherapy. Researchers have leveraged the immunostimulatory properties of OMVs to reprogram the tumor microenvironment by inducing M1 polarization of macrophages.86,87 However, the intrinsic immunotoxicity associated with OMVs derived from bacteria has considerably impeded their clinical application. A recent study addressed this issue by constructing attenuated OMVs through gene editing. This approach eliminated the inflammatory response triggered by flagellin and lipopolysaccharide on the OMVs surface while preserving its immunostimulatory function, thereby presenting a promising strategy to enhance the therapeutic potential of OMVs.88 Recently, research into fungal and parasitic EVs has expanded. However, concerns regarding their safety and therapeutic efficacy have been raised due to factors such as the potential carriage of virulence factors.83
Artificial Cell-Derived Vesicles (ACDVs)
Artificial cell-derived vesicles (ACDVs) are predominantly generated through continuous cell extrusion, enabling large-scale production with stability, distribution, and immune compatibility that closely resemble those of natural EVs.65 Fan et al have demonstrated high-yield production of EV mimetics by extruding human mesenchymal stem cells (hMSCs) through polycarbonate membrane filters with progressively decreasing pore sizes. Compared to naturally derived hMSC-EVs, these extruded EV mimetics exhibit comparable characteristics and biological internalization capabilities, while demonstrating enhanced osteogenic effects.89 However, the researchers also observed that EV mimetics contain various non-exosomal components, including GM-130, cytochrome c, and calnexin. In a recent study, immunosuppressive microvesicle mimetics (MVMs) were developed by extruding endotoxin-resistant dendritic cells and conjugating αCD3 antibodies to the vesicles to specifically target T cells. The engineered MVMs demonstrated a significant capacity to induce regulatory T cells (Tregs) and polarize anti-inflammatory M2 macrophages, leading to rapid and efficient immunosuppression in both sepsis and rheumatoid arthritis (RA) models. These findings suggest that MVMs may represent a promising therapeutic option for both acute and chronic inflammatory diseases.90
Synthetic Vesicles (SVs)
Natural EVs exhibit low drug loading efficiency,91 while liposomes do not possess the endogenous functions and low immunogenicity characteristics of natural EVs. Scientists have addressed these limitations by fusing the membranes of EVs and liposomes to construct hybrid vesicles that possess low immunogenicity, high drug loading efficiency, ease of surface modification, inherent targeting capabilities, and enhanced stability.92 Goh et al pioneered the fusion of EVs with liposomes to create EXOPLEX, which showed doxorubicin hydrochloride loading efficiencies exceeding 65%. Compared to liposomes or EVs alone, EXOPLEX exhibits superior in vitro cytotoxic effects and is anticipated to represent a new generation of engineered vesicles for efficient drug delivery.93 Moreover, tumor-derived EVs, known for their ability to target parental cancer cells,59 have been extensively studied for fusion with liposomes, emerging as promising tools for precise tumor targeting.94,95 In a recent study, Li et al developed hybrid vesicles by fusing hematopoietic stem and progenitor cell (HSPC) membranes with liposomes to specifically target the bone marrow. In a mouse model of leukemia, these hybrid vesicles facilitated targeted delivery of cytarabine to the bone marrow via the hyaluronan-CD44 axis, leading to a significant reduction in leukemic stem cell burden.96 Furthermore, the integration of thermosensitive, photosensitive, and pH-sensitive liposomes endowed the hybrid vesicles with controlled release capabilities, thereby enhancing drug delivery efficiency and mitigating adverse effects. For instance, synthetic vesicles generated by fusing macrophage-derived EVs with pH-sensitive liposomes enhanced the release of encapsulated doxorubicin in acidic microenvironments, thereby improving drug delivery to acidic tumor environments.42 However, the practical application of these hybrid vesicles is currently constrained by the complexity of their fabrication process and the challenges associated with large-scale production.
Administration Routes of EVs
The Administration Routes of EVs Influence Their Biodistribution
EVs can be administered through various routes including intravenous, intraperitoneal, oral, intranasal, subcutaneous, or local administration (eg, intratumoral, intra-articular). Numerous studies have demonstrated that different administration routes significantly influence the biodistribution of EVs (Figure 2). For instance, Wiklander et al compared tissue distribution 24 hours post-administration of HEK293T-derived EVs via intravenous, intraperitoneal, and subcutaneous injections. Their findings indicated that intravenously administered EVs accumulated more in the liver and spleen, whereas intraperitoneally and subcutaneously administered EVs exhibited less accumulation in these organs and a significantly greater presence in the gastrointestinal tract and pancreas.10 Manca et al reported that orally administered milk-derived EVs (M-EVs) predominantly accumulated in the intestines and liver, while intravenously administered M-EVs were mainly distributed to the liver and spleen.97 Twenty-four hours post-intravenous injection, mesenchymal stem cell (MSC)-derived EVs were predominantly localized in the liver, spleen, and lung. In contrast, EVs administered via nasal delivery primarily accumulated in the lung, with reduced hepatic accumulation.13 In an additional study, nebulized inhalation of MSC-derived EVs resulted in the predominant distribution in the lung and stomach after 24 hours.98 These studies suggest that optimizing routes of administration to enhance tissue distribution and improve EV delivery efficiency is a viable strategy. In the subsequent section, we provide a detailed summary and comparison of the various administration routes and their corresponding distribution profiles in EV applications.
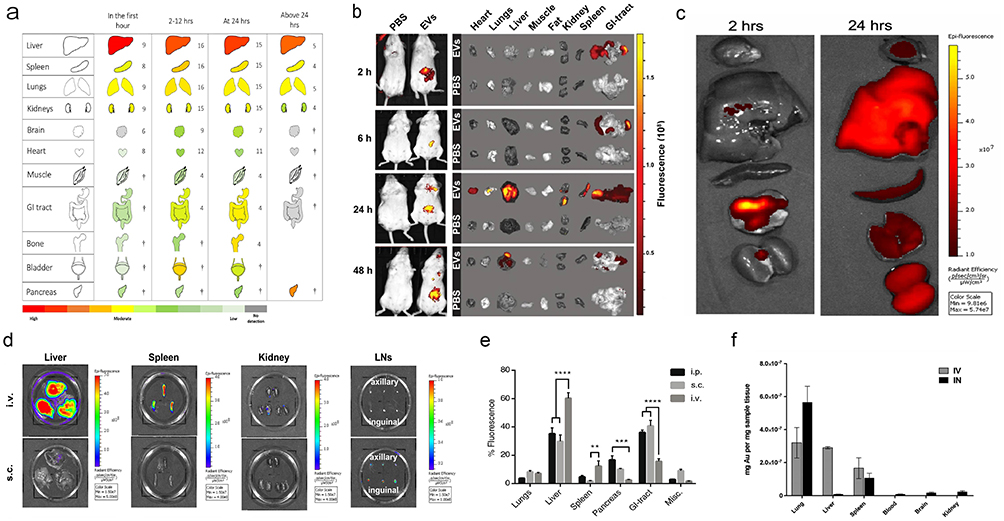 |
Figure 2 Representative examples of the biodistribution of EVs via various administration routes (a) Ex vivo organ distribution of small EVs following intravenous injection, summarized from 36 studies. The numbers adjacent to each organ indicate the number of studies for that organ, with organs marked by † represented in three or fewer studies. Reprinted with permission from Kang M, Jordan V, Blenkiron C, Chamley LW. Biodistribution of extracellular vesicles following administration into animals: a systematic review. J Extracell Vesicles. 2021;10(8):e12085. Copyright 2021, John Wiley and Sons.51 (b) In vivo imaging of mice post oral administration of milk-derived EVs. Reprinted with permission from Samuel M, Fonseka P, Sanwlani R, et al. Oral administration of bovine milk-derived extracellular vesicles induces senescence in the primary tumor but accelerates cancer metastasis. Nat Commun. 2021;12(1):3950. Copyright 2021, Springer Nature.76 (c) Organ distribution after inhalation delivery of human lung spheroid cell (LSC)-derived EVs. Reprinted with permission from Wang Z, Hu S, Popowski KD, et al. Inhalation of ACE2-expressing lung exosomes provides prophylactic protection against SARS-CoV-2. Nat Commun. 2024;15(1):2236. Copyright 2024, Springer Nature.58 (d) Organ distribution of melanoma-derived small EVs following subcutaneous and intravenous injections. Reprinted from Journal of Controlled Release, Sakurai Y, Ohtani A, Nakayama Y, et al. Logistics and distribution of small extracellular vesicles from the subcutaneous space to the lymphatic system. J Control Release. 2023;361:77–86., Copyright 2023, with permission from Elsevier.99 (e) Percentage of fluorescence signals in various organs after intraperitoneal, subcutaneous, and intravenous injections of HEK293T EVs. Misc.=brain, heart, kidneys and quadriceps muscle. N=3-4, **represents p<0.01, ***p<0.001 and ****p<0.0001. Reprinted with permission from Wiklander OPB, Nordin JZ, O’Loughlin A, et al, Extracellular vesicle in vivo biodistribution is determined by cell source, route of administration and targeting. J Extracell Vesicles. 2015;4(1):26316. © 2015 Oscar P. B. Wiklander et al10 (f) Quantitative comparison of MSC-derived small EVs delivered intranasally versus intravenously. Reprinted with permission from Betzer O, Perets N, Angel A, et al, In vivo neuroimaging of exosomes using gold nanoparticles. ACS Nano. 2017;11(11):10883–10893. Copyright 2017, American Chemical Society.13 |
