")
Back to Journals » Drug Design, Development and Therapy » Volume 19
Gut-Brain Axis-Based Polygala Tenuifolia and Magnolia Officinalis Improve D-gal-Induced Cognitive Impairment in Mice Through cAMP and NF-κB Signaling Pathways
Authors Yang D, Li W, Chen Q , Liu S, Peng C, Deng F, Meng Y, Yang Y, Yan P, Ao H, Huang L
Received 3 December 2024
Accepted for publication 1 March 2025
Published 13 March 2025 Volume 2025:19 Pages 1869—1894
DOI https://doi.org/10.2147/DDDT.S506545
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Tuo Deng
Dan Yang,1 Wen Li,1 Qiuping Chen,1 Si Liu,1 Chengjie Peng,1 Fengcheng Deng,1 Yingqi Meng,1 Yang Yang,1 Ping Yan,1 Hui Ao,1,2 Lihua Huang1,2
1School of Pharmacy, Chengdu University of Traditional Chinese Medicine, Chengdu, 611137, People’s Republic of China; 2State Key Laboratory of Southwestern Chinese Medicine Resources, Chengdu, 611137, People’s Republic of China
Correspondence: Hui Ao; Lihua Huang, School of Pharmacy, Chengdu University of Traditional Chinese Medicine, Chengdu, 611137, People’s Republic of China, Email [email protected]; [email protected]
Purpose: Polygala tenuifolia Willd. (PT) is commonly used to address cognitive impairment (CI), while Magnolia officinalis Rehd. et Wils (MO) is often prescribed for gastrointestinal issues as well as CI. This study seeks to explore the impacts and mechanisms behind the combined therapy of PT and MO (PM) in treating CI, based on the concept of the gut-brain axis.
Methods: The characteristic components of PT, MO, and PM were identified using ultra-high performance liquid chromatography-tandem triple quadrupole mass Spectrometry (UPLC-MS/MS). A mouse model was established by D-gal induction, and the effects of PT, MO, and PM on CI were evaluated through behavioral tests, pathological staining, and Enzyme-Linked Immunosorbent Assay (ELISA). Subsequently, network pharmacology was used to analyze the potential mechanisms by which PM improves CI, followed by validation through Western blotting (WB), traditional Chinese medicine (TEM), Immunofluorescence (IF), and 16S rRNA.
Results: PT, MO, and PM can each alleviate cognitive decline and neuropathological damage in D-gal mice to varying degrees, reduce the expression of pro-inflammatory factors (TNF-α, IL-1β, IL-6, IFN-γ, LPS) in serum or hippocampal tissue, and increase SOD and GSH levels. Network pharmacology analysis and molecular experiments confirmed that PM upregulates the expression of tight junction s (TJs), enhances the expression of proteins in the cAMP pathway, and inhibits p-NF-κB-p65 expression. PM reverses D-gal-induced gut microbiota dysbiosis, increases the abundance of SCFA-producing bacteria, and decreases the abundance of LPS-producing bacteria.
Conclusion: PM alleviates CI by reducing inflammation and oxidative stress, protecting the blood-brain barrier (BBB) and intestinal barrier, inhibiting the NF-κB pathway, activating the cAMP pathway, and regulating gut microbiota.
Keywords: polygala tenuifolia, magnolia officinalis, cognitive impairment, cAMP Signaling Pathway, NF-κB Signaling Pathway, gut microbiota
Graphical Abstract:

Introduction
CI denotes one or several deficits in a person’s cognitive abilities, which may include memory, attention, executive functioning, language, and visuospatial skills.1–3 The range of severity includes mild cognitive impairment, in addition to moderate and severe forms of dementia.4,5 Currently, more than 55 million individuals worldwide are affected by dementia, with around 10 million new instances recognized annually.6 The effects of dementia are significant for both the individuals diagnosed and their families, posing considerable challenges to the sustainability of elderly care and healthcare services.7
CI can arise from a multitude of factors, notably including aging, Alzheimer’s disease (AD), and depression, with AD being one of the most prevalent forms of CI.8–12 Recent scholarly research indicates that an imbalance in gut microbiota, referred to as dysbiosis, could play a role in the onset of CI.13 Animal studies utilizing mouse models have shown that when gut microbiota from Alzheimer’s mice are transplanted or when probiotics are administered, there are observable changes in amyloid beta (Aβ) pathology.14,15 This finding reinforces the idea that gut microbiota might significantly influence the fundamental processes associated with AD. In addition, numerous case-control studies have noted changes in the gut microbiota composition in people diagnosed with mild cognitive impairment and AD.16 The gut microbiome serves as an essential component of the gut-brain communication pathway, which is a sophisticated network facilitating interactions between the brain and the digestive system.17 Through various pathways and mechanisms, the gut microbiota communicates with the brain, which can significantly influence the overall health of the host.18–20 Recent studies have highlighted the significance of the gut-brain connection in controlling various physiological and pathological mechanisms, including mood regulation, cognitive functionality, and inflammatory responses.21 The functioning of this axis is influenced by the gut microbiota through the production of particular metabolites, including SCFAs, secondary bile acids, and tryptophan-derived metabolites. These metabolites may, in turn, have profound effects on cognitive functions and emotional states.22 These results collectively indicate a significant connection between gut microbiota and cognitive impairment, suggesting that gut microbiota could be pivotal in the development and advancement of cognitive impairment, potentially facilitated by the gut-brain axis mechanisms.
At present, there is a shortage of effective treatment options for CI, highlighting the importance of early diagnosis and timely intervention to enhance prognosis and overall quality of life.23–25 Although there have been numerous studies on drugs for treating cognitive impairment, commonly used drugs such as memantine and donepezil have shown limited efficacy in improving cognitive abilities.26,27 Traditional Chinese medicine offers multiple target effects and has few side effects,28 which makes it a frequently utilized method for both preventing and treating cognitive impairment, including the use of ginkgo biloba extract29 and huperzine A.30 PT refers to the dried root of Polygala tenuifolia Willd. From the Polygalaceae family, and MO refers to the dried root bark and branch bark of Magnolia Officinalis Rehd.et Wils from the Magnoliaceae family. Both PT and MO are initially recorded in “Shennong’s Materia Medica”, where PT is noted for its abilities to soothe, enhance cognition, relieve phlegm, and act as an antidepressant. This indicates that PT has a positive impact on cognitive function. Contemporary research has shown that PT provides protective benefits for the central nervous system and has the potential to ease symptoms including memory deficits, insomnia, and neurasthenia.31 The component of PT, Tenuifolin (TEN),29 has demonstrated a significant improvement in CI and memory learning, suggesting its potential utility as a therapeutic agent in the treatment of CI.32,33 TEN influences various biological processes, such as hindering the accumulation of Aβ related to neurodegenerative damage, decreasing levels of acetylcholine, lessening neuroinflammatory responses, mitigating oxidative harm to cells, and blocking the apoptosis of neurons.34–37 MO is known to dry dampness, eliminate phlegm, and alleviate fullness, which is why it is frequently used in traditional Chinese medicine to address gastrointestinal disorders.38 In traditional Chinese medicine, MO has also been used to regulate qi and blood as well as dredge the meridians. In Traditional Chinese Medicine theory, smooth flow of qi and blood and harmonious meridians are closely associated with the cognitive state of the brain. Recent research has shown that MO has a substantial effect on the nervous system, displaying properties such as anti-epileptic, anti-depressant, anti-dementia, and anti-cerebral ischemia activities. The primary active ingredients, magnolol and honokiol, can easily penetrate the BBB.39–41 Research has shown that MO can improve cognitive impairments in mice with AD by stimulating mitochondrial SIRT3,42 suggesting its promise as a versatile treatment option for neurodegenerative disorders.
The management of CI using PM has been practiced for many years. An example is the Zhu Li Decoction, which was recorded by the esteemed Tang Dynasty doctor Sun Simiao in “Qianjin Yaofang” and is applied to treat signs like confusion and speech difficulties.43,44 In a similar vein, the Wuxie Pill mentioned in Wang Tao’s “Wai Tai Mi Yao” targets symptoms of agitation and mental confusion, both of which are important indicators of CI.45 Both formulations contain PT and MO, which has traditionally been used to treat neurological conditions, thus clarifying the traditional application of PM. Previous research has indicated that PM can enhance gastrointestinal motility,46 promote the growth and proliferation of gastric stromal cells, lower NO levels in the gastric antrum, decrease the expression of VIP,47 hinder the absorption of glycosides derived from PT while facilitating their elimination,48 and regulate the gut microbiota and inhibiting the activation of the PI3K/AKT/NF - κB signaling pathway to alleviate the intestinal damage caused by PT.49 Additionally, the combination of PT with MO not only maintains the sedative and expectorant properties but also amplifies them, resulting in a synergistic effect and minimizing side effects.50 This research explores the mechanisms and effects of PM in enhancing CI as informed by the gut-brain axis, offering experimental support for the safe and rational application of PM. The graphical abstract illustrates the overall research design and main findings.
Materials and Methods
Chemicals and Reagents
D-gal (Lot No. 2023042601), methanol specifically designated for chromatography (Lot No. 2022032801), and formic acid (Lot No. 2022112001) were procured from Chengdu Cologne Chemical Co., Ltd. (Sichuan, China). Additionally, chromatographic-grade acetonitrile (Lot No. RBXA1H) was sourced from SK Chemicals (Gyeonggi-do, South Korea). 4% paraformaldehyde (Lot No. GP231030112930) and goat serum (Lot No. GA2403012) were both purchased from Wuhan Service Biotechnology Co., Ltd. (Wuhan, China). Nissl solution (Lot No. 0704A24) was obtained from Jiangsu Edison Biotechnology Co., Ltd. (Jiangsu, China). TEN (Lot No. 111849–202,207), Polygalaxanthone III (Lot No. 111850–202,207), 3.6’-diferuloyl sucrose (Lot No. 111848–202,307), and Polygalacic acid (Lot No. 111939–201201) were purchased from the National Institute for Food and Drug Control (Beijing, China). Honokiol (Lot No. MUST-17031102) and Magnolol (Lot No. MUST-17020205) were both obtained from Chengdu Master Biotechnology Co., Ltd. (Sichuan, China). Mouse TNF-α (Lot No. 202402), IL-1β (Lot No. 202402), IL-6 (Lot No. 202405), IFN-γ (Lot No. 202402), LPS (Lot No. 202402), superoxide dismutase (SOD, Lot No. 202402), GSH (Lot No. 202402), cAMP (Lot No. 202405), and SCFA (Lot No. 202405) kits were obtained from Shanghai Fankwei Industrial Co., Ltd. (Shanghai, China). The BCA protein assay kit (Lot No. 037C1500) was sourced from Shanghai EpiZyme Biotechnology Co., Ltd. (Shanghai, China). Occludin (Catalog#272260-1-AP), ZO-1 (Catalog#21773-1-AP), p65 (Catalog#10745-1-AP), and p-p65 (Catalog#82335-1-AP) were all purchased from Wuhan San Ying Biotechnology Co., Ltd. (Wuhan, China). p-PKA (Catalog#HA721864), PKA (Catalog#ER64618), p-CREB (Catalog#ET7107-93), CREB (Catalog#ET-1601-15), GAPDH (Catalog#ET1601-4), goat anti-rabbit HRP (Catalog#HA1001), and iFluor™ 488 (Catalog#HA1121) were all obtained from Hangzhou Hua’an Biotechnology Co., Ltd. (Hangzhou, China).
