")
Back to Journals » International Journal of Nanomedicine » Volume 20
Immunomodulatory Effects and Mechanisms of Two-Dimensional Black Phosphorus on Macrophage Polarization and Bone Regeneration
Authors Zou Y , Xie Q, Lin J, Dong H, Zhuang X, Xian R, Liang Y, Li S
Received 3 December 2024
Accepted for publication 18 March 2025
Published 9 April 2025 Volume 2025:20 Pages 4337—4355
DOI https://doi.org/10.2147/IJN.S508309
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Xing Zhang
Yue Zou,1,* Qinkai Xie,1,* Jiating Lin,1 Hao Dong,1 Xianxian Zhuang,1 Ruoting Xian,1 Youde Liang,2,3 Shaobing Li1
1Center of Oral Implantology, Stomatological Hospital, School of Stomatology, Southern Medical University, Guangzhou, People’s Republic of China; 2Department of stomatology center, The People’s Hospital of Baoan Shenzhen, Shenzhen, People’s Republic of China; 3The Second Affiliated Hospital of Shenzhen University, Shenzhen, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Youde Liang, Department of stomatology center, The People’ s Hospital of Baoan Shenzhen, No. 118 Longjing 2nd Road, Xin’an Street, Bao’an District, Shenzhen, 518100, People’s Republic of China, Email [email protected] Shaobing Li, Center of Oral Implantology, Stomatological Hospital, School of Stomatology, Southern Medical University, No. 366, South of Jiangnan Boulevard, Guangzhou, 510280, People’s Republic of China, Email [email protected]
Abstract: The repair of bone defects poses a significant challenge within the realm of clinical medicine. However, with the advent of various emerging biotechnologies, bone tissue engineering (BTE) has emerged as a promising discipline that offers innovative therapeutic strategies to address bone defects. Among the novel biomaterials being explored, two-dimensional (2D) black phosphorus (BP) has attracted considerable attention due to its advantageous properties, which include antimicrobial activity, drug delivery capabilities, and effective photothermal conversion. These properties render BP an excellent candidate for BTE applications. Recent studies have indicated that BP possesses remarkable immunomodulatory properties that influence bone regeneration, profoundly impacting the transformation of the osteoimmune microenvironment, thereby guiding the process of bone remodeling. Macrophage is a principal component of the osteoimmune microenvironment, and evidence suggests that BP significantly influences the polarization of macrophage M1 and M2 phenotypes. This review aims to present the regulatory effects and underlying mechanisms of 2D BP on macrophage polarization in the immune microenvironment. It highlights the ability of BP to systematically modulate the inflammatory environment and to facilitate the metabolic reprogramming of macrophages. The review concludes with a discussion of the potential applications and limitations of BP nanomaterials in the field of BTE.
Keywords: osteoimmune microenvironment, macrophage metabolic reprogramming, cell interaction, bone remodeling
Introduction
Among various systemic diseases, bone tissue defects and deformities resulting from factors such as trauma, tumors, and inflammation, along with the functional impairments they cause, have always been a significant challenge within the field of clinical medicine. Bone defects can lead to a reduction or complete loss of bone conduction and weight-bearing capabilities, significantly affecting the normal physiological functions of patients. The clinical management of bone defects presents difficulties in restoring bone integrity through anti-inflammatory repair methods alone. In addition, although autologous and allogeneic bone grafting are recognized as effective therapeutic approaches, they are still associated with risks such as immune rejection and suboptimal osseointegration outcomes.1,2 Therefore, the development of BTE and regenerative medicine has become an inevitable trend in the clinical repair of bone defects. In recent years, with the wide application of cell culture technology, advanced nanomaterials, multifunctional hydrogels and 3D-printed scaffolds have gradually become the main materials for BTE.3,4 These materials can mimic the native microenvironment and promote the recruitment, proliferation and differentiation of stem cells, as well as the construction of bone tissue structure.5,6 Accordingly, it is an effective treatment for bone regeneration.
Dynamic regulation of the bone immune system occurs throughout the bone healing process, wherein macrophages assume a complex and nuanced role. For instance, bone macrophages adhering to the bone surface induce osteogenic differentiation and promote bone anabolism,7 and these cells have the capacity to differentiate into pro-inflammatory M1 macrophages or anti-inflammatory M2 macrophages in response to both internal and external environmental stimuli, thereby influencing the homeostasis of the internal environment.8,9 In the early stages of inflammation, M1-type macrophages play a crucial role in removing of pathogenic bacteria and necrotic tissue, thereby contributing to the maintenance of metabolic balance. Subsequently, these M1-type cells transition into M2-type macrophages, which are responsible for resolving inflammation and initiating tissue regeneration and repair.9 However, an imbalance in the M1-M2 transition, coupled with the accumulation of inflammatory factors, can lead to osteoclast activation and inhibition of osteogenesis, ultimately impeding the bone healing process.10 Macrophage populations respond to local environmental changes by adjusting the interconversion of M1 and M2, which significantly influences bone metabolism.11,12 Consequently, macrophages emerge as critical targets in the context of bone regeneration facilitated by tissue engineering materials. Biomaterials enhance bone regeneration by improving the osteoimmune microenvironment and restoring the transformation balance of M1 and M2 phenotypes, thereby supporting a shift in the immune microenvironment from a pro-inflammatory to an anti-inflammatory state.13,14
As an emerging bio-nanomaterial, 2D BP exhibits a unique spatial structure and distinct physicochemical properties, including a high specific surface area, tunable band gap, and relatively facile surface functionalization. These characteristics allow it to demonstrate significant efficacy in drug delivery, photothermal therapy (PTT), and photodynamic therapy (PDT). Furthermore, since the primary degradation product of BP is non-toxic phosphate ions, the focus of clinical research has shifted from “immune evasion” to “immune interaction”, further facilitating the application of BP in the field of tissue engineering.15,16 Numerous studies have indicated that BP nanomaterials can effectively induce bone tissue regeneration. For instance, BP has been shown to activate the BMP-RUNX2 signaling pathway and the Wnt signaling pathway in stem cells, thereby effectively enhancing osteogenesis.17–19 Additionally, there is evidence that BP possesses favorable bone immunomodulatory effects on bone,20,21 that contribute to bone regeneration by modulating the overall immune microenvironment. However, to date, no comprehensive review has thoroughly examined the interactions between BP and the bone immune system. There remain significant gaps in our understanding of the bone immunomodulatory role of BP, particularly regarding the sequential regulation of the inflammatory microenvironment, the sequential activation of macrophages, and the effects on osteoclast differentiation by BP. These areas may represent significant avenues for research aimed at optimizing bone regeneration outcomes.
This paper provides a comprehensive review of the immunomodulatory effects of BP nanomaterials in the context of bone tissue regeneration. A substantial body of literature indicates that BP can enhance the inflammatory microenvironment of bone tissue, facilitate the transition of macrophages from the M1 phenotype to the M2 phenotype, and inhibit osteoclast differentiation, thereby promoting bone regeneration. Furthermore, BP has been shown to enhance neural tissue activity through immunomodulatory mechanisms, which further supports bone repair and regeneration. These findings establish a fundamental basis for the advanced application of BP in the field of BTE.
The Role of Macrophages in Bone Regeneration
In the physiological environment, bone homeostasis is dynamically regulated by the immune system. Within the immune microenvironment of bone, macrophages play a crucial role in facilitating bone anabolism or in activating osteoclasts leading to bone resorption, thereby regulating bone accumulation and influencing the prognosis of bone repair.14,22
Osteoblasts and osteoclasts play a critical role in the bone healing process, meticulously regulating bone remodeling through complex intercellular communication. And it has been demonstrated that the receptor activator of nuclear factor kappa-B ligand (RANKL)-receptor activator of NF-κB (RANK)-osteoprotegerin (OPG) signaling pathways are fundamental to bone metabolism. Various immune cells enhance RANKL expression to facilitate osteoclast differentiation and bone resorption, while OPG, mainly produced by osteoblasts, acts as a competitive antagonist to RANKL, and inhibits osteoclastogenesis.23 Furthermore, osteoblasts and osteoclasts secrete a number of active molecules that modulate each other’s activity,24 and thus work in concert to maintain the balance between bone regeneration and bone resorption.
Macrophages are capable of targeting osteoblast and osteoclast activities, thereby exerting various immunomodulatory effects on bone metabolism (as illustrated in Figure 1).25 Among them, M1-type macrophages, induced by lipopolysaccharide (LPS) or interferon-gamma (IFN-γ) secrete pro-inflammatory factors that facilitate the clearance of pathogens.26 However, under conditions of oxidative stress, M1 macrophages inhibit the proliferation and migration of pro-osteoblasts, leading to a reduction in bone mass.27 Conversely, M2 macrophages induced by interleukin-4 (IL-4) and interleukin-13 (IL-13) secrete anti-inflammatory factors to construct an immune-friendly microenvironment,26 and they can regulate the BMP2/SMAD signaling pathway in mesenchymal stem cells (MSCs) through paracrine mechanisms to enhance osteogenic differentiation and bone mass accumulation.28,29 In summary, M1 and M2 phenotypic macrophages exert distinct immunomodulatory effects on bone homeostasis.
 |
Figure 1 The role of macrophages in bone homeostasis. Macrophages secrete cytokines, including TNF-α, IL-1, and IL-6, which inhibit bone formation and promote bone resorption. They can also release cytokines such as IL-4, IL-10, and IL-13, inhibiting osteoclast differentiation. OSM, VEGF, IGF, TGF-β, and BMP-2 are secreted to stimulate osteoblast differentiation. Notes: Reproduced from Hu Y, Huang J, Chen C, et al. Strategies of Macrophages to Maintain Bone Homeostasis and Promote Bone Repair: A Narrative Review. Journal of Functional Biomaterials. 2023; 14(1):18.© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license.25 |
It’s worth noting that while M1 macrophages typically contribute to bone resorption and osteogenic inhibition during chronic inflammation, they play a beneficial role in the early stages of injury by directing and initiating the bone healing process. For instance, M1 macrophages secrete tumor necrosis factor-alpha (TNF-α), which protects hematopoietic stem cells (HSCs) from necrotic apoptosis, while supporting HSC cell cycle activation and facilitating bone marrow regeneration.30 In addition, M1 macrophages release activators that recruit bone marrow mesenchymal stem cells (BMMSCs)31 and activate both the oncostatin M (OSM)-glycoprotein 130 (gp130) signaling complex and the bone morphogenetic protein 6(BMP6)-SMAD pathway in stem cells, which are critical for early osteogenic differentiation.22 Thus, it has been shown that M1 macrophages have a significant osteogenic induction effect on stem cells. Furthermore, it has been observed that an alkaline microenvironment induces M1 polarization and enhances the secretion of BMP6 and OSM, thereby promoting the expression of early osteogenic genes. It demonstrates a more pronounced positive bone immunomodulatory effect compared to M2 polarization induced in an acidic microenvironment.32 In conclusion, sufficient evidence indicates that M1 macrophages are a critical driver of bone anabolism and occupy a unique position within the bone immune microenvironment.