The Biodistribution and Applications of EVs via Diverse Administration Routes
Intravenous Injection
Intravenous injection remains the predominant method for delivering EVs. This route allows EVs to directly enter the circulatory system, thereby enabling precise regulation of their concentration and dosage within the bloodstream. Following intravenous administration, EVs predominantly accumulate in organs such as the liver, lungs, spleen, and kidneys.51 Most studies investigating EV biodistribution have relied on imaging techniques. Vos et al conducted proteomic analysis to examine the presence of EV-derived proteins in various tissues at 1-hour and 24-hour intervals post-injection of adipose-derived mesenchymal stem cell EVs. Their findings indicated that 24 hours post-injection, EV-derived proteins were primarily localized in the liver, corroborating previous observations made using imaging methodologies.12
EVs administered via intravenous injection have diverse applications across a range of conditions, including metabolic disorders, ischemic events (such as stroke), osteoarticular pathologies, and oncological diseases, among others. Sun et al demonstrated that the intravenous administration of EVs derived from human mesenchymal stem cells (MSCs) reversed peripheral insulin resistance by restoring the phosphorylation of insulin receptor substrate 1 and protein kinase B. Concurrently, this treatment alleviated pancreatic β-cell damage, leading to a reduction in blood glucose levels in rats with type 2 diabetes.100 In the rat model of doxorubicin and trastuzumab-induced cardiotoxicity, intravenous administration of EVs derived from human cardiac-resident mesenchymal progenitor cells (CPCs) significantly attenuated myocardial oxidative stress and cardiotoxicity. This effect was attributed to the high enrichment of miR-146a-5p within the vesicles.101 As circulating EVs have demonstrated the ability to cross the blood-brain barrier (BBB), intravenously administered EVs are extensively utilized for drug delivery and as functional components in cerebral ischemia, glioma, and other neurological applications. For instance, researchers developed a curcumin delivery vehicle derived from bone marrow mesenchymal stem cell (MSC)-derived EVs conjugated with c(RGDyK), targeting the ischemic region of the brain. Following intravenous injection, this construct significantly reduced the levels of pro-inflammatory factors (TNF-α, IL-6, and IL-1β), thereby inhibiting post-ischemic brain inflammation more effectively than free curcumin.102 In the context of osteoarticular diseases, Ma et al developed hybrid EVs by fusing M2 macrophage-derived EVs with bone marrow mesenchymal stem cell-derived EVs (M2-BMSCs-Exos). Intravenous administration of M2-BMSCs-Exos targeted areas of osteolysis following joint replacement surgery, thereby inhibiting M1 polarization and promoting M2 polarization. This approach represents a promising strategy for potentially extending the lifespan of prosthetic implants in the future.62
Nonetheless, a substantial body of research demonstrates that EVs exhibit a markedly short half-life following intravenous administration.51 To address this limitation, researchers have employed strategies such as coating nanoparticles with polyethylene glycol (PEG). Additionally, incorporating molecules associated with the “don’t eat me” signal (eg, CD47, CD31, CD24) into the EV membrane has been explored. These modifications aim to reduce clearance by the mononuclear phagocyte system, thereby prolonging circulation half-life and enhancing the delivery efficiency of EVs to target tissues.52
Intraperitoneal Injection
Intraperitoneal injection leverages the extensive abdominal vasculature, providing a large absorption area and bypassing first-pass metabolism in the gastrointestinal tract. This route also accommodates larger drug volumes, making it a subject of extensive research.103 For instance, studies have shown that milk-derived EVs loaded with a suboptimal dose of the anticancer drug WFA, when administered intraperitoneally to mice bearing human lung cancer (A549) xenografts, exhibited superior tumor inhibition compared to free WFA.104 Another study proposed that intraperitoneal injection represents a promising therapeutic option for obesity, as it can effectively deliver EVs to visceral adipose tissue, in addition to the liver, lungs, and spleen.105 According to the research by Wiklander et al, EVs administered via intraperitoneal injection exhibited comparable accumulation in the gastrointestinal tract to that observed in the liver, followed by the pancreas, indicating that this route of administration could be efficacious for treating gastrointestinal diseases.10 For example, intraperitoneal injection of adipose-derived mesenchymal stem cells (MSCs) has been shown to alleviate symptoms of dextran sulfate sodium-induced acute colitis by reducing inflammatory factor levels and modulating the Treg population.106
Oral Administration
Oral administration remains the preferred route for treating various diseases owing to its cost-effectiveness, ease of use, and high patient compliance. However, the presence of enzyme degradation and strong acidic conditions in the gastrointestinal tract, along with mucosal barriers, markedly diminishes the bioavailability of orally administered drugs.107,108 Extensive evidence indicates that EVs derived from diverse sources such as plants, milk, intestinal epithelial cells, and bacteria can endure harsh gastrointestinal environments. This resilience positions EVs as promising candidates for oral drug delivery systems (Table 2). For example, EVs derived from human milk (M-EVs) have been shown to withstand simulated gastric and pancreatic digestion.108 Additionally, curcumin-loaded M-EVs exhibit resistance to degradation by human digestive enzymes.109 Researchers initially labeled M-EVs with the lipophilic near-infrared fluorescent dye DiR. They observed that the highest fluorescence signal in the bloodstream was detected at 24 hours post-administration and persisted in circulation until day 6. Furthermore, ex vivo organ imaging conducted on day 4 after oral administration revealed a relatively uniform distribution of EVs across multiple organs, including the liver, lungs, kidneys, pancreas, spleen, ovaries, colon, and brain. This contrasts with the predominant liver accumulation observed following intravenous injection.104 EVs exhibit widespread distribution across multiple organs, offering a more cost-effective and simpler alternative to intravenous administration, which has garnered significant attention from researchers. A previous study demonstrated that M-EVs were predominantly localized in the intestines and liver 24 hours post-oral administration, whereas intravenously administered M-EVs were primarily confined to the liver and spleen.97 Furthermore, subsequent studies have consistently observed the accumulation of EVs within the gastrointestinal tract following oral administration.110,111 Oral administration of EVs has demonstrated superior intestinal distribution compared to intravenous administration and holds the potential for enhancing the targeting of intestinal diseases.
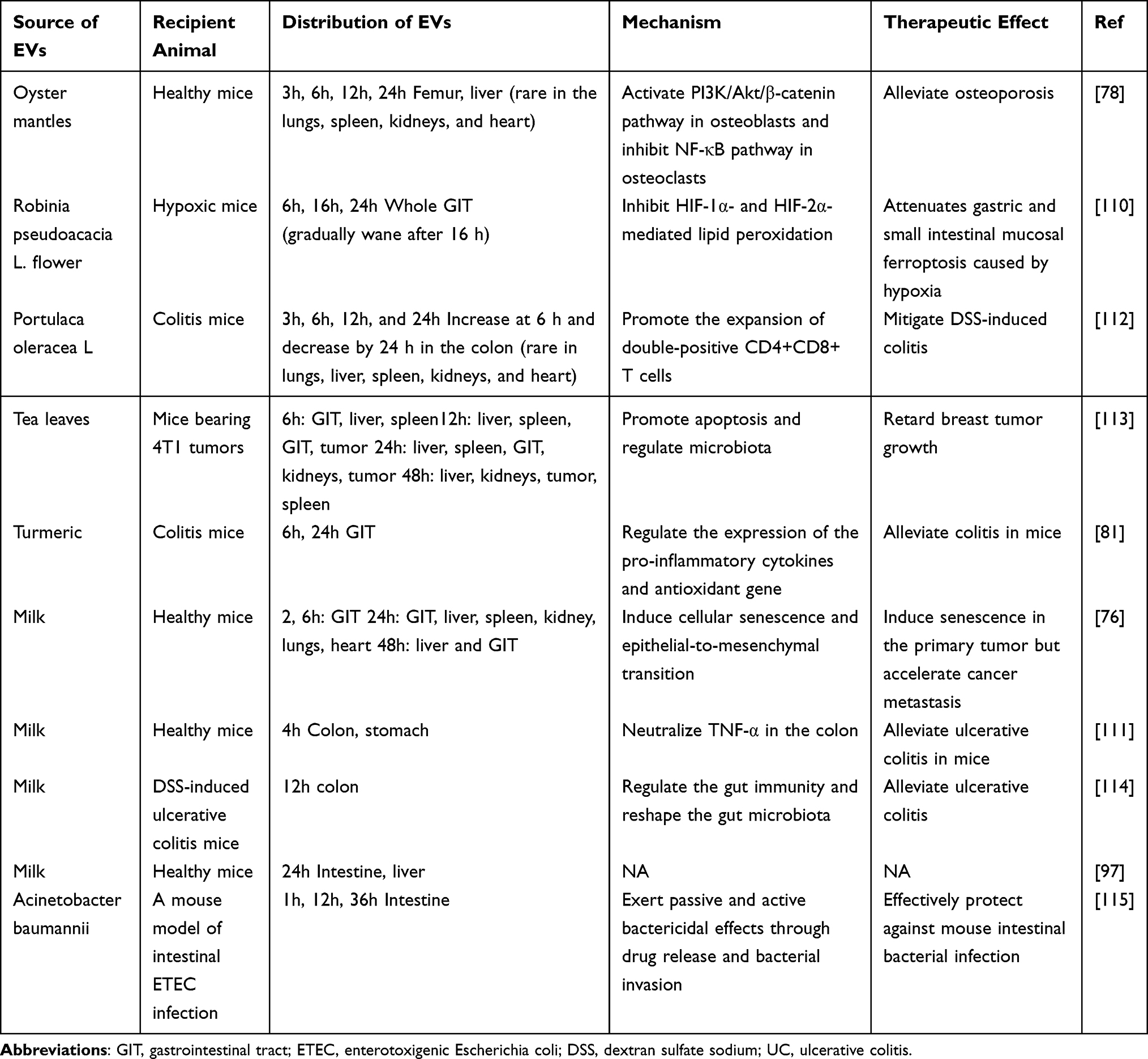 |
Table 2 Typical Examples of Oral Delivery of EVs in Recent years |
