")
Back to Journals » International Journal of Nanomedicine » Volume 20
From Plant Based Therapy to Plant-Derived Vesicle-Like Nanoparticles for Cancer Treatment: Past, Present and Future
Authors An Y, Sun JX, Ma SY, Xu MY, Xu JZ, Liu CQ, Wang SG , Xia QD
Received 9 October 2024
Accepted for publication 21 February 2025
Published 17 March 2025 Volume 2025:20 Pages 3471—3491
DOI https://doi.org/10.2147/IJN.S499893
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Professor Lijie Grace Zhang
Ye An,* Jian-Xuan Sun,* Si-Yang Ma,* Meng-Yao Xu, Jin-Zhou Xu, Chen-Qian Liu, Shao-Gang Wang, Qi-Dong Xia
Department and Institute of Urology, Tongji Hospital, Tongji Medical College, Huazhong University of Science and Technology, Wuhan, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Shao-Gang Wang; Qi-Dong Xia, Department and Institute of Urology, Tongji Hospital, Tongji Medical College, Huazhong University of Science and Technology, Wuhan, People’s Republic of China, Tel +86 027 83663460, Email [email protected]; [email protected]
Abstract: Cancer stands as a formidable malady profoundly impacting human health. Throughout history, plant-based therapies have remained pivotal in the arsenal against cancer, evolving alongside the epochs. Presently, challenges such as the arduous extraction of active components and potential safety concerns impede the progression of plant-based anticancer therapies. The isolation of plant-derived vesicle-like nanoparticles (PDVLNs), a kind of lipid bilayer capsules isolated from plants, has brought plant-based anticancer therapy into a novel realm and has led to decades of research on PDVLNs. Accumulating evidence indicates that PDVLNs can deliver plant-derived active substances to human cells and regulate cellular functions. Regulating immunity, inducing cell cycle arrest, and promoting apoptosis in cancer cells are the most commonly reported mechanisms of PDVLNs in tumor suppression. Low immunogenicity and lack of tumorigenicity make PDVLNs a good platform for drug delivery. The molecules within the PDVLNs are all from source plants, so the selection of source plants is crucial. In recent years, there has been a clear trend that the source plants have changed from vegetables or fruits to medicinal plants. This review highlights the mechanisms of medicinal plant-based cancer therapies to identify candidate source plants. More importantly, the current research on PDVLN-based cancer therapy and the applications of PDVLNs for drug delivery are systematically discussed.
Keywords: plant-derived vesicle-like nanoparticles, plant-based anticancer therapy, drug delivery system, anticancer
Graphical Abstract:

Introduction
Extracellular vesicles (EVs) are heterogeneous nano-sized lipid bilayer capsules that are naturally secreted from cells (Figure 1A). Classically, EVs can be divided into three major subgroups according to their biogenesis, biological function, size, and content namely, microvesicles (MVs), exosomes, and apoptotic bodies. In the last decades, mammal-derived EVs have been the focus of EV research, and detailed research has been conducted on their biogenesis and function. Through transporting bioactive molecules among cells, EVs adjust metabolic balance and play an important role in intercellular communications.1 In addition to widely involved in the regulation of physiological processes, EVs have been found to participate in the development and progression of many diseases.2 In recent years, EVs have emerged as a new type of therapeutic agent and have been widely applied in the treatment of numerous diseases, not just for understanding and intervening in their own biological functions. For example, mesenchymal stem cells (MSCs) derived exosomes have shown great potential in regenerative medicine because they can repair myocardial infarction and wound healing in mouse models.3,4 EVs are also widely used in cancer therapy.
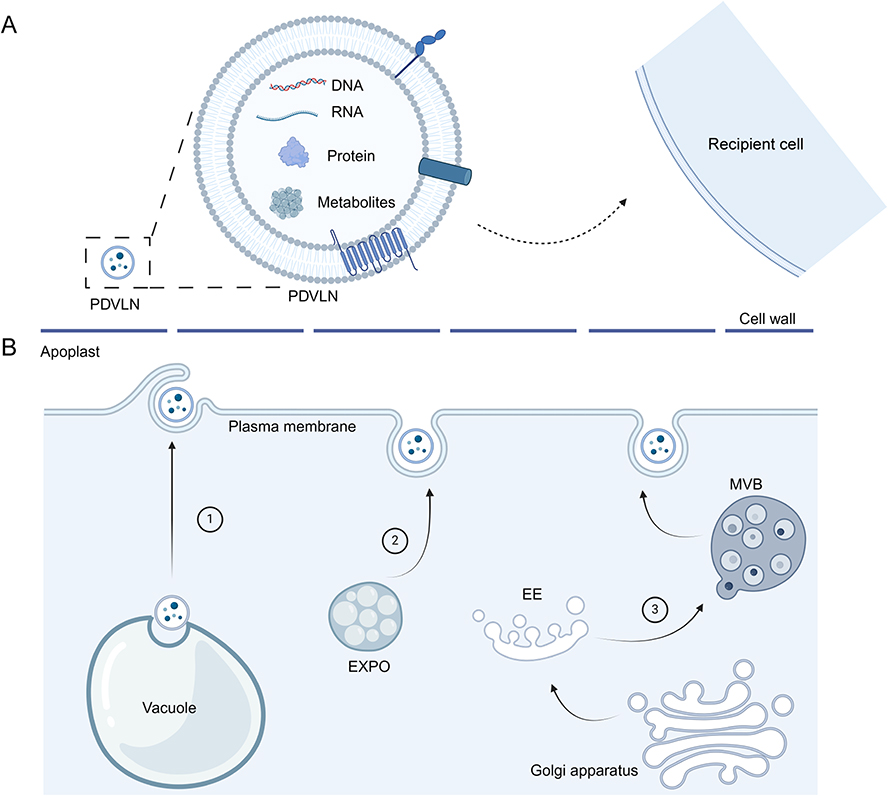 |
Figure 1 (A) PDVLNs delivery active components such as nucleic acids, metabolites and proteins to recipient cells. (B) Biogenesis pathways of PDVLNs, namely multivesicular bodies (MVB) pathway, exocyse positive organelles (EXPO) and vacuole pathways. |
Cancer is a complex disease and the leading cause of death worldwide every year. Although chemotherapy achieves good clinical cancer control, it cannot perfectly target tumor cells and may damage normal cells. Therefore, various new treatments for cancer are currently under development, and EV-based therapies have attracted considerable attention. In addition to the early direct application of EVs in cancer treatment, researchers have preferred to use EVs as a treatment tool.5 One of the most direct applications of EVs is their use as drug delivery platforms. After loading drugs into EVs, EVs can carry drugs through biological barriers, such as the blood-brain barrier, and reach tumor tissues.6 Interestingly, EVs exhibit some ability to target and accumulate in tumor tissues, which is further enhanced by adding a targeting ligand on the surface of EVs.7 For example, targeting EVs could be produced by genetically engineering their parental cells. Antibodies or targeting peptide can be linked to membrane proteins on the surface of the EVs (such as Lamp 2b).8,9 The intrinsic targeting ability of EVs is due to the inherent properties of the EVs, such as different protein components and lipid ratios on the EVS surface and different parent cells. For example, EVs expressing the β5 subunit are predominantly directed towards Kupffer cells in the liver, while those expressing the β4 subunit are preferentially taken up by CD31-positive endothelial cells in the brain.10 And EVs could acquire better targeting ability, longer retention time, and better treatment efficacy after engineering, all of which make them a promising agent for tumor treatment. However, problems such as potential tumorigenicity and immunogenicity have encouraged researchers to develop new carriers,11,12 ultimately turning their attention to plant-derived vesicle-like nanoparticles (PDVLNs).
In 2009, Regente et al first isolated PDVLNs from sunflowers, which opened a new era for extracellular vesicles and led to decades of research on PDVLNs.13 Plant-derived exosome-like nanoparticles, plant-derived EVs, and plant-derived exosome-like nanovesicles are commonly used terms describing PDVLNs in some studies, and in this review, we use PDVLNs to refer to them. PDVLNs are morphologically and biogenically similar to mammal-derived extracellular vesicles.14 PDVLNs contain various RNA, DNA, proteins, and metabolites, which perform different functions by transporting them to the recipient cell along with PDVLNs (Figure 1A). Different from mammalian EVs that are mainly secreted through membrane budding and the multivesicular body pathway, current research indicates that there are at least three ways in which plants secrete PDVLNs (Figure 1B). To date, numerous studies have demonstrated that PDVLNs can serve as drug carriers and also exert therapeutic effects on their own. B. javanica-derived PDVLNs are capable of treating breast cancer, which is ascribed to the microRNAs (miRNAs) within them.15 The contents of PDVLNs are basically all derived from the plants. For example, peu-MIR2916-p3 derived from garlic PDVLNs have been found to promote the growth of Bacteroides thetaiotaomicron in the intestines, thereby alleviating colitis.16 Therefore, the choice of the source plant is crucial.
There is an apparent trend that the choice of the source plant changes from fruits and vegetables to medicinal plants. The history of using plants as medicines can be traced back to at least 60,000 years during the Middle Paleolithic age.17 Various civilizations, such as Ancient China, Ancient India, and Ancient Greece, as well as traditional medical systems have utilized plants to treat various diseases, including cancer.17,18 Traditional Chinese Medicine (TCM) is an important system for plant-based anticancer therapy, with more than 3500 years of medical practice. Various Chinese herbal medicines are important components of TCM.19 Currently, TCM is widely used in the treatment of various tumors, such as lung cancer,19,20 triple-negative breast cancer,21 and esophageal cancer,22 and can be used in combination with other anticancer therapies, including chemotherapy, radiotherapy, tyrosine kinase inhibitors (TKIs), and immunotherapy, or as adjunctive or maintenance therapy.20,23 These TCM have also become important candidates for the sources of PDVLNs.
In this review, we summarize the mechanisms of plant-based anticancer therapy to help identify medicinal plants with the potential to serve as new sources of PDVLNs. Next, we comprehensively describe current research on PDVLNs in cancer treatment and summarize their mechanisms. In addition, we discuss the applications of PDVLNs as carriers and their modification methods.
The Mechanisms of Plant-Based Anticancer Therapies
Antioxidant Activity and Anti-Inflammatory Activity
Many plants contain abundant antioxidant compounds such as polyphenols, vitamin C, and vitamin E. Polyphenol compounds are characterized by multiple phenolic (aromatic) rings and are abundant in plants. They can be further classified into subclasses such as flavonoids, tannins, lignans, phenolic acids, and stilbenes.24 Each of them has a unique chemical structure, but almost all have antioxidant and anticancer properties. They can neutralize free radicals, reduce oxidant stress, and prevent DNA damage.25,26 Chronic inflammation is an important factor in cancer development. They can recruit inflammatory cells, induce cell proliferation, and produce reactive oxygen species (ROS), which can result in DNA damage and abnormal DNA repair, eventually leading to permanent genomic alterations and tumorigenesis.27 Compounds with antioxidant activity can also fight inflammation by reducing oxidative damage caused by ROS in the inflammatory environment. Figure 2 exhibits the mechanisms of action of some plants for cancer treatment.
