")
Back to Journals » Journal of Inflammation Research » Volume 18
IL-33 Participates in G. Vaginalis-Induced Bacterial Vaginosis: Involvement of Intravaginal IgA
Authors Zhou M, Zhou L, Liu J, Yu S
Received 20 February 2025
Accepted for publication 6 June 2025
Published 17 June 2025 Volume 2025:18 Pages 8005—8013
DOI https://doi.org/10.2147/JIR.S523880
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Dr Tara Strutt
Min Zhou,1 Lili Zhou,2 Junbo Liu,1 Shaohui Yu1
1Department of Gynecology, Changchun University of Chinese Medicine Affiliated Hospital, Changchun, Jilin, People’s Republic of China; 2Department of Obstetrics, Changchun University of Chinese Medicine Affiliated Hospital, Changchun, Jilin, People’s Republic of China
Correspondence: Shaohui Yu, Department of Gynecology, Changchun University of Chinese Medicine Affiliated Hospital, Changchun, Jilin, 130000, People’s Republic of China, Email [email protected]
Introduction: Bacterial vaginosis (BV) is a common gynecological disease characterized by an abnormal increase in vaginal secretions, odor and itching. The pathogenesis of BV is not fully understood, but it is believed that the disruption of the mucosal immune system plays a key role. We investigated the role of IL-33 in preventing BV and explored the mechanism by which IL-33 regulates intravaginal IgA.
Methods: Protein levels of IL-33 and IgA, and the pH value of vaginal secretions in healthy donors and patients with BV (14 vs 14) were determined by ELISA. G. vaginalis-induced bacterial vaginosis mouse model was established using wild-type (WT) and IL-33 knockout (KO) mice. Protein levels of IL-33, IgA and TGF-β, the pH value of vaginal secretions, and Gram-staining were measured in vivo and in vitro to investigate the role of IL-33 in BV progression.
Results: IL-33 and IgA were significantly decreased in vaginal secretions of patients with BV. IL-33 deficiency aggravated BV induced by G. vaginalis in a mouse model, while IL-33 supplementation prevented it. IL-33 modulated intravaginal IgA expression through the TGF-β signaling pathway in B cells.
Conclusion: IL-33 prevents G. vaginalis-induced BV by modulating intravaginal IgA expression through the TGF-β signaling pathway in B cells.
Keywords: bacterial vaginosis, G. vaginalis, IL-33, IgA, TGF-β
Introduction
Bacterial vaginosis (BV) is a common gynecological disease characterized by an abnormal increase in vaginal secretions, odor and itching.1,2 The general population prevalence of BV is high globally, although it varies significantly across regions and population.3 Vaginal secretions are usually grayish-white or yellow-green, thin in texture, and sometimes accompanied by foamy or cheesy secretions. The odor is usually fishy or putrid, especially after sexual intercourse.4 Itching and a burning sensation are also common symptoms of BV.5 The etiology of BV is complex, and mainly involves vaginal microecological imbalance, sexual behavior, hormonal level changes, immune system decline and other factors.6,7
BV is polymicrobial, involving a mix of bacteria. The most commonly associated pathogens include Gardnerella vaginalis (G. vaginalis) (most commonly implicated), Atopobium vaginae, etc. G. vaginalis, often considered the primary initiator of the microbial shift, is present in over 90% of BV cases. If left untreated, bacterial vaginosis—particularly when associated with G. vaginalis—can lead to several complications, including obstetric complications, gynecologic and surgical complications, increased susceptibility to sexually transmitted infections, and recurrence and antibiotic resistance.5,8 Vaginal microecological imbalance is one of the main causes of bacterial vaginosis. Under normal circumstances, there are many beneficial bacteria such as lactobacilli in the vagina, which can inhibit the growth of other pathogenic microorganisms.9 However, when certain factors (such as the use of antibiotics, changes in hormonal levels,10 sexual activity, etc). affect the number and types of beneficial bacteria, it can lead to an imbalance of vaginal flora and thus cause BV.5,11 Immune system decline is also an important cause of BV. When the body’s immune system declines, its resistance to bacteria decreases, making it easier for pathogenic bacteria to grow in the vagina and cause vaginitis.12,13
IL-33 and IgA are two molecules that have received much attention in recent years in the study of BV. IL-33 is a pro-inflammatory factor that can regulate the body’s immune response, while IgA is an immunoglobulin that is mainly present on mucosal surfaces and has important immune defense functions.14–16 Studies have shown that the functions of IL-33 and IgA are different in BV. The expression of IL-33 is significantly reduced in patients with BV, and it participates in the occurrence and development of BV.17 The level of IgA is usually high in the vagina of healthy women and can play a role in resisting invasion by pathogenic microorganisms. However, in patients with bacterial vaginosis, the level of IgA is usually decreased, which may be related to the imbalance of vaginal microecology and immune suppression.18,19 In addition, some studies have found that increasing the level of IgA can improve the symptoms of BV and prevent recurrence.12
However, the regulatory relationship between IL-33 and IgA in the context of BV remains poorly understood. This gap limits our ability to develop targeted immunotherapies for prevention and treatment. Therefore, this study aims to investigate the mechanistic role of IL-33 in modulating IgA production through TGF-β signaling during BV progression. We hypothesize that IL-33 exerts a protective effect in BV by enhancing TGF-β-mediated IgA production, thereby restoring mucosal immunity and contributing to the resolution of infection.