Since Munagala et al first demonstrated in 2016 that milk-derived EVs (M-EVs) are promising carriers for chemotherapeutic drugs and can be used for oral administration,104 researchers have been increasingly motivated to explore the potential of M-EVs in oral drug delivery. For instance, paclitaxel (PAC) exhibits limited water solubility, which restricts its oral bioavailability. Nevertheless, the oral administration of M-EVs loaded with PAC demonstrates superior anti-tumor efficacy compared to intraperitoneal administration of a lower dose of free PAC. Additionally, it exhibits reduced systemic and immune toxicity relative to intravenous administration of a lower dose.75 Similarly, oral administration of curcumin-loaded M-EVs resulted in 3–5 times higher curcumin levels in various organs and stronger inhibition of cervical tumor xenografts compared to free curcumin.116 A recent study demonstrated that VHHm3F-loaded M-EVs accumulated predominantly in the colon after four hours of oral administration, enhancing VHHm3F activity and alleviating inflammation in mice with acute ulcerative colitis compared to free VHHm3F.111 Notably, oral administration of insulin-loaded M-EVs showed better and more sustained hypoglycemic effects in type 1 diabetic rats compared to subcutaneous insulin injection.117
Plant-derived EVs, owing to their abundant availability, low immunogenicity, edibility, and inherent physiological effects, have garnered significant attention as oral drug delivery systems. Various plant sources have been explored, including ginger,80 turmeric,81 broccoli,118 garlic,119 grapefruit,36 tea flowers,120 and Robinia pseudoacacia L. flowers.110 For instance, Wang et al demonstrated that the antioxidant properties of Robinia pseudoacacia L. flower exosome-like nanoparticles (RFELNs) led to the inhibition of HIF-1α and HIF-2α-mediated peroxidation, thereby mitigating hypoxia-induced gastric and small intestinal mucosal damage following oral administration. Moreover, RFELNs were observed to accumulate predominantly in the gastrointestinal tract at 6, 16, and 24 hours post-administration, positioning them as promising candidates for natural drugs targeting this system.110 Beyond gastrointestinal applications, oral administration of garlic-derived EVs has shown preferential uptake by microglia in mouse models of high-fat diet-induced obesity. These EVs effectively inhibited brain inflammation through the IDO1-mediated AHR pathway and the c-Myc-mediated c-GAS/STING inflammatory pathway, reversing obesity and insulin resistance induced by a high-fat diet.119 Additionally, a comparative study on tea leaf-derived exosome-like nanotherapeutics (TLNTs) revealed comparable anti-tumor efficacy between oral and intravenous administration routes, with no hepatorenal toxicity observed in oral TLNTs compared to intravenous administration.113
Intestinal epithelial cell-derived EVs (IEC-EVs) have been investigated to enhance the oral bioavailability of drugs. Compared with free curcumin, curcumin-loaded IEC-EVs significantly improved cellular uptake and intestinal permeability in a Caco-2 cell model, demonstrating superior performance over M-EVs.121 Additionally, bacterial outer membrane vesicles (OMVs) have been explored for oral drug delivery applications. For instance, Huang et al developed a novel antibiotic delivery system utilizing the drug efflux resistance mechanism of OMVs. Compared with free antibiotics, OMV-encapsulated antibiotics exhibited prolonged retention in the intestine for at least 36 hours, enhanced bactericidal efficacy in a murine model of enterotoxigenic Escherichia coli-induced intestinal infection, and mitigated adverse effects on gut microbiota.115
Intranasal Administration
A substantial body of research demonstrates that intranasal administration of EVs significantly enhances their delivery to the brain. Intranasal drug delivery primarily facilitates the transport of drugs across a single epithelial cell layer into the systemic circulation or bypasses the blood-brain barrier via the olfactory and trigeminal nerves, allowing direct entry into the brain.34,122 Zhuang et al observed diffuse fluorescently labeled EVs in the brain within 30 minutes post-intranasal administration, indicating rapid translocation of EVs to the brain.123 Betzer et al found that mesenchymal stem cell (MSC)-derived EVs predominantly accumulated in the lungs 24 hours after intranasal administration, whereas intravenously administered EVs were mainly localized in the liver, lungs, and spleen. Notably, intranasal administration of MSC-derived EVs exhibited superior brain accumulation at both 1 and 24 hours compared to intravenous administration.13 Consistent with previous findings, Wang et al demonstrated that DiR-labeled grapefruit-derived EVs delivered intranasally were primarily distributed to the lungs and brain and remained stable in the brain.28 Intranasal delivery of EVs offers advantages such as rapid onset, non-invasiveness, and the ability to bypass hepatic and intestinal metabolism,124 thereby providing high bioavailability. This makes it a promising route for treating central nervous system (CNS) diseases (Table 3), including brain inflammatory diseases,123 brain tumors,125 cerebral ischemia,72 Parkinson’s disease,126 Alzheimer’s disease,127 and spinal cord injuries.128
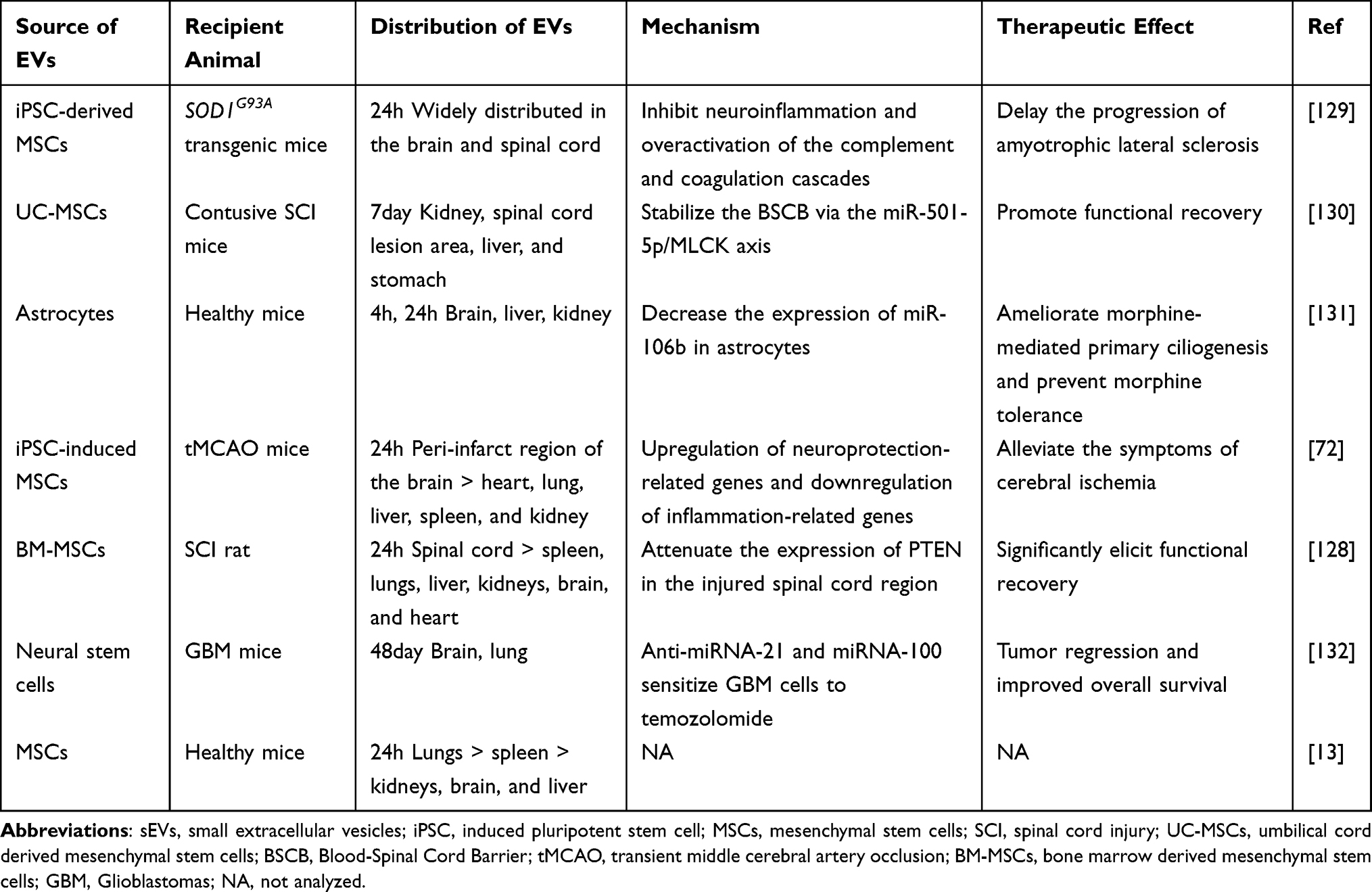 |
Table 3 Typical Examples of Intranasal Delivery of EVs in Recent years |
For instance, intranasally administered EVs were observed to be absorbed by microglia. EVs loaded with curcumin effectively inhibited the progression of lipopolysaccharide-induced brain inflammation and myelin oligodendrocyte glycoprotein-induced experimental autoimmune encephalomyelitis. Additionally, EVs carrying a Stat3 inhibitor significantly inhibited the growth of brain tumors.123 Notably, no particles larger than exosomes (30–100 nm), specifically those in the range of 500 nm to 1 μm, were detected within the brain, suggesting that EV particle size is a critical determinant for translocation into the brain via intranasal administration. In a separate study, researchers utilized folic acid ligand-modified grapefruit-derived nanovesicles (GNVs) to target folate receptor-positive GL-26 brain tumors. miR17-loaded GNVs were rapidly transported to the brain and effectively inhibited tumor growth.125 Selective targeting of the peri-infarct region was observed in a mouse model of cerebral ischemia 24 hours following intranasal administration of human urine-derived iPSC-derived small EVs (sEVs) or HEK293 cell-derived sEVs, as compared to minimal signals detected in other organs such as the heart, lung, liver, spleen, and kidney. Furthermore, sEVs loaded with brain-derived neurotrophic factor (BDNF) were found to upregulate neuroprotective genes, downregulate inflammatory genes post-administration, and activate BDNF/TrkB signaling pathways in ischemic stroke models. Consequently, these sEVs significantly improved functional outcomes, reduced infarct volume, promoted neurogenesis and angiogenesis, and enhanced synaptic plasticity and fiber preservation.72 Intranasal administration of EVs has garnered significant attention from researchers studying neurodegenerative diseases such as Parkinson’s disease34,126 and Alzheimer’s disease.127 Peng et al developed self-oriented nanocarriers termed PR-EXO/PP@Cur, which integrate bone marrow MSC-derived EVs with curcumin. These PR-EXO/PP@Cur nanocarriers were engineered to enhance membrane translocation and target diseased dopaminergic neurons following intranasal administration. This approach significantly improved motor and coordination abilities in Parkinson’s disease model mice by reducing α-synuclein aggregates, promoting neuronal function, and mitigating neuroinflammation.126 Another study on Parkinson’s disease demonstrated that macrophage-derived EVs accumulated more extensively in brain tissue four hours post-intranasal administration compared to intravenous injection. Intranasal delivery of catalase-loaded EVs markedly reduced microglial activation in 6-OHDA-intoxicated mice, offering robust neuroprotection, whereas catalase alone failed to reduce inflammation in Parkinson’s disease mice.34 Furthermore, EVs can traverse the blood-brain barrier (BBB) to reach the spinal cord region via intranasal administration. Guo et al demonstrated that intranasal delivery of MSC-EVs loaded with phosphatase and tensin homolog (PTEN) siRNA reduced PTEN expression in damaged spinal cord regions, promoted axonal growth and neovascularization, mitigated neuroinflammation and gliosis, thereby significantly enhancing functional recovery in rats with complete spinal cord injury.128 This approach appears promising as a clinical option for promoting recovery in patients with spinal cord injuries.