Preparation of Plant Materials and Aqueous Extracts
PT (batch number: 221101, report number: 5–273-02-221101) and MO (batch number: 221201, report number: 5–259-02-221201) were acquired from Sichuan Guoqiang Traditional Chinese Medicine Products Co., Ltd. (Sichuan, China). The specifications outlined in the 2020 edition of the Pharmacopoeia of the People’s Republic of China are met by both PT and MO.51 The methodologies for extraction and the ratio of the combined PM were informed by prior research conducted by our team.48,52 We utilized 50 g of PT alongside 100 g of MO. PT and MO were combined with ten times its volume of distilled water, allowed to soak for 30 minutes, and then brought to a boil. After reaching a boil, the heat was lowered; PT was simmered for one hour, then filtered and concentrated under reduced pressure to produce 50 mL, whereas MO continued to simmer for an additional 15 minutes, was filtered, and concentrated to obtain 100 mL.
Additionally, 50 g of PT was obtained and mixed with 500 mL of distilled water, allowing it to soak for half an hour, brought to a high boil, and then simmered at a lower temperature for 45 minutes. The 100 g of MO that had been previously soaked was merged with the PT infusion, which was heated for 45 minutes, brought to a boil, and then simmered for another 15 minutes. After filtering, the mixture was concentrated under reduced pressure, resulting in a yield of 50 mL. This final solution was then stored separately at −80°C.
Component Analysis
The methods of analysis were derived from earlier studies conducted by the research team.49 A 70% methanol solution was utilized to dissolve PT, MO, and PM, followed by filtration through a 0.22 μm microporous membrane. An UPLC system (Vanquish, Thermo Scientific, Waltham, MA, USA) was utilized for the analytical procedures. This UPLC system was effectively combined with a triple quadrupole mass spectrometer (TSQ Fortis, Thermo Scientific, Waltham, MA, USA). Notably, the mass spectrometer featured advanced electrospray ionization capabilities. The samples were separated using a C18 column (3×100 mm, 2.6 μm) kept at a temperature of 40°C, with a flow rate of 0.2 mL/min and an injection volume of 5 μL. The gradient elution profile was designed as follows: 0–5 minutes, 2%-10% B; 5–15 minutes, 10%-30% B; 15–20 minutes, 35%-50% B; 20–25 minutes, 50%-70% B; 25–30 minutes, 70%-95% B; 30–35 minutes, 95% B. In the mass spectrometry process, the setup included positive ionization at 4000 V, negative ionization set at 3500 V, sheath gas pressure at 35 kPa, auxiliary gas pressure at 10 kPa, and a gas temperature maintained at 350°C. For the data collection and analysis, Thermo Scientific Xcalibur (version 4.2) was employed.
Animals and Experiments
Eight-week-old male KM mice, weighing between 18 and 22 grams, were obtained from Sibeifu Biotechnology Co., Ltd. (No. 110324231107055163, Beijing, China) and housed in a facility for small animal observation, where humidity was kept at 65 ± 5% and temperature was regulated to 22 ± 2°C with 12 h/12 h light-dark cycle. Prior to the commencement of the study, all animal experiments were conducted in accordance with the principles of animal ethics and the Regulations for the Administration of Experimental Animals established by the People’s Republic of China. These experiments were reviewed and approved by the Animal Welfare and Ethics Committee of Chengdu University of Traditional Chinese Medicine, with the approval number 2024046.
The mice were randomly split into multiple groups, with each group consisting of 8 individuals: the Control group and the D-gal group, Donepezil group (0.75 mg/kg, designated as Don + D-gal), low-dose PT group (0.25 g/kg, referred to as P L + D-gal), medium-dose PT group (0.5 g/kg, labeled P M + D-gal), high-dose PT group (1 g/kg, denoted P H + D-gal), low-dose MO group (0.5 g/kg, M L + D-gal), medium-dose MO group (1 g/kg, M M + D-gal), high-dose MO group (2 g/kg, M H + D-gal), low-dose PM group (0.25 + 0.5 g/kg, PM L + D-gal), medium-dose PM group (0.5 + 1 g/kg, PM M + D-gal), and high-dose PM group (1 + 2 g/kg, PM H + D-gal). For a duration of 42 consecutive days, all mice, apart from those in the control group, were administered subcutaneous injections of D-gal dissolved in saline at a dosage of 500 mg/kg·d to create a cognitive impairment model. In contrast, the mice in the control group received an equivalent volume of saline. Based on the human drug dosage, the drug doses for mice were determined using the body surface area conversion method.51,53 After the D-gal injections, each treatment group was given the respective medications orally for 42 days, whereas the control and D-gal treated mice were provided daily with distilled water at a rate of 10 mL/kg·d.
On the 43rd day, mice underwent anesthesia using a 10% urethane solution. Following this, brain tissues, along with intestinal tissues and their respective contents, were obtained from the mice. A segment of both the brain and intestinal tissues was preserved in a 4% paraformaldehyde solution and subsequently stained using H&E or Nissl staining. The remaining hippocampal tissue was isolated from the brain and preserved in 3% glutaraldehyde for examination via transmission electron microscopy. Additionally, the extra hippocampal tissue, along with intestinal tissue and its contents, was kept at −80°C. The hippocampal tissue was allocated for ELISA and WB studies, whereas the intestinal tissue was designated for Western blot analysis, and the intestinal contents were arranged for 16S rRNA metagenomic assessment. All experimental procedures were carried out following the established protocol (Figure 1).
 |
Figure 1 Experimental flow diagram. |
Behavioral Tests
The cognitive function in mice was evaluated using the PAT (TechMan Soft, Sichuan, China) and the MWM (WMT-100S, Sichuan, China).53 Testing with the PAT occurred the day after the mice had undergone adaptive training in the apparatus. The PAT-8 video analysis system systematically captured both the latency for escape and the count of errors made. During the MWM spatial navigation assessment, all mice participated in one day of adaptive training with a visible platform, which was followed by four days of trials that involved a hidden platform in the MWM apparatus, with each trial lasting 60 seconds. On day six, the platform was removed, and a spatial exploration assessment was performed in the MWM. The WMT-100S video imaging analysis system automatically recorded the distance swum in the designated quadrant, the number of platform crossings, and the time spent in the target area.
Histological Examination
Following fixation in a solution of 4% paraformaldehyde for a duration of 48 hours, the brain and intestinal samples underwent dehydration, embedding, and sectioning. Sections of mouse brain and intestinal tissue were prepared with a uniform thickness of 5 μm. These sections, which had been embedded in paraffin, underwent staining using the H&E method. Following the staining process, the sections were examined and visualized using a microscope (Sunny, Hong Kong, China). Brain tissue sections were stained using Nissl staining technique and scanned with a digital pathology scanning system (HS6, Zhejiang, China).For IF, the paraffin sections were incubated in an oven set at 60°C for one hour, deparaffinized using a gradient of ethanol, and subjected to antigen retrieval with an EDTA solution. The sections underwent permeabilization using 0.5% Triton for thirty minutes and were subsequently blocked at room temperature with goat serum for one hour. Following this, they were incubated overnight with a primary antibody specific to p-CREB (1:100) and then treated with a secondary antibody (1:200) at room temperature for an additional hour. All sections were then counterstained with DAPI to enable visualization of the nuclei. Fluorescent images were captured employing a fluorescence microscope (Zeiss Axio Imager 2, Germany), and fluorescence intensity was measured utilizing ImageJ software. For TEM, the tissues were subjected to a 3% glutaraldehyde solution, fixed with 1% osmium tetroxide, dehydrated in acetone, and infiltrated with epoxy resin (Epox) 812 over a prolonged period. Once embedding was finalized, ultrathin sections were cut using a diamond knife, stained with uranyl acetate and lead citrate, and examined with a JEM-1400-FLASH TEM (JEOL, Japan).
ELISA Assay
Weigh the brain tissue of the mouse and homogenize it using a 1:10 ratio with physiological saline. After centrifuging, collect the supernatant to measure the protein concentration. Transfer the mouse blood into a centrifuge tube and let it rest at room temperature for a duration of two hours. Following this interval, isolate the serum using high-speed centrifugation at low temperatures. Subsequently, transfer it into a new centrifuge tube and store it at −80°C. Next, assess the levels of TNF-α, IL-1β, IL-6, IFN-γ, and LPS in the mouse serum, and evaluate TNF-α, IL-1β, SOD, GSH, cAMP, and SCFA concentrations in the mouse brain tissue following the guidelines provided by the manufacturer’s ELISA kits.
Network Pharmacology
Drug targets were identified through the utilization of various databases, which include CNKI (https://www.cnki.net/), PubMed (https://pubmed.ncbi.nlm.nih.gov/), Herb (http://herb.ac.cn/), SwissTargetPrediction (http://www.swisstargetprediction.ch/), TCMSP (https://old.tcmsp-e.com/), and BATMAN-TCM (http://bionet.ncpsb.org.cn/batman-tcm/index.php/Home/Index/index). Additionally, genes that are linked to CI were discerned using the GeneCards (https://www.genecards.org/) and OMIM databases (https://omim.org/), enabling a more focused investigation into the genetic factors contributing to CI. To further refine the analysis, the intersection between the potential drug targets for MO and PT with the CI-related genes was established using the Venny 2.1 tool (https://bioinfogp.cnb.csic.es/tools/venny/). This crossing enabled the recognition of common targets that could be involved in the therapeutic uses of MO and PT within CI contexts. The analysis was deepened by utilizing the STRING 12.0 database (https://string-db.org/) to evaluate potential therapeutic targets, from which a protein-protein interaction (PPI) network was developed using Cytoscape 3.7.2. This network serves as a visual representation of the interactions among identified proteins, thus providing insight into their functional relationships. For further visualization and analytical depth, additional tools were employed, including a bioinformatics online analysis and visualization cloud platform (https://www.bioinformatics.com.cn/). Through this platform, KEGG pathway enrichment bubble charts and GO functional enrichment bar charts were generated, highlighting significant biological pathways and functions associated with the therapeutic actions of PT. The foremost 20 pathways, selected according to their P-values, were deliberately identified to highlight essential pathways related to the treatment of CI. Ultimately, the tool Cytoscape 3.7.2 was employed to create an extensive network diagram illustrating the relationships among drugs, diseases, key targets, and important pathways. This diagram not only illustrates the interrelations but also contributes to a deeper understanding of the therapeutic landscape for cognitive impairment.
16S rRNA Sequencing of Gut Microbiota
Measure the total DNA extracted with a Nanodrop spectrophotometer. Employ PCR to amplify the variable rRNA gene regions or particular gene fragments, and subsequently purify the resulting PCR products with Vazyme VAHTS™ DNA Clean Beads, eluting them in buffer. Use the Illumina TruSeq Nano DNA LT Library Prep Kit to generate the sequencing library. Perform paired-end sequencing on the MiSeq sequencer by employing the MiSeq Reagent Kit V3 (600 cycles). Implement sequence denoising or OTU clustering per the QIIME2 DADA2 analysis protocol. Use QIIME2 (version 2019.4) software for species annotation.
Western Blot Analysis
The samples underwent lysis using RIPA buffer, after which the supernatant was obtained through centrifugation. The concentration of total protein in the samples was measured utilizing the BCA protein assay method. To separate the target proteins, SDS-PAGE was utilized, and these proteins were then transferred onto PVDF membranes. Primary antibodies, including Occludin (1:5000), ZO-1 (1:10,000), NF-κB-p65 (1:1000), p-NF-κB-p65 (1:2000), PKA (1:1000), p-PKA (1:1000), CREB (1:1000), p-CREB (1:1000), and GAPDH (1:5000), utilized to incubate the membranes overnight at 4°C. On the following day, the membranes underwent treatment with a secondary antibody diluted to 1:50,000. Visualization and semi-quantification of the bands were performed through a chemiluminescence imaging and analysis system (GelView 6000Plus, Guangzhou, China).
