")
Back to Journals » Cancer Management and Research » Volume 17
Genetic Navigation: A Narrative Review of XRCC1 Polymorphism Impact on Platinum-Based Chemotherapy Outcomes in NSCLC Patients
Authors Permatasari LI , Afifah NN , Ishmatullah MH, Intania R, Halimah E , Barliana MI
Received 17 October 2024
Accepted for publication 1 February 2025
Published 27 February 2025 Volume 2025:17 Pages 383—395
DOI https://doi.org/10.2147/CMAR.S501420
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Seema Singh
Lanny Indah Permatasari,1,2 Nadiya Nurul Afifah,1 Maryam Hasymia Ishmatullah,1 Ruri Intania,3 Eli Halimah,2 Melisa Intan Barliana1,4
1Department of Biological Pharmacy, Faculty of Pharmacy, Universitas Padjadjaran, Sumedang, Indonesia; 2Department of Pharmacology and Clinical Pharmacy, Faculty of Pharmacy, Universitas Padjadjaran, Sumedang, Indonesia; 3Department of Pulmonology and Respiratory Medicine, Dr. H.A. Rotinsulu Lung Hospital, Bandung, Indonesia; 4Center of Excellence for Pharmaceutical Care Innovation, Faculty of Pharmacy, Universitas Padjadjaran, Sumedang, Indonesia
Correspondence: Melisa Intan Barliana, Department of Biological Pharmacy, Faculty of Pharmacy, Universitas Padjadjaran, Jalan Raya Bandung-Sumedang KM 21, Jatinangor, Sumedang, West Java, 45363, Indonesia, Tel +62 22 842888888, Email [email protected]
Abstract: Non-small cell lung cancer (NSCLC) is the most common form of lung cancer, accounting for 85% of all cases, with a poor 5-year survival rate of less than 20%. The majority of NSCLC patients are diagnosed at an advanced stage, contributing to the low survival rate. Platinum-based chemotherapy, including cisplatin and carboplatin, remains the cornerstone of treatment for advanced NSCLC. However, DNA repair mechanisms often hinder treatment efficacy, notably Base Excision Repair (BER), mediated by the X-ray Repair Cross Complementing 1 (XRCC1) protein. This review aims to investigate the role of XRCC1 polymorphisms in platinum resistance, focusing on their impact on DNA repair efficiency. XRCC1’s involvement in the BER pathway is critical for repairing DNA damage caused by platinum agents, and polymorphisms in XRCC1 have been linked to altered repair capacity, influencing clinical outcomes and resistance to platinum-based chemotherapy in NSCLC patients.
Keywords: non-small cell lung cancer, DNA repair, base excision repair, XRCC1 polymorphism, platinum-based chemotherapy, response therapy, toxicity, survival rates, Platinum-based
Introduction
Non-small cell lung cancer (NSCLC) is the most common type of lung cancer worldwide, comprising approximately 85% of all reported cases.1,2 Globally, the 5-year survival rate for NSCLC is less than 20%.3 Most NSCLC patients are diagnosed at an advanced stage (70%), contributing to the low overall survival rate.4–6 Although local data from Indonesia are limited, this pattern reflects universal challenges in managing NSCLC across populations. Platinum-based chemotherapy, such as cisplatin and carboplatin, remains the mainstay of treatment for advanced NSCLC.7–9 Platinum compounds enter cancer cells via passive transport mediated by Copper Transporter 1 (CTR1)10 and direct passive diffusion across the cell membrane,11 then damage DNA. However, the efficacy of platinum agents is often hampered by resistance caused by DNA repair mechanisms such as Base Excision Repair (BER).12
The BER pathway, mediated by the XRCC1 protein, plays a critical role in repairing DNA damage and ensuring the survival of cancer cells.12 XRCC1 coordinates repair by recruiting DNA glycosylase, AP-1 enzyme (APE-1), Poly (ADP-ribose) polymerase 1 (PARP-1), DNA polymerase Beta (Pol-β), and DNA Ligase III (LIG-3) to the site of damage.12,13 Genetic variations in XRCC1 can affect protein functionality and DNA repair capacity, thereby affecting the efficacy of platinum chemotherapy. Polymorphisms on XRCC1, such as rs1799782 and rs25487, have been associated with different therapeutic responses, making them candidate biomarkers for predicting treatment outcomes.14,15
Based on data from PharmGKB, the level of evidence for the rs25487 variant is at level 2B (a combination of variant-drug with moderate evidence supporting the association) in the efficacy of platinum compounds,16 while the rs1799782 variant is at level 3.17 Global data shows that the frequency of the XRCC1 variant allele is around 20%,18,19 so it is anticipated that variant alleles with a similar frequency (>20%) are also present in the Indonesian population.20 This article aims to comprehensively examine the impact of XRCC1 gene polymorphism on the outcomes of platinum-based therapy in influencing DNA repair capacity. Variants of XRCC1 such as rs25487, rs1799782, and rs25489 are associated with diverse clinical outcomes, including chemotherapy response, toxicity profiles, and overall survival (OS) progression-free survival (PFS) rates. Consequently, the review further evaluates the clinical implications and future directions of NSCLC treatment, investigating the potential of precision medicine in disease management. This article aims to comprehensively review the role of XRCC1 polymorphisms in platinum resistance and their potential application in the precision medicine of NSCLC.
Platinum Therapy in NSCLC
Platinum-based chemotherapy is the frontline treatment option for NSCLC patients, particularly those with Epidermal Growth Factor Receptor (EGFR) and Anaplastic Lymphoma Kinase (ALK) negative tumor mutations.7–9 Platinum inhibits Deoxyribonucleic Acid (DNA) replication in the nucleus, providing a practical cytotoxic effect on cancer cells.21,22 At the molecular level, platinum agents form DNA adducts, either mono-adducts or intra- or interstrand adducts. These interactions involve covalent bonds that cause DNA lesions and cross-links between DNA strands, inhibiting cancer cell replication. Accumulation of DNA damage occurs in the G2/M phase, which ultimately triggers apoptosis of cancer cells.23 Platinum agents such as cisplatin and carboplatin show similar mechanisms of action. Cisplatin, with two amine groups (-NH3) and two chloride atoms (Cl−), and carboplatin, with a 1,1-cyclobutane dicarboxylate (CBDCA) ring, undergo hydrolysis reactions in the cell cytoplasm. This reaction replaces the Cl− atom or CBDCA ring with a water molecule (H2O), making platinum agents highly reactive. Platinum binds to the nitrogen atom (N) at position N7 in the DNA’s purine ring (adenine and guanine). This bond causes DNA damage, inhibits DNA synthesis, and ultimately triggers apoptosis.24,25
NSCLC cancer cells generally show sensitivity to platinum agents due to their tendency to have high proliferation rates and often inefficient DNA repair mechanisms.26 This makes them susceptible to platinum-induced DNA damage. However, the use of platinum agents faces several limitations. In addition to high levels of toxicity that can affect patients’ quality of life, platinum’s effectiveness often decreases over time.27 One of the main resistance mechanisms is the activation of DNA repair pathways, especially the BER pathway. The BER pathway allows cancer cells to repair DNA damage caused by platinum agents, thereby reducing the cytotoxic effects of these agents.