In addition to central nervous system diseases, intranasal EV delivery provides a suitable route for respiratory diseases due to lung accumulation. Intranasal administration of macrophage-derived EVs loaded with paclitaxel to Lewis lung carcinoma mice demonstrated significant anti-tumor efficacy, with 97.9 ± 2.0% of the EVs colocalizing with lung metastases.133
However, intranasal delivery is constrained by the mucosal layer and mucociliary clearance mechanisms, resulting in suboptimal absorption efficiency.124 To address this limitation, integrating EVs with advanced biomaterials to prolong nasal retention time appears promising. Wang et al developed a gel system comprising a glutamine amide derivative and benzaldehyde ligands conjugated with endothelial cell-derived EVs for intranasal administration. This hydrogel forms a viscous liquid that coats the nasal mucosa, facilitating prolonged contact between EVs and the nasal epithelium, thereby extending retention time and enhancing delivery efficiency. In a murine model of myocardial ischemia/reperfusion injury, this system exhibited superior anti-inflammatory properties and protection of endothelial function compared to EVs alone.134 Additionally, Sukumar et al suggested that reducing respiratory rate can minimize intranasal nanoparticle accumulation in the respiratory tract when targeting brain diseases. Under deep anesthesia, a slower respiratory rate increased the retention duration of nanoparticles in the nasal mucosa and improved their absorption efficacy relative to light anesthesia.135
Subcutaneous Injection
Subcutaneous injection, due to its operational simplicity and potential for patient self-administration, has garnered significant interest in EV delivery. However, the biodistribution of subcutaneously administered EVs exhibits variability. In early studies, Wiklander et al performed an ex vivo analysis of the biodistribution of DiR-labeled EVs derived from HEK293T cells 24 hours post subcutaneous injection. The findings revealed that EVs were predominantly localized in the gastrointestinal tract, with a secondary accumulation observed in the liver. Notably, the distribution of EVs in lymph nodes was not evaluated.10 Other studies have reported that a substantial proportion of EVs were delivered to lymph nodes. Royo et al observed considerable accumulation of 124I-labeled MLP29-derived EVs in both proximal and distal lymph nodes 72 hours after subcutaneous injection via the hoof.136 Yu et al assessed the organ distribution of melanoma-derived small EVs (sEVs) following either tail vein intravenous or tail skin subcutaneous injection. Their findings indicated that subcutaneously injected sEVs consistently accumulated primarily in lymph nodes at 6, 24, 48, and 168 hours, whereas intravenously injected sEVs were mainly distributed to the liver and spleen.99 To verify that lymphatic delivery of subcutaneously injected sEVs is a common phenomenon, the researchers examined the distribution of sEVs derived from additional cell lines, including melanoma YUMM1.7, mouse mammary carcinoma 4T1, macrophages, and lymphatic endothelial cells. Consistent results were observed across these different sources. The researchers confirmed that subcutaneously injected sEVs preferentially accumulate in lymph nodes via the lymphatic system, primarily regulated by surface membrane proteins.99 These findings suggest that targeting the lymphatic system with subcutaneous sEV delivery is feasible, particularly for immunotherapy applications, similar to liposome-based approaches. For instance, lipid-coated magnetite micelles have been shown to efficiently deliver antigens (OVA) and TLR9 agonists (CpG) to lymph nodes via subcutaneous injection, thereby enhancing antigen-specific immune responses.137 Furthermore, subcutaneous injection of sEVs has been widely utilized in dermatology. Subcutaneous administration of human umbilical cord blood-derived sEVs into skin wounds in mice reduced scar width and promoted angiogenesis, accelerating wound healing.138 Notably, another study reported that mice treated with subcutaneously injected sEVs from adipose-derived MSCs exhibited slower wound healing compared to the intravenous group.139
Nevertheless, conventional subcutaneous administration necessitates frequent drug dosing due to the limited retention duration of EVs within the body. Microneedles have attracted considerable attention for the subcutaneous delivery of EVs owing to their high penetration efficiency, minimal pain, reduced infection risk, and absence of psychological burden.140 For instance, Yang et al engineered a detachable microneedle patch composed of keratin derived from hair to co-deliver human bone marrow MSC-derived EVs and the small-molecule drug UK5099. This approach demonstrated superior efficacy in promoting hair growth compared to subcutaneous injections of EVs or topical application of UK5099.141
Inhalation Administration
Inhalation is a widely investigated route of drug delivery due to the lungs’ extensive vascular network, large surface area, and relatively permeable mucosal barrier.142 Moreover, drugs absorbed into the pulmonary circulation bypass first-pass metabolism by entering the systemic circulation directly via the pulmonary veins. Common devices for inhalation administration include nebulizers, pressurized metered-dose inhalers (pMDIs), and dry powder inhalers (DPIs). While nebulizers are cost-effective, they may compromise the integrity of lipid-based nanocarriers, leading to leakage of encapsulated molecules. In contrast, DPIs offer portability and enhanced stability of the dry powder formulation.143 Similar to intranasal delivery, mucus in the airway lining can clear drug particles. However, inhalation administration’s noninvasive nature and ability to achieve high lung deposition make it suitable for delivering EVs for respiratory diseases.144 In a recent study, Wang et al observed that human lung spheroidal cell-derived EVs (LSC-Exo) expressing ACE2 were deposited throughout the lungs at three distinct time points (2 hours, 4 hours, and 24 hours) following nebulized inhalation. These inhaled LSC-Exo bound to and neutralized SARS-CoV-2, leading to a significant reduction in lung inflammation and viral load, thereby effectively preventing SARS-CoV-2 infection.58 Dinh et al demonstrated that nebulized inhalation of LSC-derived secretome and EVs could re-establish normal alveolar structure, and reduce collagen deposition and myofibroblast proliferation in a model of bleomycin- and silica-induced fibrosis, thus restoring lung function. In another study.145 Khaled et al developed dry powders of B-cell-derived EVs loaded with miR-335 using thin-film freeze-drying technology. This formulation can down-regulate SOX4 overexpression in breast cancer and holds potential for treating primary lung cancers and lung metastases associated with SOX4 overexpression.143
Intra-Tumor Injection
The vascular system of tumors exhibits a higher propensity for leakage compared to healthy blood vessels, which facilitates the penetration of nanoparticles into the tumor stroma and results in significant retention within tumor tissue. Several studies have demonstrated that direct injection of EVs into tumors leads to superior retention compared to intravenous administration, which is associated with rapid clearance from the circulation.51 A clinical trial involving 20 patients with end-stage extrahepatic cholangiocarcinoma and malignant biliary obstruction administered methotrexate-loaded tumor-derived EVs via percutaneous transhepatic biliary drainage directly into the tumor lumen. This intra-tumoral injection of methotrexate-loaded EVs not only directly destroyed tumor cells but also recruited a substantial number of neutrophils, thereby triggering neutrophil-mediated anti-tumor activity. Consequently, five cases experienced relief from obstruction, including one case with remission lasting up to 5 months.146 Although intra-tumoral EV injection reduces toxicity to non-target tissues and enhances tumor clearance, this method is more invasive and less acceptable to patients compared to systemic administration, particularly for deep-seated tumors.147
In addition, Sakurai et al observed a substantial accumulation of B16F0-derived small EVs (sEVs) in the lymph nodes following intra-tumoral injection, which was significantly higher compared to that in the liver and spleen. This suggests that lymphatic transit may be a prevalent phenomenon after intra-tumoral administration of sEVs, potentially offering a promising strategy for preventing or treating lymph node metastasis.99
Intramuscular Injection
Intramuscular injection has been utilized as a delivery method for EVs. For instance, intramuscular administration of EVs derived from iMSCs significantly promoted microvessel formation in ischemic limbs of mice by upregulating the expression of angiogenesis-related molecules, thereby alleviating limb ischemia.71 In a follow-up study, researchers investigated the biodistribution of intramuscularly injected EVs derived from skeletal muscle 24 hours post-injection. The results indicated substantial accumulation in the gastrointestinal tract and liver, followed by the pancreas and lungs, with minimal presence in the kidneys, spleen, muscle, heart, and brain. Compared to intravenous administration at the same dose, intramuscular injection of EVs exhibited reduced distribution in the liver and spleen.148
Other in-situ Injection
Intra-Articular Injection
Intra-articular local injection represents a promising approach to enhance the accumulation of EVs at lesion sites in osteoarticular diseases. Zhang et al demonstrated that intra-articular administration of EVs derived from bone marrow mesenchymal stem cells (BMSCs) promoted macrophage polarization from the M1 to M2 phenotype, thereby mitigating synovial inflammation and cartilage damage, and slowing the progression of osteoarthritis.149 In a follow-up study, Jin et al corroborated these findings, showing that both BMSCs and their EVs, when administered intra-articularly, reduced cartilage destruction and subchondral bone remodeling in an osteoarthritis rat model. This was accompanied by a decrease in chondrocyte senescence and apoptosis, potentially elucidating their therapeutic mechanisms.17
Intrathecal Injection
Intrathecal injections, despite being highly invasive and associated with poor patient compliance, have been investigated for the delivery of EVs in neurological disorders due to their capacity to enhance drug concentration in the cerebrospinal fluid. For instance, Lu et al demonstrated that intrathecal administration of EVs derived from human placental MSCs alleviated neuropathic pain in a spared nerve injury mouse model via the miR-26a-5p/Wnt5a pathway.150
Transdermal Application
Transdermal administration of EVs has demonstrated considerable potential in dermatology. By bypassing first-pass metabolism, EVs can enhance bioavailability at the target site, thereby reducing dosing frequency and minimizing adverse effects. Zhang et al investigated the impact of topically applied Oil-in-Water Emulsions containing mesenchymal stem cell (MSC) exosomes on imiquimod (IMQ)-induced psoriasis in mice. Their study revealed significant inhibition of complement activation in the stratum corneum and a reduction in IL-17 release.151 However, EVs were predominantly confined to the stratum corneum, with limited penetration into the epidermis. Hydrogels, which are superior carriers for transdermal drug delivery, have been utilized in conjunction with EVs for topical applications. Shiekh et al developed synthetic hydrogel dressings loaded with adipose-derived stem cell EVs embedded in antioxidant polyurethane (PUAO), facilitating continuous oxygen release. This approach mitigated oxidative stress, promoted angiogenesis, enhanced collagen deposition, and accelerated re-epithelialization, leading to faster diabetic wound healing within two weeks.152
Intrathoracic Injection
A study investigated the intrapleural administration of EVs. In healthy mice, tumor-derived EVs injected intrapleurally were predominantly distributed to the lungs, liver, and spleen, suggesting the potential of this route for delivering EVs in lung-related conditions.66 In a mouse model of lung cancer-induced pleural effusion, intrapleurally administered EVs primarily accumulated in the lungs as well as in tumors, with prolonged retention in pleural cavity tumors compared to intravenous administration. A subsequent clinical trial demonstrated that intrapleural injection of methotrexate-loaded autologous tumor-derived EVs in patients with malignant pleural effusion effectively eliminated tumor cells and significantly reduced pleural effusion, with only grade 1 or 2 toxicity observed. This approach offers a promising new direction for symptom management in patients with advanced malignant pleural effusion.66
Intravitreal Injection
Pathological angiogenesis is a hallmark of various vision-threatening disorders. Tian et al developed regulatory T cell-derived EVs conjugated with anti-VEGF antibodies, which significantly prolonged the intraocular retention time of anti-VEGF antibodies following intravitreal injection. This approach effectively reduced choroidal neovascularization in a laser-induced choroidal neovascularization mouse model, presenting a promising therapeutic strategy for ocular neovascularization.153 However, Xue et al investigated the distribution of endothelial cell-derived EVs conjugated with the anti-angiogenic peptide KV11 (EXOKV11) via both intravitreal and retro-orbital injection routes. The FITC signal colocalized with retinal vessels 12 hours post-retro-orbital injection, whereas intravitreal injection did not show significant co-localization.154 To enhance intraocular EV retention, Bao et al engineered degradable polymeric microcapsules loaded with bone marrow mesenchymal stem cells (MSCs), mimicking the natural release of cellular EVs. Intravitreal injection in a retinal ischemia/reperfusion injury mouse model demonstrated that microcapsules released EVs over a period of one month, while free EVs nearly disappeared by day 7 and MSC signals nearly disappeared by day 14.155 Notably, intravitreal injection carries risks such as retinal detachment, retinal hemorrhage, and endophthalmitis,156 whereas retro-orbital injection is less invasive and poses fewer risks.157
Furthermore, localized EV delivery to the myocardium,158 kidneys,159 and prostate160 has been investigated due to its high retention in target tissues and reduced toxicity in non-target tissues. However, given that local delivery to deep tissues is more complex and invasive compared to systemic administration, researchers tend to favor high-dose systemic delivery to achieve the desired therapeutic outcomes.
Conclusions and Future Directions
In summary, EVs play crucial roles in both physiological and pathological processes. They serve as biomarkers for disease diagnosis and prognosis, as well as functional components in disease treatment. Owing to their natural origin, inherent stability, low toxicity, superior biocompatibility, and efficient biological membrane penetration—qualities that surpass those of liposomes and polymer particles—EVs have emerged as novel and exceptional nano-carriers for effective drug delivery. It is of paramount importance to elucidate the biodistribution of EVs within the body to enhance their therapeutic efficacy and minimize adverse effects. Genetic engineering and chemical modifications can improve organ targeting, thereby overcoming the rapid clearance of natural EVs and their high retention in tissues such as the liver.161 However, notable variations in biodistribution among EVs derived from different sources and administered via different routes underscore the necessity of optimizing delivery methods and selecting appropriate EV sources to improve delivery efficiency based on target tissues. As research into EVs continues to advance, investigators are increasingly focusing on more cost-effective and accessible sources, such as plant- and milk-derived EVs, which warrant further exploration of their physiological impacts. EV mimetics can mitigate large-scale production challenges to a significant extent. Moreover, synthetic vesicles derived from liposome fusion acquire enhanced controlled-release properties, thereby improving delivery efficiency.42 Additionally, the integration of EVs with advanced biomaterials for localized delivery represents an exciting frontier. This approach can prolong EV retention in target tissues, enhance therapeutic efficacy, and reduce the frequency of administration.
