")
Back to Journals » Diabetes, Metabolic Syndrome and Obesity » Volume 17
Liraglutide Ameliorates Renal Endothelial Dysfunction in Diabetic Rats Through the Inhibition of the Dll4/Notch2 Pathway
Authors Li Y, Chen Y, Zhang H , Chen W, Pan Y
Received 20 August 2024
Accepted for publication 23 October 2024
Published 30 October 2024 Volume 2024:17 Pages 4091—4104
DOI https://doi.org/10.2147/DMSO.S492252
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Prof. Dr. Juei-Tang Cheng
Yining Li, Yulin Chen, Hui Zhang, Weidong Chen, Yan Pan
Department of Nephrology, The First Affiliated Hospital of Bengbu Medical University, Bengbu, People’s Republic of China
Correspondence: Weidong Chen; Yan Pan, Department of Nephrology, The First Affiliated Hospital of Bengbu Medical University, 287 Changhuai Road, Bengbu, People’s Republic of China, Email [email protected]; [email protected]
Purpose: The glucagon-like peptide-1 receptor agonist (GLP-1RA) is a pharmacological agent utilized for the treatment of diabetes, known for its significant reno protective effects. This study aims to investigate the impact of liraglutide, a representative GLP-1RA medication, on early endothelial dysfunction in diabetic rats and elucidate its underlying mechanisms.
Methods: The present study employed a high-fat, high-sugar diet in combination with a single intraperitoneal injection of streptozotocin (STZ) to establish an experimental rat model of diabetes. Subsequently, the therapeutic efficacy of liraglutide on renal injury in this model was evaluated using various doses.
Results: Compared to the DKD rats, the rats treated with Liraglutide exhibited significant reductions in levels of blood glucose (Glu), serum creatinine (Scr), and blood urea nitrogen (BUN) (P < 0.05). Furthermore, there was a dose-dependent decrease in urinary protein levels, including 24-hour urinary protein excretion rate and microalbuminuria (m-ALB), with higher doses demonstrating more pronounced therapeutic effects (P < 0.05). In addition, treatment with Liraglutide effectively improved glomerular and interstitial damage, and suppressed the expression of CD31, CD34, and VE-cadherin associated with endothelial cell injury (P < 0.05). Furthermore, Liraglutide administration significantly increased nitric oxide (NO) production (P < 0.05). Moreover, Liraglutide treatment resulted in decreased expression of vascular endothelial growth factor (VEGF), Delta-like ligand-4(Dll4), and Notch2 protein in the Notch2 signaling pathway (P < 0.05).
Conclusion: The findings indicate that Liraglutide has a substantial effect on decreasing urinary protein excretion and improving vascular microinflammation, thus alleviating endothelial dysfunction in diabetic nephropathy. This observed mechanism can be attributed to the inhibition of the Dll4/Notch2 signaling pathway.
Keywords: diabetic kidney disease, Dll4/Notch2 signaling pathway, endothelial dysfunction, liraglutide, VEGF
Introduction
The global prevalence and mortality rate of diabetes are steadily increasing, imposing a significant burden on public health and the economy.1 It is projected that diabetes will emerge as the seventh leading cause of mortality globally by 2030.2 Diabetic kidney disease (DKD) represents the most prevalent microvascular complication associated with diabetes, characterized by endothelial dysfunction in glomerulus, thickening of GBM, mesangial matrix proliferation, and nodular glomerulosclerosis.3 In the early stages of diabetic nephropathy, endothelial dysfunction in the capillaries plays a pivotal role in initiating and advancing glomerular damage.GLP-1RAs, such as liraglutide, exhibit independent renal protective effects.4,5 The present study demonstrates that liraglutide can ameliorate renal endothelial dysfunction in Sprague-Dawley rats fed a high-salt diet by upregulating the expression of endothelial nitric oxide synthase (eNOS) and activating the adenosine monophosphate-activated protein kinase (AMPK)-eNOS pathway, thereby uncoupling the signaling axis between vascular endothelial growth factor (VEGF) and nitric oxide (NO).6,7 However, further investigations are warranted to elucidate the precise mechanisms underlying liraglutide’s beneficial effects on endothelial dysfunction in DKD.