Materials And Methods
Patient Cohort
We recruited premenopausal women aged 18 to 55 years who gave written informed consent to participate in the study (14 women in each group). At the screening visit, we collected participants’ medical history and performed physical and pelvic examinations. We also obtained a vaginal swab from women who met at least three of the four Amsel criteria for BV: thin, white, or yellow, homogeneous discharge; clue cells >20% on microscopic examination; vaginal pH >4.5; and fishy odor after adding 10% potassium hydroxide to a vaginal specimen. We used Gram staining to determine the Nugent score for BV (0 to 3: normal; 4 to 6: intermediate; 7 to 10: positive). We tested blood samples for HIV and syphilis, vaginal swabs for gonorrhea, chlamydia, and trichomonas infections, and urine samples for beta human chorionic gonadotropin and urinary tract infection. Participants were excluded if they: 1) Did not meet at least three of the four Amsel criteria for BV. 2) Had a Nugent score <7, indicating either a normal or intermediate vaginal flora. 3) Tested positive for other infections: HIV, syphilis, gonorrhea, chlamydia, trichomonas, urinary tract infection. 4) Were pregnant, as indicated by urine beta-hCG testing. Detection of G. vaginalis was inferred via Amsel and Nugent scores, and further direct method by culture. The study was approved by the Changchun University of Chinese Medicine Affiliated Hospital (#2023-CC-254), and this study was performed in accordance with the Declaration of Helsinki, Ethical Principles for Medical Research Involving Human Subjects.
Mice
We obtained wild-type (WT) mice on a C57BL/6 background and IL-33 knockout (KO) mice on an FVB/N background from GemPharmatech (Nanjing, China). The IL-33 KO mice were generated by microinjecting FVB/N zygotes with a targeting vector, and IL-33 allele was disrupted by homologous recombination. We confirmed the genotypes of the mice by PCR using the following primers: Il33 (primer1: CAGCCTCAGATTTCTCTGTGC-3, primer2: TCAGGTTTCTGTGGGATTGA, primer3: TGTCAACAATGATGCACTGG). All mice were 8 weeks old and housed at 22 °C and 50% humidity with a 12/12 h light/dark cycle (07:00–19:00) and given ad libitum access to a standard rodent chow diet and water. All studies were followed the protocols approved by the Institutional Animal Care and Use Committee of Changchun University of Chinese Medicine Affiliated Hospital (#2023/06/22-6). Gardnerella vaginalis ATCC 14018 (G. vaginalis) was cultured on Columbia Blood Agar Base Medium under anaerobic conditions in sealed jars at 37 °C for 36 h. We harvested the cultured GV and resuspended it in PBS to a density of 3×109 CFU mL−1. To induce BV in mice, we inoculated the vaginas of female mice with 3×109 CFU mL−1 of GV for 8 days according to the published methods.20 In particularly, the mice received intraperitoneal injections of 0.5 mg β-estradiol dissolved in 100 µL of filter-sterilized sesame oil, administered both three days before and on the day of bacterial inoculation. All outcome measurements (pH, cytokine, and immunoglobulin levels) were performed at Day 8 post-infection to evaluate chronic BV pathology.
Sample Collection and Processing
For human samples, vaginal secretions were collected from the participants using sterile cotton swabs. The swabs were inserted into the vaginal canal and gently rotated against the vaginal wall to absorb secretions. The collected swabs were immediately placed into sterile collection tubes containing 1 mL of sterile phosphate-buffered saline (PBS, pH 7.4), vortexed briefly, and centrifuged at 500 × g for 5 minutes at 4°C to pellet debris. The supernatants were then aliquoted and stored at −80°C until analysis.