However, the in vivo biodistribution of EVs derived from various sources remains inadequately explored. A substantial body of research is urgently required to elucidate the distribution patterns of EVs from different origins and under varying administration methods. Moreover, multiple studies have confirmed that the distribution patterns of the same type of EVs differ between normal conditions and disease models. This inherent tropism can be partially attributed to EVs inheriting membrane components and biological activities from their parental cells. Presently, studies have adopted diverse combinations of EV sources, sizes, delivery methods, and disease models, leading to variations in the observed biodistribution of EVs. When researchers aim to enhance the therapeutic efficacy of EVs by optimizing their source and delivery method, they must consider these multiple factors to avoid overgeneralization. Given the growing body of research on the distribution patterns of EVs in the future, elucidating the distinct distribution profiles of EVs from the same source across various disease models and delivery routes holds significant guiding value for advancing the clinical application of EVs. Notably, due to the small size of EVs, precise in vivo tracking is complex, driving the development of various tracking methods.162 However, it has been demonstrated that genetically modifying EVs for imaging purposes can alter their natural organ distribution. For instance, the fusion of CD63 with NanoLuc luciferase has been shown to increase EV accumulation specifically in the lungs.163 Therefore, the careful selection of an appropriate EV tracing method is crucial for ensuring the scientific validity of EV biodistribution studies.
Furthermore, given that we are still in the exploratory phase of EVs, there remain significant gaps in our understanding of their mechanisms of biogenesis, uptake, and function, as well as the heterogeneity of EVs. Ensuring the homogeneity and reproducibility of therapeutic effects necessitates controlling the source of EVs and developing standardized methods for separation and characterization to enhance exosome yield and purity.164 According to the Minimal Information for Studies of Extracellular Vesicles 2023 (MISEV2023) guidelines, recognized separation techniques include differential (ultra)centrifugation, density gradients or cushions, size exclusion chromatography, fluid flow-based separation, and charge or molecular recognition-based separations. However, each of these methods has inherent limitations and may not be optimal.165 Furthermore, research has indicated that varying separation and purification methodologies can influence the biodistribution of EVs.56 Addressing these limitations or exploring novel approaches is of paramount importance. Characterization of EVs presents unique challenges due to their diverse sources, small size, and purity concerns; consequently, no single method can comprehensively characterize all EVs. Currently, numerous advanced methodologies at the single-vesicle level have been devised to address the challenge of heterogeneity, including digital single extracellular vesicle flow cytometry. However, such high-resolution analyses tend to be time-consuming.165 MISEV2023 recommends employing multiple complementary techniques concurrently.2 As research on EVs advances, standardized protocols for EV isolation and characterization are urgently required to facilitate clinical translation. Additionally, with the increasing selectivity of EV sources and the proliferation of clinical studies, ethical concerns regarding engineered EVs, such as those modified through genetic means, have arisen. Therefore, it is imperative to establish guidelines and implement oversight for these emerging issues.166
Despite the numerous challenges that the clinical application of EVs continues to encounter, EVs undoubtedly represent a highly promising frontier in nanomedicine, providing new hope across various domains of clinical practice.
Data Sharing Statement
No data were used for the research described in the article.
Funding
This study was supported by the National Key R&D Program of China (No. 2021YFA1101500), the National Natural Science Foundation of China (Grant Nos. 81974221, 81974009, and 92049119), the Hubei Innovation Group Foundation (No. 2022CFA019), and the General Program of Health Commission of Hubei Province (No. WJ2023M126).
Disclosure
The authors declare no conflicts of interest.
References
1. van Niel G, D’Angelo G, Raposo G. Shedding light on the cell biology of extracellular vesicles. Nat Rev mol Cell Biol. 2018;19(4):213–228. doi:10.1038/nrm.2017.125
2. Welsh JA, Goberdhan DCI, O’Driscoll L, et al. Minimal information for studies of extracellular vesicles (MISEV2023): from basic to advanced approaches. J Extracell Vesicles. 2024;13(2):e12404. doi:10.1002/jev2.12404
3. Chargaff E, West R. The biological significance of the thromboplastic protein of blood. J Biol Chem. 1946;166(1):189–197.
4. Johnstone RM, Adam M, Hammond JR, Orr L, Turbide C. Vesicle formation during reticulocyte maturation. Association of plasma membrane activities with released vesicles (exosomes). J Biol Chem. 1987;262(19):9412–9420.
5. Gurung S, Perocheau D, Touramanidou L, Baruteau J. The exosome journey: from biogenesis to uptake and intracellular signalling. Cell Commun Signal. 2021;19(1):47. doi:10.1186/s12964-021-00730-1
6. Kimiz-Gebologlu I, Oncel SS. Exosomes: large-scale production, isolation, drug loading efficiency, and biodistribution and uptake. J Control Release. 2022;347:533–543. doi:10.1016/j.jconrel.2022.05.027
7. Mathieu M, Martin-Jaular L, Lavieu G, Théry C. Specificities of secretion and uptake of exosomes and other extracellular vesicles for cell-to-cell communication. Nat Cell Biol. 2019;21(1):9–17. doi:10.1038/s41556-018-0250-9
8. Mondal J, Pillarisetti S, Junnuthula V, et al. Hybrid exosomes, exosome-like nanovesicles and engineered exosomes for therapeutic applications. J Control Release. 2023;353:1127–1149. doi:10.1016/j.jconrel.2022.12.027
9. Rehman FU, Liu Y, Zheng M, Shi B. Exosomes based strategies for brain drug delivery. Biomaterials. 2023;293:121949. doi:10.1016/j.biomaterials.2022.121949
10. Wiklander OPB, Nordin JZ, O’Loughlin A, et al. Extracellular vesicle in vivo biodistribution is determined by cell source, route of administration and targeting. J Extracell Vesicles. 2015;4(1):26316. doi:10.3402/jev.v4.26316
11. Ma Y, Dong S, Grippin AJ, et al. Engineering therapeutical extracellular vesicles for clinical translation. Trends Biotechnol. 2025;43(1):61–82. doi:10.1016/j.tibtech.2024.08.007
12. Vos J, Tejeda-Mora H, Merino A, et al. Bio-distribution and longevity of mesenchymal stromal cell derived membrane particles. J Control Release. 2022;350:642–651. doi:10.1016/j.jconrel.2022.08.060
13. Betzer O, Perets N, Angel A, et al. In vivo neuroimaging of exosomes using gold nanoparticles. ACS Nano. 2017;11(11):10883–10893. doi:10.1021/acsnano.7b04495
14. Eliopoulos N, Stagg J, Lejeune L, Pommey S, Galipeau J. Allogeneic marrow stromal cells are immune rejected by MHC class I- and class II-mismatched recipient mice. Blood. 2005;106(13):4057–4065. doi:10.1182/blood-2005-03-1004
15. Gleeson BM, Martin K, Ali MT, et al. Bone marrow-derived mesenchymal stem cells have innate procoagulant activity and cause microvascular obstruction following intracoronary delivery: amelioration by antithrombin therapy. Stem Cells. 2015;33(9):2726–2737. doi:10.1002/stem.2050
16. Cheng L, Zhang K, Wu S, Cui M, Xu T. Focus on mesenchymal stem cell-derived exosomes: opportunities and challenges in cell-free therapy. Stem Cells Int. 2017;2017:6305295. doi:10.1155/2017/6305295
17. Jin Y, Xu M, Zhu H, et al. Therapeutic effects of bone marrow mesenchymal stem cells-derived exosomes on osteoarthritis. J Cell Mol Med. 2021;25(19):9281–9294. doi:10.1111/jcmm.16860
18. Ortega A, Martinez-Arroyo O, Forner MJ, Cortes R. Exosomes as drug delivery systems: endogenous nanovehicles for treatment of systemic lupus erythematosus. Pharmaceutics. 2020;13(1). doi:10.3390/pharmaceutics13010003
19. Yi YW, Lee JH, Kim SY, et al. Advances in Analysis of Biodistribution of Exosomes by Molecular Imaging. Int J mol Sci. 2020;21(2). doi:10.3390/ijms21020665
20. Zamanian MH, Norooznezhad AH, Hosseinkhani Z, et al. Human placental mesenchymal stromal cell-derived small extracellular vesicles as a treatment for severe COVID-19: a double-blind randomized controlled clinical trial. J Extracell Vesicles. 2024;13(7):e12492. doi:10.1002/jev2.12492
21. Nassar W, El-Ansary M, Sabry D, et al. Umbilical cord mesenchymal stem cells derived extracellular vesicles can safely ameliorate the progression of chronic kidney diseases. Biomater Res. 2016;20:21. doi:10.1186/s40824-016-0068-0
22. Johnson J, Law SQK, Shojaee M, et al. First-in-human clinical trial of allogeneic, platelet-derived extracellular vesicles as a potential therapeutic for delayed wound healing. J Extracell Vesicles. 2023;12(7):e12332. doi:10.1002/jev2.12332
23. Kordelas L, Rebmann V, Ludwig AK, et al. MSC-derived exosomes: a novel tool to treat therapy-refractory graft-versus-host disease. Leukemia. 2014;28(4):970–973. doi:10.1038/leu.2014.41
24. Norooznezhad AH, Yarani R, Payandeh M, et al. Treatment of persistent chemotherapy-induced hair loss (Alopecia) with human mesenchymal stromal cells exosome enriched extracellular vesicles: a case report. Heliyon. 2023;9(4):e15165. doi:10.1016/j.heliyon.2023.e15165
25. Xie X, Song Q, Dai C, et al. Clinical safety and efficacy of allogenic human adipose mesenchymal stromal cells-derived exosomes in patients with mild to moderate Alzheimer’s disease: a Phase I/II clinical trial. Gen Psychiatr. 2023;36(5):e101143. doi:10.1136/gpsych-2023-101143
26. Madhan S, Dhar R, Devi A. Plant-derived exosomes: a green approach for cancer drug delivery. J Mater Chem B. 2024;12(9):2236–2252. doi:10.1039/d3tb02752j
27. Orefice NS, Di Raimo R, Mizzoni D, Logozzi M, Fais S. Purposing plant-derived exosomes-like nanovesicles for drug delivery: patents and literature review. Expert Opin Ther Pat. 2023;33(2):89–100. doi:10.1080/13543776.2023.2195093