Base Excision Repair (BER) Pathway
The BER pathway is a type of DNA repair activated in response to minor problems such as base damage due to oxidation, deamination, or alkylation.28 DNA repair is essential for repairing errors during DNA replication or damage caused by radiation, chemicals, or metabolic byproducts. In normal cells, this DNA repair helps protect and maintain the integrity of the genetic material.29,30 However, in cancer cells, this pathway may provide a selective advantage by helping cancer cells repair DNA damage induced by platinum-based therapies, thereby increasing resistance to chemotherapy.
The BER process begins with the identification of DNA lesions by DNA glycosylases, such as Human 8-oxo guanine DNA glycosylase 1 (hOGG1), which removes the damaged nucleotide base, leaving behind an apurinic/apyrimidine site (AP site). Next, the APE-1 cleaves the DNA strand near the AP site, creating a gap with a free phosphate group at the 5’ end and a hydroxyl group at the 3’ end.31 Next, DNA POL-β fills the DNA gap created by APE-1. POL-β exhibits dual enzymatic activities: deoxyribose phosphodiesterase (dRPase) activity removes the residual sugar-phosphate group at the 5’ end of the DNA gap, and DNA polymerase activity adds a new nucleotide at the 3’ end, elongating the DNA strand and repairing the missing gap. After filling the gap, LIG-3 closes the DNA gap by forming a phosphodiester bond between the 3’-OH and 5’-phosphate ends. LIG-3 works in a complex with XRCC1. Throughout the BER process, XRCC1 acts as a scaffolding protein that regulates enzyme interactions and ensures that repair occurs quickly and accurately.31
XRCC1 Protein: Role, Mechanisms, and Interaction
XRCC1 is a protein encoded by the XRCC1 gene, functioning as a key organizer in the BER pathway.32,33 Deficiency or absence of this protein can result in inefficient DNA repair, accumulation of DNA damage, and increased risk of various diseases.34,35 XRCC1 works with PARP-1 to coordinate subsequent steps in the repair process, including the activities of POL-β and LIG-3. PARP-1 is an essential enzyme for detecting DNA damage, especially single-strand breaks (SSBs).33 Upon recognizing damage, PARP-1 initiates ADP-ribosylation, marking the site of damage with the PARP-1 chain. This serves as a recruitment signal for DNA repair proteins, including XRCC1. PARP-1 activation facilitates chromatin relaxation around the damage site, allowing XRCC1 and other enzymes to access the damaged DNA.36,37
In addition to aiding recruitment, XRCC1 prevents excessive PARP-1 activity, which can lead to DNA structural entrapment during repair and trigger additional damage.37 The interaction between XRCC1 and PARP-1 is critical for XRCC1 function during BER. Loss of PARP-1 function can impair XRCC1 recruitment, while XRCC1 deficiency can result in excessive PARP-1 activity, leading to an imbalance in the BER pathway.37 Early in the repair process, XRCC1 interacts with hOGG1 and APE-1 at unstructured junctional regions.38 This interaction ensures the transfer of DNA substrates from hOGG1 (which generates AP sites) to APE1 (which cleaves AP sites to continue the repair process). This coordination is critical for efficiency and avoids the formation of toxic intermediates that can cause additional DNA damage.39 XRCC1 also interacts with DNA by binding to its ends at SSB sites, stabilizing POL-β during the gap-filling step, and supporting LIG-3 in rejoining DNA strands. Its ability to physically interact with the enzymes involved (such as hOGG1, APE1, POL-β, and LIG-3) makes it a key component in regulating DNA repair (Figure 1).39–43 XRCC1 dysfunction or genetic mutations affecting this protein have been associated with resistance to platinum-based therapy in NSCLC.
 |
Figure 1 Mechanism of Platinum-Induced DNA Damage and BER Pathways in Cancer Cells. (A) Platinum (Pt) enters cells through passive diffusion and CTR1-mediated transport, causing DNA damage. Repair mechanisms, including base excision repair (BER) and nucleotide excision repair (NER), address single-strand breaks (SSBs) and double-strand breaks (DSBs). Persistent DNA damage leads to apoptosis. (B) BER Pathway Mechanism: DNA glycosylase removes damaged bases in BER, creating an AP site cleaved by APE-1. XRCC1 orchestrates repair by recruiting Polβ, LIG-3, and PARP-1, restoring DNA integrity after SSBs. |
XRCC1 Variants
As previously explained, the presence of the XRCC1 protein is the key to the success of the BER pathway DNA repair process. Deficiency or absence of this protein will disrupt the DNA repair process. The XRCC1 protein is encoded by the XRCC1 gene, which is located on chromosome 19q13.2. This gene consists of 17 exons that encode a protein consisting of 633 amino acids41 and has three main domains: the N-terminal domain (NTD) that interacts with Pol-β,42 the BRCT1 domain involved in interactions with PARP-1 and DNA,31,40 and the BRCT2 domain that interacts with LIG-3.43 In addition, the linker region facilitates interactions with proteins such as hOGG1 and APE-1.39