The Notch signaling pathway is a highly conserved signal transduction pathway in multicellular organisms, which governs cell differentiation, proliferation, and apoptosis by mediating receptor-ligand interactions.8,9 The recent research underscores the pivotal role of the Notch signaling pathway in the progression of vascular diseases.10–13 In a murine model of diabetes, the overexpression of endothelial cell-specific Notch ligand Jag1 impeded the activation of the Notch signaling pathway and impeded vascular remodeling.14 Additionally, modulation of glucose metabolism guides NOTCH signaling to regulate mesenchymal progenitor cells (MPC), resulting in cell cycle arrest.15,16 The ligand DLL4 (delta-like ligand 4) of Notch2 exhibits predominant expression in endothelial cells and facilitates the cascade response between vascular endothelial growth factor receptor A and vascular endothelial growth factor receptor 2 (VEGFA-VEGFR2) signaling.17–19 with elevated levels of VEGF upregulating DLL4 expression.20 Gene expression analyses have demonstrated significant expression of Notch2 in peripheral blood samples from patients with DKD, exhibiting a significant positive correlation with microalbuminuria.21 Additionally, the analysis of protein expression in renal tissues exhibiting evident proteinuria provides supplementary evidence that supports the upregulation of the Notch2 signaling pathway.22,23 Importantly, renal glomerular Notch2 expression is significantly elevated in patients with DKD and rodent experimental models.24,25 These findings suggest that inhibition of Notch2 expression may serve as a potential therapeutic target for ameliorating glomerular endothelial dysfunction in DKD. The objective of this study is to investigate the regulatory effect of Liraglutide on endothelial dysfunction in glomeruli and elucidate its potential mechanisms by establishing a streptozotocin-induced diabetic rat model.
Materials and Methods
Animals and Treatments
Fifty male Sprague-Dawley rats (aged 6–8 weeks and weighing 180–220 g) were procured from Qinglongshan Biotechnology (Jiangsu, China), meeting SPF-grade standards. These animals possessed a valid certificate with the number SCXK (Su) 2024–0001. They were housed in a well-ventilated animal facility maintained at a constant temperature (22°C ± 2°C) and humidity (50%± 5%), following a light-dark cycle of 12 hours. The rats had an libitum access to food and water, while bedding was regularly replaced.
After one week of adaptive feeding, the rats were randomly allocated into five groups (n=8). The groups established through random sampling included a normal control group (NC group), a diabetic kidney disease model group (DKD group), a low-dose liraglutide group (DKD-LL group), a medium-dose liraglutide group (DKD-ML group), and a high-dose liraglutide group (DKD-HL group). The NC group was administered a standard diet, while the remaining mice were provided with a high-fat and high-sugar diet. After 4 weeks, except for the NC group, all rats underwent STZ-induced modeling. The experimental protocol involved a 12-hour fasting period, during which participants were allowed to drink water without restriction, followed by an intraperitoneal injection of 45mg/kg 1% STZ (Solarbio Company, Beijing). Blood glucose levels were assessed by means of tail vein blood sampling within a 72-hour post-injection period. Successful modeling criteria included consecutive fasting blood glucose (FBG)≥16.7mmol/L for more than three days, urine volume exceeding original volume by >150%, and urinary protein excretion >30 mg/24 h.26,27 After successful modeling, DKD-HL, DKD-ML, and DKD-LL groups received subcutaneous injections of Liraglutide (Novo, Nordisk A/S China) at doses of 200ug/kg/d, 100ug/kg/d and 50ug/kg/d respectively. The NC and DKD groups received equivalent amounts of saline solution via subcutaneous injection. During the administration period, observations were made on the mental state, fur condition, diet, and urine output of the rats. The sample collection was conducted after 8 weeks of intervention, as it aligns with the typical stability period of early pathological changes observed in diabetic nephropathy.28,29
Biochemical Analysis
Scr, BUN, glucose (Glu), albumin (Alb), and hemoglobin A1c(HbA1c) were quantified using a fully automated biochemical analyzer. Additionally, prior to euthanasia, rats were individually housed in metabolic cages for 24 hours to collect urine samples. After centrifugation at 1000 rpm for 5 minutes at 4°C, an appropriate volume of supernatant was collected for urinary microalbumin analysis using a specific reagent kit from Jiancheng Bioengineering Research Institute (Nanjing, China). Total protein content in the 24-hour urine sample was determined using Nanjing Jiancheng’s assay. Additionally, the transparent liquid was obtained by thoroughly lysing renal tissue, and the levels of nitric oxide were quantified using a NO detection kit (Beyotime Biotechnology, Shanghai).