28. Wang Q, Zhuang X, Mu J, et al. Delivery of therapeutic agents by nanoparticles made of grapefruit-derived lipids. Nat Commun. 2013;4:1867. doi:10.1038/ncomms2886
29. Elsharkasy OM, Nordin JZ, Hagey DW, et al. Extracellular vesicles as drug delivery systems: why and how? Adv Drug Deliv Rev. 2020;159:332–343. doi:10.1016/j.addr.2020.04.004
30. Song M, Cui M, Fang Z, Liu K. Advanced research on extracellular vesicles based oral drug delivery systems. J Control Release. 2022;351:560–572. doi:10.1016/j.jconrel.2022.09.043
31. Sercombe L, Veerati T, Moheimani F, Wu SY, Sood AK, Hua S. Advances and Challenges of Liposome Assisted Drug Delivery. Front Pharmacol. 2015;6:286. doi:10.3389/fphar.2015.00286
32. Sharma S, Parveen R, Chatterji BP. Toxicology of Nanoparticles in Drug Delivery. Curr Pathobiol Rep. 2021;9(4):133–144. doi:10.1007/s40139-021-00227-z
33. Dad HA, Gu TW, Zhu AQ, Huang LQ, Peng LH. Plant exosome-like nanovesicles: emerging therapeutics and drug delivery nanoplatforms. Mol Ther. 2021;29(1):13–31. doi:10.1016/j.ymthe.2020.11.030
34. Haney MJ, Klyachko NL, Zhao Y, et al. Exosomes as drug delivery vehicles for Parkinson’s disease therapy. J Control Release. 2015;207:18–30. doi:10.1016/j.jconrel.2015.03.033
35. Xiong B, Wang Y, Chen Y, et al. Strategies for structural modification of small molecules to improve blood-brain barrier penetration: a recent perspective. J Med Chem. 2021;64(18):13152–13173. doi:10.1021/acs.jmedchem.1c00910
36. Wang B, Zhuang X, Deng ZB, et al. Targeted drug delivery to intestinal macrophages by bioactive nanovesicles released from grapefruit. Mol Ther. 2014;22(3):522–534. doi:10.1038/mt.2013.190
37. Zhang Y, Bi J, Huang J, Tang Y, Du S, Li P. Exosome: a review of its classification, isolation techniques, storage, diagnostic and targeted therapy applications. Int J Nanomed. 2020;15:6917–6934. doi:10.2147/ijn.S264498
38. Chatterjee M, Özdemir S, Fritz C, et al. Plasma extracellular vesicle tau and TDP-43 as diagnostic biomarkers in FTD and ALS. Nat Med. 2024;30(6):1771–1783. doi:10.1038/s41591-024-02937-4
39. Han Z, Peng X, Yang Y, et al. Integrated microfluidic-SERS for exosome biomarker profiling and osteosarcoma diagnosis. Biosens Bioelectron. 2022;217:114709. doi:10.1016/j.bios.2022.114709
40. Jiang S, Lu F, Chen J, et al. UPCARE: urinary extracellular vesicles-derived prostate cancer assessment for risk evaluation. J Extracell Vesicles. 2024;13(8):e12491. doi:10.1002/jev2.12491
41. Li K, Lin Y, Luo Y, et al. A signature of saliva-derived exosomal small RNAs as predicting biomarker for esophageal carcinoma: a multicenter prospective study. mol Cancer. 2022;21(1):21. doi:10.1186/s12943-022-01499-8
42. Zou Z, Li H, Xu G, Hu Y, Zhang W, Tian K. Current knowledge and future perspectives of exosomes as nanocarriers in diagnosis and treatment of diseases. Int J Nanomed. 2023;18:4751–4778. doi:10.2147/ijn.S417422
43. Xu Z, Zeng S, Gong Z, Yan Y. Exosome-based immunotherapy: a promising approach for cancer treatment. mol Cancer. 2020;19(1):160. doi:10.1186/s12943-020-01278-3
44. Liu H, Chen L, Liu J, et al. Co-delivery of tumor-derived exosomes with alpha-galactosylceramide on dendritic cell-based immunotherapy for glioblastoma. Cancer Lett. 2017;411:182–190. doi:10.1016/j.canlet.2017.09.022
45. Yu L, Zeng X, Hu X, Wen Q, Chen P. Advances and challenges in clinical applications of tumor cell-derived extracellular vesicles. Colloids Surf B Biointerfaces. 2024;234:113704. doi:10.1016/j.colsurfb.2023.113704
46. Wang H, Mu J, Chen Y, et al. Hybrid Ginseng-derived extracellular vesicles-like particles with autologous tumor cell membrane for personalized vaccination to inhibit tumor recurrence and metastasis. Adv Sci. 2024;11(17):e2308235. doi:10.1002/advs.202308235
47. Salgueiro VC, Passemar C, Vázquez-Iniesta L, Lerma L, Floto A, Prados-Rosales R. Extracellular vesicles in mycobacteria: new findings in biogenesis, host-pathogen interactions, and diagnostics. mBio. 2024;15(5):e0255223. doi:10.1128/mbio.02552-23
48. Prados-Rosales R, Carreño LJ, Batista-Gonzalez A, et al. Mycobacterial membrane vesicles administered systemically in mice induce a protective immune response to surface compartments of Mycobacterium tuberculosis. mBio. 2014;5(5):e01921–14. doi:10.1128/mBio.01921-14
49. Li C, Xue H, Du X, Nyaruaba R, Yang H, Wei H. Outer membrane vesicles generated by an exogenous bacteriophage lysin and protection against Acinetobacter baumannii infection. J Nanobiotechnology. 2024;22(1):273. doi:10.1186/s12951-024-02553-x
50. Zhang Y, Lin Y, He J, et al. Milk-derived small extracellular vesicles: a new perspective on dairy nutrition. Crit Rev Food Sci Nutr. 2023:1–22. doi:10.1080/10408398.2023.2263573
51. Kang M, Jordan V, Blenkiron C, Chamley LW. Biodistribution of extracellular vesicles following administration into animals: a systematic review. J Extracell Vesicles. 2021;10(8):e12085. doi:10.1002/jev2.12085
52. Parada N, Romero-Trujillo A, Georges N, Alcayaga-Miranda F. Camouflage strategies for therapeutic exosomes evasion from phagocytosis. J Adv Res. 2021;31:61–74. doi:10.1016/j.jare.2021.01.001
53. Zhang H, Freitas D, Kim HS, et al. Identification of distinct nanoparticles and subsets of extracellular vesicles by asymmetric flow field-flow fractionation. Nat Cell Biol. 2018;20(3):332–343. doi:10.1038/s41556-018-0040-4
54. Xiao Q, Zoulikha M, Qiu M, et al. The effects of protein Corona on in vivo fate of nanocarriers. Adv Drug Deliv Rev. 2022;186:114356. doi:10.1016/j.addr.2022.114356
55. Liam-Or R, Faruqu FN, Walters A, et al. Cellular uptake and in vivo distribution of mesenchymal-stem-cell-derived extracellular vesicles are protein Corona dependent. Nat Nanotechnol. 2024;19(6):846–855. doi:10.1038/s41565-023-01585-y
56. Nordin JZ, Lee Y, Vader P, et al. Ultrafiltration with size-exclusion liquid chromatography for high yield isolation of extracellular vesicles preserving intact biophysical and functional properties. Nanomedicine. 2015;11(4):879–883. doi:10.1016/j.nano.2015.01.003
57. Rosenkrans ZT, Thickens AS, Kink JA, et al. Investigating the in vivo biodistribution of extracellular vesicles isolated from various human cell sources using positron emission tomography. Mol Pharm. 2024;21(9):4324–4335. doi:10.1021/acs.molpharmaceut.4c00298
58. Wang Z, Hu S, Popowski KD, et al. Inhalation of ACE2-expressing lung exosomes provides prophylactic protection against SARS-CoV-2. Nat Commun. 2024;15(1):2236. doi:10.1038/s41467-024-45628-x
59. Qiao L, Hu S, Huang K, et al. Tumor cell-derived exosomes home to their cells of origin and can be used as Trojan horses to deliver cancer drugs. Theranostics. 2020;10(8):3474–3487. doi:10.7150/thno.39434
60. Hoshino A, Costa-Silva B, Shen TL, et al. Tumour exosome integrins determine organotropic metastasis. Nature. 2015;527:7578):329–35. doi:10.1038/nature15756
61. Antimisiaris SG, Mourtas S, Marazioti A. Exosomes and exosome-inspired vesicles for targeted drug delivery. Pharmaceutics. 2018;10(4). doi:10.3390/pharmaceutics10040218
62. Ma T, Chen S, Wang J, et al. Enhanced osteolysis targeted therapy through fusion of exosomes derived from M2 macrophages and bone marrow mesenchymal stem cells: modulating macrophage polarization. Small. 2024;20(7):e2303506. doi:10.1002/smll.202303506
63. Luan X, Sansanaphongpricha K, Myers I, Chen H, Yuan H, Sun D. Engineering exosomes as refined biological nanoplatforms for drug delivery. Acta Pharmacol Sin. 2017;38(6):754–763. doi:10.1038/aps.2017.12
64. Wang J, Tang W, Yang M, et al. Inflammatory tumor microenvironment responsive neutrophil exosomes-based drug delivery system for targeted glioma therapy. Biomaterials. 2021;273:120784. doi:10.1016/j.biomaterials.2021.120784
65. Sadeghi S, Tehrani FR, Tahmasebi S, Shafiee A, Hashemi SM. Exosome engineering in cell therapy and drug delivery. Inflammopharmacology. 2023;31(1):145–169. doi:10.1007/s10787-022-01115-7
66. Guo M, Wu F, Hu G, et al. Autologous tumor cell-derived microparticle-based targeted chemotherapy in lung cancer patients with malignant pleural effusion. Sci Transl Med. 2019;11(474). doi:10.1126/scitranslmed.aat5690
67. Wagner W. Implications of long-term culture for mesenchymal stem cells: genetic defects or epigenetic regulation? Stem Cell Res Ther. 2012;3(6):54. doi:10.1186/scrt145
68. Wagner W, Ho AD, Zenke M. Different facets of aging in human mesenchymal stem cells. Tissue Eng Part B Rev. 2010;16(4):445–453. doi:10.1089/ten.TEB.2009.0825
69. Fafián-Labora J, Morente-López M, Sánchez-Dopico MJ, et al. Influence of mesenchymal stem cell-derived extracellular vesicles in vitro and their role in ageing. Stem Cell Res Ther. 2020;11(1):13. doi:10.1186/s13287-019-1534-0
70. Cai Y, Wang S, Qu J, Belmonte JCI, Liu GH. Rejuvenation of Tissue Stem Cells by Intrinsic and Extrinsic Factors. Stem Cells Transl Med. 2022;11(3):231–238. doi:10.1093/stcltm/szab012
71. Hu GW, Li Q, Niu X, et al. Exosomes secreted by human-induced pluripotent stem cell-derived mesenchymal stem cells attenuate limb ischemia by promoting angiogenesis in mice. Stem Cell Res Ther. 2015;6(1):10. doi:10.1186/scrt546
72. Zhou X, Deng X, Liu M, et al. Intranasal delivery of BDNF-loaded small extracellular vesicles for cerebral ischemia therapy. J Control Release. 2023;357:1–19. doi:10.1016/j.jconrel.2023.03.033
73. Yuan X, Sun L, Jeske R, et al. Engineering extracellular vesicles by three-dimensional dynamic culture of human mesenchymal stem cells. J Extracell Vesicles. 2022;11(6):e12235. doi:10.1002/jev2.12235
74. Zhong J, Xia B, Shan S, et al. High-quality milk exosomes as oral drug delivery system. Biomaterials. 2021;277:121126. doi:10.1016/j.biomaterials.2021.121126
75. Agrawal AK, Aqil F, Jeyabalan J, et al. Milk-derived exosomes for oral delivery of paclitaxel. Nanomedicine. 2017;13(5):1627–1636. doi:10.1016/j.nano.2017.03.001
76. Samuel M, Fonseka P, Sanwlani R, et al. Oral administration of bovine milk-derived extracellular vesicles induces senescence in the primary tumor but accelerates cancer metastasis. Nat Commun. 2021;12(1):3950. doi:10.1038/s41467-021-24273-8