Studies on XRCC1 polymorphisms indicate that this genetic variation can affect DNA repair capacity. Previous studies have mainly focused on three SNP variants: rs25487, rs1799782, and rs25489. In the rs25487 variant, the polymorphism occurs in exon 10 at codon 399, which is located in the BRCT1 domain region, the binding site for PARP-1.41,44 The presence of polymorphism in this region produces a missense variant characterized by a base change from guanine (G) to adenine (A), leading to the substitution of the amino acid arginine (Arg) for glutamine (Gln).45,46 These changes may affect the interaction between XRCC1 and the PARP-1 enzyme. This interaction begins with transferring a covalent bond from the nicotinamide adenine dinucleotide (NAD+) molecule to the ADP-ribose polymer chain by the PARP enzyme. The bond formation involves the amine group (-NH3) of arginine and the polymer chain of ADP-ribose.47 Furthermore, this interaction induces PARP-1 activation at the DNA strand break site, facilitates damage recognition, and participates in the repair process.48 The rs25487 variant, which replaces arginine with glutamine, reduces the reactivity of the amine group (-NH3) on the glutamine residue to ADP-ribose. As a result, covalent bond formation is disrupted, dimming the interaction between XRCC1 and PARP-1. This reduced interaction hurts DNA repair capacity.37 Studies have reported similar findings confirming that decreased XRCC1-PAPR-1 interaction correlates with reduced DNA repair efficiency.37
Another polymorphism, namely the rs1799782 variant, occurs in exon six at codon 194,41 and also has a missense variation due to a base change from cytosine (C) to thymine (T); this leads to the replacement of arginine (Arg) with tryptophan (Trp) within the unstructured linker region. This substitution disrupts the interaction of XRCC1 with hOGG1, which is important for recruiting XRCC1 to sites of oxidative DNA damage (such as 8-oxoG).38 In addition, this variant has been reported to fail to recruit LIG-3 to the BER site, potentially interfering with subsequent steps of DNA repair.38 Besides rs1799782, another variation in the unstructured linker region is rs25489, located in exon nine at codon 280.41 This variant involves a base substitution from guanine (G) to adenine (A), resulting in arginine (Arg) being substituted for histidine (His).41 This region serves as the interaction site between XRCC1 and the APE-1 enzyme. Since XRCC1 has no enzymatic or catalytic activity, its interaction with other proteins, such as APE-1, relies entirely on direct physical interactions. This polymorphism affects the affinity of XRCC1 for APE-1, with amino acid changes, resulting in decreased XRCC1 binding affinity and the ability to interact with the APE-1 protein.49 Overall, all three polymorphism variations lead to a decrease or reduction in the interaction of XRCC1 with enzymes involved in DNA repair, which ultimately reduces the capacity of the BER pathway DNA repair and is associated with resistance and increased toxicity of platinum-based chemotherapy in NSCLC patients (Figure 2). In addition to the above variants, there is a variant of rs3213239 insertion/deletion type whose data is still very limited (Table 1).50
 |
Figure 2 XRCC1 coding region. XRCC1 interacts with several proteins participating in DNA repair processes, such as DNA polymerase beta (Polβ), Human 8-oxoguanine DNA glycosylase 1 (hOGG1), and AP endonuclease 1 (APE1). NTD: N-Terminal Domain; NLS: Nuclear Localization Signal; BRCT1/2: BRCA1 Carboxy-terminal domain. (A) Polymorphism of XRCC1 Arg399Gln. The amino acid change from Arg to Gln can disrupt DNA repair capacity. (B) Polymorphism of XRCC1 Arg194Trp and Arg280His in Linker 1 region. |
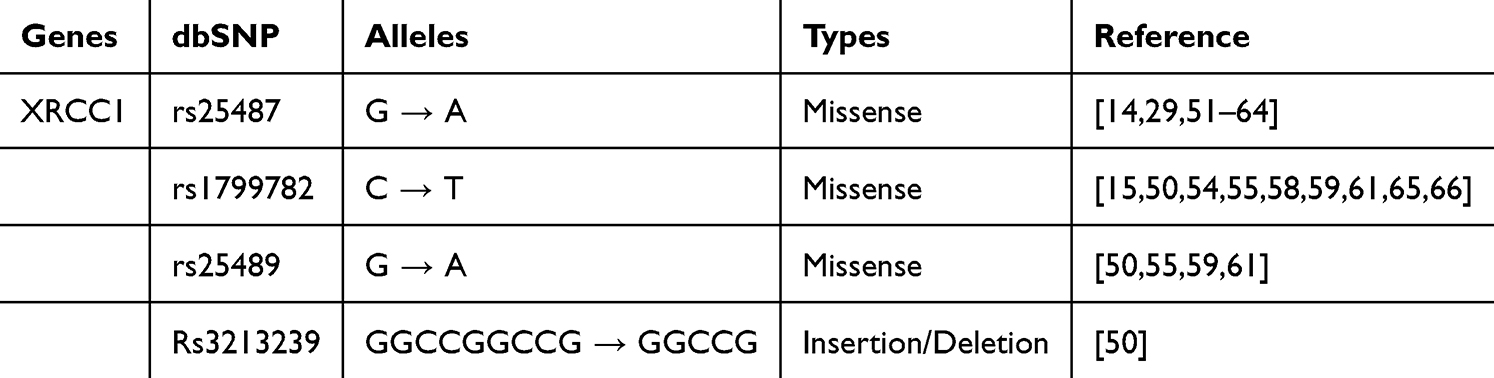 |
Table 1 Polymorphism on XRCC1 Genes |
XRCC1 Polymorphism to The Clinical Outcomes of Platinum-Based Chemotherapy in NSCLC
XRCC1 protein plays an essential role in enhancing DNA repair capacity in the BER pathway, which is responsible for repairing DNA damage caused by oxidative stress and alkylating agents, including platinum-based drugs such as cisplatin and carboplatin. Genetic variations in XRCC1, such as at positions Arg194Trp, Arg280His, or Arg399Gln, can alter the efficiency of the DNA repair process, affecting the accumulation of DNA damage in cells. This accumulation determines the effectiveness of therapy in inducing cancer cell death and can also affect toxicity to normal tissues.26 Therefore, understanding the impact of XRCC1 polymorphisms is essential for predicting clinical outcomes, including response rate to treatment, toxicity, overall survival, and progression-free survival in lung cancer patients (Table 2).