As shown above, it is clear that M1 macrophages play a dual role in the process of bone healing: they facilitate bone anabolism while simultaneously inducing the occurrence of an early inflammatory environment. It’s worth noting that the inflammatory response acts as a double-edged sword. While the initial inflammatory response is crucial for eliminating foreign bodies and bolstering tissue defenses, the sustained presence of M1 macrophages and the accompanying inflammatory cascade in damaged bone tissue can hinder the process of bone remodeling. Consequently, it is essential to address and mitigate this prolonged inflammatory response.
In conclusion, the activity of M1 and M2 type macrophages in the vicinity of bone tissue plays a significant role in the bone healing process. Therefore, the moderate modulation of M1/M2 polarization through the use of biomaterials represents an effective strategy to enhance bone regeneration.
BP Regulates the Inflammatory Microenvironment to Facilitate Bone Regeneration
The immune microenvironment plays a critical role in tissue injury, repair, and regeneration processes. Proper regulation of the immune environment is the cornerstone of bone regeneration. This is especially pertinent in the context of chronic inflammatory bone diseases, where the restoration and maintenance of bone immune homeostasis significantly influence disease prognosis. Extensive experimental data demonstrates that BP can systematically regulate the inflammatory environment and exert anti-inflammatory effects, thereby accelerating the bone remodeling process.20,33,34
Sequential Regulation of BP in the Pro-Inflammatory and Anti-Inflammatory Environment
The implantation of BP nanomaterials in the body inevitably induces an intrinsic immune response, but their excellent biocompatibility facilitates the regulation of inflammation, which is essential for the acceptance of BP by the host defense system.35,36 It should also be clear that the acute pro-inflammatory and anti-inflammatory phases are not opposed to each other, but rather are a sequential process. The acute inflammatory phase prepares for the subsequent anti-inflammatory phase and is periodically regulated by the local environment, where the sequential regulation of inflammation and macrophage activation are crucial for tissue regeneration.37–39 It has been demonstrated that BP is able to stimulate a certain degree of acute immune response and expedite the resolution of the inflammatory process, thus preventing tissue damage associated with prolonged chronic inflammation.40,41
Recent studies have demonstrated that BP can directly interact with macrophages, thereby activating early inflammatory responses and facilitating the transition from an inflammatory to a reparative environment. In the experiment, black phosphorus nanosheets (BPNSs) were co-cultured with resting macrophages (M0) and it was found that BPNSs transiently activated M0 cells, inducing a mild inflammatory response and progressively promoting an anti-inflammatory environment as the material naturally degrades. Simultaneously, the expression of the viral defense protein in macrophages was diminished, suggesting that BPNSs exhibit favorable biocompatibility.42 Additionally, BP composite scaffolds can stimulate macrophages to express interleukin-33 (IL-33),20 a crucial cytokine involved in the regulation of the immune response.43 IL-33 plays a critical role in the systematic release of pro-inflammatory and anti-inflammatory factors in the local environment, as well as in the acceleration of osteogenic differentiation of BMMSCs.20 Research on IL-33 has demonstrated that at an early stage, the IL-33/ST2 signaling axis activates the NF-κB and MAPK signaling pathways, thus inducing inflammatory responses.44 However, as the concentration of IL-33 in the local environment increases, it exhibits an inhibitory effect on osteoclast differentiation and bone resorption. Additionally, it promotes the differentiation of M2-type macrophages and the expansion of TIGIT+ regulatory T cells,45 which collectively exert a more pronounced anti-inflammatory and reparative effect. Therefore, it can be concluded that BP facilitates the transition of injured tissues from the inflammatory phase to the repair phase by inducing the release of IL-33 from macrophages.
In addition, BP sequentially modulates the transformation of the immune environment by enhancing the metabolic activity of tissue cells. Research has demonstrated that BP can specifically activate peroxisome proliferator-activated receptor γ (PPARγ) to improve cellular metabolism and facilitate the transition out of the inflammatory phase.40 PPARγ is a nuclear receptor found in various tissues and immune cells, playing a critical role in tissue glycolipid metabolism, inflammatory responses, and cell differentiation.46 Notably, black phosphorus quantum dots (BPQDs) at a concentration of 0.1 mg/kg were observed to inhibit the PPARγ signaling pathway while activating the NF-κB signaling pathway, resulting in the infiltration of inflammatory factors. Conversely, BPQDs at a concentration of 1 mg/kg were found to activate PPARγ and the glycolytic pathway while inhibiting the NF-κB pathway, thereby mitigating tissue damage caused by inflammation.40 Studies on PPARγ have revealed that crosstalk between the PPARγ and NF-κB pathways can reverse M1/M2 polarization and antagonize NF-κB signaling,47 which partially elucidates the anti-inflammatory effects of BP. Furthermore, PPARγ is instrumental in maintaining the characteristic lipid metabolism of M2 macrophages,48 and thus serves as a key mediator of the anti-inflammatory environment regulated by BP. Consequently, it is evident that upon entering the organism, BP can alleviate aspects of the acute inflammatory response by enhancing local tissue metabolism, thereby maintaining immune homeostasis.
Comprehensive analyses indicate that BP nanomaterials do not simply suppress local inflammatory responses during the repair of damaged tissue; rather, these materials initiate the inflammatory pathway early, establishing a controlled pro-inflammatory environment. This transient and moderate inflammatory response creates optimal conditions for subsequent tissue repair. Furthermore, BP possess the ability to regulate the transition from a pro-inflammatory to an anti-inflammatory environment according to a strict temporal sequence. The ultimate objective of this process is to establish an immune-friendly regenerative environment, thereby facilitating effective bone tissue regeneration.
BP Facilitates the Transformation of the Anti-Inflammatory Environment
In the treatment of bone tissue disorders, BP nanomaterials exhibit significant immunomodulatory benefits due to their antioxidant and slow degradation properties. Substantial evidence indicates that BP significantly enhances the release of numerous anti-inflammatory factors and bioactive molecules, thereby ameliorating the bone immune microenvironment.21,49,50 For instance, during the wound-healing process following tooth extraction, BPNSs mitigate oxidative stress by scavenging reactive oxygen species (ROS) and support the regeneration of blood vessels and alveolar bone.51 In the context of degenerative disc disease, BPQDs inhibit the acid-sensitive ion channel (ASIC)-mediated release of NLRP3 inflammatory vesicles and interleukin-1 beta (IL-1β), thereby fostering an anti-inflammatory environment and facilitating nucleus pulposus (NP) regeneration.33 Furthermore, in the treatment of osteoarthritis (OA), BPNSs obstruct the secretion of metalloproteinases, specifically a disintegrin and metalloproteinase with thrombospondin motifs 5 (ADAMTS5), a disintegrin and metalloproteinase with thrombospondin motifs 1 (ADAMTS1), and matrix metalloproteinase 13 (MMP13),34 while also directing the subchondral osteogenic process by modulating the inflammatory milieu. Additionally, in the treatment of hormonal osteonecrosis, periodontitis, and osteoporosis, BP has been shown to downregulate the pro-inflammatory factors such as IL-1β, TNF-α, interleukin-6 (IL-6), and inducible nitric oxide synthase (iNOS), while elevating the expression of anti-inflammatory factors such as interleukin-10 (IL-10), arginase-1 (Arg-1), and transforming growth factor-beta (TGF-β),21,49,50 (as illustrated in Figure 2).21 This regulatory mechanism ultimately establishes an immune microenvironment conducive to bone tissue repair. Meanwhile, in the context of bone tissue reconstruction following osteosarcoma resection, BP effectively stimulates the secretion of IL-10 and inhibits the expression of IL-6 and cyclooxygenase-2 (COX-2) in tumor cells,52,53 thereby initiating the processes of bone repair and regeneration. In summary, it is evident that establishing an anti-inflammatory environment is essential for the processes of osteogenic differentiation and mineralization, with the immunomodulatory and reparative functions of BP playing a crucial role in bone remodeling.