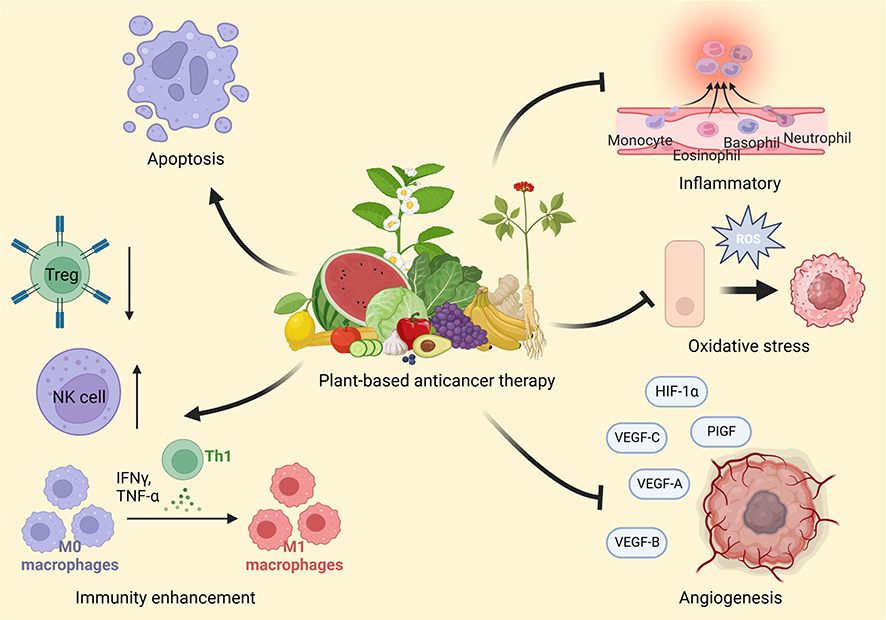 |
Figure 2 Various mechanisms of plant-based anticancer therapy. |
Regulation of Cell Cycle
Certain compounds in plants can regulate the cell cycle and interfere with DNA replication and repair in cancer cells, thus preventing the proliferation and division of cancer cells and halting their growth. One of the most famous examples is taxanes, including paclitaxel (taxol) and docetaxel, which are extracted and purified from the bark of the Pacific yew tree (Taxus brevifolia) and have been applied in the treatment of various cancers, such as breast,28 prostate29 and many other cancers. Microtubules (MTs), which consist of α-tubulin and β-tubulin heterodimers, are the main components of the cytoskeleton in eukaryotic cells and play crucial roles in cell division, cell motility, cell shape maintenance and intracellular transport. Taxanes can selectively bind to β-tubulin and promote their polymerization and assembly, thus preventing their disassembly, stabilizing the microtubules, and interfering with the formation of spindles, thereby inhibiting cell division and arresting the cell cycle at the G2/M phase.30 Other compounds, such as vinblastine and vincristine, can also exert anticancer effects by regulating the cell cycle.31,32 In addition, different plant-derived compounds can precisely target different stages of the cell cycle, thereby achieving precise regulation of the cell cycle. For example, algal sulfated polysaccharide extracted from Laurencia papillosa could target G1 phase, and xanthohumol, the major component of Humulus lupulus could target S phase.33
Induction of Apoptosis
Apoptosis is a type of programmed cell death (PCD) that can be triggered by multiple internal (release of cytochrome c from mitochondria) and external (binding of death ligands to their corresponding death receptors) signals. A series of signaling events occur once apoptosis is initiated, and all these signaling pathways finally converge on the activation of specific proteases called caspases (caspase-3, 8, and 9, etc)., which cleave essential cellular proteins and initiate the dismantling of the cell.34 Induction of apoptosis in cancer cells (eg, prostate cancer, breast cancer and colorectal cancer etc). has always been an interesting topic in the field of cancer treatment. Several plant-derived compounds have been shown to target vital molecules in the apoptotic pathways, such as Bcl-2 and caspases. Resveratrol can be extracted from more than 70 plants including grapes, peanuts, and pistachios. It has antioxidant and anti-inflammatory properties, which make it a promising candidate for cancer treatment. In particular, it can suppress the phosphorylation of Src-STAT3 and induce apoptosis in cancer cells to directly kill tumor cells.35 In addition to small-molecule anticancer agents, proteinaceous anticancer agents derived from plants, such as lectins, can also induce apoptosis to exert anticancer effects. Lectins are a class of glycoproteins which exist ubiquitously in different plant species. They can interact with sugar-binding receptors on the plasma membrane, are internalized through endocytosis, and are finally transported to the mitochondria. ROS are then generated, and cytochrome c is released into the cytoplasm to activate downstream apoptotic pathways.36 Moreover, some plant-derived compounds, such as terpenoids and flavonoids, can induce other types of PCD, such as autophagy-dependent cell death, ferroptosis and necroptosis.37,38 For example, betulin, a triterpenoid extracted from Isatidis Radix, could induces autophagy-dependent cell death in colorectal cancer cells by inhibiting PI3K/Akt/mTOR signaling pathways.39 Lou et al found Ginkgetin derived from Ginkgo biloba leaves could increased labile iron pool and lipid peroxidation, thus inducing ferroptosis in EGFR wild-type non-small-cell lung cancer.40
Inhibition of Angiogenesis
Pathological angiogenesis is a hallmark of cancer, and pro-angiogenic members of the vascular endothelial growth factor (VEGF) family (VEGF-A, VEGF-B, VEGF-C, etc). and their corresponding receptors (VEGFR-1, VEGFR-2, and VEGFR-3) play a vital role in this process. Catechins in green tea have been proven to possess antioxidant and anti-inflammatory activities, as well as their ability to inhibit angiogenesis. Epigallocatechin-3-gallate (EGCG) is the main catechin found in green tea. Studies have found that EGCG can reduce the expression of HIF-1 α, which strongly activates the expression of VEGF, and EGCG could also inhibit the binding of VEGF to its corresponding receptor, thus preventing the activation of the VEGF/VEGFR axis.41 In addition, plant-based anticancer therapies can also improve the anti-angiogenesis efficacy of other treatments. For example, Ao et al found that the extract from coix seed in combination with thalidomide significantly inhibited angiogenesis in liver cancer by inhibiting VEGF and B-FGF. Lin et al also reported that HIF-1α and VEGF, which are vital for angiogenesis, could be remarkably reduced by astragalus polysaccharide combined with cisplatin and adriamycin in live cancers.42
Enhancement of Immunity
Some plant-derived agents can not only kill tumor cells directly but can also play an anticancer role by enhancing the body’s immunity. A good example is Traditional Chinese Medicine.43–45 TCM can enhance antitumor immunity by influencing both the innate and adaptive immune systems.45 Macrophages are important components of innate immunity and can primarily be classified into M1 and M2 subtypes. M1 macrophages mainly exert anti-tumor effects, and they can secrete a large amount of pro-inflammatory cytokines, such as tumor necrosis factor-α (TNF-α), interleukin-1β (IL-1β), and interleukin-6 (IL-6), which promote inflammatory responses. Additionally, they release reactive oxygen species (ROS) and reactive nitrogen species (RNS), which can effectively kill tumor cells. Many TCMs have been reported to promote the transformation of M2 type macrophages to M1 type to enhance antitumor immunity, such as Astragaloside IV extracted from astragalus46 and soyasapogenols extracted from soybean.47 Kaneko et al also found that Xiao-chai-hu-tang augmented the activity of natural killer (NK) cells in a dose-dependent manner in mice.48 Lupeol promotes the proliferation of NK cells and increases their killing effect on gastric cancer cells.49 As for adaptive immunity, compound Chinese formula 1 (CFF-1) is a classic TCM-derived formula used in prostate cancer and has been proven to be therapeutic. Previous studies have found that CFF-1 induces apoptosis in prostate cancer by inhibiting the EGFR/PI3K/AKT signaling pathway. Interestingly, Zhang et al found that CFF-1 could also reduce PD-L1 expression in prostate cancer cells, increase the population of CD4+ T lymphocyte subsets, and simultaneously downregulate the population of regulatory T cells (Treg cells) at the same time, thereby dismantling the immunosuppressive environment in prostate cancer and enhancing anticancer immunity.50 Some TCMs can also exert antitumor effects by promoting T cell differentiation into Th1 cells, which can release several cytokines and chemokines to recruit CD8+ T cells and enhance their antitumor abilities.45
Challenges of Using Plant-Based Anticancer Therapy
Plant-based anticancer therapy has many advantages compared to other anticancer therapies; for example, they are mainly administered orally or by injection; therefore, they are more convenient and less painful for patients and have proven effective in many cancers with relatively few side effects.51 However, the challenges and drawbacks of plant-based anticancer therapies need to be addressed. It must be recognized that many compounds with anticancer activity in plants cannot be synthesized artificially, so the consumption is mainly dependent on natural resources.51 Take paclitaxel for example, paclitaxel is mainly found in the bark of the Pacific yew tree with low concentration of only 0.01%-0.05%, and the amount of paclitaxel extracted from 10 tons of bark is sufficient to treat 500 patients.30 Besides, the Pacific yew tree is a slow-growing tree, and stripping the bark usually leads to the death of the tree; therefore, over-harvesting once brought it to the brink of extinction.52 Although scientists have suggested the production of paclitaxel using plant cell cultures or genetically modified plants to reduce the consumption of natural sources, the purification efficiency is as low as 0.004%52 and many bioactive compounds are lost during the purification process. In addition, some plant-derived compounds can exhibit drug synergism and may be less effective after purification and administration as single molecules.51 The specific mechanisms underlying the anticancer effects of some plants remain unknown, so it is difficult to extract and purify their bioactive compounds. Moreover, there still exists some concerns about the bioavailability of plant-derived compounds. The absorption of many plant-derived compounds has not been well investigated; therefore, some of them may not be soluble properly in the systemic fluid and cannot be well absorbed and stably transferred to the tumor area to achieve a therapeutic concentration.51 Many methodologies have been adopted to improve bioavailability and absorption, such as encapsulation of drugs in liposomes or micelles, which can be modified to increase the targeting ability of drug delivery. However, synthetic materials also raise concerns regarding their safety; therefore, PDVLNs have unique advantages in this regard. They are less toxic or immunogenic because of their natural sources. In addition, many bioactive compounds can be retained in PDVLNs during the isolation process to exert synergistic anticancer effects, thereby significantly reducing losses during the purification process. Therefore, PDVLNs are playing increasingly crucial roles in plant based anticancer therapies.53
Characterization of the PDVLNs
As early as the 1960s, scientists first observed the release of exosome-like vesicles by the fusion of multivesicular bodies and plasma membranes from cultured carrot cells.54 In 2009, Regente et al successfully isolated exosome-like vesicles from the apoplast washing fluids of sunflower seeds,13 which led to decades of research on PDVLNs. To date, PDVLNs have been isolated from various plant species and shown to exhibit therapeutic effects in many human diseases.55,56 Similar to mammalian EVs, PDVLNs are a group of heterogeneous nano-sized lipid bilayer capsules that carry lipids, proteins, nucleic acids, and some metabolites.18
Size and Potential
Two commonly used methods for measuring the size of PDVLNs are dynamic light scattering (DLS) and nanoparticle tracking analysis (NTA). DLS serves as a non-single-particle characterization method, yet it suffers from the drawback that larger particulates can overshadow the signals emanating from smaller particles.57 Therefore, when dealing with samples that have a broad particle size distribution, the particle sizes measured by DLS may deviate from the actual values. NTA records the Brownian motion trajectories of individual particles, and quantitatively calculates particle size and concentration. However, it requires sufficient trace length and an optimal concentration range.58 The size distribution of PDVLNs varies among source plants, and the reported size ranges from 50 nm to 1000 nm.59 Notably, PDVLNs derived from the same plant source may exhibit different size distribution peaks. For example, orange PDVLNs exhibit a primary peak and a secondary smaller peak in their size distribution.60 The zeta potential of PDVLNs is mostly negative, and the zeta potential varies significantly among different PDVLNs. The reported zeta potential distribution ranges from −1.5 to −49.61
Protein Markers
Proteomics methods have been widely used to identify proteins in various PDVLNs, but our understanding of the functional roles these proteins play within PDVLNs remains limited. In contrast to the well-established marker proteins such as CD9, TSG101, and CD63 in mammalian EVs,62 there is currently a lack of consensus on specific marker proteins within PDVLNs. Proteins associated with membrane stability or PDVLN secretion may emerge as candidate marker proteins. Interestingly, in the latest research, Ma et al found that TSG101, CD63, and CD9 are highly expressed in nanoparticles derived from ginseng.63 A family of proteins that has been widely detected in PDVLNs is the heat shock proteins (HSPs). For example, HSP70 has been found to be highly expressed in nanoparticles derived from grapes and olive pollen grains.64,65 HSP90 has been reported to be present in PDVLNs from tea plants.66 PEN1 has been found to be highly expressed in PDVLNs derived from Arabidopsis and is associated with membrane surface components.67 However, to be recognized as a widely accepted vesicle surface marker protein, it still needs to be confirmed in nanoparticles from many different plant sources.