For mouse models, vaginal secretions were obtained by flushing the vaginal cavity with 50 μL of sterile PBS using a pipette tip. The lavage fluid was collected into sterile microcentrifuge tubes, briefly centrifuged at 500 × g for 5 minutes at 4°C to remove debris, and the supernatants were stored at −80°C until further use.
pH Measurement
The pH of vaginal secretions was measured immediately after collection to minimize alterations due to environmental exposure. A micro-pH electrode (Thermo Fisher Scientific) was calibrated using standard pH buffers (pH 4.0, 7.0, and 10.0) prior to each measurement session. For each sample, approximately 50–100 μL of vaginal secretion (or supernatant) was placed in a sterile microtube or well of a 96-well plate, and the pH was measured by gently inserting the probe into the sample. Care was taken to avoid contamination between samples by rinsing the electrode with distilled water and blotting dry with lint-free tissue between measurements. All measurements were performed in triplicate, and the average value was used for statistical analysis.
Gram Stain Methodology
Vaginal smears were air-dried, fixed with methanol, and stained using a standard Gram-staining kit (Sigma-Aldrich). Slides were imaged under a 100× oil-immersion objective (Nikon Eclipse Ci-L).
Reverse Transcription-Quantitative Polymerase Chain Reaction (RT-qPCR)
Tissue samples were homogenized using TissueLyzer LT (Qiagen, Hilden, Germany). Total RNA from tissue samples was extracted using QIAzol reagent (Qiagen, Hilden, Germany) and purified with RNA spin columns (Invitrogen, Waltham, USA). Complementary DNA (cDNA) synthesis was carried out with a High-Capacity cDNA Reverse Transcription kit (Thermo Fisher Scientific, Waltham, USA). The resultant cDNA was analyzed by RT-qPCR using the iCycler Sequence Detection System (Bio-Rad, Hercules, USA) and iTaq Universal SYBR Green Supermix (Bio-Rad, Hercules, USA). Relative mRNA levels were calculated by the ΔΔCt method and normalized to Actb mRNA.
ELISA Assay
We measured the levels of IL-33 and IgA in vaginal swab samples using ELISA kits. We resuspended the samples in 1 mL of PBS and assayed them with human IL-33 (BMS2048, Invitrogen), human IgA (BMS2096, Invitrogen), mouse IL-33 (88–7333-88, Invitrogen), and mouse IgA (EMIGA, Invitrogen) kits, following the manufacturers’ instructions for each kit.
Flow Cytometry
Single-cell suspensions from vaginal swabs were prepared by vortexing swabs in 1 mL PBS followed by centrifugation (300 × g, 5 min). To dissociate epithelial cells and mucus, samples were treated with 1 mg/mL collagenase D (Roche) and 0.1 mg/mL DNase I (Sigma) for 20 min at 37°C. Cells were filtered through a 70-µm strainer, washed, and stained with fluorescently labeled antibodies against IgA (C10-3, BD Biosciences) and CD19 (1D3; eBioscience). For analysis, 10,000 live cells (identified by forward/side scatter and 7-AAD exclusion) were acquired per sample using a BD FACSLyric system (Becton, Dickinson and Company, Franklin Lakes), as described previously.21 The average cell yield from vaginal swabs was 2.5 × 104 ± 0.8 × 104 cells per mouse (mean ± SD). IgA+CD19+ B cells accounted for 8.5% ± 1.8% of total live cells in wild-type mice and 4.6% ± 0.8% in IL-33 KO mice.
Statistical Analysis
The required sample size was determined using PS software (power and sample size collection version 3.0.12). The estimated sample size was 14 for each group. Statistics were performed by Prism software (version 8.3.0). Data were shown as mean ± SD. An unpaired t test (two-tailed) was used for the comparisons between two groups. One- or two-way ANOVA with appropriate post hoc tests were used for multiple comparisons. Significance level was set when p-values were less than 0.05.
Results
IL-33 and IgA are Decreased in Vaginal Secretions of Patients with BV
To evaluate the correlation between the expression levels of mucosal immune-related factors IL-33 and IgA and the onset of BV, we first collected vaginal secretions from 14 healthy volunteers and BV patients. We found that the pH level of vaginal secretions from BV patients (5.3±0.52) was significantly higher than control (4.09±0.32), which was consistent with other previous studies.22 The protein levels of IL-33 (control: 4.19±0.58 ng/mL; BV: 1.45±0.39 ng/mL) and IgA (control: 535.9±125.0 pg/mL; BV: 219.3±79.4 pg/mL) in vaginal secretions from BV patients were significantly reduced (Table 1), suggesting that these two factors were associated with the pathogenesis of BV.