 |
Figure 2 Mechanism of osteoimmune environment induced by BP degradation to accelerate bone regeneration. This study demonstrated that the PLGA/BP scaffold has osteogenic effects and bone immunomodulatory functions through in vitro and in vivo experiments. Notes: Reproduced from Long J, Yao Z, Zhang W, et al. Regulation of Osteoimmune Microenvironment and Osteogenesis by 3D-Printed PLAG/black Phosphorus Scaffolds for Bone Regeneration. Adv Sci (Weinh). 2023 Oct;10(28):e2302539. © 2023 The Authors. Advanced Science published by Wiley‐VCH GmbHThis is an open access article under the terms of the http://creativecommons.org/licenses/by/4.0/ License.21 |
A substantial body of literature indicates that BP possesses multiple mechanisms of action in modulating the anti-inflammatory environment. Firstly, as a potent photosensitizer, BP can synergistically facilitate the transformation of the anti-inflammatory milieu through photothermal therapy (PTT). Studies demonstrate that BP-based nanoplatforms can inhibit the expression of TNF-α in wounds exposed to near-infrared (NIR) radiation. The reduction of TNF-α subsequently accelerates collagen fiber deposition and angiogenesis. Concurrently, NIR light enhances the inhibitory effects of BP on IL-1β and IL-6 secretion while promoting the induction of TGF-β and IL-10 secretion. The cytokine regulation by BP composites in the immune microenvironment is further detailed in Table 1.41,54–57 In addition to the effects of PTT itself, it is noteworthy that PTT-activated heat shock protein 70 (Hsp70) exhibits beneficial immunomodulatory properties. Existing literature has shown that elevated intracellular levels of Hsp70 can inhibit NF-κB signal transduction and enhance the secretion of anti-inflammatory factors.58 BP scaffolds have been shown to effectively increase the expression levels of HSP70 in periosteal-derived stem cells (PDSCs) and human bone marrow mesenchymal stem cells (hBMSCs) under NIR light irradiation,59–61 with HSP70 further demonstrating significant immunomodulatory and osteogenic induction effects. Secondly, BP can enhance the metabolic activity of immune-responsive cells, thereby indirectly maintaining immune homeostasis. For instance, BPNSs can activate the JAK1-STAT1/3-OAS metabolic pathway in human umbilical vein endothelial cells (HUVECs),42 which subsequently improves cytokine secretion and promotes vascular regeneration. Furthermore, BP has the ability to stabilize protein structures through its electrostatic adsorption properties, thereby mitigating the inflammatory burden on the body caused by the accumulation of abnormal proteins. For instance, BP has been shown to inhibit the deposition of β-amyloid (Aβ) and the abnormal folding of α-synuclein (α-syn) monomers, thus avoiding further inflammatory responses.62,63 And BP stabilizes the structural integrity of angiotensin-converting enzyme 2 (ACE2),64 which obstructs protein interactions and effectively prevents viral infections and immune disorders. Last but not least, BP serves as an excellent carrier role in combination with drugs and can effectively control the inflammatory response within the wound, achieving good anti-inflammatory and detoxifying effects.65,66
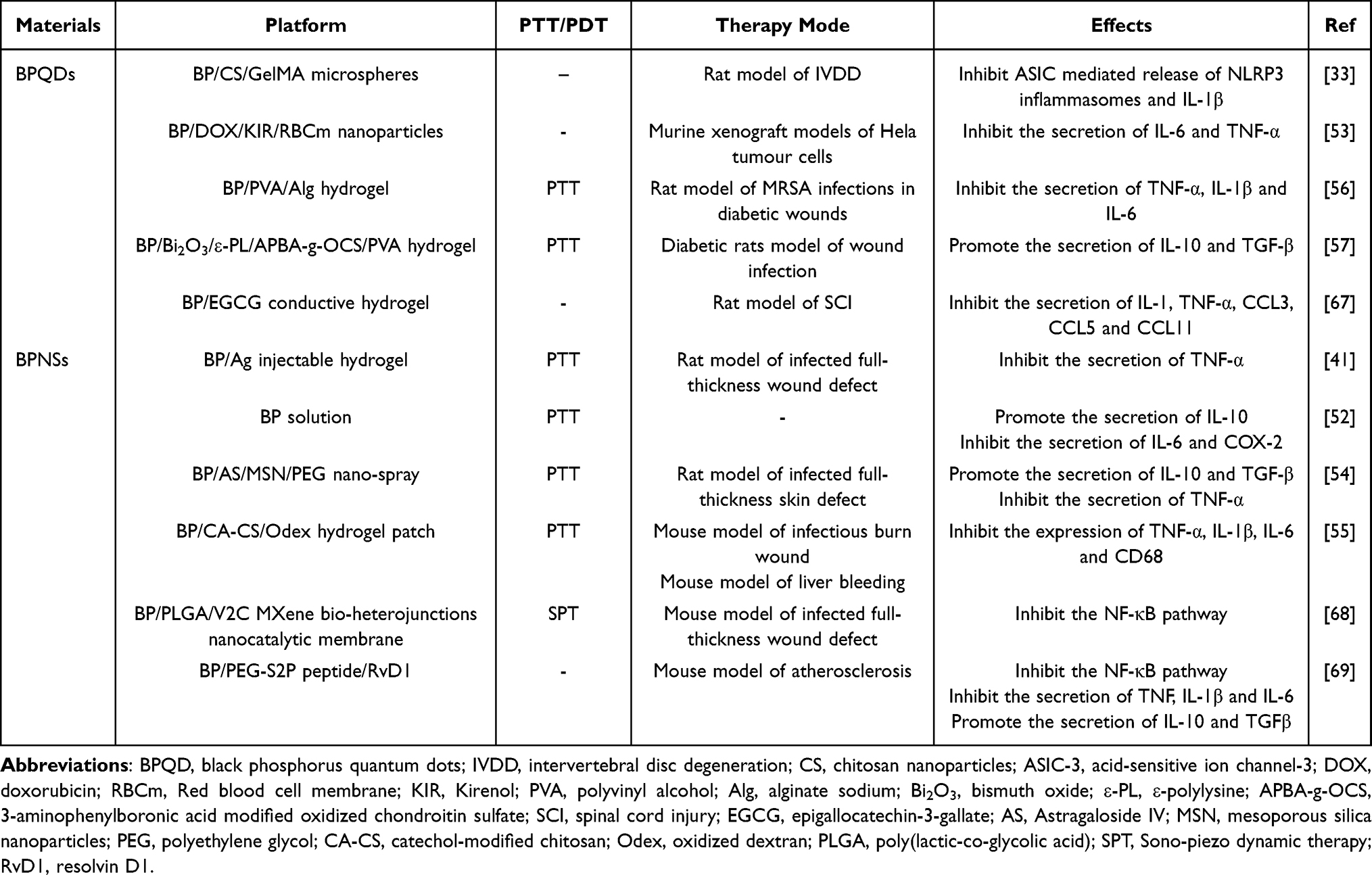 |
Table 1 Cytokine Regulation by BP Composites in the Immune Microenvironment |
While BP promotes the transformation of the anti-inflammatory environment, it also enhances the survival of damaged tissue cells. Research has indicated that small quantities of BP can mitigate ROS-induced tissue damage and apoptosis,70–72 (as illustrated in Figure 3).72 This protective mechanism may be associated with the PDK1/AKT/14-3-3/BAD and PI3K/Akt signaling pathways.73,74 Additionally, BP regulates the cell cycle through the PDK1/AKT/GSK3 signaling pathway,73 thereby facilitating tissue regeneration. Consequently, BP not only diminishes the detrimental effects of oxidative stress on the immune microenvironment but also activates the cell’s intrinsic anti-apoptotic mechanisms, ultimately promoting tissue survival.
 |
Figure 3 Schematic of the mechanism by which BP hydrogel motivates macrophage polarization and accelerates myocardial repair. CS-Gel-BP@PDA composite hydrogel can effectively attenuate oxidative stress, promote macrophage M2 phenotypic polarization, and facilitate the recovery of cardiac function. Notes: Reproduced from Zhang J, Sun D, Liao Y, et al. Time-Released Black Phosphorus Hydrogel Accelerates Myocardial Repairing through Antioxidant and Motivates Macrophage Polarization Properties. Biomater Res. 2024 May 8;28:0029. Copyright © 2024 Jiahui Zhang et al.Exclusive licensee Korean Society for Biomaterials, Republic of Korea. No claim to original U.S. Government Works. Distributed under a Creative Commons Attribution License 4.0 (CC BY 4.0).72 |
In conclusion, BP nanomaterials enhance the secretion of anti-inflammatory factors and bioactive molecules at the site of bone injury. They establish an immune-friendly microenvironment through intricate biological pathways, thereby creating optimal conditions for bone tissue regeneration.
BP Regulates Macrophages and Osteoclasts to Promote Bone Regeneration
Bone immune dysregulation represents a fundamental pathogenic factor in inflammatory bone diseases and persists throughout the progression of these conditions. Therefore, optimizing the bone immune environment is crucial for enhancing bone regeneration. Macrophages, as essential cellular components in the maintenance of bone immune homeostasis, have become a prominent focus of research in recent years. The subsequent section will provide a comprehensive discussion of the mechanisms underlying the immunomodulatory effects of BP on macrophages, ranging from passive immunity to active regulatory processes.
Pro-Inflammatory M1 Macrophages
When nanomaterials enter the bloodstream, they initially activate the body’s passive immune response and rejection mechanisms, indicating enhanced immune defenses of the host. The acute inflammatory response at the injury site is characterized by the presence of M1 macrophages and neutrophils. Consequently, BP exhibited a transient mild activation effect on M1 macrophages during the early stages of inflammation. As some BP nanomaterials are swallowed into cells and others are gradually degraded in blood, BP exerts its antioxidant effects and inhibits cellular inflammatory signaling pathways. Therefore, BP demonstrates a significant inhibitory effect on M1 in inflammatory environments. The varying immunomodulatory effects of BP on M1 will be discussed in detail below.
BP Induces Intrinsic Immunity to Facilitate M1 Polarization
BP nanomaterials enter the bloodstream and initially activate the body’s intrinsic immune response. Plasma proteins adsorb onto the surface of the nanomaterials, forming protein coronas. The black phosphorus corona complexes (BPCCs) interact with macrophage calmodulin, inducing calcium influx, which subsequently activates the NF-κB pathway within macrophages. This activation results in M1-type polarization75 and a 2-4-fold increase in the secretion of proinflammatory factors.76 Furthermore, an excess of intracellular ROS accelerates M1 production and inflammatory factors infiltration.77 Thus, the BPCCs formed by BP upon entering the bloodstream can interact with macrophages and induce M1-type polarization. Additionally, BP nanostructures can activate the integrin αvβ8 signaling pathway on macrophage membranes, triggering an immune response;78 however, they have minimal effects on the membrane, indicating that they can enhance macrophage immune response with low cytotoxicity. Furthermore, studies show that excessive levels of BPQDs result in ferroptosis in tissue cells,79 which is characterized as a mode of inflammatory cell death. In this context, dead cells release chemokines such as CCL2 and CCL7, which subsequently trigger an acute inflammatory response marked by the presence of M1-type macrophages.80 Consequently, the introduction of BP into the organism mildly activates M1 polarization and induces a shift in the inflammatory environment through various pathways.
It is worth noting that in the context of early inflammation, the generation of M1 macrophage populations is essential for strengthening host defenses. Beyond passively initiating the immune response, in certain instances, BP has the ability to actively stimulate M1 macrophages, thereby contributing to the maintenance of internal homeostasis. For instance, in the treatment of chronic osteomyelitis, BPNSs combined with iron oxides (Fe3O4) convert electromagnetic waves into thermal energy, resulting in a microwave thermal effect (MTE). MTE further activates the immune response and enhances the phagocytic activity of immune cells, thereby improving the overall bactericidal effect.81 Furthermore, in the field of tumor therapy, BP can selectively target tumor tissues, facilitate the metabolic reprogramming of M2 to M1 macrophages, and induce tumor cell apoptosis.82,83 Therefore, given the intricate relationship between macrophage activity and bone regeneration, the subsequent discussion will focus on the inhibitory role of M1 macrophages in chronic inflammation.
BP Improves the Inflammatory Environment and Inhibits M1 Polarization
In inflammatory environments, the antioxidant BP is capable of obstructing the inflammatory cascade and inhibiting the polarization of pro-inflammatory M1 macrophages. For instance, in models of osteoarthritis, periodontitis, and myocardial injury, BPNSs effectively scavenge excess ROS and inhibit the expression of M1-related cytokines, ultimately inducing the metabolic reprogramming of macrophages.34,49,84 Furthermore, BP has been shown to directly inhibit the NF-κB signaling pathway, a critical inflammatory pathway in M1 macrophages that is essential for maintaining immune homeostasis.68,69 And it is also noteworthy that PPARγ can act as a negative regulator of pro-inflammatory responses and M1 activation. Moderate concentrations of BP indirectly inhibit M1 production by targeting PPARγ activation.40,85 However, aberrant activation of PPARγ specifically promotes osteoclast differentiation while inhibiting osteoblast differentiation, which is closely associated with bone resorption in inflammatory bone diseases.85 Based on the theme of bone immunomodulation by BP, this aspect will not be further discussed here. In summary, BP, as a natural antioxidant that scavenges free oxygen radicals and limits the polarization of M1-type macrophages, plays a significant role in the regulation of inflammatory responses.