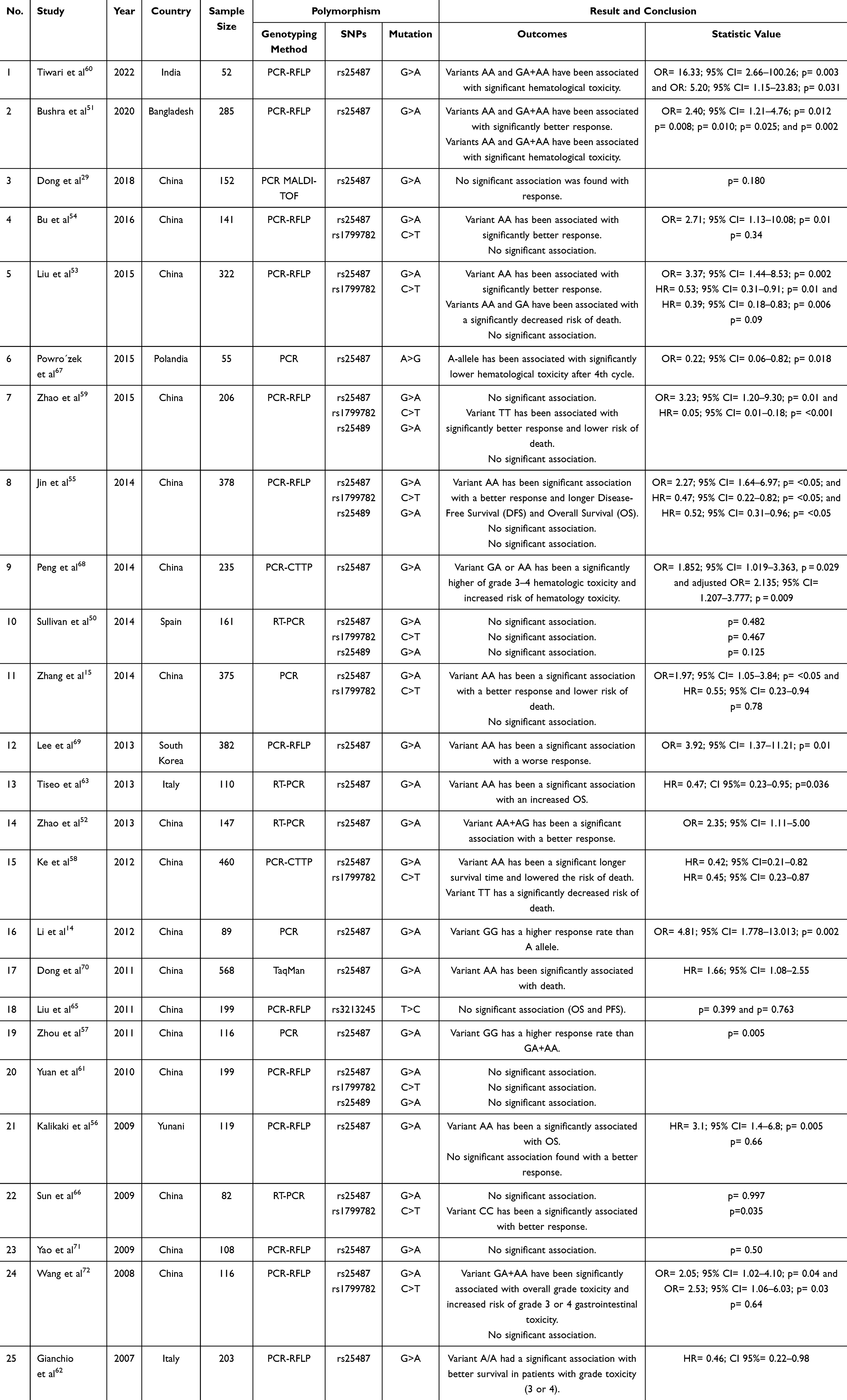 |
Table 2 The Study of XRCC1 Polymorphisms on Clinical Response to Platinum-Based Chemotherapy in NSCLC Patients |
Responses
XRCC1 gene polymorphisms have been extensively studied to determine their effect on platinum-based chemotherapy response in NSCLC patients across populations. For example, a study in Bangladesh showed that patients with the Arg/Gln (G/A) variant or Gln/Gln (A/A) variant had a better therapeutic response to platinum-based chemotherapy (OR = 2.40; 95% CI = 1.21–4.76; p = 0.012).51 Similar results were found in studies of South Korean and Chinese populations, where the Gln/Gln (A/A) variant was associated with a higher response rate.15,52–55,69 However, these results are not uniform across populations. Studies in Greek and Spanish populations did not find a significant association between the rs25487 variant and response to platinum-based chemotherapy.50,56 In contrast, other studies reported that the Gln/Gln variant decreased the response to platinum therapy in NSCLC patients.14,57 These differences in results are likely due to different genetic frequency distributions in each population and other factors such as sample size and genetic background.29,55,58
Meta-analysis and bioinformatics provide additional insights. A meta-analysis concluded that the rs25487 variant had no significant association with response to platinum-based chemotherapy.73 Bioinformatics analysis also showed that the rs25487 variant was predicted to be “benign” and did not significantly interfere with protein function. In contrast, the rs1799782 variant showed a low sorting intolerant from tolerant (SIFT) score (0.01–0.04), indicating a detrimental impact on XRCC1 protein function.73
In the rs1799782 variant, patients with the Trp/Trp (T/T) variant were significantly associated with a better response (OR = 3.23; 95% CI = 1.20–9.30; p = 0.01).59 A meta-analysis reported polymorphisms in the rs1799782 variant identified from 1145 among 2926 lung cancer patients, stating that patients with the Arg/Trp (C/T) and Trp/Trp (T/T) variants tended to have a good response (OR, 2.54; 95% CI, 1.95–3.31; p = 0.590).74 This was confirmed by seven studies including 1208 patients in the Asian population, where the Arg/Arg (C/C) genotype was associated with a worse response in those receiving platinum-based chemotherapy (OR = 0.55; 95% CI: 0.36–0.84; p = 0.013).74 Furthermore, the results of an analysis comparing 2228 NSCLC patients from nine studies reported a preference for the T allele variant in achieving a complete or partial response (OR = 1.38; 95% CI: 1.16–1.65; p < 0.001; I2 = 25.5%).75 A possible mechanism to explain how the rs1799782 variant can protect platinum resistance is the disruption of the interaction between XRCC1 and enzymes involved in repair.38
Toxicities
Platinum-based chemotherapy, such as cisplatin and carboplatin, is known to have high toxicity.76 Cisplatin often causes emesis, nephrotoxicity, and neurotoxicity. Meanwhile, carboplatin is more likely to cause hematological toxicity due to its myelosuppressive effects.77,78 A study of the XRCC1 rs25487 polymorphism reported that this variant could be used as a predictive factor for platinum-based chemotherapy toxicity, such as grade 3 and 4 anemia (p = 0.008), neutropenia (p = 0.010), thrombocytopenia (p = 0.025), along with gastrointestinal toxicity (p = 0.002).51 Another study reported that the Gln/Gln (A/A) variant carries a 16-fold higher risk (OR: 16.33, 95% CI: 2.66–100.26, P = 0.003).60 Adjusted logistic regression analysis gender, age, smoking status, disease stage, histopathology, Eastern Cooperative Oncology Group (ECOG) stage, and chemotherapy regimen showed a significantly higher incidence of grade 3–4 hematologic toxicity in those with Arg/Gln (G/A) or Gln/Gln (A/A) variants.60 Decreased DNA repair activity in XRCC1 variants is believed to be the primary cause of this increased toxicity. Platinum-based chemotherapy damages DNA, both in cancer and healthy cells. The inability to repair DNA damage in normal cells can exacerbate the side effects experienced by patients.79
Overall Survival (OS) and Progression-Free Survival (PFS)
DNA repair capacity in the BER pathway is critical in maintaining genome stability. Under normal conditions, low BER capacity increases the risk of cancer development due to the inability to repair DNA damage, resulting in cellular mutations that can drive tumor growth. Conversely, high BER capacity makes tumor cells more resistant to DNA-based therapies like platinum-based chemotherapy.80
Several studies have linked XRCC1 polymorphisms to survival in NSCLC patients.15,52,53,55,58 For example, the rs25487 polymorphism was reported to prolong OS and decrease the risk of death in patients with the Gln/Gln (A/A) or Arg/Gln (G/A) genotypes.53 These results are consistent with previous studies showing that the rs25487 polymorphism prolongs OS and reduces the risk of death in NSCLC patients.15,59 Similar results were found in the rs1799782 polymorphism, where patients with the Trp/Trp (T/T) variant showed a lower risk of death and longer OS.58