Morphology
Researchers have verified the successful isolation of PDVLNs by observing their bilayer lipid membrane structure through transmission electron microscopy. PDVLNs have been reported to have oval, spherical, or cup-shaped morphology.68 These morphological differences may originate from variations in plant species or electron microscopy imaging techniques. For instance, Mu et al used the same method to extract vesicles from grapes and grapefruits, but morphological differences could be observed.59 The transmission electron microscope (TEM), which is commonly used to observe the morphology of EVs, requires repeated dehydration during sample preparation, which may cause the EVs to take on a cup-like shape.69 Atomic force microscopy (AFM), a type of high-resolution microscope, has also been widely used for PDVLNs imaging.70
Isolation and Purification
Sample Preparation
PDVLNs are secreted into the apoplast space; therefore, before the formal isolation of PDVLNs, the juice containing PDVLNs was collected first. Figure 3 shows the general process of the PDVLNs isolation. There are two main methods to acquire PDVLNs juice: tissue disruption and tissue infiltration. For tissue disrupt method, physical methods (including grinding, blending, and squeezing) are used to disrupt plant tissues and obtain plant juice.71–73 For some plants with low juice content, phosphate buffer saline (PBS) can be added to plants and ground with plants to load PDVLNs.66,74 Following this, several low-speed centrifugation steps (usually two or three) are conducted to remove the fibers and cell debris. It is worth noting that during the duration of disruption, the extracellular matrix may contaminate the extracted juice, and subjected methods should be used to eliminate the influence of the matrix. A typical example is citrus, in which traditional differential centrifugation is difficult to separate from pectin. Tris-HCl can be added to citrus plant-derived juices to remove pectin.75 Moreover, the tissue-disrupt method inevitably destroys the cellular structure, and some organelles and cell fragments may be purified together with PDVLNs.76 As for the tissue-infiltration method, simply put, this method involves infiltrating the isolation buffer to apoplast to load the PDVLNs, and then extracting it out by centrifugation. Rutter et al successfully extracted PDVLNs from Arabidopsis using this method.67 The infiltration was driven by the pressure difference (using vacuum equipment), and centrifugation was usually performed at a low speed (700–5000 g).
 |
Figure 3 General process of the isolation of PDVLNs. The whole process can be divided into two parts: sample preparation and PDVLNs isolation. Tissue-disrupt method acquire plant juice rich in PDVLNs by grinding or squeezing, and tissue-infiltration method infiltrate buffer to load PDVLNs by pressure or vacuum. |
The tissue infiltration method causes less damage than tissue disruption and can thus obtain purer PDVLNs.53 However, the yield of PDVLNs extracted by the tissue infiltration method was lower than that extracted by the tissue disruption method. This is because the isolation buffer may dilute fluid from the apoplast. Therefore, it is important to select the appropriate method for this purpose.
Isolation and Purification of PDVLNs
As the interest of researchers in mammalian EVs has increased in the last decade, isolation methods for EVs have developed greatly, and numerous methods have been explored to isolate EVs. Similar to mammal-derived EVs, PDVLNs have been isolated based on their density, size, charge, and immunoaffinity. The most commonly reported method is ultracentrifugation (UC).61 After pre-processing the plant tissue, the juice is centrifuged one to three times at low speed (300–10,000 g) to remove impurities such as fibers and debris. Then, an ultracentrifugation step (> 40,000 g) is taken to finally collect the PDVLNs.77 The selection of centrifugation speed and time largely depends on the plant source and experience of the researchers. The advantage of UC is that researchers can simultaneously centrifuge hundreds of milliliters of juice to obtain good yields. Some organelle fragments and non-vesicular substances may have a density similar to that of PDVLNs, and may mix with PDVLNs in the last UC steps. In this context, some recent studies have combined UC and density gradient centrifugation (DGC) to obtain PDVLNs with higher purity.78 Two widely used gradient buffer are sucrose and iodixanol.79,80 However, considering the losses during the process and the potential interaction between the PDVLNs and the gradient medium, the combination of UC and DGC inevitably reduces the yield of PDVLNs.61 Notably, owing to the diversity of plant sources, specific isolation processes should be carefully evaluated and adjusted according to specific plant species. We look forward to more collaborations between researchers and botanists in the future to further improve the extraction process. The ultrafiltration method uses a membrane to separate PDVLNs from other contaminants based on their size. Lee et al isolated EVs from plant leaves using a 100kDa centrifugal filter.81 Although ultrafiltration does not require specialized equipment such as ultracentrifugation, it is usually applied in combination with other isolation methods owing to problems such as low purity and possible membrane blockage. Size exclusion chromatography (SEC) relies on the relationship between the hydrodynamic radius of the particles and pore size of the resin beads. By utilizing SEC, the PDVLNs can be separated based on their size. Kim et al combined ultrafiltration and SEC to isolate carrot derived PDVLNs and achieved relatively higher yields.82 Polymer-based precipitation uses polymers such as polyethylene glycol (PEG) to change the solubility of PDVLNs and increase their sedimentation. For example, Kalarikkal used PEG6000 to purify ginger PDVLNs and achieved a yield of 60% to 90% compared to UC.83 This method has a simple protocol and relatively high yield, but may lead to the aggregation of EVs and mixing with other proteins.84 Other methods, such as immunoaffinity, field-flow fractionation, and electrophoresis-coupled dialysis, have also been developed for PDVLN isolation; their principles and characteristics are shown in Table 1.
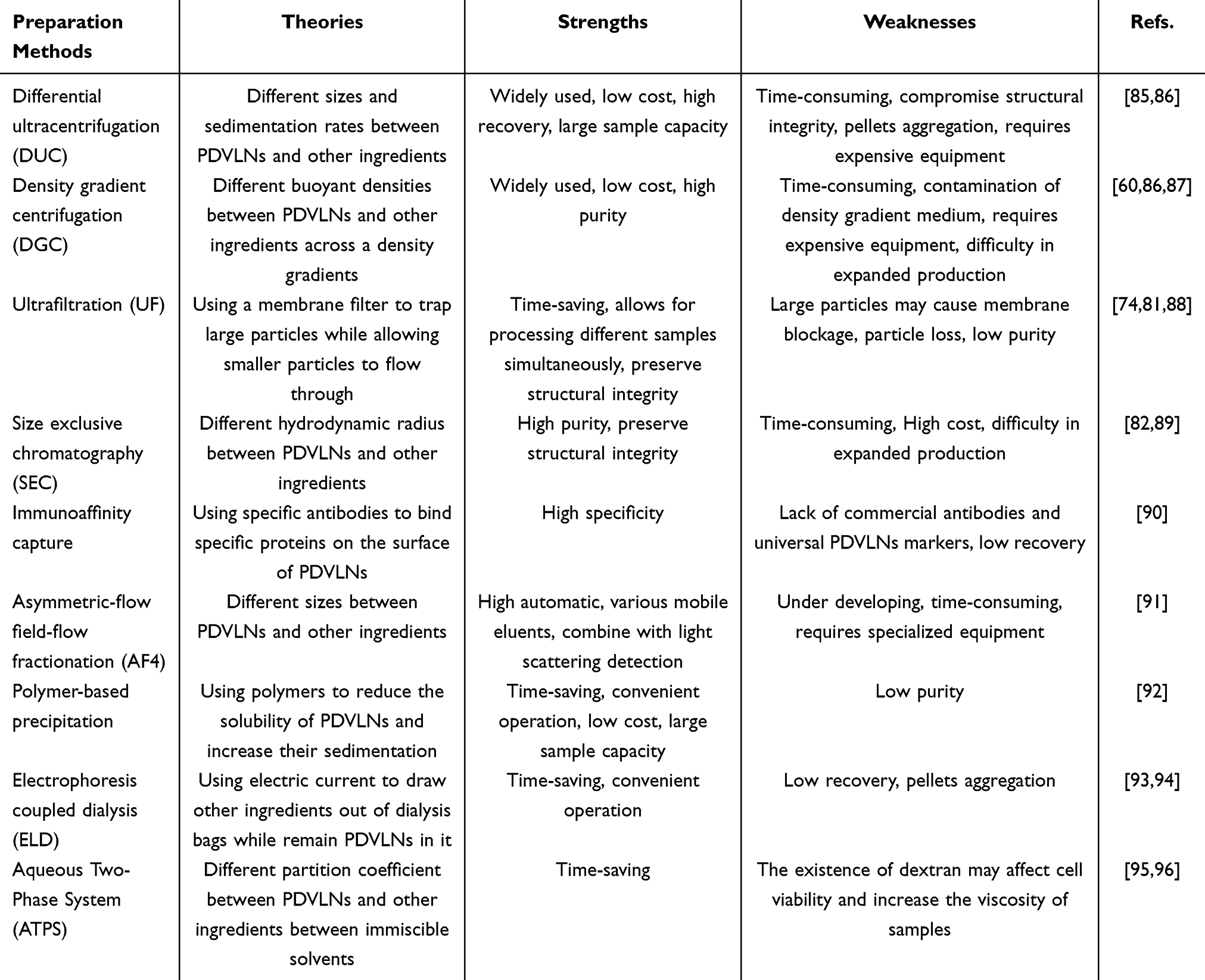 |
Table 1 Preparation Methods of PDVLNs |
Anticancer Effects of PDVLNs
In mammals, EVs play a vital role in intercellular communication and intercellular substance transport to regulate biological and pathological processes. Similarly, PDVLNs transport various bioactive substances among cells in plants. It has been proven that bioactive molecules encapsulated in PDVLNs play a role in interspecies communication,59 indicating the potential of PDVLNs for the treatment of human diseases. As mentioned earlier, there are many anticancer substances in plants, including proteins, miRNAs, and metabolites. Therefore, PDVLNs carrying anticancer substances have emerged as promising anticancer agents. Table 2 lists the current use of PDVLNs for cancer treatment, and Figure 4 shows the mechanisms underlying their anticancer activities.