Statistical Analysis
The results are displayed as mean ± SEM. Statistical analyses were performed using GraphPad Prism 8 software. To compare the groups, a one-way analysis of variance (ANOVA) was executed, alongside suitable post hoc tests. A p-value of less than 0.05 was considered statistically significant.
Result
Determination of Characteristic Components in PT, MO, and PM
The findings illustrate a comparison of mass-to-charge ratios alongside standard samples (Figure 2A). In PT, components such as TEN, Polygalaxanthone III, 3.6’-disinapoyl sucrose, and Polygalacic acid were identified (Figure 2B). In MO, the components Honokiol and Magnolol were observed (Figure 2C). Additionally, these specific components were also found in the PM combination (Figure 2D).
 |
Figure 2 UPLC-MS/MS Examination of PT, MO, and PM. (A) Typical constituents in the standard mixture of PM, (B) PT sample, (C) MO sample, (D) sample of PM. |
Effects of PT, MO, and PM on Cognitive Dysfunction and Neuropathology Induced by D-gal in Mice
The findings of the PAT study demonstrated that, when compared to the D-gal group, every dosage category of PT and PM, in conjunction with the M M and M H groups, significantly prolonged the escape latency while reducing the number of errors (P < 0.05 or P < 0.01, Figure 3A and B). In addition, compared with each dose group of MO, PM significantly increased the escape latency of mice (P < 0.01, Figure 3A and B). When compared with the P M and P H groups, both the PM M and PM H groups significantly enhanced the escape latency of mice (P < 0.05 or P < 0.01, Figure 3A and B).
 |
Figure 3 PT, MO, and PM Alleviate Cognitive Impairments Induced by D-gal. PAT results showing (A) escape latency and (B) number of errors. MWM results showing (C) escape latency, (D) accumulated swimming distance in target quadrant of the spatial exploration test, (E) times of crossing the target quadrant, (F) time in target quadrant, and (G) representative swimming trajectories. n = 8. #P < 0.05, ##P < 0.01 vs Control group; *P < 0.05, **P < 0.01 vs D-gal group, ○P < 0.05, ○○P < 0.01 vs PT group, ΔP < 0.05, ΔΔP < 0.01 vs MO group. |
When compared to mice treated with D-gal, there was a notable decrease in escape latency during the MWM navigation test for the P M, P H, M H, and all PM dosage groups (P < 0.05 or P < 0.01, Figure 3C). In the spatial exploration phase of the MWM, there was a significant enhancement in the total swimming distance within the target quadrant for the P M, P H, and all PM dosage groups (P < 0.01, Figure 3D). Compared with each corresponding dose group of PT and MO, PM significantly increased the swimming distance of mice in the target quadrant (P < 0.05 or P < 0.01, Figure 3D). Furthermore, there was a notable increase in the frequency of platform crossings observed in the P M, P H, M H, and all PM treatment groups (P < 0.05 or P < 0.01, Figure 3E). Compared with P M, M L, and M M, the corresponding dose of PM significantly increased the number of times mice crossed the target quadrant (P < 0.05 or P < 0.01, Figure 3E). In the P M, P H, M H, and all PM treatment groups, a lengthened time was spent in the target quadrant (P < 0.05 or P < 0.01, Figure 3F). Compared with P L, P H, and each dose group of MO, PM significantly increased the dwell time of mice in the target quadrant (P < 0.05 or P < 0.01, Figure 3F). It is noteworthy that PM treatment yielded better behavioral results than PT or MO treatments (Table 1).
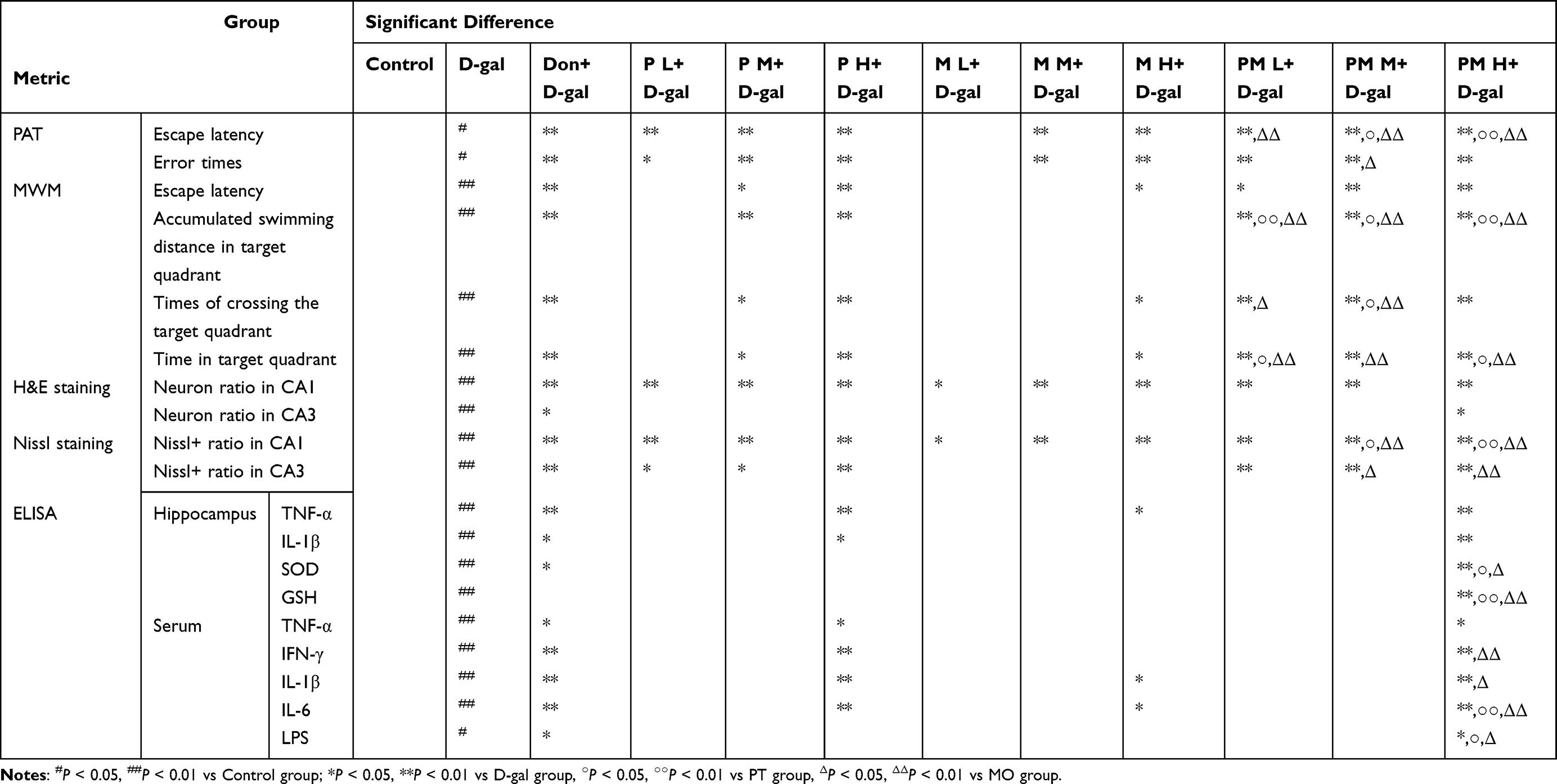 |
Table 1 Significant Differences in PT, MO, and PM in Behavioral Tests, Pathological Staining, and ELISA |
Furthermore, PT, MO, and PM interventions significantly mitigated neuropathological alterations in the hippocampus of aging mice, which included neuronal cytoplasmic shrinkage, disordered arrangement, and a decrease in neuronal counts (Figure 4A). In comparison to the D-gal group, administering PT, MO, and all PM dosages resulted in a significant increase in neuronal cells within the CA1 region (P < 0.05 or P < 0.01, Figure 4B), while the PM H group exposed a substantial enhancement of neuronal numbers in the CA3 region (P < 0.05, Figure 4C). Nissl staining results showed that the Nissl+ neuron rate in the D-gal group was significantly reduced compared to the Control group (P < 0.01, Figure 4D–F). In the CA1 region, PT, MO, and PM at all doses significantly restored the Nissl+ neuron rate reduced by D-gal (P < 0.05 or P < 0.01, Figure 4E). Compared with P M, P H, M M, and M H, the corresponding doses of PM significantly increased the Nissl+ neuron rate in the CA1 region of mice (P < 0.05 or P < 0.01, Figure 4E). In the CA3 region, PT and PM at all doses significantly increased the Nissl+ neuron rate (P < 0.05 or P < 0.01, Figure 4F). Compared with M M and M H, the corresponding dose groups of PM significantly enhanced the Nissl+ neuron rate in the CA3 region (P < 0.05 or P < 0.01, Figure 4F). The Nissl staining results further demonstrated the neuroprotective effect of PM. Notably, PM exhibited superior recovery of neuronal pathology compared to either PT or MO alone (Table 1). These results indicate that PT, MO, and PM may improve cognitive impairments in D-gal-made models by reducing neuronal damage.
 |
Figure 4 PT, MO, and PM Alleviate D-gal-Induced Neuropathological Damage. (A–C) Representative images and quantitative analysis of HE staining in the hippocampal CA1 and CA3 regions (scale bar: 200 μm and 50 μm). (D–F) Representative images and quantitative analysis of Nissl staining in the hippocampal CA1 and CA3 regions. Black arrows indicate shrunken neurons (scale bar: 500 μm and 50 μm). n = 5. ##P < 0.01 vs Control group; *P < 0.05, **P < 0.01 vs D-gal group, ○P < 0.05, ○○P < 0.01 vs PT group, ΔP < 0.05, ΔΔP < 0.01 vs MO group. |
PT, MO, and PM Treatments Alleviated Inflammation and Oxidative Stress Induced by D-gal
Neuropathological research involving mice suggests that PM is effective in diminishing neuronal damage, with PM H showing the most notable enhancement. Considering that oxidative stress and inflammation contribute to neuronal damage, we investigated the effects of P H, M H, and P M H on markers of oxidative stress and pro-inflammatory factors in serum and brain tissue. In the hippocampal area, the administration of P H, M H, and PM H resulted in a notable decrease in TNF-α levels compared to the D-gal group (P < 0.05 or P < 0.01, Figure 5A). Additionally, both P H and PM H induced a significant reduction in IL-1β level (P < 0.05 or P < 0.01, Figure 5B), while PM H specifically increased the levels of SOD and GSH (P < 0.01, Figure 5C and D). Additionally, compared with P H and M H, PM H significantly increased the levels of SOD and GSH in the hippocampus of mice (P < 0.05 or P < 0.01, Figure 5C and D). Serum analysis revealed that P H and PM H significantly reduced levels of TNF-α and IFN-γ as well (P < 0.05 or P < 0.01, Figure 5E and F). Compared with M H, PM H significantly decreased the level of IFN - γ (P < 0.01, Figure 5F). Additionally, P H, M H, and PM H effectively reduced IL-1β and IL-6 levels (P < 0.05 or P < 0.01 Figure 5G and H), with PM H also significantly decreasing the concentration of LPS (P < 0.05, Figure 5I). Compared with M H, PM H significantly decreased the levels of IL - 1β, IL - 6 and LPS (P < 0.05 or P < 0.01, Figure 5G–I). Compared with P H, PM H significantly reduced the levels of IL - 6 and LPS (P < 0.05 or P < 0.01, Figure 5H and I). These results indicate that, compared with P H and M H, PM H exhibits more potent anti - inflammatory and antioxidant effects (Table 1).