The difference in outcomes based on ethnicity is also highlighted. In the Asian population, Gln/Gln (A/A) variant in rs25487 was associated with better OS and PFS (Hazard Ratio (HR) = 0.65; 95% CI 0.43–0.98 and HR = 0.67; 95% CI 0.40–0.94), whereas in the Caucasian population, the same variant was associated with worse outcome (HR = 2.29; 95% CI 1.25–3.33).74 Furthermore, the Trp/Trp (T/T) variant in rs1799782 polymorphism contributed to longer OS in NSCLC patients (HR = 0.63, p = 0.013).75 Yuan et al (2010) conducted a haplotype study for the rs1799782, rs25487, and rs25489 variants in the Chinese population, the results of which showed that the Trp/Arg/Arg (T/G/G) haplotype has the potential to protect against death (OS) compared to the reference (Arg/Arg/Arg).61
Challenges and Limitations
Clinical implementation of XRCC1 polymorphism research results faces several challenges. Genetic testing and interpretation complexity is a significant obstacle, especially in diverse populations. Variability in the reporting of genetic data can lead to differences in interpretation, thus affecting treatment decisions.74,75 Further investigation in the broader population is needed to reduce bias and develop universally accepted clinical guidelines. In addition, integrating XRCC1 genotyping into clinical practice requires training for healthcare workers, especially in oncology, to ensure accurate and meaningful genetic interpretation. Logistical factors, such as the availability of sophisticated genetic testing facilities, also pose challenges to widespread adoption.81
Regarding the cost perspective, the expenses associated with genetic testing pose a significant barrier. Therefore, it is essential to consider the financial implications for healthcare institutions and patients, as genetic testing may not always be covered by insurance or accessible to individuals with limited reports. Addressing the cost-effectiveness of XRCC1 genotyping is crucial to ensuring equitable access to this precision method. Furthermore, ethical considerations are necessary because patient consent, privacy, and the responsible use of genetic information are essential. Clear protocols for obtaining informed consent and safeguarding patient data must be established to uphold ethical standards.
Conclusion
In conclusion, this review shows the significance of XRCC1 polymorphism in influencing treatment outcomes among NSCLC patients receiving platinum-based chemotherapy. Understanding genetic variations in XRCC1 offers potential for precision treatment methods. However, challenges related to genetic testing complexity, interpretation, cost, and ethical considerations must be addressed to maximize the clinical utility of XRCC1 genotyping. The potential of targeted therapies designed explicitly for XRCC1 variants presents an exciting avenue for improving treatment efficacy and outcomes in NSCLC patients.
Acknowledgments
We would like to express our sincere gratitude to all individuals who have contributed to the completion of this manuscript.
Funding
This work was funded by the Ministry of Higher Education, Science and Technology, Republic of Indonesia, under a Fundamental Research scheme grant for MIB. Open access funding supported by Universitas Padjadjaran.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Lu T, Yang X, Huang Y, et al. Trends in the incidence, treatment, and survival of patients with lung cancer in the last four decades. Cancer Manag Res. 2019;11:943–953. doi:10.2147/CMAR.S187317
2. WHO. Lung cancer. World Health Organization. Available from: https://www.who.int/news-room/fact-sheets/detail/lung-cancer.
3. Siegel RL, Miller KD, Jemal A. Cancer statistics, 2018. CA Cancer J Clin. 2018;68(1):7–30. doi:10.3322/caac.21442
4. Zhang Y, Simoff MJ, Ost D, et al. Understanding the patient journey to diagnosis of lung cancer. BMC Cancer. 2021;21(1):402. doi:10.1186/s12885-021-08067-1
5. Jeon DS, Kim HC, Kim SH, et al. Five-year overall survival and prognostic factors in patients with lung cancer: results from the Korean association of lung cancer registry (KALC-R) 2015. Cancer Res Treat. 2023;55(1):103–111. doi:10.4143/crt.2022.264
6. Goldstraw P, Chansky K, Crowley J, et al. The IASLC lung cancer staging project: proposals for revision of the TNM stage groupings in the forthcoming (eighth) edition of the TNM classification for lung cancer. J Thorac Oncol. 2016;11(1):39–51. doi:10.1016/j.jtho.2015.09.009
7. Kemenkes RI. Pedoman nasional pelayanan kedokteran tata laksana kanker paru. Keputusan Menteri Kesehat Republik Indones Nomor HK 01 07/MENKES/1438/2023. 2023;1–90.
8. Planchard D, Popat ST, Kerr K, et al. Metastatic non-small cell lung cancer: ESMO clinical practice guidelines for diagnosis, treatment and follow-up. Ann Oncol off J Eur Soc Med Oncol. 2018;29:iv192–iv237.
9. Ettinger DS, Wood DE, Aisner DL, et al. Non-small cell lung cancer, version 3.2022. JNCCN J Natl Compr Cancer Netw. 2022;20:497–530.