 |
Figure 4 Various mechanisms of the anticancer effect of PDVLNs. There are five main aspects: induction of apoptosis, induction of cell cycle arrest, regulation of immune system, regulation of gut microbiota and synergistical enhancement with chemotherapy. |
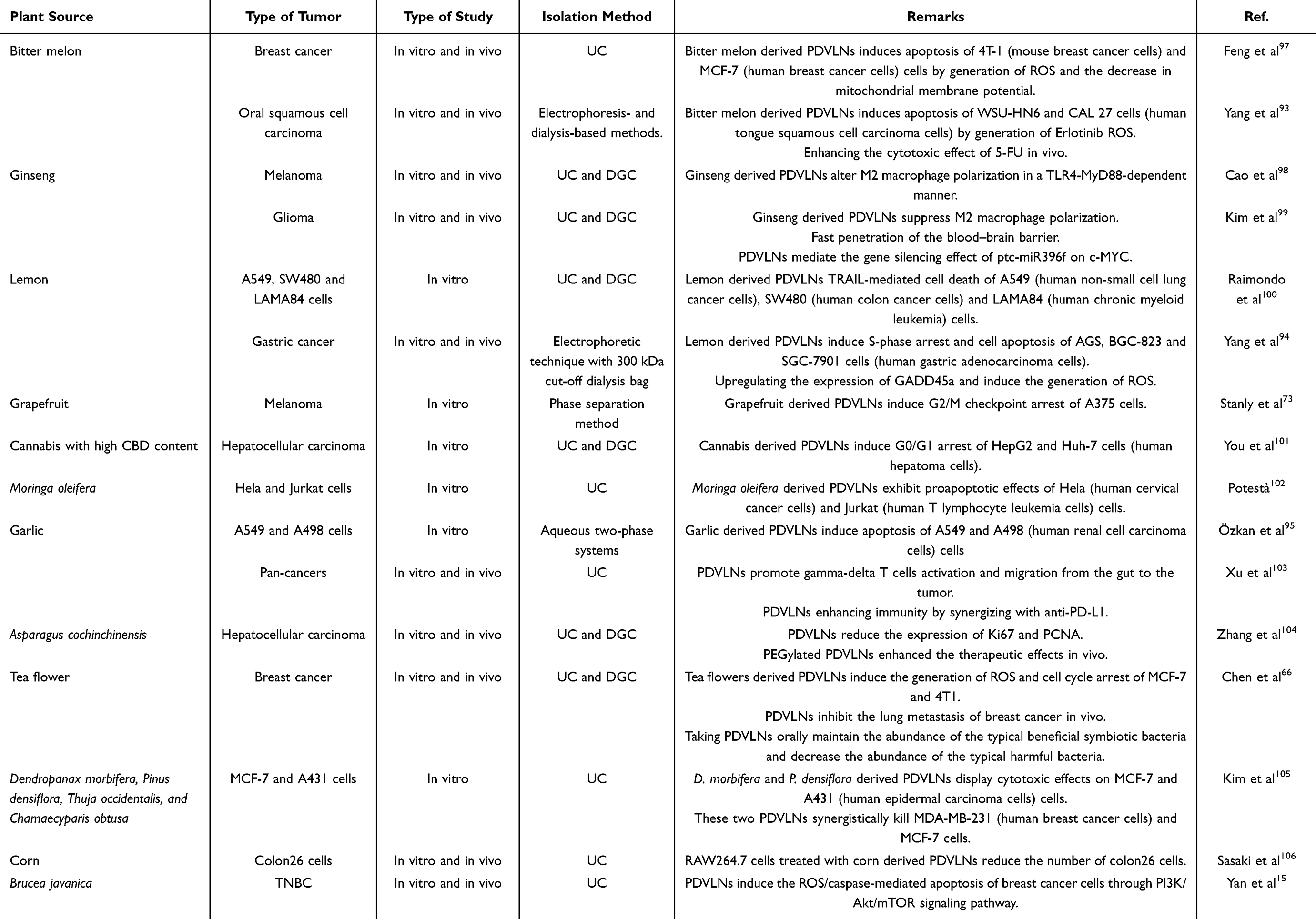 |
Table 2 The Current Use of Plant Derived Vesicle Like Nanoparticles for Tumor Treatment |
Inducing the Apoptosis of Cancer Cells
Although many PDVLNs from various plants have been reported to have anticancer activity,95,107,108 the underlying mechanisms have not been well demonstrated, and further research is needed. Briefly, the currently reported anticancer mechanisms of PDVLNs mainly involve three aspects: apoptosis, cell cycle arrest, and immune regulation. Most studies have reported the ability of PDVLNs to induce apoptosis in cancer cells, although the specific signaling pathways vary across studies. Bitter melon derived PDVLNs stimulate the generation of intracellular reactive oxygen species (ROS) in breast cancer cells, thus inducing apoptosis.97 In this study, the fluorescence intensity of DCFH-DA (20,70–dichlorofluorescin diacetate) was used to detect the level of ROS, which is a commonly used method in PDVLNs research. ROS-induced apoptosis is completed within 60 min in a caspase-dependent manner.109 Indeed, radiotherapy and chemotherapy induce the accumulation of ROS in cancer cells, resulting in apoptosis of cancer cells,110 and many types of anticancer agents can generate oxidative stress. For example, Erlotinib and Imatinib induce ROS-dependent apoptosis in non-small-cell lung cancer cells and melanoma, respectively.111,112 Furthermore, 5-fluorouracil (5-FU) induces the accumulation of ROS in a p53-dependent manner, leading to apoptosis in colorectal cancer cells.113 Interestingly, Yang et al found that bitter melon derived PDVLNs induced the apoptosis of oral squamous cell carcinoma by ROS and synergistically enhanced the cytotoxic effects of 5-FU.93 Although they pointed out that the synergistic effect was generated by the PDVLNs mediated downregulation of NLRP3, PDVLNs induced ROS may also play a role in this effect. Therefore, PDVLNs from various plants may act as promising adjuvants for enhancing the therapeutic effects of chemotherapy. Apart from ROS, Chen et al found that the levels of intracellular superoxide anions and reactive nitrogen species also increased in tea flower derived PDVLNs treated breast cancer cells.66 In the meantime, Chen et al attempted to identify the substances responsible for the induction of ROS. Using liquid chromatography coupled with tandem mass spectrometry, they found several flavonoids and polyphenols, such as epicatechin gallate and epicatechin, in tea flower derived PDVLNs, which have been reported to induce increased oxidative stress in cancer cells.114,115 Another proapoptotic effect of PDVLNs is mitochondrial damage. Although ROS can also induce mitochondrial damage,116 little is known about whether this damage is caused by ROS or PDVLNs. JC-1 (5,50,6,60-tetrachloro-1,10,3,30-tetraethyl-imidacarbocyanine iodide) assay was used to detect the membrane potential of mitochondrial to evaluate the degree of mitochondrial damage.66,97 Specifically, lemon derived PDVLNs can induce cancer cell apoptosis via the tumor necrosis factor related apoptosis-inducing ligand (TRAIL)/Dr5 pathway.100 A latest study indicated that Brucea javanica (a medicinal plant) derived PDVLNs could induce the ROS/caspase-mediated apoptosis of breast cancer cells through the PI3K/Akt/mTOR signaling pathway.15
Induction of Cell Cycle Arrest
The second anticancer effect of PDVLNs is cell cycle arrest. Several plants derived PDVLNs have exhibited the ability to induce cell cycle arrest of cancer cells. However, the specific arrest phases are different. Lemon derived PDVLNs can induce the cell cycle in gastric cancer cells at S phase.94 Kyoto Encyclopedia of Genes and Genomes (KEGG) analysis revealed that this cell cycle arrest effect might be caused by the GADD45α protein, and after treatment with lemon derived PDVLNs, the expression level of GADD45α increased. It is reported that GADD45α could inhibit the G2/M checkpoint and arrest the cell cycle at G2.117 Moreover, GADD45α can induce S-phase cell cycle arrest and further inhibit the proliferation of human hepatoma G2 cells.118 Another study on grapefruit derived PDVLNs indicated that PDVLNs induce the G2/M arrest of A375 cells (human melanoma cells).73 This effect is attributed to the dysregulation of cyclin B and CycB2. Cyclin B and CycB2 are two essential proteins that participate in the G2/M cell cycle progression, and it is reported that the downregulation of CycB2 inhibits the proliferation and metastasis of cancer cells.119,120 In general, PDVLNs derived from citrus plants appear to have a favorable ability to induce cell cycle arrest of cancer cells, and further research is needed in the future. Similarly, tea flower derived PDVLNs also induce G2/M arrest of cancer cells. Tajik et al reported that cannabis derived PDVLNs induced G0/G1 arrest of hepatocellular carcinoma cells, which might be attributed to the alteration of other cell cycle regulatory proteins.101
Regulating the Immune Microenvironment of Tumor Tissue
The immune regulatory effects of PDVLNs mainly involve two immune cells currently, namely, macrophages and T cells. Most studies have reported on the regulation of macrophages. Two studies reported the alteration of M2 macrophages by ginseng derived PDVLNs. Cao et al conducted a comprehensive study on the polarization of macrophages induced by ginseng derived PDVLNs. They found that PDVLNs altered the polarization of M2-like macrophages and inhibited melanoma growth. Using flow cytometry, they verified the polarization and function of the M1-like macrophages in vivo.98 Moreover, they found that when lacking the surface proteins (treated with proteinase K), PDVLNs did not induce alterations in M2 macrophages. Another study reported that ginseng derived PDVLNs could suppress M2 macrophage polarization, thus suppressing glioma growth.99 Meanwhile, the authors reported an alteration of T cells in tumor microenvironment, and after treatment with ginseng derived PDVLNs, the number of CD8+ T cells significantly increased, while the abundance of Treg cells (FoxP3+ and CD25+) decreased. Sasaki et al reported that corn derived PDVLNs altered the polarization of macrophages by using RAW264.7 cells (mouse monocyte macrophage leukemia cells).106 Xu et al have described the status of intestinal immune cells following oral administration of garlic PDVLNs through single-cell sequencing.103 They found that oral administration of PDVLNs could promote proliferation and activation of intestine gamma-delta T cells, thereby increasing the secretion of IFN-γ. At the same time, activated gamma-delta T increased the expression of CXCR3, which prompted them to migrate from the intestine to the tumor tissue.
Other potential anticancer effects include specific bioactive molecules (such as non-coding RNA) and regulation of the gut microbiota. Accumulating evidence has demonstrated that plant-derived miRNAs exist in the human body and play a role in human health.121,122 It has been reported that plants derived miRNAs are administered orally and are observed in the intestine.123 Due to the characteristics of interspecies communication of PDVLNs, the manner in which these miRNAs enter human bodies may also be mediated by PDVLNs. PDVLNs contain several RNAs (such as miRNAs),124,125 and these miRNAs vary among different source plants or parts of plants. Many plant-derived miRNAs have been shown to exert anticancer effects in humans.126,127 Therefore, exploring potential plants that contain anticancer miRNAs may be a promising method for identifying new PDVLNs for tumor therapy.
Another potential anticancer effect is the regulation of the gut microbiota. The oral intake of PDVLNs is considered a safe method for absorbing PDVLNs and rarely causes adverse effects. Several studies have used oral administration of PDVLNs to treat intestinal diseases, such as colitis.128 Orally administered tea flower-derived PDVLNs have been reported to remodel the gut microbiota and may play a role in anticancer effects.66 Teng et al also showed that PDVLNs could regulate gut microbiota and alleviate specific diseases.129
In general, PDVLNs are promising agents or tools for tumor therapy. Compared with mammal derived EVs, PDVLNs themselves have anticancer effects. However, unlike mammal derived EVs (cell line derived EVs), PDVLNs are extracted directly from plants rather than from cultured cells. This approach allows for the extraction of large quantities of PDVLNs in each extraction, but it also makes it difficult to genetically engineer PDVLNs.130 For example, engineered mammal derived EVs can express specific targeted proteins on their surfaces to achieve targeted delivery of drugs or fluorescent molecules to tumors.131 Notably, some PDVLNs show great biosafety because they significantly inhibit the proliferation of cancer cells while having no effect on normal cells,66,100 which compensates for the shortcomings of insufficient targeting to some extent. The engineering of PDVLNs is in its early stages, and there is still a long way to go in the future. In the next section, we review the current articles on the engineering of PDVLNs and provide insights into their future development.
Methods for PDVLNs Engineering
Although some PDVLNs were discovered to be tumor-suppressive on their own, researchers have proposed various methods to modify PDVLNs to improve their targeting ability and anticancer effects.
Loading Cargoes into PDVLNs
A crucial aspect in evaluating the feasibility of PDVLNs as delivery platforms is their ability to carry therapeutic drugs. Hydrophilic drugs can be encapsulated into PDVLNs’ cavity, whereas hydrophobic drugs can be loaded onto the lipid bilayer membrane. The drug loading strategies employed by PDVLNs are similar to those used by mammal-derived EVs, such as co-incubation, ultrasound, electroporation, freeze-thaw cycles, and co-extrusion.132,133
Co-incubation utilizes diffusion and lipophilic interactions between therapeutic drugs and the lipid bilayer of the PDVLNs. Xiao et al co-incubated doxorubicin (DOX) with lemon-derived PDVLNs and proved that encapsulation did not impair the anticancer efficiency of DOX while reducing its side effects.134 Umezu et al also attempted to package miRNA into cherry derived PDVLNs through co-incubation and observed downregulation of the miRNA target gene after oral administration.135 Although co-incubation is a widely used and simple method for drug loading, some limitations must be considered. First, co-incubation is a passive process that lacks additional forces to promote diffusion, which usually results in a relatively low encapsulation efficiency. In addition, the encapsulation efficiency may also be influenced by the characteristics of the loaded drugs because lipophilic and oppositely charged molecules make it easier to approach PDVLNs.136,137
Ultrasound, freeze-thaw cycles, and electroporation have similar mechanisms for drug loading. Ultrasound can temporarily damage the lipid bilayer of PDVLNs using mechanical energy and increase the chances for drugs to penetrate the PDVLNs.93 Similarly, freeze-thaw cycles and electroporation can form pores in the lipid bilayer, allowing drugs to enter.138 Yang et al used ultrasound to load 5- fluorouracil (5-FU) into bitter melon derived PDVLNs to enhance the therapeutic effects of 5-FU on oral squamous cell carcinoma.93 Kilasoniya et al loaded heat shock protein 70 (HSP70) into grapefruit derived PDVLNs using an ultrasonic bath and demonstrated its anti-tumor effect on glioma cells.139 These methods show a higher encapsulation efficiency than co-incubation.93,138,140 However, the damage can distort the integrity of the lipid bilayer membrane. Freeze-thaw cycles have been reported to cause less damage to vesicles and have simpler operation methods than ultrasound, but frequent temperature changes may lead to protein inactivation and an increase in vesicle size.141 Electroporation is usually applied for loading small molecules, such as siRNA, and the loading efficiency can be adjusted by adjusting the EVs, drug ratio, and voltage supplied.8
Co-extrusion refers to the extrusion of PDVLNs and drugs through a polycarbonate membrane filter, and the diameter of the obtained nanovesicles can be regulated by controlling the pore size of the filter.142 This method can produce uniform-sized vesicles with a satisfactory loading efficiency. Zhang et al encapsulated siRNA-CD98 into ginger derived PDVLNs by co-extrusion and successfully reduced the expression of CD98 in colon tissues.143 Co-extrusion can also be used to modify PDVLNs by fusing their membranes with those of other vesicles. However, this method also has several limitations. Repeated co-extrusion process may impair the biological activity of PDVLNs, which has been proved by Fuhrmann’s study.136 Loss of lipid bilayers on the filter membrane is also a vital issue that cannot be ignored.144 In addition, the microfluidic method has also been recently reported as a method for drug loading. Shoko et al used microfluidic method to load siRNA into grapefruit derived PDVLNs, achieving a loading efficiency of 11% and successfully reducing the expression of target gene in HaCaT cells.145 The advantage of microfluidic methods is that they can control the diameter of nanoparticles to obtain stable nanoparticles.