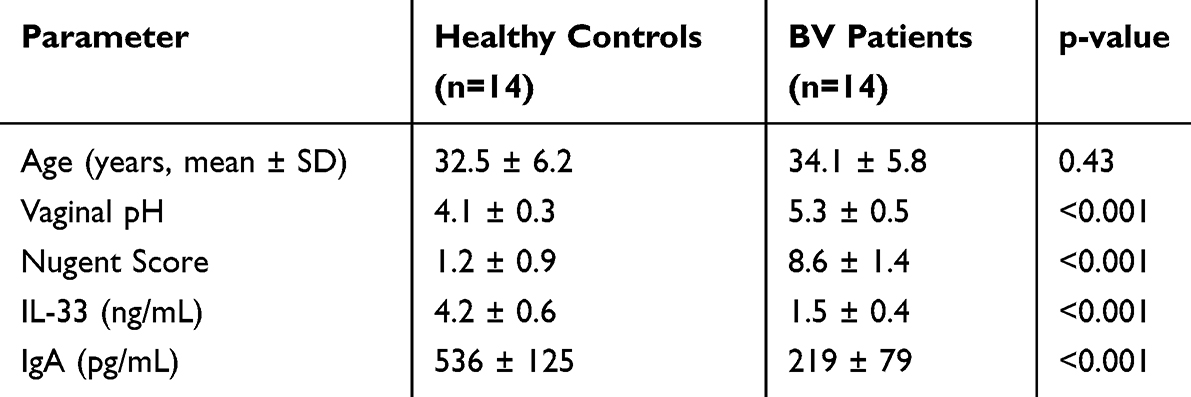 |
Table 1 Key Parameters Between the Two Cohorts. Data are Presented as Mean ± SD. Statistics Were Conducted Using Unpaired t Test. ***p < 0.005 |
IL-33 and IgA are Decreased in G. Vaginalis-Induced Bacterial Vaginosis Mouse Model
In the mouse model of BV induced by G. vaginalis, we first used Gram staining of vaginal smears to confirm that the abundance of Gram-negative bacteria increased significantly after G. vaginalis colonization compared with the untreated group (Figure 1A). The CFU of bacteria reached a peak in the first two days after colonization (Figure 1B). Consistent with clinical patients, the pH level of vaginal secretions from BV mice induced by G. vaginalis was significantly higher than mock control (Figure 1C; mock: 4.10±0.43; G. vaginalis: 5.04±0.25). The protein levels of IL-33 (mock: 7.80±1.30 ng/mL; G. vaginalis: 2.00±0.34 ng/mL) and IgA (mock: 721.0±193.8 pg/mL; G. vaginalis: 331.0±44.3 pg/mL) in vaginal secretions from BV mice induced by G. vaginalis were significantly reduced (Figure 1D and E).
 |
Figure 1 G. vaginalis-induced bacterial vaginosis model also exhibits a decrease in IL-33 and IgA. (A) Gram-stained smears of vaginal fluid collected at 2 days post-infection. (B) Time course of mean vaginal G. vaginalis titers during infection. (C) The PH value of vaginal secretions of mice infected with BV. (D and E) The protein levels of IL-33 and IgA in vaginal secretions of mice infected with BV were evaluated by ELISA assay. Data are presented as mean ± SD. Statistics were conducted using unpaired t test. **p < 0.01, ***p < 0.005. |
IL-33 Prevents the Symptoms of G. Vaginalis-Induced BV
To further explore the role of IL-33 in BV progression, we established the G. vaginalis-induced BV model using IL-33 KO mice. In the mouse model of BV induced by G. vaginalis, we found that IL-33 deficiency promoted the abundance of Gram-negative bacteria (IL-33+/+: 2.100±0.400 ng/mL; IL-33−/−: 0.005±0.011 ng/mL) (Figure 2A and B), pH value (IL-33+/+: 4.91±0.34; IL-33−/−: 5.25±0.38) (Figure 2C), G. vaginalis content (Figure 2D). IL-33 deficiency significantly reduced IgA levels (p<0.01) (Figure 2E), indicating IL-33’s essential role in maintaining mucosal IgA.