 |
Figure 5 PT, MO, and PM alleviate inflammation and oxidative stress in D-gal mice. Levels of (A) TNF-α, (B) IL-1β, (C) SOD, and (D) GSH in the hippocampus of mice, and (E) TNF-α, (F) IFN-γ, (G) IL-1β, (H) IL-6, and (I) LPS in mouse serum. n = 6. #P < 0.05, ##P < 0.01 vs Control group; *P < 0.05, **P < 0.01 vs D-gal group, ○P < 0.05, ○○P < 0.01 vs PT group, ΔP < 0.05, ΔΔP < 0.01 vs MO group. |
Network Pharmacology Analysis
Using the Venny 2.1 tool to analyze the intersection between pulmonary PM and CI, A total of 232 overlapping targets were identified, which could act as possible therapeutic options for tackling CI within the framework of PM (Figure 6A). This finding underscores the relevance of these targets in the pursuit of effective treatments. Furthermore, in our examination of the protein-protein interaction (PPI) network, we noted that nodes that are larger in size and display darker colors correspond to higher degree values, thus emphasizing their importance within the overall network of interactions. Our comprehensive data analysis indicated that among all the identified nodes, the five core nodes exhibiting the highest degree values were INS, TNF, PPKACA, CASP3, and IL1B (Figure 6B). These core nodes are critical for understanding the biochemical pathways at play and could be central to developing targeted interventions for managing cognitive impairment associated with pulmonary metastasis.
 |
Figure 6 Network pharmacology analysis of PM. (A) Venn diagram, (B) PM’s medicinal effects on key target protein-protein interaction (PPI) network diagram, (C) GO enrichment analysis, (D) KEGG enrichment analysis, (E) PM for CI disease key target protein-key pathway network diagram, (F) cAMP and NF-κB KEGG pathway diagrams. The red box represents the signaling pathway validated in our study. |
The data was subsequently entered into the Metascape database to conduct an enrichment analysis, resulting in a total of 2730 Gene Ontology (GO) annotations. The total includes 2269 biological processes (BP), 189 cellular components,54 and 272 molecular functions (MF). For each category, the ten highest-ranked entries were chosen based on their P-value rankings and visualized using the online tool available on the Microbioinformatics website. In terms of BP, the primary functions that were identified included the regulation of membrane potential, system process regulation, chemical synapse function, and the processes of synaptic signaling. In the context of CC, the primary components recognized included the synaptic membrane, the complex of monatomic ion channels, the transport complex, and the transmembrane transporter assembly. Within the MF category, the key functionalities encompassed activities related to monatomic ion transmembrane transporters, transmembrane transport for inorganic substances, monatomic cation transmembrane transport activity, and functions associated with monatomic ion channels (Figure 6C).
The KEGG analysis proceeded to select the twenty pathways that displayed the smallest P-values and the highest counts of gene enrichment for the purposes of visualization, as illustrated (Figure 6D). In this representation, pathways characterized by more intense red colors indicate the presence of smaller P-values, elevated gene enrichment counts, and a correspondingly stronger significance of the pathways themselves. The visual data suggests that the therapeutic impact of PM on CI may be closely linked to certain biological pathways, particularly the cAMP signaling pathway and the NF-κB signaling pathway (Figure 6F). To further elucidate these relationships, we imported the top twenty pathways along with their relevant targets into Cytoscape 3.7.2 software. This action allowed us to construct a comprehensive network diagram that highlights the interconnections among the disease, significant biological targets, and critical pathways (Figure 6E). In this network, the red nodes signify the crucial biological targets, while the blue nodes depict the important pathways, and the yellow node designates the disease itself. In total, the diagram encompasses 149 nodes and 607 edges, underscoring that the mechanism through which PM operates against CI involves a complex interplay of various components, biological targets, and significant pathways.
PM Improved D-gal-Induced Barrier Damage
D-gal induction has the potential to cause harm to the intestines, which includes compromising the intestinal barrier. Histological assessments utilizing H&E staining revealed that PM treatment markedly ameliorated the pathological damage induced by D-gal, including disarranged villi architecture, fractures, and apical detachment (Figure 7A). To enhance the understanding of intestinal barrier integrity, WB analysis was conducted to assess the levels of TJs proteins, particularly Occludin and ZO-1. In comparison to the Control group, a notable reduction in the levels of Occludin and ZO-1 was observed after treatment with D-gal. Importantly, administration of PM at both medium and high doses notably augmented the levels of these proteins in the hippocampus (P < 0.01, Figure 7B and C). Additionally, D-gal treatment resulted in damage to the BBB, which resulted in tight junction dysfunction; however, this effect was mitigated by PM treatment (Figure 7D). Moreover, the levels of Occludin and ZO-1 proteins in the hippocampus experienced a significant reduction after D-gal treatment, while PM treatment led to a notable increase in their expression (P < 0.05 or P < 0.01, Figure 7E and F).
 |
Figure 7 PM regulates the integrity of the barrier. (A) H&E staining of mouse intestinal tissues (scale bar = 200 μm), (B and C) WB analysis of Occludin and ZO-1 in the intestine, (D) Representative TEM images of the hippocampal region (scale bar = 200 nm), (E and F) WB analysis of Occludin and ZO-1 in the hippocampus. Red arrows indicate villus breakage, black arrows indicate apical detachment of villi, and Orange arrows indicate TJs. n = 3. ##P < 0.01 vs Control group; *P < 0.05, **P < 0.01 vs D-gal group. |
PM Treatment Modulates NF-κB and cAMP Signaling Pathways in the Hippocampus of D-gal-Induced CI Mice
The findings from the WB analysis indicated that, when contrasted with the D-gal group, there was a significant reduction in the relative expression levels of p-NF-κB-p65 in the hippocampal tissue (P < 0.01, Figure 8A–C). In contrast, there was a notable elevation in the levels of p-PKA and p-CREB (P < 0.05 or P < 0.01, Figure 8A–C). Further evaluations through ELISA revealed that treatment with PM resulted in a significant enhancement of both cAMP and SCFA concentrations within the hippocampus in comparison to the D-gal group (P < 0.01, Figure 8D). Moreover, IF studies demonstrated that PM treatment significantly increased the levels of p-CREB expression in the CA1 and CA3 regions (P < 0.05 or P < 0.01, Figure 8E and F). These results suggest that PM is involved in inhibiting the NF-κB inflammatory signaling pathway within the hippocampus while also promoting the cAMP signaling pathway.
 |
Figure 8 PM Regulates NF-κB and cAMP Signaling Pathways. (A–C) WB analysis of p-NF-κB-p65, p-PKA, and p-CREB in the hippocampus, n=3, (D) Expression of SCFA and cAMP in the mouse hippocampus, n=6, (E and F) Immunofluorescence analysis and quantification of p-CREB (S133) in the CA1 and CA3 regions (scale bar = 20 μm), n = 3. ##P < 0.01 vs Control group; *P < 0.05, **P < 0.01 vs D-gal group. |
PM Treatment Modulates Gut Microbiota in D-gal-Induced CI Mice
The rarefaction curve for α-diversity, which is derived from the count of operational taxonomic units (OTUs) across various samples, levels off on the right, suggesting that saturation has been achieved in the sequencing data (Figure 9A). The assessment of α-diversity, utilizing the Shannon and Simpson indices, revealed that the PM group demonstrated a reduction in species richness compared to the D-gal group (P > 0.05, Figure 9B). Results from Principal Coordinate Analysis (PCoA) clearly displayed distinct separations between the D-gal, Control, and PM groups, highlighting significant differences in the gut microbiota structure of the PM group (Figure 9C). Additionally, the Venn diagram depicts the quantity of shared and distinct OTUs among the groups, revealing that there are 497 OTUs common across all three groups (Figure 9D).
Figure 9 Continued. Figure 9 PM Alleviates D-gal-Induced Gut Microbiota Dysbiosis. (A) Diversity rarefaction curve, (B) Diversity indices based on OTU levels, (C) β-diversity of gut microbiota based on Principal Coordinate Analysis (PCoA) and unweighted UniFrac algorithm, (D) Pie chart of OTU numbers for each group, (E) Bar chart at the phylum level, (F) Comparison of relative abundance of different phylum among groups, (G) Firmicutes/Bacteroidetes ratio, (H) Bar chart at the families level, (I) Comparison of relative abundance of different families among groups, (J) Bar chart at the genus level, (K) Comparison of relative abundance of different genus among groups. (L) LDA value distribution histogram and LEfSe circular phylogenetic tree. n = 5. #P < 0.05, ##P < 0.01 vs Control group; *P < 0.05, **P < 0.01 vs D-gal group.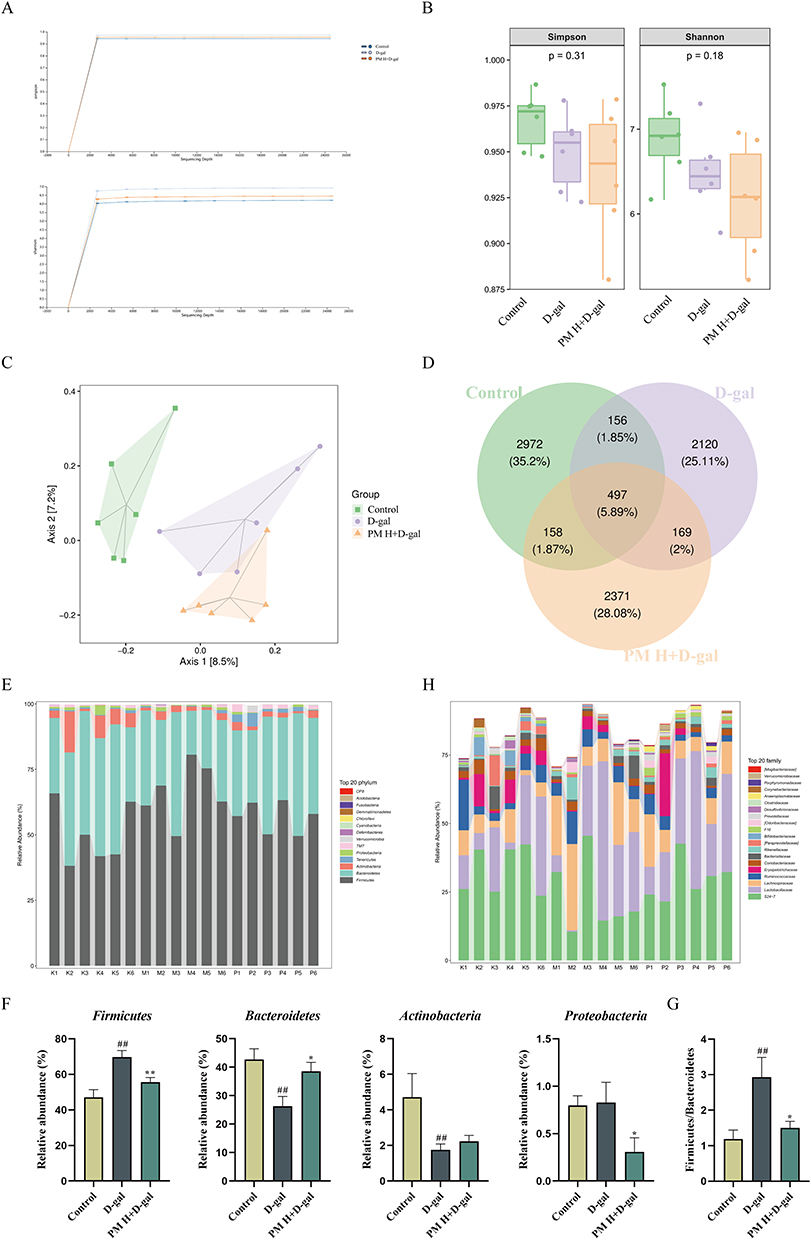

Further analysis of the species diversity at different taxonomic tiers revealed notable changes in the gut microbiome. At the phylum level, the introduction of PM resulted in a substantial differences in the relative abundance of Bacteroidetes and Proteobacteria compared to the group treated with D-gal (P < 0.05, Figure 9E and F). Additionally, a noticeable trend was observed indicating a rise in Actinobacteria (P > 0.05, Figure 9E and F). Conversely, the treatment with PM corresponded to a significant decline in the relative abundance of Firmicutes (P < 0.01, Figure 9E and F), along with a lower Firmicutes/Bacteroidetes ratio, which suggests a modification in the composition of the microbiota (P < 0.05, Figure 9G). At the family taxonomic level, the relative abundance of Desulfovibrionaceae decreased with PM treatment when compared to the D-gal group (P > 0.05, Figure 9H and I). In contrast, a marked reduction in the relative abundance of Enterobacteriaceae was observed following PM treatment (P < 0.05, Figure 9H and I). When analyzing the data at the genus level, there was a trend observed with PM treatment that indicated a rise in the relative abundance of Bacteroides and Coprococcus in comparison to the D-gal group (P > 0.05, Figure 9J and K). Notably, PM led to significantly higher relative abundances of Prevotella, Anaeroplasma, and Clostridium (P < 0.05 or P < 0.01, Figure 9J and K). These results demonstrate the significant impact of particulate matter on the composition of gut microbiota at various taxonomic levels.