In general, these are the main strategies for loading anti-tumor molecules into PDVLNs. Co-incubation can protect the integrity of the membrane, but has a lower loading efficiency. In contrast, ultrasound, freeze-thaw cycles, and electroporation have better loading efficiency, but damage the membrane of nanoparticles. Co-extrusion can produce uniform-sized vesicles but may diminish the biological activity of PDVLNs. The characteristics of anti-tumor molecules can also affect the loading process. Researchers should take these aspects into consideration before deciding on a proper encapsulation method.
Modifying PDVLNs
Unlike mammal-derived EVs isolated from cell culture medium, PDVLNs are directly isolated from plants, indicating the absence of a cell culture process. This characteristic makes PDVLNs easier to isolate and less costly but blocks the possibility of engineering PDVLNs from their biogenesis, such as using lentivirus to transduce genes into cells and express targeting peptides on their membrane. The culture of plant cells is a complex and expensive process. Thus, it is rarely considered in PDVLN production. Most studies that engineered PDVLNs have focused on modifying their lipid bilayer membranes. Table 3 compares the advantages and disadvantages of the three types of PDVLNs in terms of preparation and cost.
 |
Table 3 Comparison of Three Kinds of PDVLNs in Property |
To enhance the targeting capability of PDVLNs, a common strategy involves immobilizing molecules such as cyclic arginine-glycine-aspartic acid (cRGD) onto their surface. cRGD can recognize and competitively bind to αvβ3 integrins that are highly expressed on the surface of many solid tumors,146 which can also block the adhesion of tumor cells to the extracellular matrix or directly induce tumor cell apoptosis. Taking advantage of these features, Chen et al modified lemon derived PDVLNs with cRGD using the EDC/NHS chemistry coupling method and identified an increased accumulation at the tumor site.147 Some studies have also applied heparin as a bridge between cRGD and PDVLNs since it can increase the stability and biocompatibility of EVs in vivo owing to its good anti-complement activation ability.134 Niu et al first linked pH-sensitive adipic acid dihydrazide (ADH) to DOX to promote its release in an acidic tumor microenvironment. They then conjugated ADH-DOX and cRGD to the carboxyl group of heparin. The remaining carboxyl groups of heparin were then bound to the active amino groups on the membrane of grapefruit derived PDVLNs to establish a novel delivery platform that could not only target gliomas but also control drug release.148 Folic acid (FA) is another frequently used targeting ligand, since folate receptors are reported to be highly expressed in many cancer cells but have limited distribution in normal organs.149 Zhang et al mixed total lipids extracted from ginger derived nanovesicles (GDNV) with DOX and FA in dimethyl sulfoxide (DMSO) in chloroform, and then dried the mixture under nitrogen to obtain a thin lipid film. A standard method based on lipid film hydration was applied to obtain DOX-FA-GDNVs, which exhibited an improved targeting ability towards colon tumors.137
Another strategy for modifying PDVLNs involves the infusion of different types of lipid membranes. Wang et al coated grapefruit derived nanovector with a leukocyte-derived plasma membrane (IGNV) by extruding the mixture 20 times through a 200 nm polycarbonate membrane. They found that IGNV showed an enhanced ability to home to inflammatory tumor tissues.150 Chen et al proposed a conceptually novel design of a lemon derived PDVLNs engineered structural droplet drug (ESDD). They mixed cRGD- PDVLNs with DOX in a PBS solution as the aqueous phase, used squalene as the organic phase, and then used an ultrasonic emulsification method to fabricate the ESDD. The structure of the ESDD was similar to that of encapsulating DOX into a squalene droplet and coating it with cRGD-PDVLNs. ESDD could carry more DOX than PDVLNs and had better deformability, which could benefit the transcytosis process and improve its penetration into tumor tissues. ESDD also inherits the advantages of PDVLNs, such as their good bioavailability and targeting capability.147
Clinical Trials of PDVLNs
Currently, research on PDVLNs is primarily preclinical. Given the promising performance of PDVLNs in disease treatment, there is an urgent need to know whether PDVLNs will demonstrate good efficacy and safety when applied in humans. There are currently four registered clinical trials related to PDVLNs, two of which have been completed, one is recruiting participants, and one had to be withdrawn due to lack of patient enrollment. Table 4 summarizes the details of these four clinical trials.
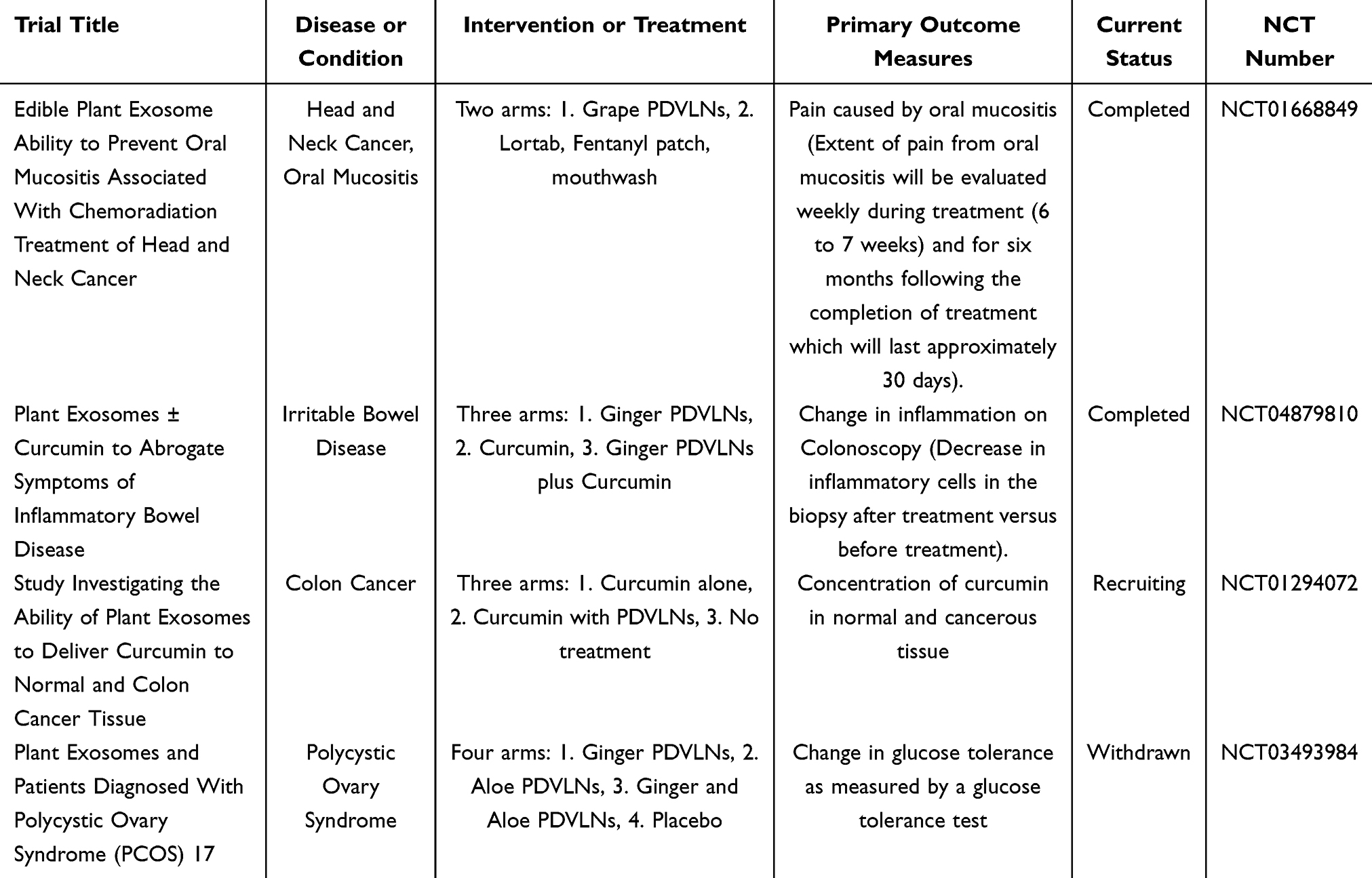 |
Table 4 Clinical Trials of PDVLNs |
Conclusion and Prospective
Before the first successful isolation of PDVLNs, plant-based anticancer therapy was mainly focused on the extraction of specific anticancer molecules or adjuvant therapy with other therapies. Although these two methods have yielded valuable achievements, the development of plant-based anticancer therapies has been limited by several factors. Along with the successful isolation of PDVLNs, plant-based anticancer therapy is a new development. Several plants with potential medicinal properties have been used to treat cancer. Currently, PDVLNs are primarily isolated from fruits and vegetables. Given the diversity of medicinal plants used for cancer therapy, medicinal plant-derived PDVLNs can be isolated in future research to further concentrate on effective substances and reduce their toxicities. Bioactive molecules in PDVLNs are derived from source plants. Therefore, plants that have been proven to have active molecules that work against specific diseases may be the focus of future PDVLNs research.
PDVLNs have emerged as promising agents and tools for cancer therapies. PDVLNs exhibit ant-cancer effects and can act as anticancer agents. Moreover, similar to mammal derived EVs, PDVLNs can be designed as drug delivery platforms to carry drugs to the lesion area. The characteristic of PDVLNs to resist digestive enzymes in the gastrointestinal tract makes them ideal for oral medicine. However, compared to mammal derived EVs, research on PDVLNs is still in its infancy, and many problems need to be further studied. Although it is relatively easy to obtain PDVLNs that can be isolated directly from plant leaves, seeds, and stems of fruit juice, the process of planting plants requires a lot of manpower and resources. In the meantime, different soils, climates, and seasons may have an impact on the production of plants and the abundance of bioactive molecules. It is currently unclear whether these natural factors affect the composition of the PDVLNs. The isolation methods for PDVLNs are highly similar to those used for mammal derived EVs. However, this does not mean that these methods are suitable for extracting PDVLNs, because they originate from plants.
To further improve the quality of PDVLNs research and enhance the translational applications of PDVLNs, efforts can be made in several directions. The first is the development of more suitable isolation methods for PDVLNs and the improvement of the yield and purity of PDVLNs. Exploring the most suitable isolation method for specific plants. Second, qualitative and quantitative analyses of PDVLN components and the identification of effective substances in PDVLNs were performed. Multi-omics analysis can help to explore the differences in components among different PDVLNs and the mechanisms of PDVLNs in the biological effects of PDVLNs. Quantitative analysis of specific bioactive molecules can aid the translational application of PDVLNs. Third, comprehensively evaluating the safety of PDVLNs. Although some studies have reported the selective killing effect of PDVLNs on tumor cells, it is unclear whether PDVLNs are safe for all normal cells. Due to the complexity of PDVLNs components, the potential interactions between PDVLNs and other drugs should be carefully investigated.
Data Sharing Statement
All relevant data are within the paper.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This work was supported by the China Postdoctoral Science Foundation (2023M741278), Key R & D Program of Hubei Province (2023BCB149), Tongji Hospital Medical Innovation and Transformation Incubation Project (2022ZHFY02), and the National Natural Science Foundation of China (NSFC, 82370770).
Disclosure
The authors have declared no conflicts of interest.