77. Zhang C, Zhang H, Millán Cotto HA, et al. Milk exosomes anchored with hydrophilic and zwitterionic motifs enhance mucus permeability for applications in oral gene delivery. Biomater Sci. 2024;12(3):634–649. doi:10.1039/d3bm01089a
78. Hu Y, Hou Z, Liu Z, et al. Oyster mantle-derived exosomes alleviate osteoporosis by regulating bone homeostasis. Biomaterials. 2024;311:122648. doi:10.1016/j.biomaterials.2024.122648
79. Halperin W, Jensen WA. Ultrastructural changes during growth and embryogenesis in carrot cell cultures. J Ultrastruct Res. 1967;18(3):428–443. doi:10.1016/s0022-5320(67)80128-x
80. Zhang M, Viennois E, Prasad M, et al. Edible ginger-derived nanoparticles: a novel therapeutic approach for the prevention and treatment of inflammatory bowel disease and colitis-associated cancer. Biomaterials. 2016;101:321–340. doi:10.1016/j.biomaterials.2016.06.018
81. Liu C, Yan X, Zhang Y, et al. Oral administration of turmeric-derived exosome-like nanovesicles with anti-inflammatory and pro-resolving bioactions for murine colitis therapy. J Nanobiotechnology. 2022;20(1):206. doi:10.1186/s12951-022-01421-w
82. Şahin F, Koçak P, Güneş MY, Özkan İ, Yıldırım E, Kala EY. In vitro wound healing activity of wheat-derived nanovesicles. Appl Biochem Biotechnol. 2019;188(2):381–394. doi:10.1007/s12010-018-2913-1
83. Schuh C, Cuenca J, Alcayaga-Miranda F, Khoury M. Exosomes on the border of species and kingdom intercommunication. Transl Res. 2019;210:80–98. doi:10.1016/j.trsl.2019.03.008
84. Liu C, Yazdani N, Moran CS, et al. Unveiling clinical applications of bacterial extracellular vesicles as natural nanomaterials in disease diagnosis and therapeutics. Acta Biomater. 2024;180:18–45. doi:10.1016/j.actbio.2024.04.022
85. Jiang L, Driedonks TAP, Jong WSP, et al. A bacterial extracellular vesicle-based intranasal vaccine against SARS-CoV-2 protects against disease and elicits neutralizing antibodies to wild-type and Delta variants. J Extracell Vesicles. 2022;11(3):e12192. doi:10.1002/jev2.12192
86. Feng Q, Ma X, Cheng K, et al. Engineered bacterial outer membrane vesicles as controllable two-way adaptors to activate macrophage phagocytosis for improved tumor immunotherapy. Adv Mater. 2022;34(40):e2206200. doi:10.1002/adma.202206200
87. Qing S, Lyu C, Zhu L, et al. Biomineralized bacterial outer membrane vesicles potentiate safe and efficient tumor microenvironment reprogramming for anticancer therapy. Adv Mater. 2020;32(47):e2002085. doi:10.1002/adma.202002085
88. Bai X, Li C, Qiu J, et al. A “plug-and-display” nanoparticle based on attenuated outer membrane vesicles enhances the immunogenicity of protein antigens. J Control Release. 2024;378:687–700. doi:10.1016/j.jconrel.2024.12.022
89. Fan J, Lee CS, Kim S, Chen C, Aghaloo T, Lee M. Generation of small RNA-modulated exosome mimetics for bone regeneration. ACS Nano. 2020;14(9):11973–11984. doi:10.1021/acsnano.0c05122
90. Lin M, Lei S, Chai Y, et al. Immunosuppressive microvesicles-mimetic derived from tolerant dendritic cells to target T-lymphocytes for inflammation diseases therapy. J Nanobiotechnology. 2024;22(1):201. doi:10.1186/s12951-024-02470-z
91. Vader P, Mol EA, Pasterkamp G, Schiffelers RM. Extracellular vesicles for drug delivery. Adv Drug Deliv Rev. 2016;106(Pt A):148–156. doi:10.1016/j.addr.2016.02.006
92. Liu A, Yang G, Liu Y, Liu T. Research progress in membrane fusion-based hybrid exosomes for drug delivery systems. Front Bioeng Biotechnol. 2022;10:939441. doi:10.3389/fbioe.2022.939441
93. Goh WJ, Zou S, Lee CK, et al. EXOPLEXs: Chimeric drug delivery platform from the fusion of cell-derived nanovesicles and liposomes. Biomacromolecules. 2018;19(1):22–30. doi:10.1021/acs.biomac.7b01176
94. Li L, He D, Guo Q, et al. Exosome-liposome hybrid nanoparticle codelivery of TP and miR497 conspicuously overcomes chemoresistant ovarian cancer. J Nanobiotechnology. 2022;20(1):50. doi:10.1186/s12951-022-01264-5
95. Yang B, Chen Y, Shi J. Exosome biochemistry and advanced nanotechnology for next-generation theranostic platforms. Adv Mater. 2019;31(2):e1802896. doi:10.1002/adma.201802896
96. Li J, Wu H, Yu Z, et al. Hematopoietic stem and progenitor cell membrane-coated vesicles for bone marrow-targeted leukaemia drug delivery. Nat Commun. 2024;15(1):5689. doi:10.1038/s41467-024-50021-9
97. Manca S, Upadhyaya B, Mutai E, et al. Milk exosomes are bioavailable and distinct microRNA cargos have unique tissue distribution patterns. Sci Rep. 2018;8(1):11321. doi:10.1038/s41598-018-29780-1
98. Shi MM, Yang QY, Monsel A, et al. Preclinical efficacy and clinical safety of clinical-grade nebulized allogenic adipose mesenchymal stromal cells-derived extracellular vesicles. J Extracell Vesicles. 2021;10(10):e12134. doi:10.1002/jev2.12134
99. Sakurai Y, Ohtani A, Nakayama Y, et al. Logistics and distribution of small extracellular vesicles from the subcutaneous space to the lymphatic system. J Control Release. 2023;361:77–86. doi:10.1016/j.jconrel.2023.07.043
100. Sun Y, Shi H, Yin S, et al. Human mesenchymal stem cell derived exosomes alleviate Type 2 diabetes mellitus by reversing peripheral insulin resistance and relieving β-cell destruction. ACS Nano. 2018;12(8):7613–7628. doi:10.1021/acsnano.7b07643
101. Milano G, Biemmi V, Lazzarini E, et al. Intravenous administration of cardiac progenitor cell-derived exosomes protects against doxorubicin/trastuzumab-induced cardiac toxicity. Cardiovasc Res. 2020;116(2):383–392. doi:10.1093/cvr/cvz108
102. Tian T, Zhang HX, He CP, et al. Surface functionalized exosomes as targeted drug delivery vehicles for cerebral ischemia therapy. Biomaterials. 2018;150:137–149. doi:10.1016/j.biomaterials.2017.10.012
103. Kučuk N, Primožič M, Knez Ž, Leitgeb M. Exosomes engineering and their roles as therapy delivery tools, therapeutic targets, and biomarkers. Int J mol Sci. 2021;22(17). doi:10.3390/ijms22179543
104. Munagala R, Aqil F, Jeyabalan J, Gupta RC. Bovine milk-derived exosomes for drug delivery. Cancer Lett. 2016;371(1):48–61. doi:10.1016/j.canlet.2015.10.020
105. Zhou X, Li Z, Sun W, Yang G, Xing C, Yuan L. Delivery efficacy differences of intravenous and intraperitoneal injection of exosomes: perspectives from tracking dye labeled and MiRNA encapsulated exosomes. Curr Drug Deliv. 2020;17(3):186–194. doi:10.2174/1567201817666200122163251
106. Heidari N, Abbasi-Kenarsari H, Namaki S, et al. Adipose-derived mesenchymal stem cell-secreted exosome alleviates dextran sulfate sodium-induced acute colitis by Treg cell induction and inflammatory cytokine reduction. J Cell Physiol. 2021;236(8):5906–5920. doi:10.1002/jcp.30275
107. Homayun B, Lin X, Choi HJ. Challenges and recent progress in oral drug delivery systems for biopharmaceuticals. Pharmaceutics. 2019;11(3). doi:10.3390/pharmaceutics11030129
108. Ensign LM, Cone R, Hanes J. Oral drug delivery with polymeric nanoparticles: the gastrointestinal mucus barriers. Adv Drug Deliv Rev. 2012;64(6):557–570. doi:10.1016/j.addr.2011.12.009
109. Vashisht M, Rani P, Onteru SK, Singh D. Curcumin Encapsulated in Milk Exosomes Resists Human Digestion and Possesses Enhanced Intestinal Permeability in Vitro. Appl Biochem Biotechnol. 2017;183(3):993–1007. doi:10.1007/s12010-017-2478-4
110. Wang D, Zhang H, Liao X, et al. Oral administration of Robinia pseudoacacia L. flower exosome-like nanoparticles attenuates gastric and small intestinal mucosal ferroptosis caused by hypoxia through inhibiting HIF-1α- and HIF-2α-mediated lipid peroxidation. J Nanobiotechnology. 2024;22(1):479. doi:10.1186/s12951-024-02663-6
111. Jing R, Zhang L, Li R, et al. Milk-derived extracellular vesicles functionalized with anti-tumour necrosis factor-α nanobody and anti-microbial peptide alleviate ulcerative colitis in mice. J Extracell Vesicles. 2024;13(6):e12462. doi:10.1002/jev2.12462
112. Zhu MZ, Xu HM, Liang YJ, et al. Edible exosome-like nanoparticles from portulaca oleracea L mitigate DSS-induced colitis via facilitating double-positive CD4(+)CD8(+)T cells expansion. J Nanobiotechnology. 2023;21(1):309. doi:10.1186/s12951-023-02065-0
113. Chen Q, Zu M, Gong H, et al. Tea leaf-derived exosome-like nanotherapeutics retard breast tumor growth by pro-apoptosis and microbiota modulation. J Nanobiotechnology. 2023;21(1):6. doi:10.1186/s12951-022-01755-5
114. Tong L, Hao H, Zhang Z, et al. Milk-derived extracellular vesicles alleviate ulcerative colitis by regulating the gut immunity and reshaping the gut microbiota. Theranostics. 2021;11(17):8570–8586. doi:10.7150/thno.62046
115. Huang W, Zhang Q, Li W, et al. Development of novel nanoantibiotics using an outer membrane vesicle-based drug efflux mechanism. J Control Release. 2020;317:1–22. doi:10.1016/j.jconrel.2019.11.017
116. Aqil F, Munagala R, Jeyabalan J, Agrawal AK, Gupta R. Exosomes for the Enhanced Tissue Bioavailability and Efficacy of Curcumin. Aaps j. 2017;19(6):1691–1702. doi:10.1208/s12248-017-0154-9
117. Wu L, Wang L, Liu X, et al. Milk-derived exosomes exhibit versatile effects for improved oral drug delivery. Acta Pharm Sin B. 2022;12(4):2029–2042. doi:10.1016/j.apsb.2021.12.015
118. Deng Z, Rong Y, Teng Y, et al. Broccoli-derived nanoparticle inhibits mouse colitis by activating dendritic cell AMP-activated protein kinase. Mol Ther. 2017;25(7):1641–1654. doi:10.1016/j.ymthe.2017.01.025
119. Sundaram K, Mu J, Kumar A, et al. Garlic exosome-like nanoparticles reverse high-fat diet induced obesity via the gut/brain axis. Theranostics. 2022;12(3):1220–1246. doi:10.7150/thno.65427
120. Chen Q, Li Q, Liang Y, et al. Natural exosome-like nanovesicles from edible tea flowers suppress metastatic breast cancer via ROS generation and microbiota modulation. Acta Pharm Sin B. 2022;12(2):907–923. doi:10.1016/j.apsb.2021.08.016
121. Carobolante G, Mantaj J, Ferrari E, Vllasaliu D. Cow milk and intestinal epithelial cell-derived extracellular vesicles as systems for enhancing oral drug delivery. Pharmaceutics. 2020;12(3). doi:10.3390/pharmaceutics12030226
122. Lochhead JJ, Davis TP. Perivascular and perineural pathways involved in brain delivery and distribution of drugs after intranasal administration. Pharmaceutics. 2019;11(11). doi:10.3390/pharmaceutics11110598