Renal Pathology Observation
Renal tissue was excised and divided into three portions: one portion was immersed in pre-chilled electron microscopy fixative, provided by Servicebio Biotechnology (Wuhan, China) for the observation of ultrastructural changes in renaltissue; another portion was fixed with 4% paraformaldehyde (Servicebio) for histopathological examination; the remaining portion was stored at −80°C for Western blot analysis.
Light Microscope
Renal tissue specimens fixed in formalin were retrieved from the polyformaldehyde fixative solution, dehydrated, and cleansed. Subsequently, the tissues were embedded in paraffin and sectioned into approximately 3.5μm thick slices. Morphological changes of glomeruli and interstitial injury were observed under an Olympus microscope using Hematoxylin-eosin (HE), periodic acid-Schiff (PAS), and Masson’s trichrome staining techniques. For HE staining, sections were dewaxed, hydrated, stained with hematoxylin for 15 minutes followed by rinsing; then stained with eosin for 5 minutes, rinsed again, dehydrated, and mounted following standard procedures. For PAS staining, sections were dewaxed, hydrated; immersed in a high iodine acid solution for 10 minutes followed by triple rinsing; subsequently incubated at room temperature in a dark place with Schiff reagent for 40 minutes before being rinsed again. Staining with hematoxylin was performed for 1 minute followed by another rinse prior to dehydration and mounting. For Masson’s trichrome staining technique, sections were first dewaxed and hydrated; then stained with fuchsin under acidic conditions followed by immersion in phosphotungstic solution and subsequent washing with ammonium iron alum solution before dehydration and mounting.
Transmission Electron Microscope
The renal tissue was fixed in the electron microscopy fixative for 2 hours, rinsed overnight, and subsequently fixed with 10 g/L osmium tetroxide (pH: 7.3–7.4) for 1.5 hours. Following a 20-minute wash with buffer solution, dehydration was performed using a gradient of acetone before soaking and embedding prior to ultra-thin sectioning. The sections were stained with uranyl acetate and lead citrate, washed, dehydrated, mounted on slides, and observed under a transmission electron microscope to analyze the morphological changes of glomerular endothelial cells and thickness of GBM.
Immunohistochemistry
The renal tissue frozen sections were incubated overnight at 4°C with primary antibodies, including CD31, CD34, and VE-cadherin (Biosharp Biotechnology, Hefei). Subsequently, the sections were incubated for 50 minutes with horseradish peroxidase-conjugated secondary antibodies (Biosharp), followed by contrast staining using the nuclear dye DAB (Zsbio, Beijing). The sliced images were observed and captured under a microscope at a magnification of 400x and analyzed using Image J software.
Western Blot
Renaltissue was lysed using RIPA and PMSF (Biosharp), and protein concentration was determined by the BCA method (GIPBIO, USA). Uniform loading of 60μg protein was achieved on 7.5% and 10% SDS-PAGE gels, which were subsequently transferred onto a 0.45 μm PVDF membrane. The membrane was blocked with Biosharp’s rapid blocking solution for 30 minutes followed by overnight incubation at 4°C with anti-VEGF antibody (Zen-Bioscience, Chengdu) at a dilution of 1:1000, anti-DLL4 antibody (Protech, Wuhan) at a dilution of 1:500, Notch2 antibody (Zen-Bioscience) at a dilution of 1:500 and β-actin antibody (Zen-Bioscience) at a dilution of 1:10000. Subsequently, the membrane was incubated with a goat anti-rabbit secondary antibody conjugated with horseradish peroxidase (Biosharp) and visualized after one hour. Semi-quantitative analysis was conducted using Image J software.
Statistical Analysis
The experimental data were statistically analyzed and graphed using GraphPad Prism 10 software. Quantitative data are reported as the mean ± standard error of the mean (SEM). After confirming normality in data distribution and conducting homogeneity tests, paired sample t-tests are employed to compare two sets of measurement data. One-way analysis of variance (ANOVA) is utilized for multiple sets of measurement data. In cases where there is heterogeneity in variances, rank sum tests are applied. A significance level of P<0.05 was considered statistically significant.