 |
Figure 2 IL-33 prevents the symptoms of G. vaginalis-induced bacterial vaginosis. (A) Gram-stained smears of vaginal fluid collected from WT and IL-33 KO mice at 2 days post-infection of G. vaginalis. (B) The protein levels of IL-33 in vaginal secretions of WT and IL-33 KO mice infected with BV were evaluated by ELISA assay. (C) The PH value of vaginal secretions of WT and IL-33 KO mice infected with BV. (D) Time course of mean vaginal G. vaginalis titers in WT and IL-33 KO mice during infection. (E) The protein levels of IgA in vaginal secretions of WT and IL-33 KO mice infected with BV were evaluated by ELISA assay. Data are presented as mean ± SD. Statistics were conducted using two-way ANOVA analysis with a Bonferroni post hoc test for the panel C, or unpaired t test for others. **p < 0.01, ***p < 0.005. |
IL-33 Enhances the Induction of IgA Through Promoting TGF-β Expression
To investigate the regulatory role of IL-33 on IgA, we compared the expression level of IgA in B cells in the vagina of WT and IL-33 KO mice. FACS results showed that IL-33 deficiency significantly decreased the proportion of IgA-positive B cells (IL-33+/+: 8.50±1.83%; IL-33−/−: 4.59±0.79%) (Figure 3A and B). Meanwhile, qPCR detection showed that the level of α-GLT and AID mRNA was also significantly reduced in intravaginal B cells isolated from G. vaginalis-exposed IL-33 KO mice compared to that in WT mice (Figure 3C). Because TGF-β is a key cytokine for inducing IgA,23 we compared the protein level of TGF-β in the mouse vagina and found that IL-33 deficiency significantly reduced TGF-β level (IL-33+/+: 346.2±61.7 pg/mL; IL-33−/−: 132.8±47.0 pg/mL) (Figure 3D). In vitro, recombinant IL-33 could significantly stimulate TGF-β production upon LPS treatment (Figure 3E). Moreover, flow cytometry analysis showed that supplementing TGF-β in IL-33 knockout mice could eliminate the difference between them and WT mice (Figure 3F and G). These results suggested that IL-33 had a significant regulatory role on TGF-β-induced IgA production by B cells.
 |
Figure 3 IL-33 enhances the induction of IgA via promoting TGF-b expression. (A and B) Frequency of IgA+ B cells that infiltrate in vagina of G. vaginalis-exposed WT and IL-33 KO mice, assessed by flow cytometry, and quantification of the left results. (C) Quantitative PCR analysis of α-GLT and AID mRNA in intravaginal B cells isolated from G. vaginalis-exposed WT and IL-33 KO mice. (D) The protein levels of TGF-b in vaginal secretions of WT and IL-33 KO mice infected with BV were evaluated by ELISA assay. (E) ELISA of TGF-b in the supernatant of BMDMs stimulated with LPS plus different doses of IL-33. (F and G) Frequency of IgA+ B cells that infiltrate in vagina of G. vaginalis-exposed WT and IL-33 KO mice which Intravaginal administration of recombinant TGF-b (10 µg/kg), assessed by flow cytometry, and quantification of the left results. Data are presented as mean ± SD. Statistics were conducted using one-way ANOVA analysis with a Tukey post hoc test for the panel E, or unpaired t test for others. *p < 0.05, **p < 0.01. |
Discussion
BV is a common vaginal disorder characterized by an imbalance of the vaginal microbiota, which can increase the risk of various reproductive and sexual health problems.2 The pathogenesis of BV is not fully understood, but it is believed that the disruption of the mucosal immune system plays a key role.24–26 In this study, we investigated the role of IL-33, a cytokine involved in mucosal immunity and inflammation,27 in preventing BV induced by G. vaginalis, a major causative agent of BV.28 We also explored the mechanism by which IL-33 regulates intravaginal IgA, an antibody that can protect against bacterial infection.29 Here, we demonstrated that IL-33 deficiency aggravated BV induced by G. vaginalis in a mouse model, while IL-33 supplementation prevented it. We also found that IL-33 modulated intravaginal IgA expression through TGF-β signaling pathway in B cells. These results answered our research question on how IL-33 affects BV and its underlying mechanism.
Using G. vaginalis to construct a BV model in mice is based on its significant role in human BV. First, it is because of the pathogenic relevance in BV. G. vaginalis is one of the primary bacteria associated with human BV and is often detected at high levels in affected individuals.30,31 It is involved in disrupting the vaginal microbiota, reducing beneficial Lactobacillus populations, and creating a dysbiotic environment that mimics BV conditions.30,31 Second, it could induce key BV features. G. vaginalis produces biofilms on the vaginal epithelium, which are characteristic of BV and provide a niche for other anaerobic bacteria. It releases virulence factors, such as hemolysins and cytotoxins, that can damage host tissues and contribute to inflammation.30,31 Using G. vaginalis can replicate several hallmark features of BV, such as vaginal dysbiosis, immune response alterations, etc. Third, G. vaginalis is relatively easy to culture and administer to mice, making it feasible for constructing reproducible experimental BV models.