Furthermore, BP has the capacity to eliminate M1-associated inflammatory cells and indirectly inhibit M1 activation. Research has demonstrated that the local inflammatory milieu in rheumatoid arthritis (RA) fosters the production of invasive synovial fibroblasts (SFs),86 while elevated levels of aerobic glycolysis support their increased proliferative activity.87 SFs further interact with synovial macrophages (SMs) to enhance the metabolism of SM cells and increase their expression of pro-inflammatory factors.88 This synergistic interaction exacerbates the inflammatory response within the synovium. In the context of RA treatment, BP delivers substantial amounts of ROS to the inflamed joints under NIR, thereby eliminating excess proliferative synovial tissue.89 Additionally, BP induces apoptosis or necrosis of SFs in the joint cavity of RA,36 and promotes the M1-M2 phenotype transition in local macrophages,90 which significantly mitigates the inflammatory response and preserves metabolic homeostasis. This mechanism may be associated with the activation of the P53 apoptotic pathway in inflammatory cells by BP.91,92
The immunomodulatory effects of BP can be further enhanced by surface modification or drug loading. For instance, the surface modification of BP with titanium sulfonate ligand (TiL4) significantly reduces the secretion of pro-inflammatory factors by macrophages and inhibits the infiltration of peripheral neutrophils.35 Furthermore, the combination of BP with Icaritin (ICT) or the incorporation of procyanidin B2 (OPC B2) can effectively manage M1-type acute inflammation and promote the conversion of M1 (CD86+) macrophages to M2 (CD206+) macrophages,93,94 thereby contributing to the resolution of inflammation.
The biocompatibility of BP nanomaterials has been demonstrated by various studies. Assessment of apoptotic markers following the co-culture of BP with cells indicated that BP did not exhibit significant cytotoxicity.63,95 Although naked BP (BPQDs) resulted in a transient increase in the levels of peripheral neutrophils, pro-inflammatory factors, and inflammatory macrophages in the target organs of mice, the overall inflammatory response was found to be reversible, gradually returning to baseline levels over time.35,36 Furthermore, research has demonstrated that BPQDs can be effectively eliminated from the body via renal excretion,96 without causing damage to vital organs or inducing inflammatory lesions.68,96 These findings support the potential application of BPQDs in biomedical fields.
Anti-Inflammatory M2 Macrophages
The Direct Role of BP in Promoting M2 Polarization
In the dynamic regulation of the immune system, anti-inflammatory M2 macrophages typically emerge later in the immune response and play a crucial role in facilitating bone tissue repair. A substantial body of literature indicates that BP can target macrophages to induce M2 polarization and remodel the anti-inflammatory environment. For instance, in models of periodontitis and skull defects, BPNSs promote the reprogramming of M1 to M2 macrophages and enhance the secretion of M2-specific anti-inflammatory factors,19,49,97,98 (as illustrated in Figure 4),49 which contributes to improved disease prognosis. In the treatment of osteoporosis, BP facilitates the M1 to M2 transition while simultaneously inhibiting the differentiation of macrophage-derived osteoclasts,50 thereby guiding bone regeneration through general immunomodulation. In the treatment of infected and diabetic wounds, BP administration has been observed to drive the production of M2 (CD206) macrophages. Subsequently, these M2 macrophages release various active factors that facilitate angiogenesis and tissue healing.19,99–101 The promoting effect of BP composites on macrophage phenotypic transformation is further described in Table 2. Consequently, the polarization of M2 macrophages induced by BP is critical for bone regeneration. Furthermore, the PI3K-Akt signaling pathway has been shown to be a crucial pathway for M2 polarization,102 and the downstream factor mechanistic target of rapamycin complex 2 (mTORC2) serves as a marker for M2 cells.103 Evidence suggests that the intracellular PI3K/Akt signaling pathway activated by BP is intricately linked to the metabolic reprogramming between M1 and M2 macrophages.21,104,105 Therefore, it is hypothesized that BP may enhance M2 polarization via the PI3K-Akt-mTORC2 signaling axis, thereby modulating the bone immune microenvironment. However, further validation through extensive experimentation is required.
 |
Table 2 Promotional Effect of BP Composites on Macrophage Phenotype Transformation |
 |
Figure 4 BPNSs promote the metabolic reprogramming of macrophages in local periodontal inflammation. Administration of BPNS during aPDT-mediated periodontitis treatment protects against ROS-induced oxidative damage and polarizes pro-inflammatory M1 macrophages into anti-inflammatory M2 macrophages. Notes: Reproduced from Li X, Ren S, Song L, et al. Combined Black Phosphorus Nanosheets with ICG/aPDT is an Effective Anti-Inflammatory Treatment for Periodontal Disorders. Int J Nanomedicine. 2023 Feb 15;18:813-827. © 2023 Li et al.This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution – Non Commercial (unported, v3.0) License.49 |
Furthermore, it has been suggested that BP facilitates M2 polarization by activating the cellular autophagy pathway. Cellular autophagy, which occurs at basal levels in all tissues, plays a crucial role in alleviating oxidative stress and exhibiting anti-apoptotic effects.107 Previous studies have indicated that the autophagy pathway is involved in IL-10-induced M2 polarization.108,109 BP has been shown to induce cellular autophagy, as evidenced by the induction of autophagosome formation, upregulation of Beclin-1 gene expression and promotion of the conversion from LC3-I to LC3-II.110,111 The mechanism involved may be related to the activation of the PERK-ATF4-CHOP pathway.111 And BPQDs can activate macrophage autophagy to reduce the inflammatory response and promote lipid metabolism.112 Therefore, it can be posited that the process of BP-induced M2 polarization may be partially dependent on the activation of autophagy. However, the precise mechanisms involved require further investigation.
The Indirect Role of BP in Promoting M2 Polarization
In addition to the direct induction of M2 polarization, the indirect effects of BP on macrophages deserve considerable attention. First, PTT has been demonstrated to upregulate cytosolic integrin α4 and activate the PI3K-Akt1 signaling pathway, which subsequently facilitates M2 polarization.28 The induction of M2 polarization by BP is markedly enhanced when combined with PTT,93,113 ultimately accelerating bone remodeling. Second, electrical stimulation serves as an effective promoter of M2 polarization. BP-based electroactive hydrogels, when supported by electrical stimulation, can improve the immune microenvironment and inhibit the activation of inflammatory cells,67 thereby promoting M2 polarization, (as illustrated in Figure 5a–c)114 and aiding in the repair of tissue damage. Consequently, the exceptional photothermal and conductive properties of BP are advantageous for establishing an anti-inflammatory microenvironment and promoting tissue regeneration.
 |
Figure 5 Electroactive BP hydrogels inhibit cellular oxidative stress damage and promote macrophage M2 polarization.(a) Injection process of the BHGD hydrogel into the ischemic myocardium. (b) Preparation and properties of the BHGD hydrogel. (c) Modulation of the MI microenvironment and bioresorption mechanism of the BHGD hydrogel in vivo. Notes: Reproduced from Qiu Y, Yu C, Yue Z, et al. Chronological-programmed black phosphorus hydrogel for responsive modulation of the pathological microenvironment in myocardial infarction. ACS Appl Mater Interfaces. 2024. Copyright © 2024 American Chemical Society.114 |
Furthermore, BP has the ability to activate the intracellular antioxidant pathway, thereby inducing the production of M2 macrophages. The Nrf2/Keap pathway functions as a critical defense mechanism for cells, facilitating the maintenance of homeostasis and playing a significant role in the survival of tissue cells. Research has demonstrated that the Nrf2/Keap pathway promotes the differentiation of M2 macrophages115 while simultaneously antagonizing NF-κB receptors and activating PPARγ receptors,116 thereby indirectly modulating the immune microenvironment. Evidence indicates that BP activates the intracellular Nrf2 pathway,117 which contributes to the maintenance of tissue homeostasis and results in the upregulation of Nrf2 protein and heme oxygenase-1 (HO-1) expression. This enhancement in tissue metabolic activity ultimately fosters vascularization and neural regeneration.95,118 Concurrently, the increase in ROS also triggers the activation of the Nrf2/Keap pathway, which enhances cellular adaptation to oxidative stress.119 In conclusion, BP effectively activates the intracellular Nrf2/Keap pathway in response to oxidative stress, facilitating the establishment of an anti-inflammatory environment characterized by M2-type macrophages.
Furthermore, the metabolites of BP can indirectly enhance M2 polarization. The exceptional bioelectrical conductivity of BP increases the intracellular concentration of free Ca2+94 and promotes the formation of calcium phosphate through in situ phosphorus-driven calcium extraction.120 Calcium phosphate nanoparticles interact with the cell surface integrin β1, activating the PI3K/Akt signaling pathway, which induces M2 polarization and subsequent osteoblast differentiation.121 Consequently, both BP monomers and the metabolite calcium phosphate are capable of promoting M2 polarization, thereby accelerating the process of bone regeneration.
Finally, BP can serve as a drug reservoir to facilitate the conversion of M1 to M2 macrophages. For instance, the co-loading of graphene oxide (GO) layers and BPNSs with IL-4 can alleviate local inflammation while driving M2 polarization.122 Furthermore, the combination of carbon nanotubes (sdCNTs), BPNSs and dexamethasone (DEX) is used to induce osteogenesis, while regulating the transition to an M2 polarization and an anti-inflammatory environment,123 ultimately accelerating vascularization and osseointegration. Therefore, it is evident that BP can enhance the efficacy of bone immunomodulatory drugs, helping them to achieve their optimal therapeutic effects.
It is essential to recognize that the role of M2 macrophages cannot be generalized in a single statement. Regenerative repair associated with excessive M2 polarization may also contribute to the development of certain diseases. For instance, after skeletal muscle injury, TGFβ1-producing macrophages are directly linked to the aberrant differentiation of progenitor cells, which ultimately leading to heterotopic ossification (HO).124 Notably, BP has been shown to inhibit M2 polarization and preserve metabolic homeostasis under pathological conditions. For example, after acute ocular surface injury, BPQDs scavenge ROS and impede the production of intracellular neutrophil extracellular traps (NETs), thereby mitigating pathological corneal fibrosis associated with M2 polarization.125,126 In conclusion, BP does not exert a singular regulatory effect on macrophages; rather, it dynamically modulates the M1/M2 ratio and contributes to the maintenance of immune system homeostasis.