Results
The Body Weight Change Before and After Treatment
Compared to the NC group, the DKD group exhibited significant increases in urine volume, water intake, and food consumption. Additionally, they displayed symptoms such as fur yellowing and lethargy, which were alleviated in the treatment groups. We assessed the body weight of rats prior to and following treatment (Figure 1A) and observed a significant increase in the NC group after 8 weeks (327.06±12.46g). Conversely, the DKD group (238.15±16 0.84 g) as well as all treatment groups exhibited a reduction in body weight, with the DKD group demonstrating the most pronounced decrease (P<0.05). This finding suggests that liraglutide significantly improves low body weight status among rats with DKD.As depicted in Figure 1D and Table 1, the renal index (double renal weight(g)/bodyweight ratio(kg)) further corroborated these outcomes.
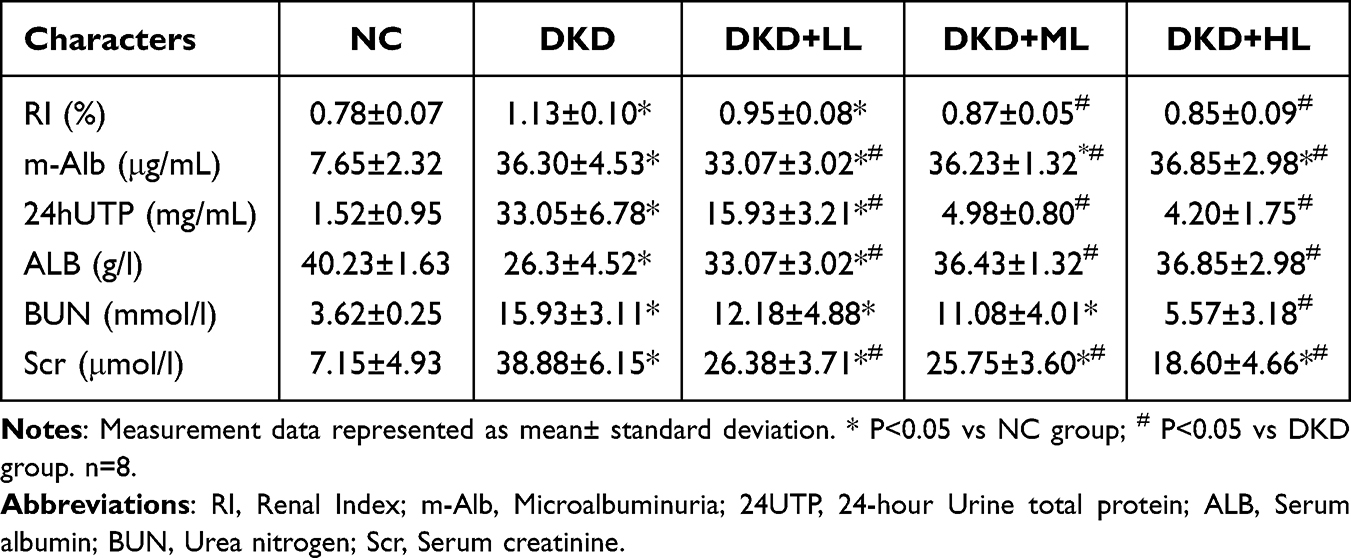 |
Table 1 Changes in Renal Function After Treatment |
 |
Figure 1 The impact of Liraglutide treatment on various parameters in each group of rats. Abbreviations: Glu, Glucose; HbA1c, Glycated hemoglobin; RI, Renal Index; m-Alb, Microalbuminuria; 24h UTP, 24-hour Urine total protein; Alb, Serum albumin; Scr, Serum creatinine; BUN, Urea nitrogen. Notes: (A) Body weight of rats in each group before and after treatment; (B) Glu; (C) HbA1c; (D) RI; (E) m-Alb; (F) 24h UTP; (G) Alb; (H) Scr; (I) BUN. Measurement data represented as mean± standard deviation. *P<0.05, ** P<0.01, *** P<0.001 vs NC group; # P<0.05, ## P<0.01, ### P<0.001 vs DKD group. n=8. |
Effect of Liraglutide on Blood Glucose
In comparison to the NC group with blood glucose levels of 5.475 ± 2.13 mmol/L and HbA1c levels of 4.06 ± 0 0.71%, the DKD group showed significantly increased blood glucose levels (Figure 1B; 26.84 ± 10.80 mmol /L) and HbA1c levels (Figure 1C; 17.15 ± 2.15%). Nevertheless, these elevations were effectively reversed after liraglutide treatment administration. The noteworthy finding was that the DKD-HL group displayed a more substantial decrease in HbA1c levels (6 0.33 ± 0 0.63%), indicating that higher dosage treatments may lead to superior long-term glycemic control effects.