10. Song I-S, Savaraj N, Siddik ZH, et al. Role of human copper transporter Ctr1 in the transport of platinum-based antitumor agents in cisplatin-sensitive and cisplatin-resistant cells. mol Cancer Ther. 2004;3(12):1543–1549. doi:10.1158/1535-7163.1543.3.12
11. Gately DP, Howell SB. Cellular accumulation of the anticancer agent cisplatin: a review. Br J Cancer. 1993;67(6):1171–1176. doi:10.1038/bjc.1993.221
12. Slyskova J, Sabatella M, Ribeiro-Silva C, et al. Base and nucleotide excision repair facilitate resolution of platinum drugs-induced transcription blockage. Nucleic Acids Res. 2018;46(18):9537–9549. doi:10.1093/nar/gky764
13. Cuneo MJ, London RE. Oxidation state of the XRCC1 N-terminal domain regulates DNA polymerase β binding affinity. Proc Natl Acad Sci U S A. 2010;107(15):6805–6810. doi:10.1073/pnas.0914077107
14. Li D, Zhou Q, Liu Y, Yang Y, Li Q. DNA repair gene polymorphism associated with sensitivity of lung cancer to therapy. Med Oncol. 2012;29:1622–1628. doi:10.1007/s12032-011-0033-7
15. Zhang L, Ma W, Li Y, Wu J, Shi GY. Pharmacogenetics of DNA repair gene polymorphisms in non-small-cell lung carcinoma patients on platinum-based chemotherapy. Genet Mol Res. 2014;13:228–236. doi:10.4238/2014.January.14.2
16. Huang M-Y, Huang M-L, Chen M-J, et al. Multiple genetic polymorphisms in the prediction of clinical outcome of metastatic colorectal cancer patients treated with first-line FOLFOX-4 chemotherapy. Pharmacogenet Genomics. 2011;21(1):18–25. doi:10.1097/FPC.0b013e3283415124
17. Liu L, Wu J, Zhong R, et al. Multi-loci analysis reveals the importance of genetic variations in sensitivity of platinum-based chemotherapy in non-small-cell lung cancer. Mol Carcinog. 2013;52(12):923–931. doi:10.1002/mc.21942
18. PharmGKB. rs1799782. PharmGKB. 2024. Available from: https://www.pharmgkb.org/variant/PA166155370.
19. PharmGKB. rs25487. PharmGKB. 2024. Available from: https://www.pharmgkb.org/variant/PA166155346.
20. Wright AF. Genetic variation: polymorphisms and mutations. Encycl Life Sci. 2005;1–10. doi:10.1038/npg.els.0005005
21. Corte-Rodríguez M, Espina M, Sierra LM, et al. Quantitative evaluation of cellular uptake, DNA incorporation and adduct formation in cisplatin sensitive and resistant cell lines: comparison of different Pt-containing drugs. Biochem Pharmacol. 2015;98(1):69–77. doi:10.1016/j.bcp.2015.08.112
22. Dasari S, Tchounwou PB. Cisplatin in cancer therapy: molecular mechanisms of action. Eur J Pharmacol. 2014;740:364–378. doi:10.1016/j.ejphar.2014.07.025
23. Rottenberg S, Disler C, Perego P. The rediscovery of platinum-based cancer therapy. Nat Rev Cancer. 2021;21:37–50. doi:10.1038/s41568-020-00308-y
24. Bhat A, Verma S, Chander G, et al. Cisplatin-based combination therapy for cancer. J Cancer Res Ther. 2023;19.
25. Brown A, Kumar S, Tchounwou PB. Cisplatin-based chemotherapy of human cancers. J Cancer Sci Ther. 2019;11.
26. Kim ES, Lee JJ, He G, et al. Tissue platinum concentration and tumor response in non-small-cell lung cancer. J Clin Oncol. 2012;30(27):3345–3352. doi:10.1200/JCO.2011.40.8120
27. Eskandari A, Kundu A, Ghosh S, Suntharalingam K. A triangular platinum(II) multinuclear complex with cytotoxicity towards breast cancer stem cells. Angew Chem Int Ed Engl. 2019;58:12059–12064. doi:10.1002/anie.201905389
28. Dexheimer TS. DNA Repair Pathways and Mechanisms BT - DNA Repair of Cancer Stem Cells. Mathews LA, Cabarcas SM, Hurt EM eds. 19–32: Springer Netherlands. 2013. doi:10.1007/978-94-007-4590-2_2
29. Dong J, Wang X, Cui JW. Association of base excision repair gene polymorphisms with response to chemotherapy of advanced non small-cell lung cancer in a Chinese population. J Clin Oncol. 2018;36(15_suppl):e20504–e20504. doi:10.1200/JCO.2018.36.15_suppl.e20504
30. Cleaver JE. Xeroderma pigmentosum: a human disease in which an initial stage of DNA repair is defective. Proc Natl Acad Sci U S A. 1969;63(2):428–435. doi:10.1073/pnas.63.2.428
31. Polo LM, Xu Y, Hornyak P, et al. Efficient single-strand break repair requires binding to both poly(ADP-Ribose) and DNA by the central BRCT domain of XRCC1. Cell Rep. 2019;26(3):573–581.e5. doi:10.1016/j.celrep.2018.12.082
32. Campalans A, Marsin S, Nakabeppu Y, et al. XRCC1 interactions with multiple DNA glycosylases: a model for its recruitment to base excision repair. DNA Repair. 2005;4(7):826–835. doi:10.1016/j.dnarep.2005.04.014
33. Thompson LH, Brookman KW, Jones NJ, Allen SA, Carrano AV. Molecular cloning of the human XRCC1 gene, which corrects defective DNA strand break repair and sister chromatid exchange. mol Cell Biol. 1990;10(12):6160–6171. doi:10.1128/mcb.10.12.6160-6171.1990
34. Hoeijmakers HJ. DNA damage, aging, and cancer. N Engl J Med. 2025;361(15):1475–1485. doi:10.1056/NEJMra0804615
35. Hoeijmakers JH. Genome maintenance mechanisms for preventing cancer. Nature. 2001;411(6835):366–374. doi:10.1038/35077232
36. Polo SE, Kaidi A, Baskcomb L, Galanty Y, Jackson SP. Regulation of DNA‐damage responses and cell‐cycle progression by the chromatin remodelling factor CHD4. EMBO J. 2010;29(18):3130–3139. doi:10.1038/emboj.2010.188
37. Demin AA, Hirota K, Tsuda M, et al. XRCC1 prevents toxic PARP1 trapping during DNA base excision repair. Mol Cell. 2021;81(14):3018–3030.e5. doi:10.1016/j.molcel.2021.05.009
38. Campalans A, Moritz E, Kortulewski T, et al. Interaction with OGG1 is required for efficient recruitment of XRCC1 to base excision repair and maintenance of genetic stability after exposure to oxidative stress. mol Cell Biol. 2015;35(9):1648–1658. doi:10.1128/MCB.00134-15
39. Marsin S, Vidal AE, Sossou M, et al. Role of XRCC1 in the coordination and stimulation of oxidative DNA damage repair initiated by the DNA glycosylase Hogg1. J Biol Chem. 2003;278(45):44068–44074. doi:10.1074/jbc.M306160200
40. Masson M, Niedergang C, Schreiber V, et al. XRCC1 is specifically associated with poly(ADP-Ribose) polymerase and negatively regulates its activity following DNA damage. mol Cell Biol. 1998;18(6):3563–3571. doi:10.1128/MCB.18.6.3563
41. Norjmaa B, Tulgaa K, Saitoh T. Base excision repair pathway and polymorphisms of XRCC1 gene human genetic factor of disease susceptibility view project multi drug resistant organisms view project. 2016.