In summary, the immunomodulatory strategy of BP includes several key aspects. First, BP activates the body’s innate immune system. As the material is gradually degraded in vivo, BP effectively eliminates ROS that accumulate in inflammatory environments. This process inhibits the activation of inflammatory signaling pathways and the production of M1-type macrophages, thereby mitigating the inflammatory response at the site of infection. Second, BP promotes the production of M2-type macrophages, particularly when combined with PTT and electrical stimulation, ultimately fostering an immune environment conducive to bone tissue repair.
Osteoclasts
Monocytes and macrophages serve as the primary source of osteoclast differentiation. The overactivation of osteoclasts is linked to a reduction in bone mass85 and plays a pivotal role in inflammation-induced bone loss.127,128 Furthermore, osteoclasts can induce H-type angiogenesis, which enhances the communication and interaction between immune cells and tissues.129 Consequently, targeted modulation of osteoclasts represents a significant strategy for maintaining bone immune homeostasis and metabolic balance.
In addition to its role in regulating macrophage polarization, BP can also directly influence osteoclast differentiation, thereby affecting osseointegration. For instance, oxidized black phosphorus nanosheets (oBPNSs) have been shown to inhibit the phosphorylation of p38 mitogen-activated protein kinase (MAPK) in RAW264.7 cells and subsequently downregulate the NF-κB pathway and the expression of the NFATc1 gene. This effect ultimately leads to a reduction in RANKL-induced osteoclast differentiation.130 Additionally, the degradation product of BP, inorganic phosphate (Pi), exerts a negative feedback regulation on RANK-RANKL signaling, downregulates osteoclast gene expression, and antagonizes the activity of the NF-κB pathway.131 Furthermore, BP stimulates the secretion of IL-33 by macrophages, and elevated levels of IL-33 have been found to inhibit osteoclast formation while promoting environmental changes conducive to bone regeneration.20 In conclusion, osteoclast differentiation is intricately regulated by BP, a process that is crucial for enhancing the bone immune environment and facilitating bone mass accumulation. Moreover, both the reduced state and the oxidative degradation products of BP can modulate the bone immune environment in a manner that promotes beneficial bone regeneration, demonstrating the favorable biocompatibility and biosafety of the nanomaterials, which establishes a foundation for their advanced applications in biomedical fields.
Conclusion and Future Perspectives
Over the past few years, research on 2D nanomaterials has progressively deepened, leading to a paradigm shift in the field of BTE. This shift has transitioned from a singular emphasis on the osteogenic properties of materials to a more comprehensive investigation of their effects on the bone immune environment. BPNSs, as a novel class of nanomaterials, have exhibited significant potential in promoting bone regeneration and modulating immune responses within the bone. A substantial body of research has demonstrated that BPNSs can interact with the bone immune microenvironment to elicit a specific immune response. Furthermore, the inherent antioxidant properties of BP contribute to the establishment of an anti-inflammatory repair environment and facilitate targeted alterations in macrophage phenotypes, ultimately enhancing bone regeneration.
Recent studies have demonstrated that BP can also enhance the immune microenvironment surrounding nerves and indirectly facilitate innervated bone regeneration, thus filling a gap in its regulatory mechanisms in bone remodeling. For instance, magnesium ion-modified BPNSs (BP@Mg) have been shown to diminish the accumulation of inflammatory cells in infected environments, improve the functionality of Schwann cells (SC), and promote innervated bone regeneration.132,133 Meanwhile, BPNSs have been shown to activate the Nrf2 pathway in neural progenitor cells (NPCs), which directly modulates intracellular oxidative stress responses and promotes neural differentiation.117 Additionally, Studies have shown BP-mediated electrical stimulation effectively promotes the transformation of Schwann cells into an active repair phenotype and supports nerve-induced osteogenesis.134 In conclusion, there is considerable evidence that BP guides nerve regeneration during the early stages of bone repair through immune modulation.
Despite the established anti-inflammatory and pro-repair properties of BP, several critical challenges remain that must be addressed. Current research trends predominantly emphasize the role of BP-induced polarization of M2-type macrophages in bone regeneration. However, it is essential to acknowledge that during the actual bone healing process, both M1- and M2-type macrophages work synergistically to enhance stem cell recruitment, promote osteoblastic activity, and facilitate matrix mineralization, ultimately leading to effective bone remodeling. Consequently, it is imperative to elucidate the differential regulatory effects of BP on M1 and M2 macrophages at all stages of bone healing, as well as their respective contributions to bone remodeling, in order to develop optimal strategies for bone regeneration. Furthermore, the bone immune microenvironment is a dynamic and self-regulating ecosystem in which a diverse array of immune cells collaborate to maintain environmental homeostasis. Significant gaps remain in our understanding of the effects of BP on other immune cell types, including neutrophils, mast cells, monocytes, dendritic cells, and lymphocytes, as well as their collective regulation of osteogenic activity, which warrants further investigation.
Furthermore, the mechanisms by which BP nanomaterials influence immune cell metabolism remain inadequately understood. Research indicates that the regulation of bone immune homeostasis is fundamentally linked to the modulation of metabolic pathways in immune cells. Specifically, M0 and M2 macrophages predominantly utilize the tricarboxylic acid cycle and oxidative phosphorylation to sustain their cellular functions, whereas M1 macrophages exhibit a greater reliance on glycolysis, which facilitates a prompt and robust inflammatory response.26,135 Additionally, the hypermetabolic profile characteristic of M1 macrophages is closely associated with a refractory inflammatory environment.103 Evidence suggests that BP nanomaterials specifically target the respiratory metabolism of immune cells, promoting M2 polarization in macrophages21,104 and enhancing the immune response of natural killer (NK) cells.136 Consequently, the regulation of immune cell metabolism to induce alterations in cellular phenotype and function, thereby fostering a favourable immune microenvironment, may represent a promising avenue for future research into the role of BP in bone immunomodulation.
Finally, the biological application limitations of BP nanomaterials cannot be overlooked. These limitations include a rapid degradation rate in vivo, cytotoxicity to specific cell types, and the properties of BP nanomaterials—such as surface morphology, surface charge, porosity, wettability, and particle size—that significantly influence their interactions with immune cells. Therefore, only through appropriate surface and chemical modifications can the degradation of BP be delayed, and protein adsorption and subsequent immune responses be precisely regulated, thereby optimizing bone regeneration. However, current research on the combined applications of multiple biomaterials remains limited. It is still to be determined which dimensions of BPNSs are most effective for bone immunomodulation and regeneration, how to align the optimal timing for immune modulation with the degradation rate of materials through surface modification, and the impact of biomaterial modifications on immune cell activities. Consequently, future research should prioritize the joint application of BP and various nanomaterials to advance the development of bone tissue engineering by leveraging the complementary strengths of multiple materials.
Data Sharing Statement
Data availability is not applicable to this article as no new data were created or analyzed in this study.
Acknowledgments
This work was supported by the National Natural Science Foundation of China, Grant/AwardNumber: 81801008; Science Research Cultivation Program of Stomatological Hospital, Southern Medical University, Grant/Award Number: PY2024024.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Disclosure
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
References
1. Wang M, Li Y, Su Z, Mo A. Clinical and radiographic outcomes of customized allogeneic bone block versus autogenous bone block for ridge augmentation: 6 Month results of a randomized controlled clinical trial. J Clin Periodontol. 2023;50(1):22–35. doi:10.1111/jcpe.13714
2. Heimes D, Pabst A, Becker P, et al. Comparison of morbidity-related parameters between autologous and allogeneic bone grafts for alveolar ridge augmentation from patients’ perspective-A questionnaire-based cohort study. Clin Implant Dent Relat Res. 2024;26(1):170–182. doi:10.1111/cid.13242
3. Wang C, Huang W, Zhou Y, et al. 3D printing of bone tissue engineering scaffolds. Bioact Mater. 2020;5(1):82–91. doi:10.1016/j.bioactmat.2020.01.004
4. Wang Z, Zhao J, Tang W, et al. Multifunctional nanoengineered hydrogels consisting of black phosphorus nanosheets upregulate bone formation. Small. 2019;15(41):e1901560. doi:10.1002/smll.201901560