Effect of Liraglutide on Renal Function
We measured a series of indicators reflecting changes in renal function (Table 1). Compared to the NC group, the DKD group exhibited significant increases in urinary microalbumin (U-Alb), 24UTP, Scr, and BUN levels (Figure 1E, F, H and I). Treatment with liraglutide mitigated these elevations, and the extent of reduction was positively correlated with the dosage administered. These findings suggest that higher doses of liraglutide within a certain range yield superior therapeutic effect. Moreover, there was a notable improvement in Alb levels (Figure 1G), which was consistent with the observed reduction in urinary protein excretion, thereby indicating its effective enhancement of Alb levels in rats with DKD.
Histopathological Morphological Changes of Renal
Significant pathological changes were observed in the rat renal of the DKD group compared to the NC group during observation under an optical microscope. HE staining (Figure 2A) revealed an increase in mesangial cells; PAS staining (Figure 2C) demonstrated significant enlargement of glomeruli (Table 2), mild thickening of the basement membrane, and proliferation of mesangial matrix; Masson staining (Figure 2E) exhibited hypertrophy of glomerular capillary loops and expansion of the mesangial area. TEM observation reveals significant pathological alterations in the glomeruli of the DKD group (Figure 3B), characterized by uniform thickening of the basement membrane and extensive fusion or even loss of foot processes. However, treatment with liraglutide resulted in varying degrees of improvement in these pathological changes, with the most notable improvement observed in the DKD-HL group. Furthermore, measurement of GBM thickness revealed a significant increase from baseline values (0.14±0.01μm) for DKD group (0.48±0.06μm). Compared to the DKD group, all treatment groups demonstrated a prominent trend of decreased basement membrane thickness. Importantly, this reduction was particularly pronounced in the high-dose liraglutide treatment group (0.22±0.02μm). These results indicate that liraglutide therapy contributes to restoring normal GBM structure and improving early-stage renal damage caused byDKD.
 |
Table 2 Changes in Glomerular Diameter and Surface Area of PAS |
 |
Figure 2 Light microscopic pathology of renal tissue in each group of rats. Notes: (A) HE staining of the renal tissues; (B) PAS staining of the renal tissue; (C) Masson staining of the renal tissues. Original magnification: ×400, bar=50μm. |
 |
Figure 3 Ultrastructure of renal tissue in each group of rats. Notes: (A–E) The changes of renal tissue detected via the microscope. (A) NC group; (B) DKD group; (C) DKD+LL group; (D) DKD+MLgroup; (E) DKD+HL group. (F) Comparison of the thickness of GBM in each group (n=6). Measurement data represented as mean± standard deviation. **P<0.01, ***P<0.001 vs NC group; #P<0.05, ##P<0.01 vs DKD group. Original magnification: ×1000, bar=5.0μm. |
Expression of Proteins Associated with Glomerular Endothelial Injury
Immunohistochemistry results demonstrated that CD31 staining (Figure 4D) was predominantly localized in the glomerular capillaries. Compared to the NC group, DKD exhibited enhanced expression of endothelial cell staining intensity within glomerular capillaries. Quantitative analysis revealed a significant increase in the number of CD31-positive cells (Figure 4A), which showed a dose-dependent decrease after liraglutide treatment. The CD34 staining (Figure 4E) showed predominant localization to glomerular walls and mesangial areas, with additional expression detected on proximal tubule walls. Quantitative analysis demonstrated a significant increase in CD34-positive cell numbers in the DKD group (Figure 4B), which subsequently decreased following liraglutide treatment. VE-cadherin staining (Figure 4F) primarily displayed expression within endothelial cells and glomerular mesangial area. Compared to the NC group, there was a noticeable rise in positive cell counts in the DKD group (Figure 4C); however, improvement was observed after administration of liraglutide.