G. vaginalis elicits both local and systemic effects. It stimulates epithelial cells to produce pro-inflammatory cytokines such as IL-1β, IL-6, and IL-8, which recruit neutrophils and other immune cells to the infection site.32 On a molecular level, G. vaginalis has been shown to activate Toll-like receptor 2, leading to NF-κB activation and a cascade of inflammatory responses.33 These immune interactions contribute to the pathogenesis of BV by enhancing microbial colonization, tissue disruption, and creating an environment conducive to secondary infections and complications such as increased susceptibility to STIs and adverse pregnancy outcomes.34
Our findings are consistent with previous studies that showed that IL-33 is involved in mucosal immunity and inflammation in various tissues.27,35 IL-33 has been reported to regulate IgA production by B cells in the gut and the lung, as well as to enhance TGF-β signaling in macrophages.36–38 Our study extended these findings to the vagina and showed that IL-33 plays a protective role against BV by modulating intravaginal IgA expression through TGF-β signaling pathway in B cells. Moreover, our findings have important implications for the understanding and treatment of BV. BV is a complex condition that involves multiple factors, such as vaginal microbiota, host immunity, hormones, and sexual behavior.2 Our study revealed a novel mechanism by which mucosal immunity protects against bacterial infection and inflammation in the vagina. IL-33 may be used as a biomarker for diagnosing BV or assessing its severity.
The treatment strategies for BV, both traditional and emerging, aim to restore the vaginal microbiome and alleviate symptoms. The traditional therapies include the use of antibiotics, metronidazole, clindamycin and boric acid.8 Despite their effectiveness, antibiotic treatments for BV are limited by high recurrence rates, potential side effects, and the risk of antimicrobial resistance. To counter the high recurrence rates of BV, probiotics (specifically Lactobacillus species) are increasingly being explored as adjunctive or preventive treatments. Immunotherapy for BV is a newer, innovative approach that targets the immune system’s role in the pathogenesis of BV.12 Since BV is not only a microbial imbalance but also involves immune dysregulation, immunotherapeutic strategies aim to modulate host responses to restore balance. For example, cytokine modulation is the focus of immunotherapy. BV is often associated with an inflammatory response, particularly an increase in pro-inflammatory cytokines like TNF-α and IL-1β.39 Immunotherapies that target these cytokines or their signaling pathways could potentially reduce the inflammation and promote the recovery of a healthy microbiota. For example, topical or systemic administration of anti-inflammatory agents might be used to suppress the inflammatory environment that allows pathogenic bacteria to thrive. Traditional therapies for BV, particularly antibiotics, remain the most widely used treatment options but face challenges related to recurrence and antimicrobial resistance. Probiotics have emerged as a promising adjunct to antibiotic therapy, helping to restore the vaginal microbiome. In contrast, immunotherapy offers an exciting, albeit still experimental, avenue for more targeted treatment. By modulating the immune response or developing vaccines and immune-based therapies, immunotherapy could potentially provide a more sustainable, long-term solution to BV, reducing recurrence and improving patient outcomes. However, more research is needed to establish the safety, efficacy, and practical applications of these newer treatments.
However, our study also has some limitations that should be acknowledged. First, we used a mouse model of BV induced by G. vaginalis, which may not fully reflect the complexity and diversity of human BV. Human BV is usually caused by a polymicrobial community that includes various anaerobes, such as Prevotella, Atopobium, and Sneathia.40 Therefore, further studies are needed to confirm our findings in human samples and with other BV-associated bacteria. Second, we focused on the role of IL-33 in regulating intravaginal IgA expression through TGF-β signaling pathway in B cells, but we did not explore other possible mechanisms or pathways that may be involved in IL-33-mediated protection against BV. Therefore, future research should investigate the role of IL-33 in regulating other aspects of mucosal immunity and inflammation in the vagina.
Conclusion
Our study demonstrated that IL-33 prevents BV induced by G. vaginalis by modulating intravaginal IgA expression through TGF-β signaling pathway in B cells. This reveals a novel mechanism by which mucosal immunity protects against bacterial infection and inflammation in the vagina. Further research is warranted to explore its potential relevance in future therapeutic strategies.
Data Sharing Statement
The raw data could be obtained upon reasonable request to the corresponding author.
Funding
This study was funded by the Natural Science Foundation of Jilin (YDZJ202501ZYTS176).
Disclosure
The authors declare that there is no conflict of interest associated with this publication and there has been no significant financial support for this work that could have influenced its outcome.