References
1. Couch Y, Buzàs EI, Di Vizio D, et al. A brief history of nearly EV-erything – the rise and rise of extracellular vesicles. J Extracell Vesicles. 2021;10(14):e12144. doi:10.1002/jev2.12144
2. Raposo G, Stoorvogel W. Extracellular vesicles: exosomes, microvesicles, and friends. J Cell Biol. 2013;200(4):373–383. doi:10.1083/jcb.201211138
3. Lai RC, Arslan F, Lee MM, et al. Exosome secreted by MSC reduces myocardial ischemia/reperfusion injury. Stem Cell Res. 2010;4(3):214–222. doi:10.1016/j.scr.2009.12.003
4. Zhang J, Guan J, Niu X, et al. Exosomes released from human induced pluripotent stem cells-derived MSCs facilitate cutaneous wound healing by promoting collagen synthesis and angiogenesis. J Transl Med. 2015;13:49. doi:10.1186/s12967-015-0417-0
5. Zhang X, Zhang H, Gu J, et al. Engineered Extracellular Vesicles for Cancer Therapy. Adv Mater. 2021;33(14):e2005709. doi:10.1002/adma.202005709
6. Morad G, Carman CV, Hagedorn EJ, et al. Tumor-derived extracellular vesicles breach the intact blood-brain barrier via transcytosis. ACS Nano. 2019;13(12):13853–13865. doi:10.1021/acsnano.9b04397
7. Wiklander OP, Nordin JZ, O’Loughlin A, et al. Extracellular vesicle in vivo biodistribution is determined by cell source, route of administration and targeting. J Extracell Vesicles. 2015;4:26316. doi:10.3402/jev.v4.26316
8. Alvarez-Erviti L, Seow Y, Yin H, Betts C, Lakhal S, Wood MJA. Delivery of siRNA to the mouse brain by systemic injection of targeted exosomes. Nat Biotechnol. 2011;29(4):341–345. doi:10.1038/nbt.1807
9. Elsharkasy OM, Nordin JZ, Hagey DW, et al. Extracellular vesicles as drug delivery systems: why and how? Adv Drug Deliv Rev. 2020;159:332–343. doi:10.1016/j.addr.2020.04.004
10. Murphy DE, de Jong OG, Brouwer M, et al. Extracellular vesicle-based therapeutics: natural versus engineered targeting and trafficking. Exp mol Med. 2019;51(3):1–12. doi:10.1038/s12276-019-0223-5
11. McVey MJ, Weidenfeld S, Maishan M, et al. Platelet extracellular vesicles mediate transfusion-related acute lung injury by imbalancing the sphingolipid rheostat. Blood. 2021;137(5):690–701. doi:10.1182/blood.2020005985
12. Herrmann IK, Wood MJA, Fuhrmann G. Extracellular vesicles as a next-generation drug delivery platform. Nat Nanotechnol. 2021;16(7):748–759. doi:10.1038/s41565-021-00931-2
13. Regente M, Corti-Monzón G, Maldonado AM, Pinedo M, Jorrín J, de la Canal L. Vesicular fractions of sunflower apoplastic fluids are associated with potential exosome marker proteins. FEBS Lett. 2009;583(20):3363–3366. doi:10.1016/j.febslet.2009.09.041
14. An Q, van Bel AJ, Hückelhoven R. Do plant cells secrete exosomes derived from multivesicular bodies? Plant Signal Behav. 2007;2(1):4–7. doi:10.4161/psb.2.1.3596
15. Yan G, Xiao Q, Zhao J, et al. Brucea javanica derived exosome-like nanovesicles deliver miRNAs for cancer therapy. J Control Release. 2024;367:425–440. doi:10.1016/j.jconrel.2024.01.060
16. Wang X, Liu Y, Dong X, et al. peu-MIR2916-p3-enriched garlic exosomes ameliorate murine colitis by reshaping gut microbiota, especially by boosting the anti-colitic Bacteroides thetaiotaomicron. Pharmacol Res. 2024;200:107071. doi:10.1016/j.phrs.2024.107071
17. Fabricant DS, Farnsworth NR. The value of plants used in traditional medicine for drug discovery. Environ Health Perspect. 2001;109(Suppl 1):69–75. doi:10.1289/ehp.01109s169
18. Yang AK, He SM, Liu L, Liu JP, Wei MQ, Zhou SF. Herbal interactions with anticancer drugs: mechanistic and clinical considerations. Curr Med Chem. 2010;17(16):1635–1678. doi:10.2174/092986710791111279
19. Su XL, Wang JW, Che H, et al. Clinical application and mechanism of traditional Chinese medicine in treatment of lung cancer. Chinese Med J. 2020;133(24):2987–2997. doi:10.1097/CM9.0000000000001141
20. Li Z, Feiyue Z, Gaofeng L. Traditional Chinese medicine and lung cancer--From theory to practice. Biomed Pharmacother. 2021;137:111381. doi:10.1016/j.biopha.2021.111381
21. Yang Z, Zhang Q, Yu L, Zhu J, Cao Y, Gao X. The signaling pathways and targets of traditional Chinese medicine and natural medicine in triple-negative breast cancer. J Ethnopharmacol. 2021;264:113249. doi:10.1016/j.jep.2020.113249
22. Cao L, Wang X, Zhu G, et al. Traditional Chinese medicine therapy for esophageal cancer: a literature review. Integr Cancer Ther. 2021;20:15347354211061720. doi:10.1177/15347354211061720
23. Zhang X, Qiu H, Li C, Cai P, Qi F. The positive role of traditional Chinese medicine as an adjunctive therapy for cancer. Biosci Trends. 2021;15(5):283–298. doi:10.5582/bst.2021.01318
24. Dini I, Grumetto L. Recent advances in natural polyphenol research. Molecules. 2022;27(24):8777. doi:10.3390/molecules27248777
25. Azmi AS, Bhat SH, Hanif S, Hadi SM. Plant polyphenols mobilize endogenous copper in human peripheral lymphocytes leading to oxidative DNA breakage: a putative mechanism for anticancer properties. FEBS Lett. 2006;580(2):533–538. doi:10.1016/j.febslet.2005.12.059
26. Apostolou A, Stagos D, Galitsiou E, et al. Assessment of polyphenolic content, antioxidant activity, protection against ROS-induced DNA damage and anticancer activity of Vitis vinifera stem extracts. Food Chem Toxicol. 2013;61:60–68. doi:10.1016/j.fct.2013.01.029
27. Coussens LM, Werb Z. Inflammation and cancer. Nature. 2002;420(6917):860–867. doi:10.1038/nature01322
28. Abu Samaan TM, Samec M, Liskova A, Kubatka P, Büsselberg D. Paclitaxel’s mechanistic and clinical effects on breast cancer. Biomolecules. 2019;9(12):789. doi:10.3390/biom9120789
29. Hashemi M, Zandieh MA, Talebi Y, et al. Paclitaxel and docetaxel resistance in prostate cancer: molecular mechanisms and possible therapeutic strategies. Biomed Pharmacother. 2023;160:114392. doi:10.1016/j.biopha.2023.114392
30. Yang YH, Mao JW, Tan XL. Research progress on the source, production, and anti-cancer mechanisms of paclitaxel. Chin J Nat Med. 2020;18(12):890–897. doi:10.1016/S1875-5364(20)60032-2
31. Salerni BL, Bates DJ, Albershardt TC, Lowrey CH, Eastman A. Vinblastine induces acute, cell cycle phase-independent apoptosis in some leukemias and lymphomas and can induce acute apoptosis in others when Mcl-1 is suppressed. Mol Cancer Ther. 2010;9(4):791–802. doi:10.1158/1535-7163.MCT-10-0028
32. Rovsing AB, Thomsen EA, Nielsen I, et al. Resistance to vincristine in DLBCL by disruption of p53-induced cell cycle arrest and apoptosis mediated by KIF18B and USP28. Br J Haematol. 2023;202(4):825–839. doi:10.1111/bjh.18872
33. Çoban EA, Tecimel D, Şahin F, Deniz AAH. Targeting cancer metabolism and cell cycle by plant-derived compounds. Adv Exp Med Biol. 2020;1247:125–134.
34. Wong RS. Apoptosis in cancer: from pathogenesis to treatment. J Exp Clin Cancer Res. 2011;30(1):87. doi:10.1186/1756-9966-30-87
35. Ren B, Kwah MX, Liu C, et al. Resveratrol for cancer therapy: challenges and future perspectives. Cancer Lett. 2021;515:63–72. doi:10.1016/j.canlet.2021.05.001
36. Bhutia SK, Panda PK, Sinha N, et al. Plant lectins in cancer therapeutics: targeting apoptosis and autophagy-dependent cell death. Pharmacol Res. 2019;144:8–18. doi:10.1016/j.phrs.2019.04.001
37. Yang Y, Chen Y, Wu JH, et al. Targeting regulated cell death with plant natural compounds for cancer therapy: a revisited review of apoptosis, autophagy-dependent cell death, and necroptosis. Phytother Res. 2023;37(4):1488–1525. doi:10.1002/ptr.7738
38. Xu C, Wen S, Du X, et al. Targeting regulated cell death (RCD) with naturally derived sesquiterpene lactones in cancer therapy. Pharmacol Res. 2025;211:107553. doi:10.1016/j.phrs.2024.107553
39. Han YH, Mun JG, Jeon HD, Kee JY, Hong SH. Betulin inhibits lung metastasis by inducing cell cycle arrest, autophagy, and apoptosis of metastatic colorectal cancer cells. Nutrients. 2019;12(1):66. doi:10.3390/nu12010066
40. Lou JS, Zhao LP, Huang ZH, et al. Ginkgetin derived from Ginkgo biloba leaves enhances the therapeutic effect of cisplatin via ferroptosis-mediated disruption of the Nrf2/HO-1 axis in EGFR wild-type non-small-cell lung cancer. Phytomedicine. 2021;80:153370. doi:10.1016/j.phymed.2020.153370
41. Rashidi B, Malekzadeh M, Goodarzi M, Masoudifar A, Mirzaei H. Green tea and its anti-angiogenesis effects. Biomed Pharmacother. 2017;89:949–956. doi:10.1016/j.biopha.2017.01.161
42. Wang Y, Li J, Xia L. Plant-derived natural products and combination therapy in liver cancer. Front Oncol. 2023;13:1116532. doi:10.3389/fonc.2023.1116532
43. Efferth T, Li PC, Konkimalla VS, Kaina B. From traditional Chinese medicine to rational cancer therapy. Trends Mol Med. 2007;13(8):353–361. doi:10.1016/j.molmed.2007.07.001
44. Wang D, Calabrese EJ, Lian B, Lin Z, Calabrese V. Hormesis as a mechanistic approach to understanding herbal treatments in traditional Chinese medicine. Pharmacol Ther. 2018;184:42–50. doi:10.1016/j.pharmthera.2017.10.013
45. Wang Y, Zhang Q, Chen Y, et al. Antitumor effects of immunity-enhancing traditional Chinese medicine. Biomed Pharmacother. 2020;121:109570. doi:10.1016/j.biopha.2019.109570
46. Xu F, Cui WQ, Wei Y, et al. Astragaloside IV inhibits lung cancer progression and metastasis by modulating macrophage polarization through AMPK signaling. J Exp Clin Cancer Res. 2018;37(1):207. doi:10.1186/s13046-018-0878-0
47. Fujiwara Y, Shiraishi D, Yoshitomi M, et al. Soyasapogenols contained in soybeans suppress tumour progression by regulating macrophage differentiation into the protumoural phenotype. J Funct Foods. 2015;19:594–605. doi:10.1016/j.jff.2015.09.055
48. Kaneko M, Kawakita T, Tauchi Y, Saito Y, Suzuki A, Nomoto K. Augmentation of NK activity after oral administration of a traditional Chinese medicine, Xiao-chai-hu-tang (Shosaiko-to). Immuno Immunotoxicol. 1994;16(1):41–53. doi:10.3109/08923979409029899
49. Wu X-T, Liu J-Q, Lu X-T, et al. The enhanced effect of lupeol on the destruction of gastric cancer cells by NK cells. Int Immunopharmacol. 2013;16(2):332–340. doi:10.1016/j.intimp.2013.04.017
50. Zhang Y, Wei Y, Jiang S, et al. Traditional Chinese medicine CFF-1 exerts a potent anti-tumor immunity to hinder tumor growth and metastasis in prostate cancer through EGFR/JAK1/STAT3 pathway to inhibit PD-1/PD-L1 checkpoint signaling. Phytomedicine. 2022;99:153939. doi:10.1016/j.phymed.2022.153939
51. Esmeeta A, Adhikary S, Dharshnaa V, et al. Plant-derived bioactive compounds in colon cancer treatment: an updated review. Biomed Pharmacother. 2022;153:113384. doi:10.1016/j.biopha.2022.113384
52. Buyel JF. Plants as sources of natural and recombinant anti-cancer agents. Biotechnol Adv. 2018;36(2):506–520. doi:10.1016/j.biotechadv.2018.02.002
53. Feng J, Xiu Q, Huang Y, Troyer Z, Li B, Zheng L. Plant-derived vesicle-like nanoparticles as promising biotherapeutic tools: present and future. Adv Mater. 2023;35(24):e2207826. doi:10.1002/adma.202207826
54. Halperin W, Jensen WA. Ultrastructural changes during growth and embryogenesis in carrot cell cultures. J Ultrastruct Res. 1967;18(3):428–443. doi:10.1016/S0022-5320(67)80128-X
55. Cai H, Huang LY, Hong R, et al. Momordica charantia exosome-like nanoparticles exert neuroprotective effects against ischemic brain injury via inhibiting matrix Metalloproteinase 9 and activating the AKT/GSK3β signaling pathway. Front Pharmacol. 2022;13:908830. doi:10.3389/fphar.2022.908830
56. Sundaram K, Mu J, Kumar A, et al. Garlic exosome-like nanoparticles reverse high-fat diet induced obesity via the gut/brain axis. Theranostics. 2022;12(3):1220–1246. doi:10.7150/thno.65427
57. Bhattacharjee S. DLS and zeta potential - What they are and what they are not? J Control Release. 2016;235:337–351. doi:10.1016/j.jconrel.2016.06.017
58. Gardiner C, Ferreira YJ, Dragovic RA, Redman CW, Sargent IL. Extracellular vesicle sizing and enumeration by nanoparticle tracking analysis. J Extracell Vesicles. 2013;2.