The results suggest that PM significantly influences dysbiosis and modifies the alterations in gut microbiota caused by D-gal. Biomarkers with an LDA score greater than 4 were identified by employing linear discriminant analysis effect size (LEFSE) (Figure 9L). For the PM group, the identified biomarkers comprised Parabacteroides and Gemmiger, whereas the D-gal group presented biomarkers like [Ruminococcus] and Ruminococcus. Additionally, the PM group showed the presence of other biomarkers including Bifidobacterium and Desulfovibrio.
Discussion
Both PT and MO notably enhance CI and show promise as treatment options; their synergistic pharmacological effects can work in harmony, resulting in improved therapeutic results.33 The pairing of PT and MO amplifies the soothing and cognitive-boosting benefits of PT, while MO contributes its dampness-drying and phlegm-eliminating attributes to support PT’s expectorant actions. Together, they more effectively balance heart and kidney functions, thus alleviating symptoms associated with CI.46,52,55 Regarding the interactions and synergistic effects of the bioactive components of PT and MO, studies have shown that after their combination, significant changes occur in the content and composition ratio of chemical components, laying a material foundation for the synergistic effects.56 MO can effectively alleviate the gastrointestinal motility inhibition induced by PT, and the mechanism may be related to the changes in component content.55 In regulating gastrointestinal smooth muscle, their combination can significantly enhance the contraction amplitude, reduce the frequency inhibition rate, and weaken the inhibition on intestinal muscles.57 PT polysaccharides alleviate gut microbiota dysbiosis and neuroinflammation in AD models by increasing the abundance of SCFA-producing bacteria (eg, Bacteroides) and decreasing pro - inflammatory bacteria (eg, Proteobacteria).58 PT scavenges free radicals and reduces the increased intestinal permeability induced by oxidative stress.59,60 The ethanol extract of MO alleviates intestinal inflammation and barrier damage by inhibiting the PI3K/AKT/NF-κB pathway.49 Moreover, magnolol, the main component of MO, can reverse gut microbiota imbalance (eg, increasing Ruminococcaceae and decreasing Firmicutes) and improve lipid metabolism disorders (eg, reducing serum LPS and regulating bile acid metabolism), thereby protecting the intestinal barrier.61–64 PT and MO have different focuses on the gut - brain axis. Their combination may optimize the microbiota structure, target oxidative stress and inflammatory cascades simultaneously, comprehensively supporting the hypothesis of PM’s multi-target treatment for CI. These interactions can explain the enhanced effects of PM.
The fundamental principle by which D - gal induces CI is to simulate the biochemical changes of CI via long - term or high - dose D - gal injection. During the metabolic process in animals, a substantial amount of reactive oxygen species (ROS) is generated. This leads to an elevation in oxidative stress levels, the occurrence of neuroinflammation, neuronal damage, and a decline in cognitive function, thereby mimicking the process of human cognitive decline.65–69 D-gal induction may also lead to the activation of intestinal inflammation. Pro-inflammatory cytokines such as TNF-α, IL-1β, IL-6, and IFN-γ can enter the central nervous system through the blood circulation.70 These cytokines act on nerve cells and glial cells in the brain, triggering neuroinflammation, affecting the synthesis, release, and metabolism of neurotransmitters, interfering with neural signal transduction, and subsequently leading to cognitive impairment.71,72 After LPS enters the blood circulation, it can activate the immune system and inflammatory signaling pathways, further exacerbating the systemic inflammatory response and indirectly affecting brain function, thus forming an intestinal-immune-brain signaling pathway.73,74
The imbalance of the gut microbiota caused by D-gal induction may lead to an increase in the level of intestinal oxidative stress, a decrease in SOD activity, and a reduction in GSH content.75 Oxidative products generated by oxidative stress can reach the brain through the blood circulation, damaging neurons and glial cells and affecting neural plasticity and neurotransmitter function.76 Meanwhile, the oxidative stress response in the brain can also affect the redox state of the intestine through neuroendocrine signal feedback, forming an interaction between the intestine, oxidative stress, and the brain.77 The research indicated that mice treated with D-gal displayed deficits in learning and memory when subjected to behavioral assessments, whereas PM significantly alleviated these impairments. Furthermore, pathological staining indicated that PM effectively countered the reduction in neuron counts and neuronal damage associated with D-gal treatment. As a reaction to the oxidative stress and the systemic inflammatory response triggered by D-gal, PM significantly reduced the levels of pro-inflammatory mediators such as TNF-α and IL-1β in the serum and hippocampus of subjects treated with D-gal. Additionally, PM significantly increased SOD activity and elevated GSH levels within the hippocampus, thereby mitigating oxidative stress in these mice. Moreover, PM reduced the content of pro - inflammatory substances in the serum. Through these pharmacological investigations, we established that PM was more effective than PT or MO individually in enhancing cognitive functions, pathological outcomes, and molecular markers in D-gal-treated mice. This highlights that the combination of PT and MO produces a pharmacological synergy, leading to improved therapeutic results.
The appropriate organization and operation of the intestinal barrier are associated with various diseases affecting the central nervous system, including Alzheimer’s disease and Parkinson’s disease.78 In the CI model, D - gal induces damage to the intestinal barrier. The underlying mechanism may involve the production of a large amount of ROS, which triggers oxidative stress. Ultimately, this leads to apoptosis of epithelial cells or damage to the structure of TJs, thus disrupting the integrity of the intestinal barrier.79,80 Research has shown that the active ingredients of traditional Chinese medicine can regulate the gut microbiota, significantly reduce the LPS content in the serum of model mice, improve the apoptosis and disordered arrangement of intestinal epithelial cells, upregulate the mRNA expression levels of ZO - 1, Occludin, and Claudin - 1, and contribute to the restoration of TJs between cells. This, in turn, helps maintain the integrity and permeability of the intestinal epithelial barrier.79,81,82 SCFAs are produced by specific gut bacteria during the fermentation of dietary fiber. They provide energy for intestinal epithelial cells, promote their growth and differentiation, and are crucial for maintaining the integrity of the intestinal barrier.83 In our study, PM increased the abundance of SCFA - producing bacteria such as Actinobacteria, Bacteroides, Prevotella, Anaeroplasma, and Clostridium, thereby enhancing the intestinal barrier function and maintaining the stability of the gut - brain axis. LPS is the main component of the cell wall of Gram - negative bacteria. Our research indicates that PM can decrease the abundance of LPS - producing bacteria. When the intestinal barrier is impaired, the LPS generated by these bacteria enters the bloodstream, activating the immune system and triggering a systemic inflammatory response.84 Subsequently, inflammatory cytokines can reach the brain via the bloodstream, activating microglia and astrocytes in the brain. These activated cells release a large number of pro - inflammatory cytokines, such as TNF - α, IL - 1β, etc., thus leading to neuroinflammation.85,86 Neuroinflammation can damage neurons, affect the synthesis and release of neurotransmitters, disrupt neural signal transduction, and ultimately lead to cognitive impairment.87 PM reduces the abundance of LPS - producing bacteria such as Proteobacteria, Desulfovibrionaceae, and Enterobacteriaceae, thereby decreasing the release of LPS. This alleviates both systemic and neuroinflammation, and further improves cognitive function. This demonstrates the impact of gut microbiota and their products on the brain’s inflammatory state within the gut - brain axis.
BBB is a pivotal protective structure that shields the brain from noxious substances and the integrity of the BBB is intricately linked to cognitive function.88 Damage to the BBB induced by D-gal may stem from different factors, such as the release of inflammatory cytokines, changes in astrocyte activation, and a decline in the expression of tight junction proteins among endothelial cells. These changes can significantly improve the permeability of the BBB, allowing LPS and pathogen molecules from the bloodstream to enter brain tissue, potentially leading to neuroinflammation and neurodegeneration.89,90 Once in the brain, LPS stimulates brain endothelial cells and activates the caspase - 4/11 - GSDMD signaling pathway. This leads to the inflammatory disruption of the BBB, making it easier for harmful substances to enter the brain, thus further exacerbating neuroinflammation and neuronal damage.90 In addition, LPS activates immune cells in the brain, such as microglia, triggering an inflammatory response in the brain. It also binds to Toll - like receptor 4, activating the downstream NF - κB signaling pathway, which generates inflammatory cytokines like TNF - α, IL - 1β, and IL - 6, thus contributing to the development of neurodegenerative diseases.91,92 Although there are certain differences in tissue structure and physiological functions between the BBB and the intestinal barrier, they share similar biological requirements in maintaining the stability of the body’s internal environment and resisting the invasion of external harmful substances. Moreover, both rely on TJs to ensure the integrity and selective permeability of the barriers.93 In this research, PM mitigated the pathological injury to the intestine in D-gal mice and restored the reduction in the levels of Occludin and ZO-1 in both the intestine and hippocampus induced by D-gal, thus safeguarding both the intestinal barrier and the BBB. This suggests that PM may have the potential to widely protect the body’s barrier functions. On one hand, PM strengthens the BBB, reduces the entry of harmful substances into the CNS, decreases neuroinflammation and nerve damage, and improves cognitive function. On the other hand, it protects the intestinal barrier, prevents the translocation of LPS and pathogens, and maintains the homeostasis of the intestinal environment. Thus, it influences the functions and health of the brain through the gut - brain axis.
SCFAs are metabolites generated by the gut microbiota during the fermentation of dietary fiber. They can reach the brain via the bloodstream and are closely linked to the onset and progression of cognitive function and neurodegenerative diseases through the gut - brain axis.94 In neurodegenerative diseases like AD and PD, a reduction in the quantity of SCFA - producing bacteria and a decrease in the concentration of SCFAs in feces have been observed.95 SCFAs exert their physiological functions via GPCRs, especially GPR41 and GPR43. Activation of these receptors can trigger the activation of multiple intracellular signaling pathways, including that of adenylate cyclase, thus increasing the intracellular cAMP level and activating the cAMP signaling pathway.96 SCFAs can impact the synthesis and metabolism of neurotransmitters, such as gamma - aminobutyric acid and serotonin, which are closely associated with cognitive function.97 SCFAs also have immunomodulatory properties. They inhibit the production of inflammatory factors and promote the secretion of anti - inflammatory cytokines, thereby maintaining the balance of the immune system.98 PM boosts the number of SCFA - producing bacteria, influences and regulates the brain’s immune response through the gut - brain axis, mitigates the occurrence and development of neuroinflammation, shields neurons from inflammatory damage, and ultimately enhances cognitive function.