 |
Figure 4 Immunohistochemical evaluation of the expression level of CD31/CD34/VE-cadherin in each group of rats. Notes: (A–C) Expression analysis of CD31/CD34/VE-cadherin in IHC for each group. (D) CD31 staining; (E) CD34 staining; (F) VE-cadherin staining (n=6). Measurement data represented as mean± standard deviation. ***P<0.001 vs NC group; #P<0.05, ##P<0.01, vs DKD group. Original magnification: ×400, bar=50μm. |
Changes in the Protein Levels of VEGF/Dll4/Notch2
We assessed the levels of proteins involved in the VEGF/Dll4/Notch2 pathway associated with endothelial dysfunction. Compared to the NC group (Figure 5), the DKD group exhibited elevated expression of VEGF, Dll4, and Notch2, which subsequently decreased across all three groups following liraglutide intervention. Notably, both VEGF and Dll4 expression showed a significantly greater reduction in the DKD-ML and DKD-HL groups However, there was no statistically significant difference observed in the expression of Notch2 among the three intervention groups. To further investigate the potential therapeutic effect of liraglutide on endothelial function in diabetic kidney disease (DKD), we quantified renal tissue levels of nitric oxide (NO). Our findings revealed a significant reduction in NO content within the DKD group compared to the non-diabetic control (NC) group (P<0.01). However, treatment with liraglutide resulted in an augmentation of NO production, suggesting that this pharmacological agent may possess the capacity to protect and restore endothelial function. This phenomenon could potentially be attributed to liraglutide’s ability to enhance NO synthesis or release through yet unidentified mechanisms, thereby ameliorating impaired endothelial cell function in DKD.
 |
Figure 5 The effect of Liraglutide on renal glomerular endothelial function in each group of rats. Notes: (A) The Western blotting; (B) the analysis of expression level of Notch2; (C) the analysis of expression level of Dll4; (D) the analysis of expression level of VEGF. (E) Nitric oxide (NO) content within the renal endothelium (n=6). Measurement data represented as mean± standard deviation. *P<0.05, **P<0.01, ***P<0.001 vs NC group; #P<0.05, ##P<0.01, ###P<0.001 vs DKD group. |
Discussion
GLP-1RA, an antidiabetic medication, has been extensively utilized in clinical diabetes treatment. The representative drug liraglutide not only effectively reduces blood sugar levels but also ameliorates endothelial dysfunction in various diseases.30–33 It can prevent the decrease of eNOS phosphorylation induced by free fatty acids and maintain endothelial cell function.34 Furthermore, it exhibits a regulatory effect on endothelial dysfunction in a mouse model of angiotensin II–induced hypertension.35 However, the underlying mechanisms responsible for endothelial dysfunction in DKD remain unclear. In this study, we observed that liraglutide significantly improved Glu levels, reduced urinary protein excretion, increased Alb levels, and enhanced renal function in diabetic rats. These effects were positively correlated with the therapeutic dosage. Further histopathological examination of renal tissues revealed that liraglutide alleviated STZ-induced renal pathology damage in DKD rats through downregulation of proteins associated with endothelial injury. Moreover, for the first time in this study, elevated expression of VEGF, Dll4, and Notch2 proteins was observed in DKD renal tissues; however, liraglutide treatment was able to reduce their expression. The research findings demonstrate that liraglutide can ameliorate glomerular endothelial dysfunction and confer renoprotective effects in DKD through the inhibition of the Dll4/Notch2 signaling pathway.
In this study, we initially focused on renal function indicators and administered three different doses of liraglutide to treat DKD rats in order to assess its therapeutic effects on DKD. The results demonstrated that liraglutide significantly ameliorated the overall condition of DKD rats, regulated serum glucose metabolism, and reduced urinary protein and Scr levels, with the highest dose exhibiting the most favorable treatment outcome. The findings from this study indicated that liraglutide enhanced renal function indicators, with its efficacy being positively correlated with dosage. These observations are consistent with previous research findings.36–39
In addition to functional improvements, we investigated the pathological protective effects and potential mechanisms of liraglutide in DKD. Renal histopathological analysis of DKD rats included HE, PAS, Masson staining, and transmission electron microscopy observations, which revealed pathological changes such as glomerular hypertrophy, thickening of the basement membrane, and mesangial matrix proliferation in DKD renal tissues. These symptoms were significantly ameliorated following treatment with liraglutide. Furthermore, we observed a significant upregulation in the protein expression levels of CD31, CD34, and VE-cadherin, key markers associated with endothelial injury in diabetic nephropathy. Endothelial cells at the first barrier of the glomerular filtration membrane are susceptible to damage from high glucose levels abnormal lipids and inflammatory factors,40,41 potentially leading to functional and structural changes within the glomerulus through cross-interference mechanisms.42 Existing research suggests that compared to podocyte injury, endothelial cell injury is more closely associated with increased urinary albumin excretion,43,44 indicating that endothelial cell damage may have a greater impact on glomerular changes in DKD than podocyte injury. The administration of Liraglutide led to a significant decrease in the expression levels of CD31, CD34, and VE-cadherin proteins, providing further evidence that Liraglutide may ameliorate endothelial dysfunction through specific molecular pathways, thereby exerting protective effects on renal function. To further elucidate this mechanism of action, we conducted additional studies.