59. Mu J, Zhuang X, Wang Q, et al. Interspecies communication between plant and mouse gut host cells through edible plant derived exosome-like nanoparticles. mol Nutr Food Res. 2014;58(7):1561–1573. doi:10.1002/mnfr.201300729
60. Stanly C, Moubarak M, Fiume I, Turiák L, Pocsfalvi G. Membrane transporters in citrus clementina fruit juice-derived nanovesicles. Int J mol Sci. 2019;20(24):6205. doi:10.3390/ijms20246205
61. Lian MQ, Chng WH, Liang J, et al. Plant-derived extracellular vesicles: recent advancements and current challenges on their use for biomedical applications. J Extracell Vesicles. 2022;11(12):e12283. doi:10.1002/jev2.12283
62. Liang Y, Lehrich BM, Zheng S, Lu M. Emerging methods in biomarker identification for extracellular vesicle-based liquid biopsy. J Extracell Vesicles. 2021;10(7):e12090. doi:10.1002/jev2.12090
63. Ma C, Liu K, Wang F, et al. Neutrophil membrane-engineered Panax ginseng root-derived exosomes loaded miRNA 182-5p targets NOX4/Drp-1/NLRP3 signal pathway to alleviate acute lung injury in sepsis: experimental studies. Int J Surg. 2024;110(1):72–86. doi:10.1097/JS9.0000000000000789
64. Prado N, Alché Jde D, Casado-Vela J, et al. Nanovesicles are secreted during pollen germination and pollen tube growth: a possible role in fertilization. Mol Plant. 2014;7(3):573–577. doi:10.1093/mp/sst153
65. Pérez-Bermúdez P, Blesa J, Soriano JM, Marcilla A. Extracellular vesicles in food: experimental evidence of their secretion in grape fruits. Eur J Pharm Sci. 2017;98:40–50. doi:10.1016/j.ejps.2016.09.022
66. Chen Q, Li Q, Liang Y, et al. Natural exosome-like nanovesicles from edible tea flowers suppress metastatic breast cancer via ROS generation and microbiota modulation. Acta Pharm Sin B. 2022;12(2):907–923. doi:10.1016/j.apsb.2021.08.016
67. Rutter BD, Innes RW. Extracellular vesicles isolated from the leaf apoplast carry stress-response proteins. Plant Physiol. 2017;173(1):728–741. doi:10.1104/pp.16.01253
68. Karamanidou T, Tsouknidas A. Plant-derived extracellular vesicles as therapeutic nanocarriers. Int J mol Sci. 2021;23(1):191. doi:10.3390/ijms23010191
69. Yuana Y, Koning RI, Kuil ME, et al. Cryo-electron microscopy of extracellular vesicles in fresh plasma. J Extracell Vesicles. 2013;2.
70. Eghiaian F, Rico F, Colom A, Casuso I, Scheuring S. High-speed atomic force microscopy: imaging and force spectroscopy. FEBS Lett. 2014;588(19):3631–3638. doi:10.1016/j.febslet.2014.06.028
71. Chen X, Zhou Y, Yu J. Exosome-like nanoparticles from ginger rhizomes inhibited NLRP3 inflammasome activation. Mol Pharm. 2019;16(6):2690–2699. doi:10.1021/acs.molpharmaceut.9b00246
72. Zhuang X, Deng ZB, Mu J, et al. Ginger-derived nanoparticles protect against alcohol-induced liver damage. J Extracell Vesicles. 2015;4:28713. doi:10.3402/jev.v4.28713
73. Stanly C, Alfieri M, Ambrosone A, Leone A, Fiume I, Pocsfalvi G. Grapefruit-derived micro and nanovesicles show distinct metabolome profiles and anticancer activities in the A375 human melanoma cell line. Cells. 2020;9(12):2722. doi:10.3390/cells9122722
74. You JY, Kang SJ, Rhee WJ. Isolation of cabbage exosome-like nanovesicles and investigation of their biological activities in human cells. Bioact Mater. 2021;6(12):4321–4332. doi:10.1016/j.bioactmat.2021.04.023
75. Pocsfalvi G, Turiák L, Ambrosone A, et al. Protein biocargo of citrus fruit-derived vesicles reveals heterogeneous transport and extracellular vesicle populations. J Plant Physiol. 2018;229:111–121. doi:10.1016/j.jplph.2018.07.006
76. Liu Y, Wu S, Koo Y, et al. Characterization of and isolation methods for plant leaf nanovesicles and small extracellular vesicles. Nanomedicine. 2020;29:102271. doi:10.1016/j.nano.2020.102271
77. Cong M, Tan S, Li S, et al. Technology insight: plant-derived vesicles-How far from the clinical biotherapeutics and therapeutic drug carriers? Adv Drug Deliv Rev. 2022;182:114108. doi:10.1016/j.addr.2021.114108
78. Bokka R, Ramos AP, Fiume I, et al. Biomanufacturing of tomato-derived nanovesicles. Foods. 2020;9(12):1852. doi:10.3390/foods9121852
79. Mammadova R, Maggio S, Fiume I, et al. Protein biocargo and anti-inflammatory effect of tomato fruit-derived nanovesicles separated by density gradient ultracentrifugation and loaded with curcumin. Pharmaceutics. 2023;15(2):333. doi:10.3390/pharmaceutics15020333
80. Seo K, Yoo JH, Kim J, et al. Ginseng-derived exosome-like nanovesicles extracted by sucrose gradient ultracentrifugation to inhibit osteoclast differentiation. Nanoscale. 2023;15(12):5798–5808. doi:10.1039/D2NR07018A
81. Lee R, Ko HJ, Kim K, et al. Anti-melanogenic effects of extracellular vesicles derived from plant leaves and stems in mouse melanoma cells and human healthy skin. J Extracell Vesicles. 2020;9(1):1703480. doi:10.1080/20013078.2019.1703480
82. Kim DK, Rhee WJ. Antioxidative effects of carrot-derived nanovesicles in cardiomyoblast and neuroblastoma cells. Pharmaceutics. 2021;13(8):1203. doi:10.3390/pharmaceutics13081203
83. Kalarikkal SP, Prasad D, Kasiappan R, Chaudhari SR, Sundaram GM. A cost-effective polyethylene glycol-based method for the isolation of functional edible nanoparticles from ginger rhizomes. Sci Rep. 2020;10(1):4456. doi:10.1038/s41598-020-61358-8
84. Webber J, Clayton A. How pure are your vesicles? J Extracell Vesicles. 2013;2.
85. Théry C, Witwer KW, Aikawa E, et al. Minimal information for studies of extracellular vesicles 2018 (MISEV2018): a position statement of the International Society for Extracellular Vesicles and update of the MISEV2014 guidelines. J Extracell Vesicles. 2018;7(1):1535750. doi:10.1080/20013078.2018.1535750
86. Gardiner C, Di Vizio D, Sahoo S, et al. Techniques used for the isolation and characterization of extracellular vesicles: results of a worldwide survey. J Extracell Vesicles. 2016;5:32945. doi:10.3402/jev.v5.32945
87. Li Z, Wang H, Yin H, Bennett C, Zhang HG, Guo P. Arrowtail RNA for ligand display on ginger exosome-like nanovesicles to systemic deliver siRNA for cancer suppression. Sci Rep. 2018;8(1):14644. doi:10.1038/s41598-018-32953-7
88. Liga A, Vliegenthart ADB, Oosthuyzen W, Dear JW, Kersaudy-Kerhoas M. Exosome isolation: a microfluidic road-map. Lab Chip. 2015;15(11):2388–2394. doi:10.1039/C5LC00240K
89. Gámez-Valero A, Monguió-Tortajada M, Carreras-Planella L, Franquesa M, Beyer K, Borràs FE. Size-exclusion chromatography-based isolation minimally alters extracellular vesicles’ characteristics compared to precipitating agents. Sci Rep. 2016;6:33641. doi:10.1038/srep33641
90. He B, Cai Q, Qiao L, et al. RNA-binding proteins contribute to small RNA loading in plant extracellular vesicles. Nat Plants. 2021;7(3):342–352. doi:10.1038/s41477-021-00863-8
91. Sitar S, Kejžar A, Pahovnik D, et al. Size characterization and quantification of exosomes by asymmetrical-flow field-flow fractionation. Anal Chem. 2015;87(18):9225–9233. doi:10.1021/acs.analchem.5b01636
92. Suresh AP, Kalarikkal SP, Pullareddy B, Sundaram GM. Low pH-based method to increase the yield of plant-derived nanoparticles from fresh ginger rhizomes. ACS Omega. 2021;6(27):17635–17641. doi:10.1021/acsomega.1c02162
93. Yang M, Luo Q, Chen X, Chen F. Bitter melon derived extracellular vesicles enhance the therapeutic effects and reduce the drug resistance of 5-fluorouracil on oral squamous cell carcinoma. J Nanobiotechnology. 2021;19(1):259. doi:10.1186/s12951-021-00995-1
94. Yang M, Liu X, Luo Q, Xu L, Chen F. An efficient method to isolate lemon derived extracellular vesicles for gastric cancer therapy. J Nanobiotechnology. 2020;18(1):100. doi:10.1186/s12951-020-00656-9
95. Özkan İ, Koçak P, Yıldırım M, et al. Garlic (Allium sativum)-derived SEVs inhibit cancer cell proliferation and induce caspase mediated apoptosis. Sci Rep. 2021;11(1):14773. doi:10.1038/s41598-021-93876-4
96. Savcı Y, Kırbaş OK, Bozkurt BT, et al. Grapefruit-derived extracellular vesicles as a promising cell-free therapeutic tool for wound healing. Food Funct. 2021;12(11):5144–5156. doi:10.1039/D0FO02953J
97. Feng T, Wan Y, Dai B, Liu Y. Anticancer activity of bitter melon-derived vesicles extract against breast cancer. Cells. 2023;12(6):824. doi:10.3390/cells12060824
98. Cao M, Yan H, Han X, et al. Ginseng-derived nanoparticles alter macrophage polarization to inhibit melanoma growth. J Immunother Cancer. 2019;7(1):326. doi:10.1186/s40425-019-0817-4
99. Kim J, Zhu Y, Chen S, et al. Anti-glioma effect of ginseng-derived exosomes-like nanoparticles by active blood-brain-barrier penetration and tumor microenvironment modulation. J Nanobiotechnology. 2023;21(1):253. doi:10.1186/s12951-023-02006-x
100. Raimondo S, Naselli F, Fontana S, et al. Citrus limon-derived nanovesicles inhibit cancer cell proliferation and suppress CML xenograft growth by inducing TRAIL-mediated cell death. Oncotarget. 2015;6(23):19514–19527. doi:10.18632/oncotarget.4004
101. Tajik T, Baghaei K, Moghadam VE, Farrokhi N, Salami SA. Extracellular vesicles of cannabis with high CBD content induce anticancer signaling in human hepatocellular carcinoma. Biomed Pharmacother. 2022;152:113209. doi:10.1016/j.biopha.2022.113209
102. Potestà M, Roglia V, Fanelli M, et al. Effect of microvesicles from Moringa oleifera containing miRNA on proliferation and apoptosis in tumor cell lines. Cell Death Discov. 2020;6:43. doi:10.1038/s41420-020-0271-6
103. Xu J, Yu Y, Zhang Y, et al. Oral administration of garlic-derived nanoparticles improves cancer immunotherapy by inducing intestinal IFNγ-producing γδ T cells. Nat Nanotechnol. 2024;19:1569–1578. doi:10.1038/s41565-024-01722-1
104. Zhang L, He F, Gao L, et al. Engineering exosome-like nanovesicles derived from Asparagus cochinchinensis can inhibit the proliferation of hepatocellular carcinoma cells with better safety profile. Int J Nanomed. 2021;16:1575–1586. doi:10.2147/IJN.S293067
105. Kim K, Yoo HJ, Jung JH, et al. Cytotoxic effects of plant sap-derived extracellular vesicles on various tumor cell types. J Funct Biomater. 2020;11(2):22. doi:10.3390/jfb11020022