The transcription factor NF-κB is essential for cellular immune responses, inflammation, cell growth, and programmed cell death.99 Activation of the NF-κB signaling pathway leads to the release of pro-inflammatory cytokines such as TNF-α, IL-1β, and IL-6, all of which play significant roles in neuroinflammation and CI. Inhibiting this NF-κB signaling pathway may reduce microglial activation and decrease neuroinflammation, which can subsequently improve cognitive function.100 Moreover, the impairment of the blood-brain barrier has been associated with the triggering of the NF-κB signaling pathway, which may exacerbate cognitive impairment.100 This research indicates that PM substantially lowered the levels of p-NF-κB-p65 in the hippocampal region of mice, alongside a decrease in pro-inflammatory cytokines in both the hippocampus and serum. These findings suggest that PM could mitigate CI induced by D-gal by inhibiting the NF-κB signaling pathway.
The cAMP signaling pathway plays an essential role as an intracellular mechanism that participates in numerous physiological functions, such as memory and learning, immune response, motor regulation, and metabolic processes.101 CREB, an important transcription factor located in the nucleus, is predominantly found in the brain, especially in areas related to memory and learning, including the cortex and hippocampus. Changes in the expression levels of CREB might be associated with cognitive impairments.102 For example, in AD, disruptions in the cAMP signaling pathway may result in lower phosphorylation levels of CREB, adversely affecting neuronal health and impairing learning and memory capacities.103 cAMP plays a crucial role in supporting neuronal survival and regeneration via PKA, which contributes to the enhancement of cognitive abilities.104 Neuroinflammation represents a significant pathological characteristic of AD, and influencing the cAMP signaling pathway might assist in regulating inflammation linked to this condition.105 In our research, the administration of PM notably enhanced the levels of p-PKA and p-CREB in the hippocampal region of mice treated with D-gal, while also causing a significant increase in cAMP concentrations in this area. These findings suggest that PM stimulates the cAMP signaling pathway within the hippocampus of D-gal-treated mice and may mitigate cognitive deficits via this process.
Individuals with CI frequently encounter dysbiosis within their gut microbiota.106 Numerous probiotic preparations can modulate the structure of the gut microbiota, increasing the relative abundance of beneficial bacteria such as Bifidobacterium and Lactobacillus. Moreover, they can alleviate intestinal inflammation and influence neurotransmitter metabolism.107,108 The ratio of Firmicutes to Bacteroidetes serves as a widely utilized measure for evaluating the balance of gut microbiota.109 In healthy individuals, the gut microbiome predominantly consists of Firmicutes and Bacteroidetes, which together account for a considerable fraction of gut bacteria. The relative presence of these two phyla can indicate the condition of the gut microbiota, and any imbalance in this ratio might be associated with a range of diseases.110 A heightened Firmicutes/Bacteroidetes ratio is linked to inflammatory responses and can impact both the gut and additional organs.111 Our study found a notable decrease in the Firmicutes/Bacteroidetes ratio in mice that underwent D-gal treatment, in comparison to those in the Control group. This alteration was linked to heightened levels of inflammatory markers in both serum and the hippocampus and the administration of PM significantly mitigated this effect. Additionally, D-gal treatment resulted in lower levels of SCFAs within the hippocampus and an elevation of LPS concentrations in the serum. These effects may be linked to D-gal’s disruption of gut microbiota balance, resulting in barrier impairment and heightened permeability of the BBB, which permits the LPS from specific gut bacteria to penetrate the intestinal lining and enter the bloodstream, thereby exacerbating BBB compromise and enter the brain to exacerbate the inflammatory response. This microbial imbalance reduces the population of beneficial bacteria that synthesize SCFAs leading to diminished SCFAs levels available in the hippocampus via circulation and consequently inhibiting the cAMP signaling pathway. In conclusion, PM has similar effects to probiotic preparations. By restoring the gut microbiota, inhibiting the NF - κB pathway, and activating the cAMP signaling pathway, PM alleviates cognitive impairment CI. This study has limitations. Mechanistic experiments did not directly compare PM with individual PT or MO, precluding clear distinction between synergistic and additive effects. The long - term impacts of PM are unassessed, and 16S rRNA sequencing limitations may bias gut microbiota and CI relationship analysis. Future research should extend the observation, and use multi-omics and more techniques to analyze PM’s mechanisms. To validate the synergistic effects and explore PM’s therapeutic potential in humans, experiments are needed to clarify the contributions of PT and MO, along with cellular assays and small-scale clinical trials.
Conclusion
In this research, we showed that PM outperforms both PT and MO individually in enhancing CI. PM notably reduced the deterioration of learning and memory, neuronal injury, inflammation, and oxidative stress caused by D-gal. Our results suggest that PM mitigates CI by stimulating the cAMP-PKA and NF-κB signaling cascades, reinstating the integrity of the BBB and gastrointestinal barrier, as well as adjusting gut microbiota.
Abbreviations
AD, Alzheimer’s disease; BBB, blood-brain barrier; CI, Cognitive impairment; cAMP, cyclic Adenosine Monophosphate; D-gal, D-galactose; ELISA, Enzyme-Linked Immunosorbent Assay; GPCRs, G-protein-coupled receptors; GSH, glutathione; H&E, Hematoxylin and Eosin; IF, Immunofluorescence; IFN-γ, Interferon-γ; IL-1β, interleukin-1β; IL-6, interleukin-6; LEFSE, Linear discriminant analysis effect size; LPS, Lipopolysaccharide; MWM, Morris Water Maze; NF-κB, Nuclear factor kappa-B; ANOVA, one-way analysis of variance; OTU, operational taxonomic unit; PAT, Passive avoidance test; CREB, p-cyclic Adenosine Monophosphate Response Element Binding; PCoA, principal co-ordinates analysis; PKA, Protein Kinase A; ROS, reactive oxygen species; SCFAs, short-chain fatty acids; SOD, superoxide dismutase; TJs, tight junctions; TEM, Transmission Electron Microscopy; TNF-α, tumor necrosis factor-α; UPLC-MS/MS, ultra-high performance liquid chromatography-tandem triple quadrupole mass Spectrometry; WB, Western blotting; ZO-1, Zonula occludens protein 1; Aβ, β-protein.
Data Sharing Statement
The datasets PRJNA1191803 for this study can be found in the National Center for Biotechnology Information (NCBI). https://www.ncbi.nlm.nih.gov/sra/PRJNA1191803.
Funding
This research was funded by the National Natural Science Foundation of China (82003965), Chengdu University of Traditional Chinese Medicine - Xinglin Scholar Program (QJRC2022033), Sichuan Provincial Administration of Traditional Chinese Medicine Project (2024MS167), The Seventh Batch of National Academic Experience Inheritance Project for Old Chinese Medicine Experts by the National Administration of Traditional Chinese Medicine {Guo Zhong Yi Yao Ren Jiao Han (2022) No. 76}, National-level project of the College Student Innovation and Entrepreneurship Training Program (202310633028) and Provincial-level project of the College Student Innovation and Entrepreneurship Training Program (S202410633056).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Inoue S, Nakanishi N, Amaya F, et al. Post‐intensive care syndrome: recent advances and future directions. Acute Med Surg. 2024;11(1):e929. doi:10.1002/ams2.929
2. Zhou L, Huang X, Wang J, Wang F, Liu J, Liu N. The influence of eight cognitive training regimes upon cognitive screening tool performance in post-stroke survivors: a network meta-analysis. Front Aging Neurosci. 2024;16:1374546. doi:10.3389/fnagi.2024.1374546
3. Duan D, Dong Y, Zhang H, et al. Empty-nest-related psychological distress is associated with progression of brain white matter lesions and cognitive impairment in the elderly. Sci Rep. 2017;7(1):43816. doi:10.1038/srep43816
4. Hung L-P, Huang W, Shih J-Y, Liu C-L. A novel IoT based positioning and shadowing system for dementia training. Int J Environ Res Public Health. 2021;18(4):1610. doi:10.3390/ijerph18041610
5. Li YX, Li JC, Tian M, et al. Efficacy and safety of dengyinnaotong capsule in patients with cognitive impairment caused by cerebral small vessel Disease: study protocol of a multicenter, randomized, open-label, controlled trial (De-CSVD trial). Trials. 2022;23(1):676. doi:10.1186/s13063-022-06646-6
6. World Health Organization. Dementia. World Health Organization. 2023. Available from: https://www.who.int/news-room/fact-sheets/detail/dementia. Accessed March 6, 2025.
7. Ding G, Zhao X, Wang Y, et al. Evaluation of the relationship between cognitive impairment and suboptimal health status in a northern Chinese population: a cross-sectional study. J Glob Health. 2020;10(1). doi:10.7189/jogh.10.010804.