5. Shi H, Zhou K, Wang M, et al. Integrating physicomechanical and biological strategies for BTE: biomaterials-induced osteogenic differentiation of MSCs. Theranostics. 2023;13(10):3245–3275. doi:10.7150/thno.84759
6. Miao Y, Chen Y, Luo J, et al. Black phosphorus nanosheets-enabled DNA hydrogel integrating 3D-printed scaffold for promoting vascularized bone regeneration. Bioact Mater. 2023;21:97–109. doi:10.1016/j.bioactmat.2022.08.005
7. Batoon L, Millard SM, Wullschleger ME, et al. CD169(+) macrophages are critical for osteoblast maintenance and promote intramembranous and endochondral ossification during bone repair. Biomaterials. 2019;196:51–66. doi:10.1016/j.biomaterials.2017.10.033
8. Mohamad SF, Gunawan A, Blosser R, et al. Neonatal osteomacs and bone marrow macrophages differ in phenotypic marker expression and function. J Bone Miner Res. 2021;36(8):1580–1593. doi:10.1002/jbmr.4314
9. Gibon E, Lu LY, Nathan K, Goodman SB. Inflammation, ageing, and bone regeneration. J Orthop Translat. 2017;10:28–35. doi:10.1016/j.jot.2017.04.002
10. Zhang J, Akiyama K, Mun AY, et al. Age-related effects on msc immunomodulation, macrophage polarization, apoptosis, and bone regeneration correlate with IL-38 expression. Int J mol Sci. 2024;25(6). doi:10.3390/ijms25063252
11. Quero L, Tiaden AN, Hanser E, et al. miR-221-3p drives the shift of M2-macrophages to a pro-inflammatory function by suppressing JAK3/STAT3 activation. Front Immunol. 2019;10:3087. doi:10.3389/fimmu.2019.03087
12. Cui Y, Hong S, Xia Y, et al. Melatonin engineering M2 macrophage-derived exosomes mediate endoplasmic reticulum stress and immune reprogramming for periodontitis therapy. Adv Sci. 2023;10(27):e2302029. doi:10.1002/advs.202302029
13. Lee J, Byun H, Madhurakkat Perikamana SK, Lee S, Shin H. Current advances in immunomodulatory biomaterials for bone regeneration. Adv Healthc Mater. 2019;8(4):e1801106. doi:10.1002/adhm.201801106
14. Qiao W, Xie H, Fang J, et al. Sequential activation of heterogeneous macrophage phenotypes is essential for biomaterials-induced bone regeneration. Biomaterials. 2021;276:121038. doi:10.1016/j.biomaterials.2021.121038
15. Hou Y, Fei Y, Liu Z, Liu Y, Li M, Luo Z. Black phosphorous nanomaterials as a new paradigm for postoperative tumor treatment regimens. J Nanobiotechnol. 2022;20(1):366. doi:10.1186/s12951-022-01579-3
16. Liu W, Zhu Y, Tao Z, Chen Y, Zhang L, Dong A. Black phosphorus-based conductive hydrogels assisted by electrical stimulus for skin tissue engineering. Adv Healthc Mater. 2023;12(29):e2301817. doi:10.1002/adhm.202301817
17. Huang K, Wu J, Gu Z. Black phosphorus hydrogel scaffolds enhance bone regeneration via a sustained supply of calcium-free phosphorus. ACS Appl Mater Interfaces. 2019;11(3):2908–2916. doi:10.1021/acsami.8b21179
18. Miao Y, Liu X, Luo J, Yang Q, Chen Y, Wang Y. Double-network DNA macroporous hydrogel enables aptamer-directed cell recruitment to accelerate bone healing. Adv Sci. 2024;11(1):e2303637. doi:10.1002/advs.202303637
19. He Y, Tang Y, Zeng B, et al. Black phosphorus quantum dot-modified ADSCs as a novel therapeutic for periodontitis bone loss coupling of osteogenesis and osteoimmunomodulation. Mater Today Bio. 2024;27:101122. doi:10.1016/j.mtbio.2024.101122
20. Qiu M, Tulufu N, Tang G, et al. Black phosphorus accelerates bone regeneration based on immunoregulation. Adv Sci. 2024;11(1):e2304824. doi:10.1002/advs.202304824
21. Long J, Yao Z, Zhang W, et al. Regulation of osteoimmune microenvironment and osteogenesis by 3D-printed PLAG/black phosphorus scaffolds for bone regeneration. Adv Sci. 2023;10(28):e2302539. doi:10.1002/advs.202302539
22. Wang Q, Xu L, Willumeit-Römer R, Luthringer-Feyerabend BJC. Macrophage-derived oncostatin M/bone morphogenetic protein 6 in response to Mg-based materials influences pro-osteogenic activity of human umbilical cord perivascular cells. Acta Biomater. 2021;133:268–279. doi:10.1016/j.actbio.2020.12.016
23. Park-Min KH. Mechanisms involved in normal and pathological osteoclastogenesis. Cell mol Life Sci. 2018;75(14):2519–2528. doi:10.1007/s00018-018-2817-9
24. Liang H, Chen K, Xie J, et al. A bone-penetrating precise controllable drug release system enables localized treatment of osteoporotic fracture prevention via modulating osteoblast-osteoclast communication. Small. 2023;19(26):e2207195. doi:10.1002/smll.202207195
25. Hu Y, Huang J, Chen C, et al. Strategies of macrophages to maintain bone homeostasis and promote bone repair: a narrative review. J Funct Biomater. 2022;14(1):18. doi:10.3390/jfb14010018
26. Ishida K, Nagatake T, Saika A, et al. Induction of unique macrophage subset by simultaneous stimulation with LPS and IL-4. Front Immunol. 2023;14:1111729. doi:10.3389/fimmu.2023.1111729
27. Yu H, Wang H, Liu J, Huang T, Man Y, Xiang L. The effect of ROS-YAP crosstalk on osteoimmune response orchestrating osteogenesis. Cell Cycle. 2023;22(11):1391–1405. doi:10.1080/15384101.2023.2211830
28. Li B, Liu F, Ye J, et al. Regulation of macrophage polarization through periodic photo-thermal treatment to facilitate osteogenesis. Small. 2022;18(38):e2202691. doi:10.1002/smll.202202691
29. Kang M, Huang CC, Lu Y, et al. Bone regeneration is mediated by macrophage extracellular vesicles. Bone. 2020;141:115627. doi:10.1016/j.bone.2020.115627
30. Yamashita M, Passegué E. TNF-α coordinates hematopoietic stem cell survival and myeloid regeneration. Cell Stem Cell. 2019;25(3):357–372.e7. doi:10.1016/j.stem.2019.05.019
31. Liang B, Wang H, Wu D, Wang Z. Macrophage M1/M2 polarization dynamically adapts to changes in microenvironment and modulates alveolar bone remodeling after dental implantation. J Leukoc Biol. 2021;110(3):433–447. doi:10.1002/jlb.1ma0121-001r
32. Wu H, Yin Y, Hu X, et al. Effects of environmental pH on macrophage polarization and osteoimmunomodulation. ACS Biomater Sci Eng. 2019;5(10):5548–5557. doi:10.1021/acsbiomaterials.9b01181
33. Li Z, Cai F, Tang J, et al. Oxygen metabolism-balanced engineered hydrogel microspheres promote the regeneration of the nucleus pulposus by inhibiting acid-sensitive complexes. Bioact Mater. 2023;24:346–360. doi:10.1016/j.bioactmat.2022.12.025
34. Lu H, Wei J, Liu K, et al. Radical-scavenging and subchondral bone-regenerating nanomedicine for osteoarthritis treatment. ACS Nano. 2023;17(6):6131–6146. doi:10.1021/acsnano.3c01789
35. Qu G, Liu W, Zhao Y, et al. Improved biocompatibility of black phosphorus nanosheets by chemical modification. Angew Chem Int Ed Engl. 2017;56(46):14488–14493. doi:10.1002/anie.201706228
36. Zhao C, Song W, Ma J, Wang N. Macrophage-derived hybrid exosome-mimic nanovesicles loaded with black phosphorus for multimodal rheumatoid arthritis therapy. Biomater Sci. 2022;10(23):6731–6739. doi:10.1039/d2bm01274j
37. Martin KE, García AJ. Macrophage phenotypes in tissue repair and the foreign body response: implications for biomaterial-based regenerative medicine strategies. Acta Biomater. 2021;133:4–16. doi:10.1016/j.actbio.2021.03.038
38. Meizlish ML, Franklin RA, Zhou X, Medzhitov R. Tissue homeostasis and inflammation. Annu Rev Immunol. 2021;39:557–581. doi:10.1146/annurev-immunol-061020-053734
39. Dai K, Geng Z, Zhang W, et al. Biomaterial design for regenerating aged bone: materiobiological advances and paradigmatic shifts. Natl Sci Rev. 2024;11(5):nwae076. doi:10.1093/nsr/nwae076
40. Shen C, Ding X, Ruan J, et al. Black phosphorus quantum dots induce myocardial inflammatory responses and metabolic disorders in mice. J Environ Sci. 2024;137:53–64. doi:10.1016/j.jes.2023.01.004
41. Zhao Y, Chen Z, Shao W, et al. Black phosphorus-enhanced injectable hydrogel for infected soft tissue healing. APL Bioeng. 2023;7(1):016103. doi:10.1063/5.0121241
42. Bai X, Wang R, Hu X, et al. Two-dimensional biodegradable black phosphorus nanosheets promote large full-thickness wound healing through in situ regeneration therapy. ACS Nano. 2024;18(4):3553–3574. doi:10.1021/acsnano.3c11177
43. Dwyer GK, D’Cruz LM, Turnquist HR. Emerging functions of IL-33 in homeostasis and immunity. Annu Rev Immunol. 2022;40:15–43. doi:10.1146/annurev-immunol-101320-124243
44. Wang M, Gao M, Yi Z. Biological effects of IL-33/ST2 axis on oral diseases: autoimmune diseases and periodontal diseases. Int Immunopharmacol. 2023;122:110524. doi:10.1016/j.intimp.2023.110524
45. de Lima MHF, Hiroki CH, de Fátima Borges V, et al. Sepsis-induced immunosuppression is marked by an expansion of a highly suppressive repertoire of FOXP3+ T-regulatory cells expressing TIGIT. J Infect Dis. 2022;225(3):531–541. doi:10.1093/infdis/jiab405
46. Bertschi NL, Steck O, Luther F, et al. PPAR-γ regulates the effector function of human T helper 9 cells by promoting glycolysis. Nat Commun. 2023;14(1):2471. doi:10.1038/s41467-023-38233-x
47. Luo W, Xu Q, Wang Q, Wu H, Hua J. Effect of modulation of PPAR-γ activity on Kupffer cells M1/M2 polarization in the development of non-alcoholic fatty liver disease. Sci Rep. 2017;7:44612. doi:10.1038/srep44612
48. Lee YJ, Kim K, Kim M, Ahn YH, Kang JL. Inhibition of STAT6 activation by AS1517499 inhibits expression and activity of PPARγ in macrophages to resolve acute inflammation in mice. Biomolecules. 2022;12(3). doi:10.3390/biom12030447
49. Li X, Ren S, Song L, et al. Combined black phosphorus nanosheets with ICG/aPDT is an effective anti-inflammatory treatment for periodontal disorders. Int J Nanomed. 2023;18:813–827. doi:10.2147/ijn.S394861
50. Wang J, Ye J, Yang G, et al. Fenton-like reaction inspired “·OH catalyzed” osteogenic process for the treatment of osteoporosis. Adv Healthc Mater. 2024;13(15):e2304091. doi:10.1002/adhm.202304091