Recent research has revealed that the Notch1 signal modulates VE-cadherin levels by regulating transcription factors SNAI1 and ERG, resulting in glomerular filtration barrier dysfunction and proteinuria induction.45 In both developing and adult mammalian kidneys, Notch1 and Notch2 exhibit high structural similarity with almost overlapping expression patterns.32 Based on current findings and existing literature,46 we hypothesize that Liraglutide may exert its renal protective effects by modulating specific signaling pathways. Using Western blot technique, we assessed the expression of proteins involved in the VEGF/DLL4/Notch2 pathway. Our results demonstrated increased levels of VEGF, DLL4, and Notch2 proteins in the DKD group compared to the NC group, while different doses of Liraglutide intervention groups showed varying degrees of decreased expression. Therefore, it is speculated that regulation of the VEGF/DLL4/Notch2 signaling pathway by Liraglutide may represent one of its key mechanisms for improving DKD.47,48 Furthermore, no significant differences were observed in Notch2 expression within the VEGF/DLL4/Notch2 pathway when comparing protein expression among the three intervention groups. Some studies have suggested a lack of correlation between gamma-secretase inhibitor dosage and inhibition of notch pathway protein expression in glioblastoma stem cells;49 others have found that the impact of Notch2 on DKD depends on Notch1 expression.46 This phenomenon may be attributed to a lack of cascading amplification in signal transduction between notch receptors and ligands; however, further investigation is warranted due to limitations inherent in existing research.
In this study, we observed therapeutic effects of different doses of liraglutide on DKD. The DKD-HL group exhibited the most significant effect, followed by the DKD-ML group. However, we also noted some inconsistent results. Specifically, Glu levels were higher in the DKD-HL group compared to the DKD-ML group. Interestingly, CD34 expression in renal tissues was higher in the DKD-ML group than in the DKD-LL group. These discrepancies may be attributed to variations in baseline characteristics among randomly assigned groups, which could have influenced an accurate assessment of treatment efficacy. It is important to note that our research primarily focuses on animal models and does not directly elucidate the mechanism of action of liraglutide on renal glomerular endothelial cells under in vitro conditions. Furthermore, additional comprehensive clinical trials are necessary to validate the sustained efficacy of liraglutide in improving endothelial function among patients with DKD, as well as to determine the optimal dosage and safety profile. Additionally, it is worth exploring whether a synergistic effect exists between liraglutide and other treatment modalities, such as ACE inhibitors or ARBs, when used in combination. These investigations will facilitate the optimized utilization of this drug in clinical practice and provide personalized precision treatment strategies for individuals DKD.
Conclusion
In conclusion, we emphasize liraglutide as a promising therapeutic option for DKD and have demonstrated its significant efficacy in renal protection through innovative mechanisms and essential preliminary data. Further elucidation of the potential mechanisms of action of liraglutide or conducting clinical trials to investigate the impact of different doses and treatment durations would greatly enhance the drug’s value in DKD clinical practice.
Data Sharing Statement
The data that support the findings of this study are available from the corresponding author upon reasonable request.
Ethics Approval and Consent to Participate
The experimental animals and their breeding conditions are in accordance with the Guide for Laboratory Animal Care and Use as well as the Animal Welfare Act, while strictly adhering to the guidelines established by the Ethics Review Committee of Bengbu Medical College (Approval No. 2024-611).
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
The funding for this study was provided by the Key Project of Provincial Natural Science Research in Anhui Province Higher Education Institutions (kJ2013A191), the Key Project of Natural Science Foundation of Bengbu Medical College (2022byzd041), and the Key Project of Provincial Natural Science Research in Anhui Province Higher Education Institutions (2023AH051936). The authors declare no conflicts of interest associated with this publication.
Disclosure
The authors declare that they have no competing interests in this work.