8. la Monte SM D. Insulin resistance and neurodegeneration: progress towards the development of new therapeutics for Alzheimer’s disease. Drugs. 2017;77:47–65. doi:10.1007/s40265-016-0674-0
9. Le PG, Choi SH, Cho S. Alzheimer’s disease biomarker detection using field effect transistor-based biosensor. Biosensors. 2023;13(11):987. doi:10.3390/bios13110987
10. Livingston G, Huntley J, Liu KY, et al. Dementia prevention, intervention, and care: 2024 report of the Lancet standing Commission. Lancet. 2024;404(10452):572–628. doi:10.1016/S0140-6736(24)01296-0
11. Singh M, Agarwal V, Pancham P, et al. A comprehensive review and androgen deprivation therapy and its impact on alzheimer’s disease risk in older men with prostate cancer. Degener Neurol Neuromuscul Dis. 2024;14:33–46. doi:10.2147/dnnd.S445130
12. Ramakrishna K, Nalla LV, Naresh D, et al. WNT-β catenin signaling as a potential therapeutic target for neurodegenerative diseases: current status and future perspective. Diseases. 2023;11(3):89. doi:10.3390/diseases11030089
13. Ren S, Wang X, Qin J, et al. Altered gut microbiota correlates with cognitive impairment in Chinese children with Down’s syndrome. Eur Child Adolesc Psych. 2022;31(1):189–202. doi:10.1007/s00787-021-01799-2
14. Wang M, Cao J, Gong C, Amakye WK, Yao M, Ren J. Exploring the microbiota-Alzheimer’s disease linkage using short-term antibiotic treatment followed by fecal microbiota transplantation. Brain Behav Immun. 2021;96:227–238. doi:10.1016/j.bbi.2021.06.003
15. Bonfili L, Cecarini V, Berardi S, et al. Microbiota modulation counteracts Alzheimer’s disease progression influencing neuronal proteolysis and gut hormones plasma levels. Sci Rep. 2017;7(1):2426. doi:10.1038/s41598-017-02587-2
16. Liang X, Fu Y, Wt C, et al. Gut microbiome, cognitive function and brain structure: a multi-omics integration analysis. Transl Neurodegener. 2022;11(1):49. doi:10.1186/s40035-022-00323-z
17. Singh N, Singh V, Rai SN, Mishra V, Vamanu E, Singh MP. Deciphering the gut microbiome in neurodegenerative diseases and metagenomic approaches for characterization of gut microbes. Biomed Pharmacother. 2022;156:113958. doi:10.1016/j.biopha.2022.113958
18. Feng H, Hu X, Lin Y, et al. Dexmedetomidine attenuates acute stress-impaired learning and memory in mice by maintaining the homeostasis of intestinal flora. Eur J Med Res. 2024;29(1):271. doi:10.1186/s40001-024-01832-5
19. Li Y, Peng Y, Shen Y, Zhang Y, Liu L, Yang X. Dietary polyphenols: regulate the advanced glycation end products-RAGE axis and the microbiota-gut-brain axis to prevent neurodegenerative diseases. Crit Rev Food Sci Nutr. 2023;63(29):9816–9842. doi:10.1080/10408398.2022.2076064
20. Pinacho-Guendulain B, Montiel-Castro AJ, Ramos-Fernández G, Pacheco-López G. Social complexity as a driving force of gut microbiota exchange among conspecific hosts in non-human primates. Front Integr Neurosci. 2022;16:876849. doi:10.3389/fnint.2022.876849
21. Plassmann H, Schelski DS, Simon MC, Koban L. How we decide what to eat: toward an interdisciplinary model of gut–brain interactions. Wiley Interdiscip Rev Cogn Sci. 2022;13(1):e1562. doi:10.1002/wcs.1562
22. Cheatham CL, Nieman DC, Neilson AP, Lila MA. Enhancing the cognitive effects of flavonoids with physical activity: is there a case for the gut microbiome? Front Neurosci. 2022;16:833202. doi:10.3389/fnins.2022.833202
23. Guerrero A, De Strooper B, Arancibia-Cárcamo IL. Cellular senescence at the crossroads of inflammation and Alzheimer’s disease. Trends Neurosci. 2021;44(9):714–727. doi:10.1016/j.tins.2021.06.007
24. Guimarães V, Sousa I, de Bruin ED, Pais J, Correia MV. Minding your steps: a cross-sectional pilot study using foot-worn inertial sensors and dual-task gait analysis to assess the cognitive status of older adults with mobility limitations. BMC Geriatr. 2023;23(1):329. doi:10.1186/s12877-023-04042-6
25. Hurden A, Cotter I, Mowszowski L, Naismith S, Anderson CS. Perspectives of general practitioners and memory clinic patients on ageing and cognitive decline to inform the design of a decentralised antihypertensive dementia prevention trial. Cereb Circ Cogn Behav. 2024;6:100215. doi:10.1016/j.cccb.2024.100215
26. Zeng J, Zhang X, Wang J, Cheng X, Zhang Y, Zhou W. Comparison of donepezil, memantine, melatonin, and liuwei dihuang decoction on behavioral and immune endocrine responses of aged senescence-accelerated mouse resistant 1 mice. Front Pharmacol. 2020;11:350. doi:10.3389/fphar.2020.00350
27. Ramakrishna K, Karuturi P, Siakabinga Q, et al. Indole-3 carbinol and diindolylmethane mitigated β-amyloid-induced neurotoxicity and acetylcholinesterase enzyme activity: in silico, in vitro, and network pharmacology study. Diseases. 2024;12(8):184. doi:10.3390/diseases12080184
28. Tripathi PN, Lodhi A, Rai SN, et al. Review of pharmacotherapeutic targets in alzheimer’s disease and its management using traditional medicinal plants. Degener Neurol Neuromuscul Dis. 2024;14:47–74. doi:10.2147/dnnd.S452009
29. Tan M-S, Yu J-T, Tan -C-C, et al. Efficacy and adverse effects of ginkgo biloba for cognitive impairment and dementia: a systematic review and meta-analysis. J Alzheimers Dis. 2015;43(2):589–603. doi:10.3233/JAD-140837
30. Zheng W, Xiang YQ, Li XB, et al. Adjunctive huperzine A for cognitive deficits in schizophrenia: a systematic review and meta‐analysis. Hum Psychopharmacol Clin Exp. 2016;31(4):286–295. doi:10.1002/hup.2537
31. Moreira J, Machado M, Dias-Teixeira M, Ferraz R, Delerue-Matos C, Grosso C. The neuroprotective effect of traditional Chinese medicinal plants—A critical review. Acta Pharm Sin B. 2023;13(8):3208–3237. doi:10.1016/j.apsb.2023.06.009
32. Li C, Gao F, Qu Y, Zhao P, Wang X, Zhu G. Tenuifolin in the prevention of Alzheimer’s disease‐like phenotypes: investigation of the mechanisms from the perspectives of calpain system, ferroptosis, and apoptosis. Phytother Res. 2023;37(10):4621–4638. doi:10.1002/ptr.7930
33. Wang L, Jin GF, Yu HH, et al. Protective effects of tenuifolin isolated from Polygala tenuifolia Willd roots on neuronal apoptosis and learning and memory deficits in mice with Alzheimer’s disease. Food Funct. 2019;10(11):7453–7460. doi:10.1039/C9FO00994A
34. Kuboyama T, Yang X, Tohda C. Natural medicines and their underlying mechanisms of prevention and recovery from amyloid β-induced axonal degeneration in Alzheimer’s Disease. Int J mol Sci. 2020;21(13):4665. doi:10.3390/ijms21134665
35. Jiang N, Zhang Y, Yao C, et al. Tenuifolin ameliorates the sleep deprivation‐induced cognitive deficits. Phytother Res. 2023;37(2):464–476. doi:10.1002/ptr.7627
36. Cao Q, Jiang Y, Cui S-Y, et al. Tenuifolin, a saponin derived from Radix Polygalae, exhibits sleep-enhancing effects in mice. Phytomedicine. 2016;23(14):1797–1805. doi:10.1016/j.phymed.2016.10.015
37. Chen S, Jia J. Tenuifolin attenuates amyloid-β 42-induced neuroinflammation in microglia through the NF-κB signaling pathway. J Alzheimers Dis. 2020;76(1):195–205. doi:10.3233/JAD-200077
38. Wang X, Fu S, Zhang C, et al. gastroprotective effect of ethanol extracts from bark of magnolia officinalis on ethanol‐induced gastric mucosal damage in rats. Biomed Res Int. 2021;2021(1):6688414. doi:10.1155/2021/6688414
39. Zhang M, Shen Y. Research progress on pharmacological effects and mechanisms of Magnolia officinalis and its active ingredients in the prevention and treatment of Alzheimer ‘s disease and Parkinson ‘s disease. Drug Evaluation Study. 2023;46(09):2054–2060.
40. Zhang Z, Shu X, Cao Q, et al. Compound from Magnolia officinalis ameliorates white matter injury by promoting oligodendrocyte maturation in chronic cerebral ischemia models. Neurosci Bull. 2023;39(10):1497–1511. doi:10.1007/s12264-023-01068-z
41. Wang X, Jia J. Magnolol improves Alzheimer’s disease-like pathologies and cognitive decline by promoting autophagy through activation of the AMPK/mTOR/ULK1 pathway. Biomed Pharmacother. 2023;161:114473. doi:10.1016/j.biopha.2023.114473
42. Li H, Jia J, Wang W, et al. Honokiol alleviates cognitive deficits of Alzheimer’s disease (PS1 V97L) transgenic mice by activating mitochondrial SIRT3. J Alzheimers Dis. 2018;64(1):291–302. doi:10.3233/JAD-180126
43. Xue M. An overview of the etiology and treatment of stroke. Shaanxi Tradit Chin Med. 1980;(06):20–23.
44. Sun S. Qianjin Yaofang. Art China Network; 1955.
45. Wang T. Wai Tai Mi Yao. People’s Medical Publishing House; 1955.
46. Jiang J, Jian W, Da L, Liu L, Wu X, Ma X. Effects of radix polygalae and magnolia of different decoction time and compatibility proportion on mouse gastrointestinal motility. J Xi’an Jiaotong Univ. 2009;(3):380.
47. Fu Y, Xiao W, Wang J, Liu L, Wang D, Wang X. The effects of the combination of magnolia officinalis and polygala tenuifolia on NO in gastrointestinal tissues and vasoactive intestinal peptide in the gastric antrum. Food Med. 2011;13(11):392–395.
48. Huang L. Study of Mechanisms That Magnolia Officinalis Alleviates the Side Effects of Polygala Tenuifolia Base on Method of Pharmacokinetics. Chengdu University of Traditional Chinese Medicine; 2015.
49. Liu S, Yang D, Li W, et al. Magnolia officinalis alcohol extract alleviates the intestinal injury induced by polygala tenuifolia through regulating the PI3K/AKT/NF-κB signaling pathway and intestinal flora. Drug Des Devel Ther. 2024;Volume 18:1695–1710. doi:10.2147/DDDT.S461152
50. Ma X, Wang J, Huang C, et al. Study on the sedative and expectorant effects of radix polygonae radix decoctum prepared by thick park. Pharmacol Clin Chin Med. 2013;29(1):90–93.
51. Committee NP. Pharmacopoeia of the People’s Republic of China.
52. Ma R, Xie Q, Wang J, et al. Metabolomics study on the effects of honokiol and yuanzhi combined ethanol extracts on urinary metabolites. J.Pharm.Pharmacol. 2019.
53. Wei W, Wu X, Li Y. Pharmacological Experimental Methodology.
54. Yang Z, Le TD, Simovic MO, et al. Traumatized triad of complementopathy, endotheliopathy, and coagulopathy˗ Impact on clinical outcomes in severe polytrauma patients. Front Immunol. 2022;13:991048. doi:10.3389/fimmu.2022.991048
55. Jiang J. Research on extraction methods and proportions of magnolia officinalis for alleviating gastrointestinal adverse reactions caused by polygala tenuifolia. Master. 2009.
56. Li D. A preliminary study on the comparison of chemical components before and after the compatibility of polygala tenuifolia and magnolia officinalis and the preliminary exploration of the detoxification mechanism. 2009.
57. Ma X. Research on the Effects of the Compatibility of Magnolia Officinalis and Polygala Tenuifolia on Gastrointestinal Electrophysiology and Isolated Gastrointestinal Smooth Muscle. Master. Chengdu University of Traditional Chinese Medicine; 2011.