51. Guo F, Li J, Chen Z, et al. An injectable black phosphorus hydrogel for rapid tooth extraction socket healing. ACS Appl Mater Interfaces. 2024. doi:10.1021/acsami.4c03278
52. Fasolino I, Soriente A, Caporali M, et al. 2D exfoliated black phosphorus influences healthy and cancer prostate cell behaviors. Sci Rep. 2021;11(1):5856. doi:10.1038/s41598-021-85310-6
53. Huang X, Wu B, Li J, et al. Anti-tumour effects of red blood cell membrane-camouflaged black phosphorous quantum dots combined with chemotherapy and anti-inflammatory therapy. Artif Cells Nanomed Biotechnol. 2019;47(1):968–979. doi:10.1080/21691401.2019.1584110
54. Liu L, Wang W, Hong W, et al. Photothermal 2D nanosheets combined with astragaloside IV for antibacterial properties and promoting angiogenesis to treat infected wounds. Front Bioeng Biotechnol. 2021;9:826011. doi:10.3389/fbioe.2021.826011
55. Wang X, Tang X, Li N, et al. A multifunctional black phosphorus-based adhesive patch intrinsically induces partial EMT for effective burn wound healing. Biomater Sci. 2022;11(1):235–247. doi:10.1039/d2bm01625g
56. Huang S, Xu S, Hu Y, et al. Preparation of NIR-responsive, ROS-generating and antibacterial black phosphorus quantum dots for promoting the MRSA-infected wound healing in diabetic rats. Acta Biomater. 2022;137:199–217. doi:10.1016/j.actbio.2021.10.008
57. Zhang Y, Chen W, Feng W, Fang W, Han X, Cheng C. Multifunctional chondroitin sulfate based hydrogels for promoting infected diabetic wounds healing by chemo-photothermal antibacterial and cytokine modulation. Carbohydr Polym. 2023;314:120937. doi:10.1016/j.carbpol.2023.120937
58. Tukaj S. Heat shock protein 70 as a double agent acting inside and outside the cell: insights into autoimmunity. Int J mol Sci. 2020;21(15). doi:10.3390/ijms21155298
59. Tong L, Liao Q, Zhao Y, et al. Near-infrared light control of bone regeneration with biodegradable photothermal osteoimplant. Biomaterials. 2019;193:1–11. doi:10.1016/j.biomaterials.2018.12.008
60. Li W, Li S, Zhang J, et al. Fabrication and evaluation of bone morphogenetic protein-2 microspheres coated black phosphorus nanosheets@polylactic-glycolic acid copolymers scaffold: a multifunctional antibacterial photothermal scaffold for bone regeneration. Int J Biol Macromol. 2022;210:350–364. doi:10.1016/j.ijbiomac.2022.05.028
61. Zhao Y, Peng X, Xu X, et al. Chitosan based photothermal scaffold fighting against bone tumor-related complications: recurrence, infection, and defects. Carbohydr Polym. 2023;300:120264. doi:10.1016/j.carbpol.2022.120264
62. Yang J, Liu W, Sun Y, Dong X. LVFFARK-PEG-stabilized black phosphorus nanosheets potently inhibit amyloid-β fibrillogenesis. Langmuir. 2020;36(7):1804–1812. doi:10.1021/acs.langmuir.9b03612
63. Cheng G, Li Z, Liu Y, et al. “Swiss Army Knife” black phosphorus-based nanodelivery platform for synergistic antiparkinsonian therapy via remodeling the brain microenvironment. J Control Release. 2023;353:752–766. doi:10.1016/j.jconrel.2022.12.024
64. Liu RR, Zhai HL, Shao HP, Wang TH. Biological effect of black phosphorus nanosheets on the interaction between SARS-CoV-2 S protein and ACE2. Phys Chem Chem Phys. 2022;24(44):27388–27393. doi:10.1039/d2cp03994j
65. Luan X, Zhang X, Nie M, Zhao Y. Traditional Chinese medicine integrated responsive microneedles for systemic sclerosis treatment. Research. 2023;6:0141. doi:10.34133/research.0141
66. Ouyang J, Ji X, Zhang X, et al. In situ sprayed NIR-responsive, analgesic black phosphorus-based gel for diabetic ulcer treatment. Proc Natl Acad Sci U S A. 2020;117(46):28667–28677. doi:10.1073/pnas.2016268117
67. Liu M, Zhang W, Han S, et al. Multifunctional conductive and electrogenic hydrogel repaired spinal cord injury via immunoregulation and enhancement of neuronal differentiation. Adv Mater. 2024;36(21):e2313672. doi:10.1002/adma.202313672
68. Geng C, He S, Yu S, et al. Achieving clearance of drug-resistant bacterial infection and rapid cutaneous wound regeneration using an ROS-balancing-engineered heterojunction. Adv Mater. 2024;36(16):e2310599. doi:10.1002/adma.202310599
69. He Z, Chen W, Hu K, et al. Resolvin D1 delivery to lesional macrophages using antioxidative black phosphorus nanosheets for atherosclerosis treatment. Nat Nanotechnol. 2024;19(9):1386–1398. doi:10.1038/s41565-024-01687-1
70. Wang D, Zhao Q, Qin J, Guo Y, Zhang C, Li Y. Urokinase loaded black phosphorus nanosheets for sequential thrombolysis and reactive oxygen species scavenging in ischemic stroke treatment. Biomater Sci. 2022;10(16):4656–4666. doi:10.1039/d2bm00746k
71. Hou J, Wang H, Ge Z, et al. Treating acute kidney injury with antioxidative black phosphorus nanosheets. Nano Lett. 2020;20(2):1447–1454. doi:10.1021/acs.nanolett.9b05218
72. Zhang J, Sun D, Liao Y, et al. Time-released black phosphorus hydrogel accelerates myocardial repairing through antioxidant and motivates macrophage polarization properties. Biomater Res. 2024;28:0029. doi:10.34133/bmr.0029
73. Xie DM, Sun C, Tu Q, et al. Modified black phosphorus quantum dots promotes spinal cord injury repair by targeting the AKT signaling pathway. J Tissue Eng. 2023;14:20417314231180033. doi:10.1177/20417314231180033
74. Gao X, Wang B, Li J, et al. Catalytic tunable black phosphorus/ceria nanozyme: a versatile oxidation cycle accelerator for alleviating cisplatin-induced acute kidney injury. Adv Healthc Mater. 2023;12(30):e2301691. doi:10.1002/adhm.202301691
75. Mo J, Xu Y, Wang X, Wei W, Zhao J. Exploiting the protein Corona: coating of black phosphorus nanosheets enables macrophage polarization via calcium influx. Nanoscale. 2020;12(3):1742–1748. doi:10.1039/c9nr08570j
76. Mo J, Xie Q, Wei W, Zhao J. Revealing the immune perturbation of black phosphorus nanomaterials to macrophages by understanding the protein Corona. Nat Commun. 2018;9(1):2480. doi:10.1038/s41467-018-04873-7
77. Guo C, Zhao X, Ma R, et al. Silica nanoparticles promoted pro-inflammatory macrophage and foam cell transformation via ROS/PPARγ/NF-κB signaling. Sci Total Environ. 2023;881:163430. doi:10.1016/j.scitotenv.2023.163430
78. Liao Z, Ma X, Kai J-J, Fan J. Molecular mechanisms of integrin αvβ8 activation regulated by graphene, boron nitride and black phosphorus nanosheets. Colloids Surf B Biointerfaces. 2023;222:113139. doi:10.1016/j.colsurfb.2023.113139
79. Ruan F, Zeng J, Yin H, et al. RNA m6A modification alteration by black phosphorus quantum dots regulates cell ferroptosis: implications for nanotoxicological assessment. Small Methods. 2021;5(3):e2001045. doi:10.1002/smtd.202001045
80. Wang Y, Quan F, Cao Q, et al. Quercetin alleviates acute kidney injury by inhibiting ferroptosis. J Adv Res. 2021;28:231–243. doi:10.1016/j.jare.2020.07.007
81. Zhan YL, Wen KC, Li ZA, Sun P, Li FQ. Dielectric and magnetic composites of Fe(3)O(4)@APNs for superior microwave thermal effect. ACS Biomater Sci Eng. 2023;10(2):791–799. doi:10.1021/acsbiomaterials.3c01341
82. Zhang X, Tang J, Li C, Lu Y, Cheng L, Liu J. A targeting black phosphorus nanoparticle based immune cells nano-regulator for photodynamic/photothermal and photo-immunotherapy. Bioact Mater. 2021;6(2):472–489. doi:10.1016/j.bioactmat.2020.08.024
83. Wu Q, Ma Q, Ma J, et al. Cascade amplification of pyroptosis and apoptosis for cancer therapy through a black phosphorous-doped thermosensitive hydrogel. Pharmaceutics. 2023;15(7). doi:10.3390/pharmaceutics15071830
84. Li W, Chen P, Pan Y, et al. Construction of a band-aid like cardiac patch for myocardial infarction with controllable H(2) S release. Adv Sci. 2022;9(35):e2204509. doi:10.1002/advs.202204509
85. Yao Y, Cai X, Ren F, et al. The macrophage-osteoclast axis in osteoimmunity and osteo-related diseases. Front Immunol. 2021;12:664871. doi:10.3389/fimmu.2021.664871
86. Symons RA, Colella F, Collins FL, et al. Targeting the IL-6-Yap-Snail signalling axis in synovial fibroblasts ameliorates inflammatory arthritis. Ann Rheum Dis. 2022;81(2):214–224. doi:10.1136/annrheumdis-2021-220875
87. Damerau A, Kirchner M, Pfeiffenberger M, et al. Metabolic reprogramming of synovial fibroblasts in osteoarthritis by inhibition of pathologically overexpressed pyruvate dehydrogenase kinases. Metab Eng. 2022;72:116–132. doi:10.1016/j.ymben.2022.03.006
88. Saeki N, Imai Y. Reprogramming of synovial macrophage metabolism by synovial fibroblasts under inflammatory conditions. Cell Commun Signal. 2020;18(1):188. doi:10.1186/s12964-020-00678-8
89. Pan W, Dai C, Li Y, et al. PRP-chitosan thermoresponsive hydrogel combined with black phosphorus nanosheets as injectable biomaterial for biotherapy and phototherapy treatment of rheumatoid arthritis. Biomaterials. 2020;239:119851. doi:10.1016/j.biomaterials.2020.119851
90. Zhuang C, Sun R, Zhang Y, et al. Treatment of rheumatoid arthritis based on the inherent bioactivity of black phosphorus nanosheets. Aging Dis. 2024. doi:10.14336/ad.2024.0319
91. Cao X, Fu M, Du Q, Chang Z. Developmental toxicity of black phosphorus quantum dots in zebrafish (Danio rerio) embryos. Chemosphere. 2023;335:139029. doi:10.1016/j.chemosphere.2023.139029
92. Dong H, Wen Y, Lin J, et al. Cytotoxicity induced by black phosphorus nanosheets in vascular endothelial cells via oxidative stress and apoptosis activation. J Funct Biomater. 2023;14(5):284. doi:10.3390/jfb14050284
93. Xue P, Chang Z, Chen H, et al. Macrophage membrane (MMs) camouflaged near-infrared (NIR) responsive bone defect area targeting nanocarrier delivery system (BTNDS) for rapid repair: promoting osteogenesis via phototherapy and modulating immunity. J Nanobiotechnol. 2024;22(1):87. doi:10.1186/s12951-024-02351-5