References
1. Muzny CA, Balkus J, Mitchell C, et al. Diagnosis and management of bacterial vaginosis: summary of evidence reviewed for the 2021 centers for disease control and prevention sexually transmitted infections treatment guidelines. Clin Infect Dis. 2022;74(Suppl_2):S144–S151. doi:10.1093/cid/ciac021
2. Sobel JD. Bacterial vaginosis. Annu Rev Med. 2000;51:349–356. doi:10.1146/annurev.med.51.1.349
3. Peebles K, Velloza J, Balkus JE, McClelland RS, Barnabas RV. High global burden and costs of bacterial vaginosis: a systematic review and meta-analysis. Sex Transm Dis. 2019;46(5):304–311. doi:10.1097/OLQ.0000000000000972
4. Bagnall P, Rizzolo D. Bacterial vaginosis: a practical review. JAAPA. 2017;30(12):15–21. doi:10.1097/01.JAA.0000526770.60197.fa
5. Abou Chacra L, Fenollar F, Diop K. Bacterial vaginosis: what do we currently know? Front Cell Infect Microbiol. 2021;11:672429. doi:10.3389/fcimb.2021.672429
6. Khedkar R, Pajai S. Bacterial vaginosis: a comprehensive narrative on the etiology, clinical features, and management approach. Cureus. 2022;14(11):e31314. doi:10.7759/cureus.31314
7. Kumar N, Behera B, Sagiri SS, Pal K, Ray SS, Roy S. Bacterial vaginosis: etiology and modalities of treatment-A brief note. J Pharm Bioallied Sci. 2011;3(4):496–503. doi:10.4103/0975-7406.90102
8. Abbe C, Mitchell CM. Bacterial vaginosis: a review of approaches to treatment and prevention. Front Reprod Health. 2023;5:1100029. doi:10.3389/frph.2023.1100029
9. Vicariotto F, Mogna L, Del Piano M. Effectiveness of the two microorganisms Lactobacillus fermentum LF15 and Lactobacillus plantarum LP01, formulated in slow-release vaginal tablets, in women affected by bacterial vaginosis: a pilot study. J Clin Gastroenterol. 2014;48(Suppl 1):S106–12. doi:10.1097/MCG.0000000000000226
10. Ai F, Wang Y, Wang J, Zhou L, Wang S. Effect of estrogen on vaginal complications of pessary use: a systematic review and meta-analysis. Climacteric. 2022;25(6):533–542. doi:10.1080/13697137.2022.2079973
11. Haggerty CL, Hillier SL, Bass DC, Ness RB, Evaluation PID. Clinical health study i. Bacterial vaginosis and anaerobic bacteria are associated with endometritis. Clin Infect Dis. 2004;39(7):990–995. doi:10.1086/423963
12. Amabebe E, Anumba DOC. Mechanistic insights into immune suppression and evasion in bacterial vaginosis. Curr Microbiol. 2022;79(3):84. doi:10.1007/s00284-022-02771-2
13. Muzny CA, Laniewski P, Schwebke JR, Herbst-Kralovetz MM. Host-vaginal microbiota interactions in the pathogenesis of bacterial vaginosis. Curr Opin Infect Dis. 2020;33(1):59–65. doi:10.1097/QCO.0000000000000620
14. Hapfelmeier S, Lawson MA, Slack E, et al. Reversible microbial colonization of germ-free mice reveals the dynamics of IgA immune responses. Science. 2010;328(5986):1705–1709. doi:10.1126/science.1188454
15. Milovanovic M, Volarevic V, Radosavljevic G, et al. IL-33/ST2 axis in inflammation and immunopathology. Immunol Res. 2012;52(1–2):89–99. doi:10.1007/s12026-012-8283-9
16. Selvaskandan H, Shi S, Twaij S, Cheung CK, Barratt J. Monitoring immune responses in iga nephropathy: biomarkers to guide management. Front Immunol. 2020;11:572754. doi:10.3389/fimmu.2020.572754
17. Oh JE, Kim BC, Chang DH, et al. Dysbiosis-induced IL-33 contributes to impaired antiviral immunity in the genital mucosa. Proc Natl Acad Sci U S A. 2016;113(6):E762–71. doi:10.1073/pnas.1518589113
18. Lewis WG, Robinson LS, Perry J, et al. Hydrolysis of secreted sialoglycoprotein immunoglobulin A (IgA) in ex vivo and biochemical models of bacterial vaginosis. J Biol Chem. 2012;287(3):2079–2089. doi:10.1074/jbc.M111.278135
19. Cauci S, Driussi S, Guaschino S, Isola M, Quadrifoglio F. Correlation of local interleukin-1beta levels with specific IgA response against Gardnerella vaginalis cytolysin in women with bacterial vaginosis. Am J Reprod Immunol. 2002;47(5):257–264. doi:10.1034/j.1600-0897.2002.01096.x
20. Gilbert NM, Lewis WG, Lewis AL. Clinical features of bacterial vaginosis in a murine model of vaginal infection with Gardnerella vaginalis. PLoS One. 2013;8(3):e59539. doi:10.1371/journal.pone.0059539