58. Li Z, Li YY, Zhang J, et al. Serum metabolomics combined with gut microbiota reveals the effects of Polygala tenuifolia polysaccharide on the metabolic and microbial profiles in SAMP8 mouse. J Pharm Biomed Anal. 2024:251116442. doi:10.1016/j.jpba.2024.116442
59. Xue C, He Z, Zeng M, et al. The protective effects of polygala tenuifolia and tenuifolin on corticosterone-evoked ferroptosis, oxidative stress, and neuroinflammation: insights from molecular dynamics simulations and in vitro experiments. Foods. 2024;13(21):3358. doi:10.3390/foods13213358
60. Ali Q, Ma S, Farooq U, et al. Pasture intake protects against commercial diet-induced lipopolysaccharide production facilitated by gut microbiota through activating intestinal alkaline phosphatase enzyme in meat geese. Front Immunol. 2022;13:1041070. doi:10.3389/fimmu.2022.1041070
61. Chen F, Zhang H, Du EC, et al. Supplemental magnolol or honokiol attenuates adverse effects in broilers infected with Salmonella pullorum by modulating mucosal gene expression and the gut microbiota. J Anim Sci Biotechnol. 2021;12(1):87. doi:10.1186/s40104-021-00611-0
62. Mo JH, Xiang J, Li J, et al. Natural magnolol ameliorates coccidiosis infected with Eimeria tenella by affecting antioxidant, anti-inflammatory, and gut microbiota of chicks. Poultry Sci. 2023;102(11):102975. doi:10.1016/j.psj.2023.102975
63. Niu L, Hou YY, Jiang M, Bai G. The rich pharmacological activities of Magnolia officinalis and secondary effects based on significant intestinal contributions. J Ethnopharmacol. 2021;281114524. doi:10.1016/j.jep.2021.114524
64. Zhai T, Wang JJ, Chen Y. Honokiol affects the composition of gut microbiota and the metabolism of lipid and bile acid in methionine-choline deficiency diet-induced NASH mice. Sci Rep. 2023;13(1):15203. doi:10.1038/s41598-023-42358-w
65. Wu Y, Wu Y, Yang Y, et al. Lysyl oxidase-like 2 inhibitor rescues D-galactose-induced skeletal muscle fibrosis. Aging Cell. 2022;21(7):e13659. doi:10.1111/acel.13659
66. Li P, Ma Y, Wang X, et al. The protective effect of PL 1-3 on D-galactose-induced aging mice. Front Pharmacol. 2023;14:1304801. doi:10.3389/fphar.2023.1304801
67. Liang Y, Chen X, Teng Z, Wang X, Yang J, Liu G. Discovery of a 4-Hydroxy-3’-trifluoromethoxy-substituted resveratrol derivative as an anti-aging agent. Molecules. 2023;29(1):86. doi:10.3390/molecules29010086
68. Lail H, Mabb AM, Parent MB, Pinheiro F, Wanders D. Effects of dietary methionine restriction on cognition in mice. Nutrients. 2023;15(23):4950. doi:10.3390/nu15234950
69. Li R, Huang Z, Luo J, Luo H, Wang W. Downregulation of the CB1-mediated endocannabinoid signaling underlies d-galactose-induced memory impairment. Front Mol Neurosci. 2020;13:130. doi:10.3389/fnmol.2020.00130
70. Jia XY, Liu HM, Yin GL, et al. Arctium lappa L. polysaccharides alleviate oxidative stress and inflammation in the liver and kidney of aging mice by regulating intestinal homeostasis. Int J Biol Macromol. 2024:280135802. doi:10.1016/j.ijbiomac.2024.135802
71. Liu N, Jiang X, Li H. The viral hypothesis in Alzheimer’s disease: SARS-CoV-2 on the cusp. Front Aging Neurosci. 2023;15:1129640. doi:10.3389/fnagi.2023.1129640
72. Song Y, Wang X, Ma W, et al. Graves’ disease as a driver of depression: a mechanistic insight. Front Endocrinol. 2023;14:1162445. doi:10.3389/fendo.2023.1162445
73. Lin LK, Li C, Zhang YJ, et al. Effects of an Akt-activating peptide obtained from walnut protein degradation on the prevention of memory impairment in mice. Food Funct. 2024;15(4):2115–2130. doi:10.1039/d3fo04479c
74. Agirman G, Yu KB, Hsiao EY. Signaling inflammation across the gut-brain axis. Science. 2021;374(6571):1087–1092. doi:10.1126/science.abi6087
75. Liu XL, Zhao YC, Zhu HY, et al. Taxifolin retards the D-galactose-induced aging process through inhibiting Nrf2-mediated oxidative stress and regulating the gut microbiota in mice. Food Funct. 2021;12(23):12142–12158. doi:10.1039/d1fo01349a
76. Sun W, Zhu J, Qin G, et al. Lonicera japonica polysaccharides alleviate D-galactose-induced oxidative stress and restore gut microbiota in ICR mice. Int J Biol Macromol. 2023;245:125517. doi:10.1016/j.ijbiomac.2023.125517
77. Li K, Li J. The research progress of interaction between the intestinal microbiota-gut-brain axis and stress. Chin J Int Med. 2020;59(3):247–249. doi:10.3760/cma.j.issn.0578-1426.2020.03.018
78. Pellegrini C, Fornai M, D’Antongiovanni V, Antonioli L, Bernardini N, Derkinderen P. The intestinal barrier in disorders of the central nervous system. review. Lancet Gastroenterol Hepatol. 2023;8(1):66–80. doi:10.1016/s2468-1253(22)00241-2
79. He Y, Huang X, Zheng W, Hao Q, Ding X. The regulatory effect of mulberry leaf alkaloids on D-galactose-induced dysbiosis in mice. Food Sci. 2022;43(5):118–126.
80. Gao Y, Liu Y, Ma F, et al. Lactobacillus plantarum Y44 alleviates oxidative stress by regulating gut microbiota and colonic barrier function in Balb/C mice with subcutaneous d-galactose injection. Food Funct. 2021;12(1):373–386. doi:10.1039/D0FO02794D
81. Ruan Q, Hu X, Ao H, et al. The neurovascular protective effects of huperzine A on D-galactose-induced inflammatory damage in the rat hippocampus. Gerontology. 2014;60(5):424–439. doi:10.1159/000358235
82. Zhao H, Li N, Wang Q, Cheng X, Li X, Liu T. Resveratrol decreases the insoluble Aβ1–42 level in hippocampus and protects the integrity of the blood–brain barrier in AD rats. Neuroscience. 2015;310:641–649. doi:10.1016/j.neuroscience.2015.10.006
83. Wang M, Fu R, Xu D, et al. Traditional Chinese Medicine: a promising strategy to regulate the imbalance of bacterial flora, impaired intestinal barrier and immune function attributed to ulcerative colitis through intestinal microecology. J Ethnopharmacol. 2024;318(Pt A):116879. doi:10.1016/j.jep.2023.116879
84. Wang M, Jia J, Cui Y, Wang Q. Advances in research on lipopolysaccharide - induced intestinal barrier proteins and their associated signaling pathways. ChinJ Animal Nutr. 2020;32(5):6.
85. Han Y, Ding L, Cheng X, et al. Hypoxia augments cerebral inflammation in a dextran sulfate sodium-induced colitis mouse model. Front Cell Neurosci. 2020;14:611764. doi:10.3389/fncel.2020.611764
86. Aktar S, Ferdousi F, Kondo S, Kagawa T, Isoda H. Transcriptomics and biochemical evidence of trigonelline ameliorating learning and memory decline in the senescence-accelerated mouse prone 8 (SAMP8) model by suppressing proinflammatory cytokines and elevating neurotransmitter release. Geroscience. 2024;46(2):1671–1691. doi:10.1007/s11357-023-00919-x
87. Chen Y, Yu Y. Tau and neuroinflammation in Alzheimer’s disease: interplay mechanisms and clinical translation. J Neuroinflammation. 2023;20(1):165. doi:10.1186/s12974-023-02853-3
88. Montagne A, Nation DA, Pa J, Sweeney MD, Toga AW, Zlokovic BV. Brain imaging of neurovascular dysfunction in Alzheimer’s disease. Review Acta Neuropathol. 2016;131(5):687–707. doi:10.1007/s00401-016-1570-0
89. Dang Q, Wu D, Li Y, et al. Walnut-derived peptides ameliorate d-galactose-induced memory impairments in a mouse model via inhibition of MMP-9-mediated blood–brain barrier disruption. Food Res Int. 2022;162:112029. doi:10.1016/j.foodres.2022.112029
90. Wei C, Jiang W, Wang R, et al. Brain endothelial GSDMD activation mediates inflammatory BBB breakdown. Nature. 2024;1–8.
91. Mz W, Jf J, Yz C, Qj W. Research progress on lipopolysaccharide-induced intestinal barrier signaling pathway. 2020.
92. Zhao Y, Lukiw WJ. Bacteroidetes neurotoxins and inflammatory neurodegeneration. review. mol Neurobiol. 2018;55(12):9100–9107. doi:10.1007/s12035-018-1015-y
93. Li Y, Zhou F, You J, Gong X. Annexin A1 mitigates blood-brain barrier disruption in a sepsis-associated encephalopathy model by enhancing the expression of occludin and zonula occludens-1 (ZO-1). CNS Neurosci Ther. 2024;30(12):e70173. doi:10.1111/cns.70173
94. Lu Q, Yu A, Pu J, et al. Post-stroke cognitive impairment: exploring molecular mechanisms and omics biomarkers for early identification and intervention. Front Mol Neurosci. 2024;17:1375973. doi:10.3389/fnmol.2024.1375973
95. Zhu L, Xing J, j W, w W, Zhang A. Research progress on the mechanisms of short-chain fatty acids in neurodegenerative diseases. Chin Gen Pract. 2023;26(24):3061–3066.
96. Sebastiao I, Candeias E, Santos MS, De oliveira CR, Moreira PI, Duarte AI. Insulin as a bridge between type 2 diabetes and Alzheimer disease - how anti-diabetics could be a solution for dementia. Front Endocrinol. 2014;5110. doi:10.3389/fendo.2014.00110
97. Luo T, Che Q, Guo Z, Song T, Zhao J, Xu D. Modulatory effects of traditional Chinese medicines on gut microbiota and the microbiota-gut-x axis. Front Pharmacol. 2024;15:1442854. doi:10.3389/fphar.2024.1442854
98. Chen Y, Xu J, Chen Y. Regulation of neurotransmitters by the gut microbiota and effects on cognition in neurological disorders. Nutrients. 2021;13(6). doi:10.3390/nu13062099
99. Yang B, Zhang Z, Yang Z, et al. Chanling gao attenuates bone cancer pain in rats by the IKKβ/NF-κB signaling pathway. Front Pharmacol. 2020;11:525. doi:10.3389/fphar.2020.00525
100. Nakano-Kobayashi A, Canela A, Yoshihara T, Hagiwara M. Astrocyte-targeting therapy rescues cognitive impairment caused by neuroinflammation via the Nrf2 pathway. Proc Natl Acad Sci USA. 2023;120(33):e2303809120. doi:10.1073/pnas.2303809120
101. Wang L, Wu C, Peng W, et al. A high-performance genetically encoded fluorescent indicator for in vivo cAMP imaging. Nat Commun. 2022;13(1):5363. doi:10.1038/s41467-022-32994-7
102. Gu X, Quan J, Guo Y, Li S. The role of cAMP response element-binding protein in regulating the differentiation of human apical papilla stem cells via transforming growth factor β1. Prevention Treat Oral Dis. 2018;26(7):428–433.
103. Lj X, Deng T, Ying X, Sun T, Js LI. Research progress and countermeasures of Alzheimer’s disease signaling pathways. Chin Pharm J. 2021;37(2):165–170.
104. Chen F, Wang Y, Xiang Q. The effect of gastrodin on cognitive impairment in alzheimer’s disease rats via the cAMP/PKA/CREB signaling pathway. Chin J Geriatr. 2024.
105. Guo H, Cheng Y, Wang C, et al. FFPM, a PDE4 inhibitor, reverses learning and memory deficits in APP/PS1 transgenic mice via cAMP/PKA/CREB signaling and anti-inflammatory effects. Neuropharmacology. 2017;116:260–269. doi:10.1016/j.neuropharm.2017.01.004
106. Vamanu E, Rai SN. The link between obesity, microbiota dysbiosis, and neurodegenerative pathogenesis. Diseases. 2021;9(3):45. doi:10.3390/diseases9030045
107. Yang XQ, Yu DK, Xue L, Li H, Du JR. Probiotics modulate the microbiota-gut-brain axis and improve memory deficits in aged SAMP8 mice. Acta Pharm Sin B. 2020;10(3):475–487. doi:10.1016/j.apsb.2019.07.001
108. Sorboni SG, Moghaddam HS, Jafarzadeh-Esfehani R, Soleimanpour S. A comprehensive review on the role of the gut microbiome in human neurological disorders. Clin Microbiol Rev. 2022;35(1):e00338–20. doi:10.1128/cmr.00338-20
109. Chen J, Chen X, Zhang Y, et al. Bioactivity and influence on colonic microbiota of polyphenols from noni (Morinda citrifolia L.) fruit under simulated gastrointestinal digestion. Food Chem. 2024;21:101076.
110. Wei S, Bahl MI, Baunwall SMD, Hvas CL, Licht TR. Determining gut microbial dysbiosis: a review of applied indexes for assessment of intestinal microbiota imbalances. Appl Environ Microbiol. 2021;87(11):e00395–21. doi:10.1128/AEM.00395-21
111. Ramos-Romero S, Santocildes G, Piñol-Piñol D, et al. Implication of gut microbiota in the physiology of rats intermittently exposed to cold and hypobaric hypoxia. PLoS One. 2020;15(11):e0240686. doi:10.1371/journal.pone.0240686
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms.php
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms.php
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.