123. Zhuang X, Xiang X, Grizzle W, et al. Treatment of brain inflammatory diseases by delivering exosome encapsulated anti-inflammatory drugs from the nasal region to the brain. Mol Ther. 2011;19(10):1769–1779. doi:10.1038/mt.2011.164
124. Tai J, Han M, Lee D, Park IH, Lee SH, Kim TH. Different methods and formulations of drugs and vaccines for nasal administration. Pharmaceutics. 2022;14(5). doi:10.3390/pharmaceutics14051073
125. Zhuang X, Teng Y, Samykutty A, et al. Grapefruit-derived nanovectors delivering therapeutic miR17 through an intranasal route inhibit brain tumor progression. Mol Ther. 2016;24(1):96–105. doi:10.1038/mt.2015.188
126. Peng H, Li Y, Ji W, et al. Intranasal administration of self-oriented nanocarriers based on therapeutic exosomes for synergistic treatment of Parkinson’s disease. ACS Nano. 2022;16(1):869–884. doi:10.1021/acsnano.1c08473
127. Losurdo M, Pedrazzoli M, D’Agostino C, et al. Intranasal delivery of mesenchymal stem cell-derived extracellular vesicles exerts immunomodulatory and neuroprotective effects in a 3xTg model of Alzheimer’s disease. Stem Cells Transl Med. 2020;9(9):1068–1084. doi:10.1002/sctm.19-0327
128. Guo S, Perets N, Betzer O, et al. Intranasal delivery of mesenchymal stem cell derived exosomes loaded with phosphatase and tensin homolog siRNA repairs complete spinal cord injury. ACS Nano. 2019;13(9):10015–10028. doi:10.1021/acsnano.9b01892
129. Zhou J, Li F, Jia B, et al. Intranasal delivery of small extracellular vesicles reduces the progress of amyotrophic lateral sclerosis and the overactivation of complement-coagulation cascade and NF-ĸB signaling in SOD1(G93A) mice. J Nanobiotechnology. 2024;22(1):503. doi:10.1186/s12951-024-02764-2
130. Xie Y, Sun Y, Liu Y, et al. Targeted delivery of RGD-CD146(+)CD271(+) human umbilical cord mesenchymal stem cell-derived exosomes promotes blood-spinal cord barrier repair after spinal cord injury. ACS Nano. 2023;17(18):18008–18024. doi:10.1021/acsnano.3c04423
131. Ma R, Kutchy NA, Wang Z, Hu G. Extracellular vesicle-mediated delivery of anti-miR-106b inhibits morphine-induced primary ciliogenesis in the brain. Mol Ther. 2023;31(5):1332–1345. doi:10.1016/j.ymthe.2023.03.030
132. Wang K, Kumar US, Sadeghipour N, Massoud TF, Paulmurugan R. A microfluidics-based scalable approach to generate extracellular vesicles with enhanced therapeutic MicroRNA loading for intranasal delivery to mouse glioblastomas. ACS Nano. 2021;15(11):18327–18346. doi:10.1021/acsnano.1c07587
133. Kim MS, Haney MJ, Zhao Y, et al. Development of exosome-encapsulated paclitaxel to overcome MDR in cancer cells. Nanomedicine. 2016;12(3):655–664. doi:10.1016/j.nano.2015.10.012
134. Wang J, Tan Y, Dai Y, et al. Intranasal delivery of endothelial cell-derived extracellular vesicles with Supramolecular gel attenuates myocardial ischemia-reperfusion injury. Int J Nanomed. 2023;18:5495–5510. doi:10.2147/ijn.S420301
135. Sukumar UK, Bose RJC, Malhotra M, et al. Intranasal delivery of targeted polyfunctional gold-iron oxide nanoparticles loaded with therapeutic microRNAs for combined theranostic multimodality imaging and presensitization of glioblastoma to temozolomide. Biomaterials. 2019;218:119342. doi:10.1016/j.biomaterials.2019.119342
136. Royo F, Cossío U, Ruiz de Angulo A, Llop J, Falcon-Perez JM. Modification of the glycosylation of extracellular vesicles alters their biodistribution in mice. Nanoscale. 2019;11(4):1531–1537. doi:10.1039/c8nr03900c
137. Ruiz-de-angulo A, Zabaleta A, Gómez-Vallejo V, Llop J, Mareque-Rivas JC. Microdosed lipid-coated (67)Ga-magnetite enhances antigen-specific immunity by image tracked delivery of antigen and CpG to lymph nodes. ACS Nano. 2016;10(1):1602–1618. doi:10.1021/acsnano.5b07253
138. Hu Y, Rao SS, Wang ZX, et al. Exosomes from human umbilical cord blood accelerate cutaneous wound healing through miR-21-3p-mediated promotion of angiogenesis and fibroblast function. Theranostics. 2018;8(1):169–184. doi:10.7150/thno.21234
139. Hu L, Wang J, Zhou X, et al. Exosomes derived from human adipose mensenchymal stem cells accelerates cutaneous wound healing via optimizing the characteristics of fibroblasts. Sci Rep. 2016;6:32993. doi:10.1038/srep32993
140. Fathi-Karkan S, Heidarzadeh M, Narmi MT, et al. Exosome-loaded microneedle patches: promising factor delivery route. Int J Biol Macromol. 2023;243:125232. doi:10.1016/j.ijbiomac.2023.125232
141. Yang G, Chen Q, Wen D, et al. A therapeutic microneedle patch made from hair-derived keratin for promoting hair regrowth. ACS Nano. 2019;13(4):4354–4360. doi:10.1021/acsnano.8b09573
142. Peswani Sajnani SL, Zhang Y, Vllasaliu D. Exosome-based therapies for mucosal delivery. Int J Pharm. 2021;608:121087. doi:10.1016/j.ijpharm.2021.121087
143. AboulFotouh K, Almanza G, Yu YS, et al. Inhalable dry powders of microRNA-laden extracellular vesicles prepared by thin-film freeze-drying. Int J Pharm. 2024;651:123757. doi:10.1016/j.ijpharm.2023.123757
144. Zhang Q, Pan J, Xiong D, et al. Pulmonary aerosol delivery of Let-7b microRNA confers a striking inhibitory effect on lung carcinogenesis through targeting the tumor immune microenvironment. Adv Sci. 2021;8(17):e2100629. doi:10.1002/advs.202100629
145. Dinh PC, Paudel D, Brochu H, et al. Inhalation of lung spheroid cell secretome and exosomes promotes lung repair in pulmonary fibrosis. Nat Commun. 2020;11(1):1064. doi:10.1038/s41467-020-14344-7
146. Gao Y, Zhang H, Zhou N, et al. Methotrexate-loaded tumour-cell-derived microvesicles can relieve biliary obstruction in patients with extrahepatic cholangiocarcinoma. Nat Biomed Eng. 2020;4(7):743–753. doi:10.1038/s41551-020-0583-0
147. Rezaie J, Ajezi S, Ç B A, et al. Exosomes and their application in biomedical field: difficulties and advantages. mol Neurobiol. 2018;55(4):3372–3393. doi:10.1007/s12035-017-0582-7
148. Jalabert A, Vial G, Guay C, et al. Exosome-like vesicles released from lipid-induced insulin-resistant muscles modulate gene expression and proliferation of beta recipient cells in mice. Diabetologia. 2016;59(5):1049–1058. doi:10.1007/s00125-016-3882-y
149. Zhang J, Rong Y, Luo C, Cui W. Bone marrow mesenchymal stem cell-derived exosomes prevent osteoarthritis by regulating synovial macrophage polarization. Aging. 2020;12(24):25138–25152. doi:10.18632/aging.104110
150. Lu Y, Zhang J, Zeng F, et al. Human PMSCs-derived small extracellular vesicles alleviate neuropathic pain through miR-26a-5p/Wnt5a in SNI mice model. J Neuroinflammation. 2022;19(1):221. doi:10.1186/s12974-022-02578-9
151. Zhang B, Lai RC, Sim WK, Choo ABH, Lane EB, Lim SK. Topical application of mesenchymal stem cell exosomes alleviates the imiquimod induced psoriasis-like inflammation. Int J mol Sci. 2021;22(2). doi:10.3390/ijms22020720
152. Shiekh PA, Singh A, Kumar A. Exosome laden oxygen releasing antioxidant and antibacterial cryogel wound dressing OxOBand alleviate diabetic and infectious wound healing. Biomaterials. 2020;249:120020. doi:10.1016/j.biomaterials.2020.120020
153. Tian Y, Zhang F, Qiu Y, et al. Reduction of choroidal neovascularization via cleavable VEGF antibodies conjugated to exosomes derived from regulatory T cells. Nat Biomed Eng. 2021;5(9):968–982. doi:10.1038/s41551-021-00764-3
154. Dong X, Lei Y, Yu Z, et al. Exosome-mediated delivery of an anti-angiogenic peptide inhibits pathological retinal angiogenesis. Theranostics. 2021;11(11):5107–5126. doi:10.7150/thno.54755
155. Bao H, Tian Y, Wang H, et al. Exosome-loaded degradable polymeric microcapsules for the treatment of vitreoretinal diseases. Nat Biomed Eng. 2023. doi:10.1038/s41551-023-01112-3
156. van Wijngaarden P, Coster DJ, Williams KA. Inhibitors of ocular neovascularization: promises and potential problems. JAMA. 2005;293(12):1509–1513. doi:10.1001/jama.293.12.1509
157. Nayak K, Misra M. A review on recent drug delivery systems for posterior segment of eye. Biomed Pharmacother. 2018;107:1564–1582. doi:10.1016/j.biopha.2018.08.138
158. Potz BA, Scrimgeour LA, Pavlov VI, Sodha NR, Abid MR, Sellke FW. Extracellular vesicle injection improves myocardial function and increases angiogenesis in a swine model of chronic ischemia. J Am Heart Assoc. 2018;7(12). doi:10.1161/jaha.117.008344
159. Song T, Eirin A, Zhu X, et al. Mesenchymal stem cell-derived extracellular vesicles induce regulatory T Cells to ameliorate chronic kidney injury. Hypertension. 2020;75(5):1223–1232. doi:10.1161/hypertensionaha.119.14546
160. Goudarzi F, Kiani A, Moradi M, et al. Intraprostatic injection of exosomes isolated from adipose-derived mesenchymal stem cells for the treatment of chronic non-bacterial prostatitis. J Tissue Eng Regen Med. 2021;15(12):1144–1154. doi:10.1002/term.3251
161. Ming-Kun C, Zi-Xian C, Mao-Ping C, Hong C, Zhuang-Fei C, Shan-Chao Z. Engineered extracellular vesicles: a new approach for targeted therapy of tumors and overcoming drug resistance. Cancer Commun. 2024;44(2):205–225. doi:10.1002/cac2.12518
162. Aimaletdinov AM, Gomzikova MO. Tracking of extracellular vesicles’ biodistribution: new methods and approaches. Int J mol Sci. 2022;23(19). doi:10.3390/ijms231911312
163. Lázaro-Ibáñez E, Faruqu FN, Saleh AF, et al. Selection of fluorescent, bioluminescent, and radioactive tracers to accurately reflect extracellular vesicle biodistribution in vivo. ACS Nano. 2021;15(2):3212–3227. doi:10.1021/acsnano.0c09873
164. Li G, Chen T, Dahlman J, et al. Current challenges and future directions for engineering extracellular vesicles for heart, lung, blood and sleep diseases. J Extracell Vesicles. 2023;12(2):e12305. doi:10.1002/jev2.12305
165. Manno M, Bongiovanni A, Margolis L, Bergese P, Arosio P. The physico-chemical landscape of extracellular vesicles. Nat Rev Bioeng. 2025;3(1):68–82. doi:10.1038/s44222-024-00255-5
166. Tian J, Han Z, Song D, et al. Engineered exosome for drug delivery: recent development and clinical applications. Int J Nanomed. 2023;18:7923–7940. doi:10.2147/ijn.S444582
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms.php
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms.php
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.