94. Zhou LP, Pi W, Cheng SY, et al. Multifunctional DNA hydrogels with hydrocolloid-cotton structure for regeneration of diabetic infectious wounds. Adv Funct Mater. 2021;31(48):2106167. doi:10.1002/adfm.202106167
95. Qian Y, Yuan WE, Cheng Y, Yang Y, Qu X, Fan C. Concentrically integrative bioassembly of a three-dimensional black phosphorus nanoscaffold for restoring neurogenesis, angiogenesis, and immune homeostasis. Nano Lett. 2019;19(12):8990–9001. doi:10.1021/acs.nanolett.9b03980
96. Guo T, Wu Y, Lin Y, et al. Black phosphorus quantum dots with renal clearance property for efficient photodynamic therapy. Small. 2018;14(4). doi:10.1002/smll.201702815
97. Wang Y, Yuan Y, Wang R, et al. Injectable thermosensitive Gel CH-BPNs-NBP for effective periodontitis treatment through ROS-scavenging and jaw vascular unit protection. Adv Healthc Mater. 2024;13(22):e2400533. doi:10.1002/adhm.202400533
98. Huang JF, Tan QC, Bai H, et al. Harnessing immunomodulation for efficient bone regeneration: bioengineered black phosphorus-incorporated self-healing hydrogel. Chem Eng J. 2023:470144117. 10.1016/j.cej.2023.144117
99. Zhou L, Zhou L, Wei C, Guo R. A bioactive dextran-based hydrogel promote the healing of infected wounds via antibacterial and immunomodulatory. Carbohydr Polym. 2022;291:119558. doi:10.1016/j.carbpol.2022.119558
100. Zhao Y, Tian C, Liu Y, et al. All-in-one bioactive properties of photothermal nanofibers for accelerating diabetic wound healing. Biomaterials. 2023;295:122029. doi:10.1016/j.biomaterials.2023.122029
101. Luo X, Zhang L, Luo YP, et al. Charge-driven self-assembled microspheres hydrogel scaffolds for combined drug delivery and photothermal therapy of diabetic wounds. Adv Funct Mater. 2023;33(26). doi:10.1002/adfm.202214036
102. Xiao L, Liu H, Huang H, et al. 3D nanofiber scaffolds from 2D electrospun membranes boost cell penetration and positive host response for regenerative medicine. J Nanobiotechnol. 2024;22(1):322. doi:10.1186/s12951-024-02578-2
103. Wang S, Liu R, Yu Q, Dong L, Bi Y, Liu G. Metabolic reprogramming of macrophages during infections and cancer. Cancer Lett. 2019;452:14–22. doi:10.1016/j.canlet.2019.03.015
104. Zhou L, Liu N, Feng L, et al. Multifunctional electrospun asymmetric wettable membrane containing black phosphorus/Rg1 for enhancing infected wound healing. Bioeng Transl Med. 2022;7(2):e10274. doi:10.1002/btm2.10274
105. Li Q, Wang R, Xue J, et al. ZIF-8-modified black phosphorus nanosheets incorporated into injectable dual-component hydrogels for enhanced photothermal antibacterial and osteogenic activities. ACS Appl Mater Interfaces. 2024;16(25):32058–32077. doi:10.1021/acsami.4c05298
106. Su X, Chi Y, Liu T, et al. Black phosphorus nanosheets-based hydrogel for efficient bacterial inhibition and accelerating wound healing. ACS Appl Mater Interfaces. 2024;16(33):43892–43906. doi:10.1021/acsami.4c06075
107. Prerna K, Dubey VK. Beclin1-mediated interplay between autophagy and apoptosis: new understanding. Int J Biol Macromol. 2022;204:258–273. doi:10.1016/j.ijbiomac.2022.02.005
108. Sanjurjo L, Aran G, Téllez É, et al. CD5L promotes M2 macrophage polarization through autophagy-mediated upregulation of ID3. Front Immunol. 2018;9:480. doi:10.3389/fimmu.2018.00480
109. Liu K, Zhao E, Ilyas G, et al. Impaired macrophage autophagy increases the immune response in obese mice by promoting proinflammatory macrophage polarization. Autophagy. 2015;11(2):271–284. doi:10.1080/15548627.2015.1009787
110. Shang Y, Wang Q, Wu B, et al. Platelet-membrane-camouflaged black phosphorus quantum dots enhance anticancer effect mediated by apoptosis and autophagy. ACS Appl Mater Interfaces. 2019;11(31):28254–28266. doi:10.1021/acsami.9b04735
111. Wang L, Lin M, Hou X, et al. Black phosphorus quantum dots induce autophagy and apoptosis of human bronchial epithelial cells via endoplasmic reticulum stress. Chemosphere. 2023;327:138463. doi:10.1016/j.chemosphere.2023.138463
112. Ji Y, Wen Y, Zhang S, et al. Black phosphorus quantum dots prevent atherosclerosis in high-fat diet-fed apolipoprotein E knockout mice. Aging. 2024;16(13):10784–10798. doi:10.18632/aging.205874
113. Wu M, Liu H, Li D, et al. Smart-responsive multifunctional therapeutic system for improved regenerative microenvironment and accelerated bone regeneration via mild photothermal therapy. Adv Sci. 2024;11(2):e2304641. doi:10.1002/advs.202304641
114. Qiu Y, Yu C, Yue Z, et al. Chronological-programmed black phosphorus hydrogel for responsive modulation of the pathological microenvironment in myocardial infarction. ACS Appl Mater Interfaces. 2024. doi:10.1021/acsami.4c01956
115. Sha W, Zhao B, Wei H, et al. Astragalus polysaccharide ameliorates vascular endothelial dysfunction by stimulating macrophage M2 polarization via potentiating Nrf2/HO-1 signaling pathway. Phytomedicine. 2023;112:154667. doi:10.1016/j.phymed.2023.154667
116. Luo J, Wang J, Zhang J, et al. Nrf2 deficiency exacerbated CLP-induced pulmonary injury and inflammation through autophagy- and NF-κB/PPARγ-mediated macrophage polarization. Cells. 2022;11(23):3927. doi:10.3390/cells11233927
117. He FM, Cheng K, Qi JY, et al. Black phosphorus nanosheets enhance differentiation of neural progenitor cells for improved treatment in spinal cord injury. Chem Eng J. 2023:472144977. 10.1016/j.cej.2023.144977
118. Tan H, Cao K, Zhao Y, et al. Brain-targeted black phosphorus-based nanotherapeutic platform for enhanced hypericin delivery in depression. Small. 2024;20(31):e2310608. doi:10.1002/smll.202310608
119. Nguyen LXT, Troadec E, Kalvala A, et al. The Bcl-2 inhibitor venetoclax inhibits Nrf2 antioxidant pathway activation induced by hypomethylating agents in AML. J Cell Physiol. 2019;234(8):14040–14049. doi:10.1002/jcp.28091
120. Yang B, Yin J, Chen Y, et al. 2D-black-phosphorus-reinforced 3D-printed scaffolds: a stepwise countermeasure for osteosarcoma. Adv Mater. 2018;30(10). doi:10.1002/adma.201705611
121. Li M, Guo X, Qi W, et al. Macrophage polarization plays roles in bone formation instructed by calcium phosphate ceramics. J Mater Chem B. 2020;8(9):1863–1877. doi:10.1039/c9tb02932j
122. Liu X, Gaihre B, Park S, et al. 3D-printed scaffolds with 2D hetero-nanostructures and immunomodulatory cytokines provide pro-healing microenvironment for enhanced bone regeneration. Bioact Mater. 2023;27:216–230. doi:10.1016/j.bioactmat.2023.03.021
123. Liu X, Astudillo Potes MD, Dashtdar B, et al. 3D stem cell spheroids with 2D hetero-nanostructures for in vivo osteogenic and immunologic modulated bone repair. Adv Healthc Mater. 2024;13(12):e2303772. doi:10.1002/adhm.202303772
124. Sorkin M, Huber AK, Hwang C, et al. Regulation of heterotopic ossification by monocytes in a mouse model of aberrant wound healing. Nat Commun. 2020;11(1):722. doi:10.1038/s41467-019-14172-4
125. Eghbalzadeh K, Georgi L, Louis T, et al. Compromised anti-inflammatory action of neutrophil extracellular traps in PAD4-deficient mice contributes to aggravated acute inflammation after myocardial infarction. Front Immunol. 2019;10:2313. doi:10.3389/fimmu.2019.02313
126. Zhang J, Su L, Liu Z, et al. A responsive hydrogel modulates innate immune cascade fibrosis to promote ocular surface reconstruction after chemical injury. J Control Release. 2024;365:1124–1138. doi:10.1016/j.jconrel.2023.12.025
127. Andreev D, Liu M, Weidner D, et al. Osteocyte necrosis triggers osteoclast-mediated bone loss through macrophage-inducible C-type lectin. J Clin Invest. 2020;130(9):4811–4830. doi:10.1172/jci134214
128. Yu B, Gao Q, Sheng S, et al. Smart osteoclasts targeted nanomedicine based on amorphous CaCO(3) for effective osteoporosis reversal. J Nanobiotechnol. 2024;22(1):153. doi:10.1186/s12951-024-02412-9
129. Zhang H, Wang L, Cui J, et al. Maintaining hypoxia environment of subchondral bone alleviates osteoarthritis progression. Sci Adv. 2023;9(14):eabo7868. doi:10.1126/sciadv.abo7868
130. Xu Y, Mo JB, Wei W, Zhao J. Oxidized black phosphorus nanosheets as an inorganic antiresorptive agent. Ccs Chemistry. 2021;3(4):1105–1115. doi:10.31635/ccschem.020.202000212
131. Mozar A, Haren N, Chasseraud M, et al. High extracellular inorganic phosphate concentration inhibits RANK-RANKL signaling in osteoclast-like cells. J Cell Physiol. 2008;215(1):47–54. doi:10.1002/jcp.21283
132. Xu Y, Xu C, He L, et al. Stratified-structural hydrogel incorporated with magnesium-ion-modified black phosphorus nanosheets for promoting neuro-vascularized bone regeneration. Bioact Mater. 2022;16:271–284. doi:10.1016/j.bioactmat.2022.02.024
133. Jing X, Xu C, Su W, et al. Photosensitive and conductive hydrogel induced innerved bone regeneration for infected bone defect repair. Adv Healthc Mater. 2023;12(3):e2201349. doi:10.1002/adhm.202201349
134. Su Y, Zeng L, Deng R, et al. Endogenous electric field-coupled PD@BP biomimetic periosteum promotes bone regeneration through sensory nerve via Fanconi anemia signaling pathway. Adv Healthc Mater. 2023;12(12):e2203027. doi:10.1002/adhm.202203027
135. Palsson-McDermott EM, Curtis AM, Goel G, et al. Pyruvate kinase M2 regulates Hif-1α activity and IL-1β induction and is a critical determinant of the Warburg effect in LPS-activated macrophages. Cell Metab. 2015;21(2):347. doi:10.1016/j.cmet.2015.01.017
136. He L, Zhao J, Li H, et al. Metabolic reprogramming of NK cells by black phosphorus quantum dots potentiates cancer immunotherapy. Adv Sci. 2023;10(8):e2202519. doi:10.1002/advs.202202519
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms.php
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms.php
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.