42. Marintchev A, Marintchev A, Mullen MA, et al. Solution structure of the single-strand break repair protein XRCC1 N-terminal domain. Nat Struct Biol. 1999;6(9):884–893. doi:10.1038/12347
43. Nash RA, Caldecott KW, Barnes DE, Lindahl T. XRCC1 protein interacts with one of two distinct forms of DNA ligase III. Biochemistry. 1997;36(17):5207–5211. doi:10.1021/bi962281m
44. Seibold P, Schmezer P, Behrens S, et al. A polymorphism in the base excision repair gene PARP2 is associated with differential prognosis by chemotherapy among postmenopausal breast cancer patients. BMC Cancer. 2015;15(1):978. doi:10.1186/s12885-015-1957-7
45. Kang CH, Jang BG, Kim D-W, et al. The prognostic significance of ERCC1, BRCA1, XRCC1, and βIII-tubulin expression in patients with non-small cell lung cancer treated by platinum- and taxane-based neoadjuvant chemotherapy and surgical resection. Lung Cancer. 2010;68(3):478–483. doi:10.1016/j.lungcan.2009.07.004
46. Lunn RM, Langlois RG, Hsieh LL, Thompson CL, Bell DA. XRCC1 polymorphisms: effects on aflatoxin B1-DNA adducts and glycophorin A variant frequency. Cancer Res. 1999;59(11):2557–2561.
47. Reber JM, Božić-Petković J, Lippmann M, et al. PARP1 and XRCC1 exhibit a reciprocal relationship in genotoxic stress response. Cell Biol Toxicol. 2023;39(1):345–364. doi:10.1007/s10565-022-09739-9
48. El-Khamisy SF, Masutani M, Suzuki H, Caldecott KW. A requirement for PARP-1 for the assembly or stability of XRCC1 nuclear foci at sites of oxidative DNA damage. Nucleic Acids Res. 2003;31(19):5526–5533. doi:10.1093/nar/gkg761
49. Vidal AE, Boiteux S, Hickson ID, Radicella JP. XRCC1 coordinates the initial and late stages of DNA abasic site repair through protein-protein interactions. EMBO J. 2001;20(22):6530–6539. doi:10.1093/emboj/20.22.6530
50. Sullivan I, Salazar J, Majem M, et al. Pharmacogenetics of the DNA repair pathways in advanced non-small cell lung cancer patients treated with platinum-based chemotherapy. Cancer Lett. 2014;353(2):160–166. doi:10.1016/j.canlet.2014.07.023
51. Bushra MU, Rivu SF, Sifat AE, et al. Genetic polymorphisms of GSTP1, XRCC1, XPC and ERCC1: prediction of clinical outcome of platinum-based chemotherapy in advanced non-small cell lung cancer patients of Bangladesh. Mol Biol Rep. 2020;47(9):7073–7082. doi:10.1007/s11033-020-05771-2
52. Zhao W, Hu L, Xu J, et al. Polymorphisms in the base excision repair pathway modulate prognosis of platinum-based chemotherapy in advanced non-small cell lung cancer. Cancer Chemother Pharmacol. 2013;71(5):1287–1295. doi:10.1007/s00280-013-2127-8
53. Liu JY, Liu QM, Li LR. Association of GSTP1 and XRCC1 gene polymorphisms with clinical outcomes of patients with advanced non-small cell lung cancer. Genet Mol Res. 2015;14(3):10331–10337. doi:10.4238/2015.August.28.19
54. Bu L, Zhang LB, Mao X, Wang P. GSTP1 Ile105Val and XRCC1 Arg399Gln gene polymorphisms contribute to the clinical outcome of patients with advanced non-small cell lung cancer. Genet Mol Res. 2016;15(2):1–9. doi:10.4238/gmr.15027611
55. Jin ZY, Zhao XT, Zhang LN, et al. Effects of polymorphisms in the XRCC1, XRCC3, and XPG genes on clinical outcomes of platinum-based chemotherapy for treatment of non-small cell lung cancer. Genet Mol Res. 2014;13(3):7617–7625. doi:10.4238/2014.March.31.13
56. Kalikaki A, Kanaki M, Vassalou H, et al. DNA repair gene polymorphisms predict favorable clinical outcome in advanced non-small-cell lung cancer. Clin Lung Cancer. 2009;10(2):118–123. doi:10.3816/CLC.2009.n.015
57. zhou F, Yu Z, Jiang T, et al. Genetic polymorphisms of GSTP1 and XRCC1: prediction of clinical outcome of platinum-based chemotherapy in advanced non-small cell lung cancer (NSCLC) patients. Swiss Med Wkly. 2011;141:w13275. doi:10.4414/smw.2011.13275
58. Ke HG, Li J, Shen Y, et al. Prognostic significance of GSTP1, XRCC1 and XRCC3 polymorphisms in non-small cell lung cancer patients. Asian Pacific J Cancer Prev. 2012;13(9):4413–4416. doi:10.7314/APJCP.2012.13.9.4413
59. Zhao R, Chen G. Role of GSTP1 Ile105Val and XRCC1 Arg194Trp, Arg280His and Arg399Gln gene polymorphisms in the clinical outcome of advanced non-small cell lung cancer. Int J Clin Exp Pathol. 2015;8(11):14909–14916.