106. Sasaki D, Kusamori K, Takayama Y, Itakura S, Todo H, Nishikawa M. Development of nanoparticles derived from corn as mass producible bionanoparticles with anticancer activity. Sci Rep. 2021;11(1):22818. doi:10.1038/s41598-021-02241-y
107. Han X, Wei Q, Lv Y, et al. Ginseng-derived nanoparticles potentiate immune checkpoint antibody efficacy by reprogramming the cold tumor microenvironment. Mol Ther. 2022;30(1):327–340. doi:10.1016/j.ymthe.2021.08.028
108. Chen Q, Zu M, Gong H, et al. Tea leaf-derived exosome-like nanotherapeutics retard breast tumor growth by pro-apoptosis and microbiota modulation. J Nanobiotechnology. 2023;21(1):6. doi:10.1186/s12951-022-01755-5
109. Perillo B, Di Donato M, Pezone A, et al. ROS in cancer therapy: the bright side of the moon. Exp Mol Med. 2020;52(2):192–203. doi:10.1038/s12276-020-0384-2
110. Brenneisen P, Reichert AS. Nanotherapy and Reactive Oxygen Species (ROS) in cancer: a novel perspective. Antioxidants. 2018;7(2). doi:10.3390/antiox7020031
111. Shan F, Shao Z, Jiang S, Cheng Z. Erlotinib induces the human non-small-cell lung cancer cells apoptosis via activating ROS-dependent JNK pathways. Cancer Med. 2016;5(11):3166–3175. doi:10.1002/cam4.881
112. Chang SP, Shen SC, Lee WR, Yang LL, Chen YC. Imatinib mesylate induction of ROS-dependent apoptosis in melanoma B16F0 cells. J Dermatol Sci. 2011;62(3):183–191. doi:10.1016/j.jdermsci.2011.03.001
113. Hwang PM, Bunz F, Yu J, et al. Ferredoxin reductase affects p53-dependent, 5-fluorouracil-induced apoptosis in colorectal cancer cells. Nat Med. 2001;7(10):1111–1117. doi:10.1038/nm1001-1111
114. Batra P, Sharma AK. Anti-cancer potential of flavonoids: recent trends and future perspectives. 3 Biotech. 2013;3(6):439–459. doi:10.1007/s13205-013-0117-5
115. Stepanic V, Gasparovic AC, Troselj KG, Amic D, Zarkovic N. Selected attributes of polyphenols in targeting oxidative stress in cancer. Curr Top Med Chem. 2015;15(5):496–509. doi:10.2174/1568026615666150209123100
116. Zeeshan M, Murugadas A, Ghaskadbi S, Ramaswamy BR, Akbarsha MA. Ecotoxicological assessment of cobalt using Hydra model: ROS, oxidative stress, DNA damage, cell cycle arrest, and apoptosis as mechanisms of toxicity. Environ Pollut. 2017;224:54–69. doi:10.1016/j.envpol.2016.12.042
117. Wang XW, Zhan Q, Coursen JD, et al. GADD45 induction of a G2/M cell cycle checkpoint. Proc Natl Acad Sci U S A. 1999;96(7):3706–3711. doi:10.1073/pnas.96.7.3706
118. Sun Y, Tang S, Xiao X. The effect of GADD45a on Furazolidone-induced S-Phase cell-cycle arrest in human hepatoma G2 cells. J Biochem mol Toxicol. 2015;29(10):489–495. doi:10.1002/jbt.21719
119. King RW, Jackson PK, Kirschner MW. Mitosis in transition. Cell. 1994;79(4):563–571. doi:10.1016/0092-8674(94)90542-8
120. Lei CY, Wang W, Zhu YT, Fang WY, Tan WL. The decrease of cyclin B2 expression inhibits invasion and metastasis of bladder cancer. Urol Oncol. 2016;34(5):237.e1–10. doi:10.1016/j.urolonc.2015.11.011
121. Chin AR, Fong MY, Somlo G, et al. Cross-kingdom inhibition of breast cancer growth by plant miR159. Cell Res. 2016;26(2):217–228. doi:10.1038/cr.2016.13
122. Yang J, Farmer LM, Agyekum AA, Hirschi KD. Detection of dietary plant-based small RNAs in animals. Cell Res. 2015;25(4):517–520. doi:10.1038/cr.2015.26
123. Zhang L, Hou D, Chen X, et al. Exogenous plant MIR168a specifically targets mammalian LDLRAP1: evidence of cross-kingdom regulation by microRNA. Cell Res. 2012;22(1):107–126. doi:10.1038/cr.2011.158
124. Zhao Z, Yu S, Li M, Gui X, Li P. Isolation of exosome-like nanoparticles and analysis of MicroRNAs derived from coconut water based on small RNA high-throughput sequencing. J Agric Food Chem. 2018;66(11):2749–2757. doi:10.1021/acs.jafc.7b05614
125. Xiao J, Feng S, Wang X, et al. Identification of exosome-like nanoparticle-derived microRNAs from 11 edible fruits and vegetables. PeerJ. 2018;6:e5186. doi:10.7717/peerj.5186
126. Zhang WJ, Li YY, Xiang ZH, et al. Emerging evidence on the effects of plant-derived microRNAs in colorectal cancer: a review. Food Funct. 2023;14(2):691–702. doi:10.1039/D2FO03477H
127. Minutolo A, Potestà M, Gismondi A, et al. Olea europaea small RNA with functional homology to human miR34a in cross-kingdom interaction of anti-tumoral response. Sci Rep. 2018;8(1):12413. doi:10.1038/s41598-018-30718-w
128. Zhu MZ, Xu HM, Liang YJ, et al. Edible exosome-like nanoparticles from portulaca oleracea L mitigate DSS-induced colitis via facilitating double-positive CD4(+)CD8(+)T cells expansion. J Nanobiotechnology. 2023;21(1):309. doi:10.1186/s12951-023-02065-0
129. Teng Y, Ren Y, Sayed M, et al. Plant-derived exosomal MicroRNAs shape the gut microbiota. Cell Host Microbe. 2018;24(5):637–52.e8. doi:10.1016/j.chom.2018.10.001
130. He W, Zheng S, Zhang H, et al. Plant-derived vesicle-like nanoparticles: clinical application exploration and challenges. Int J Nanomed. 2023;18:5671–5683. doi:10.2147/IJN.S428647
131. Sun JX, Xu JZ, An Y, et al. Future in precise surgery: fluorescence-guided surgery using EVs derived fluorescence contrast agent. J Control Release. 2023;353:832–841. doi:10.1016/j.jconrel.2022.12.013
132. Wang Q, Zhuang X, Mu J, et al. Delivery of therapeutic agents by nanoparticles made of grapefruit-derived lipids. Nat Commun. 2013;4:1867. doi:10.1038/ncomms2886
133. Dad HA, Gu TW, Zhu AQ, Huang LQ, Peng LH. Plant exosome-like nanovesicles: emerging therapeutics and drug delivery nanoplatforms. Mol Ther. 2021;29(1):13–31. doi:10.1016/j.ymthe.2020.11.030
134. Xiao Q, Zhao W, Wu C, et al. Lemon-derived extracellular vesicles nanodrugs enable to efficiently overcome cancer multidrug resistance by endocytosis-triggered energy dissipation and energy production reduction. Adv Sci. 2022;9(20):e2105274. doi:10.1002/advs.202105274
135. Umezu T, Takanashi M, Murakami Y, et al. Acerola exosome-like nanovesicles to systemically deliver nucleic acid medicine via oral administration. mol Ther Methods Clin Dev. 2021;21:199–208. doi:10.1016/j.omtm.2021.03.006
136. Fuhrmann G, Serio A, Mazo M, Nair R, Stevens MM. Active loading into extracellular vesicles significantly improves the cellular uptake and photodynamic effect of porphyrins. J Control Release. 2015;205:35–44. doi:10.1016/j.jconrel.2014.11.029
137. Zhang M, Xiao B, Wang H, et al. Edible ginger-derived nano-lipids loaded with doxorubicin as a novel drug-delivery approach for colon cancer therapy. Mol Ther. 2016;24(10):1783–1796. doi:10.1038/mt.2016.159
138. Haney MJ, Klyachko NL, Zhao Y, et al. Exosomes as drug delivery vehicles for Parkinson’s disease therapy. J Control Release. 2015;207:18–30. doi:10.1016/j.jconrel.2015.03.033
139. Kilasoniya A, Garaeva L, Shtam T, et al. Potential of plant exosome vesicles from grapefruit (Citrus × paradisi) and tomato (Solanum lycopersicum) juices as functional ingredients and targeted drug delivery vehicles. Antioxidants. 2023;12(4):943. doi:10.3390/antiox12040943
140. Kim MS, Haney MJ, Zhao Y, et al. Development of exosome-encapsulated paclitaxel to overcome MDR in cancer cells. Nanomedicine. 2016;12(3):655–664. doi:10.1016/j.nano.2015.10.012
141. Gelibter S, Marostica G, Mandelli A, et al. The impact of storage on extracellular vesicles: a systematic study. J Extracell Vesicles. 2022;11(2):e12162. doi:10.1002/jev2.12162
142. Xu X, Xu L, Wen C, Xia J, Zhang Y, Liang Y. Programming assembly of biomimetic exosomes: an emerging theranostic nanomedicine platform. Mater Today Bio. 2023;22:100760. doi:10.1016/j.mtbio.2023.100760
143. Zhang M, Wang X, Han MK, Collins JF, Merlin D. Oral administration of ginger-derived nanolipids loaded with siRNA as a novel approach for efficient siRNA drug delivery to treat ulcerative colitis. Nanomedicine. 2017;12(16):1927–1943. doi:10.2217/nnm-2017-0196
144. Ko K-W, Yoo Y-I, Kim JY, et al. Attenuation of tumor necrosis factor-α induced inflammation by umbilical cord-mesenchymal stem cell derived exosome-mimetic nanovesicles in endothelial cells. Tissue Eng Regen Med. 2020;17(2):155–163. doi:10.1007/s13770-019-00234-7
145. Itakura S, Shohji A, Amagai S, et al. Gene knockdown in HaCaT cells by small interfering RNAs entrapped in grapefruit-derived extracellular vesicles using a microfluidic device. Sci Rep. 2023;13(1):3102. doi:10.1038/s41598-023-30180-3
146. Li G, Wang M, Ding T, et al. cRGD enables rapid phagocytosis of liposomal vancomycin for intracellular bacterial clearance. J Control Release. 2022;344:202–213. doi:10.1016/j.jconrel.2022.02.030
147. Chen J, Pan J, Liu S, et al. Fruit-derived extracellular vesicles engineered structural droplet drugs for enhanced glioblastoma chemotherapy. Adv Mater. 2023;35:e2304187. doi:10.1002/adma.202304187
148. Niu W, Xiao Q, Wang X, et al. A biomimetic drug delivery system by integrating grapefruit extracellular vesicles and doxorubicin-loaded heparin-based nanoparticles for glioma therapy. Nano Lett. 2021;21(3):1484–1492. doi:10.1021/acs.nanolett.0c04753
149. Pillai JJ, Thulasidasan AKT, Anto RJ, Chithralekha DN, Narayanan A, Kumar GSV. Folic acid conjugated cross-linked acrylic polymer (FA-CLAP) hydrogel for site specific delivery of hydrophobic drugs to cancer cells. J Nanobiotechnol. 2014;12:25. doi:10.1186/1477-3155-12-25
150. Wang Q, Ren Y, Mu J, et al. Grapefruit-derived nanovectors use an activated leukocyte trafficking pathway to deliver therapeutic agents to inflammatory tumor sites. Cancer Res. 2015;75(12):2520–2529. doi:10.1158/0008-5472.CAN-14-3095
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms.php
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms.php
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
In vitro Development of Controlled-Release Nanoniosomes for Improved Delivery and Anticancer Activity of Letrozole for Breast Cancer Treatment

Ahmadi S, Seraj M, Chiani M, Hosseini S, Bazzazan S, Akbarzadeh I, Saffar S, Mostafavi E
International Journal of Nanomedicine 2022, 17:6233-6255
Published Date: 11 December 2022