21. Omana-Zapata I, Mutschmann C, Schmitz J, et al. Accurate and reproducible enumeration of T-, B-, and NK lymphocytes using the BD FACSLyric 10-color system: a multisite clinical evaluation. PLoS One. 2019;14(1):e0211207. doi:10.1371/journal.pone.0211207
22. Wilson J. Managing recurrent bacterial vaginosis. Sex Transm Infect. 2004;80(1):8–11. doi:10.1136/sti.2002.002733
23. Cazac BB, Roes J. TGF-beta receptor controls B cell responsiveness and induction of IgA in vivo. Immunity. 2000;13(4):443–451. doi:10.1016/s1074-7613(00)00044-3
24. Guaschino S, De Seta F, Piccoli M, Maso G, Alberico S. Aetiology of preterm labour: bacterial vaginosis. BJOG. 2006;113(Suppl 3):46–51. doi:10.1111/j.1471-0528.2006.01122.x
25. Mitchell C, Marrazzo J. Bacterial vaginosis and the cervicovaginal immune response. Am J Reprod Immunol. 2014;71(6):555–563. doi:10.1111/aji.12264
26. Onderdonk AB, Delaney ML, Fichorova RN. The human microbiome during bacterial vaginosis. Clin Microbiol Rev. 2016;29(2):223–238. doi:10.1128/CMR.00075-15
27. De la Fuente M, MacDonald TT, Hermoso MA. The IL-33/ST2 axis: role in health and disease. Cytokine Growth Factor Rev. 2015;26(6):615–623. doi:10.1016/j.cytogfr.2015.07.017
28. Machado A, Cerca N. Influence of biofilm formation by Gardnerella vaginalis and other anaerobes on bacterial vaginosis. J Infect Dis. 2015;212(12):1856–1861. doi:10.1093/infdis/jiv338
29. Corthesy B. Role of secretory IgA in infection and maintenance of homeostasis. Autoimmun Rev. 2013;12(6):661–665. doi:10.1016/j.autrev.2012.10.012
30. Kwak J, Pandey S, Cho J, et al. Development of the standard mouse model for human bacterial vaginosis induced by Gardnerella vaginalis. Front Vet Sci. 2023;10:1226859. doi:10.3389/fvets.2023.1226859
31. Morrill S, Gilbert NM, Lewis AL. Gardnerella vaginalis as a cause of bacterial vaginosis: appraisal of the evidence from in vivo models. Front Cell Infect Microbiol. 2020;10:168. doi:10.3389/fcimb.2020.00168
32. Gillet E, Meys JF, Verstraelen H, et al. Bacterial vaginosis is associated with uterine cervical human papillomavirus infection: a meta-analysis. BMC Infect Dis. 2011;11:10. doi:10.1186/1471-2334-11-10
33. Zhang I, Pletcher SD, Goldberg AN, Barker BM, Cope EK. Fungal microbiota in chronic airway inflammatory disease and emerging relationships with the host immune response. Front Microbiol. 2017;8:2477. doi:10.3389/fmicb.2017.02477
34. Hillier SL, Nugent RP, Eschenbach DA, et al. Association between bacterial vaginosis and preterm delivery of a low-birth-weight infant. The vaginal infections and prematurity study group. N Engl J Med. 1995;333(26):1737–1742. doi:10.1056/NEJM199512283332604
35. Miller AM. Role of IL-33 in inflammation and disease. J Inflamm. 2011;8(1):22. doi:10.1186/1476-9255-8-22
36. Le Goffic R, Arshad MI, Rauch M, et al. Infection with influenza virus induces IL-33 in murine lungs. Am J Respir Cell Mol Biol. 2011;45(6):1125–1132. doi:10.1165/rcmb.2010-0516OC
37. Sattler S, Ling GS, Xu D, et al. IL-10-producing regulatory B cells induced by IL-33 (Breg(IL-33)) effectively attenuate mucosal inflammatory responses in the gut. J Autoimmun. 2014;50(100):107–122. doi:10.1016/j.jaut.2014.01.032
38. Stolarski B, Kurowska-Stolarska M, Kewin P, Xu D, Liew FY. IL-33 exacerbates eosinophil-mediated airway inflammation. J Immunol. 2010;185(6):3472–3480. doi:10.4049/jimmunol.1000730
39. Thurman AR, Doncel GF. Innate immunity and inflammatory response to Trichomonas vaginalis and bacterial vaginosis: relationship to HIV acquisition. Am J Reprod Immunol. 2011;65(2):89–98. doi:10.1111/j.1600-0897.2010.00902.x
40. Turovskiy Y, Sutyak Noll K, Chikindas ML. The aetiology of bacterial vaginosis. J Appl Microbiol. 2011;110(5):1105–1128. doi:10.1111/j.1365-2672.2011.04977.x
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms.php
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms.php
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.