60. Tiwari S, Vijayaraghavalu S, Siddiqui MA, Al-Khedhairy AA, Kumar M. Clinical response of carboplatin-based chemotherapy and its association to genetic polymorphism in lung cancer patients from North India – a clinical pharmacogenomics study. J Cancer Res Ther. 2022;18(1):109–118. doi:10.4103/jcrt.JCRT_925_20
61. Yuan P, Liu L, Wu C, et al. No association between XRCC1 polymorphisms and survival in non-small-cell lung cancer patients treated with platinum-based chemotherapy. Cancer Biol Ther. 2010;10(9):854–859. doi:10.4161/cbt.10.9.13238
62. Giachino DF, Ghio P, Regazzoni S, et al. Prospective assessment of XPD Lys751Gln and XRCC1 Arg399Gln single nucleotide polymorphisms in lung cancer. Clin Cancer Res. 2007;13(10):2876–2881. doi:10.1158/1078-0432.CCR-06-2543
63. Tiseo M, Bordi P, Bortesi B, et al. ERCC1/BRCA1 expression and gene polymorphisms as prognostic and predictive factors in advanced NSCLC treated with or without cisplatin. Br J Cancer. 2013;108(8):1695–1703. doi:10.1038/bjc.2013.127
64. Chen J, Zhao Q, Shi G, Wang L. XRCC1 Arg399Gln and clinical outcome of platinum-based treatment for advanced non-small cell lung cancer: a meta-analysis in 17 studies. J Zhejiang Univ Sci B. 2012;13(11):875–883. doi:10.1631/jzus.B1200083
65. Liu L, Yuan P, Wu C, et al. Assessment of XPD Lys751Gln and XRCC1 T–77C polymorphisms in advanced non-small-cell lung cancer patients treated with platinum-based chemotherapy. Lung Cancer. 2011;73(1):110–115. doi:10.1016/j.lungcan.2010.11.004
66. Sun X, Li F, Sun N, et al. Polymorphisms in XRCC1 and XPG and response to platinum-based chemotherapy in advanced non-small cell lung cancer patients. Lung Cancer. 2009;65(2):230–236. doi:10.1016/j.lungcan.2008.11.014
67. Powrózek T, Mlak R, Krawczyk P, et al. The relationship between polymorphisms of genes regulating DNA repair or cell division and the toxicity of platinum and vinorelbine chemotherapy in advanced NSCLC patients. Clin Transl Oncol. 2016;18(2):125–131. doi:10.1007/s12094-015-1343-6
68. Peng Y, Li Z, Zhang S, et al. Association of DNA base excision repair genes (OGG1, APE1 and XRCC1) polymorphisms with outcome to platinum-based chemotherapy in advanced nonsmall-cell lung cancer patients. Int J Cancer. 2014;135(11):2687–2696. doi:10.1002/ijc.28892
69. Lee SY, Kang H-G, Yoo SS, et al. Polymorphisms in DNA repair and apoptosis-related genes and clinical outcomes of patients with non-small cell lung cancer treated with first-line paclitaxel-cisplatin chemotherapy. Lung Cancer. 2013;82(2):330–339. doi:10.1016/j.lungcan.2013.07.024
70. Dong J, Hu Z, Shu Y, et al. Potentially functional polymorphisms in DNA repair genes and non-small-cell lung cancer survival: a pathway-based analysis. Mol Carcinog. 2012;51(7):546–552. doi:10.1002/mc.20819
71. Yao CY, Huang XE, Li C, et al. Lack of influence of XRCC1 and XPD gene polymorphisms on outcome of platinum-based chemotherapy for advanced non small cell lung cancers. Asian Pacific J Cancer Prev. 2009;10:859–864.
72. Wang Z, Xu B, Lin D, et al. XRCC1 polymorphisms and severe toxicity in lung cancer patients treated with cisplatin-based chemotherapy in Chinese population. Lung Cancer. 2008;62(1):99–104. doi:10.1016/j.lungcan.2008.02.019
73. Sito H, Sharzehan MAK, Islam MA, Tan SC. Genetic variants associated with response to platinum-based chemotherapy in non-small cell lung cancer patients: a field synopsis and meta‐analysis. Br J Biomed Sci. 2024;81:1–12. doi:10.3389/bjbs.2024.11835
74. Li D-J, Xiao D. Association between the XRCC1 polymorphisms and clinical outcomes of advanced NSCLC treated with platinum-based chemotherapy: a meta-analysis based on the PRISMA statement. BMC Cancer. 2017;17(1):501. doi:10.1186/s12885-017-3487-y
75. Zhang N, Ouyang Y, Chang J, Liu P, Tian X, Yu J. Pharmacogenetic association between XRCC1 polymorphisms and response to platinum-based chemotherapy in Asian patients with NSCLC: a meta-analysis. BioMed Res Int. 2020;2020:3520764.
76. Gao G, Jiang J, Liang X, et al. A meta-analysis of platinum plus gemcitabine or vinorelbine in the treatment of advanced non-small-cell lung cancer. Lung Cancer. 2009;65(3):339–344. doi:10.1016/j.lungcan.2008.11.019
77. Griesinger F, Korol EE, Kayaniyil S, et al. Efficacy and safety of first-line carboplatin-versus cisplatin-based chemotherapy for non-small cell lung cancer: a meta-analysis. Lung Cancer. 2019;135:196–204. doi:10.1016/j.lungcan.2019.07.010
78. Ho GY, Woodward N, Coward JIG. Cisplatin versus carboplatin: comparative review of therapeutic management in solid malignancies. Crit Rev Oncol Hematol. 2016;102:37–46. doi:10.1016/j.critrevonc.2016.03.014
79. van den Boogaard WMC, Komninos DSJ, Vermeij WP. Chemotherapy side-effects: not all DNA damage is equal. Cancers. 2022;14(3):1–27. doi:10.3390/cancers14030627
80. Rosell R, Lord RVN, Taron M, Reguart N. DNA repair and cisplatin resistance in non-small-cell lung cancer. Lung Cancer. 2002;38(3):217–227. doi:10.1016/S0169-5002(02)00224-6
81. Bangash H, Kullo IJ. Implementation science to increase adoption of genomic medicine: an urgent need. J Pers Med. 2021;11:1–3.
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms.php
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms.php
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.