References
1. Khan MAB, Hashim MJ, King JK. et al. Epidemiology of type 2 diabetes–global burden of disease and forecasted trends. J Epidemiol Glob Health. 2020;10(1):107–111. doi:10.2991/jegh.k.191028.001
2. Cho NH, Shaw JE, Karuranga S, et al. IDF Diabetes Atlas: global estimates of diabetes prevalence for 2017 and projections for 2045. Diabet Res Clin Pract. 2018;138:271–281. doi:10.1016/j.diabres.2018.02.023
3. Chen X, Tan H, Xu J, et al. Klotho-derived peptide 6 ameliorates diabetic kidney disease by targeting Wnt/β-catenin signaling. Kidney Int. 2022;102(3):506–520. doi:10.1016/j.kint.2022.04.028
4. Su K, Yi B, Yao BQ, et al. Liraglutide attenuates renal tubular ectopic lipid deposition in rats with diabetic nephropathy by inhibiting lipid synthesis and promoting lipolysis. Pharmacol Res. 2020;156:104778. doi:10.16/j.phrs.2020.104778
5. Xiao S, Yang Y, Liu YT, Zhu J. Liraglutide regulates the kidney and liver in diabetic nephropathy rats through the miR-34a/SIRT1 pathway. J Diabetes Res. 2021;2021:8873956. doi:10.1155/2021/8873956
6. Sukumaran V, Tsuchimochi H, Sonobe T, et al. Liraglutide improves renal endothelial function in Obese Zucker rats on a high-salt diet. J Pharmacol Exp Ther. 2019;369(3):375–388. doi:10.1124/jpet.118.254821
7. Li K, Sun J, Huang N, et al. Liraglutide improves obesity-induced renal injury by alleviating uncoupling of the glomerular VEGF-NO axis in obese mice. Clin Exp Pharmacol Physiol. 2020;47(12):1978–1984. doi:10.1111/1440-1681.13391
8. Ma T, Li X, Zhu Y, et al. Excessive activation of notch signaling in macrophages promote kidney inflammation, fibrosis, and necroptosis. Front Immunol. 2022;13:835879. doi:10.3389/fimmu.2022.835879
9. Mack JJ, Iruela-Arispe ML. NOTCH regulation of the endothelial cell phenotype. Curr Opin Hematol. 2018;25(3):212–218. doi:10.1097/MOH.0000000000000425
10. Xiong Z, Hu Y, Jiang M, et al. Hypoxic bone marrow mesenchymal stem cell exosomes promote angiogenesis and enhance endometrial injury repair through the miR-424-5p-mediated DLL4/Notch signaling pathway. PeerJ. 2024;12:e16953. doi:10.7717/peerj.16953
11. Zheng Y, Wang S, Xue X, et al. Notch signaling in regulating angiogenesis in a 3D biomimetic environment. Lab Chip. 2017;17(11):1948–1959. doi:10.1039/C7LC00186J
12. Wang Y, Singh AR, Zhao Y, et al. TRIM28 regulates sprouting angiogenesis through VEGFR-DLL4-Notch signaling circuit. FASEB J. 2020;34(11):14710–14724. doi:10.1096/fj.202000186RRR
13. Tanajak P, Pintana H, Siri-Angkul N, et al. Vildagliptin and caloric restriction for cardioprotection in pre-diabetic rats. J Endocrinol. 2017;232(2):189–204. doi:10.1530/JOE-16-0406
14. Yoon CH, Choi YE, Cha YR, et al. Diabetes-induced Jagged1 overexpression in endothelial cells causes retinal capillary regression in a murine model of diabetes mellitus: insights into diabetic retinopathy. Circulation. 2016;134(3):233–247.
15. Hilton MJ, Tu X, Wu X, et al. Notch signaling maintains bone marrow mesenchymal progenitors by suppressing osteoblast differentiation. Nat Med. 2008;14(3):306–314. doi:10.1038/nm1716
16. Hankenson KD, Gagne K, Shaughnessy M. Extracellular signaling molecules to promote fracture healing and bone regeneration. Adv Drug Deliv Rev. 2015;94:3–12. doi:10.1016/j.addr.2015.09.008
17. Luna-Zurita L, Prados B, Grego-Bessa J, et al. Integration of a Notch-dependent mesenchymal gene program and Bmp2-driven cell invasiveness regulates murine cardiac valve formation. J Clin Invest. 2010;120(10):3493–3507. doi:10.1172/JCI42666
18. Pitulescu ME, Schmidt I, Giaimo BD, et al. Dll4 and Notch signalling couples sprouting angiogenesis and artery formation. Nat Cell Biol. 2017;19(8):915–927. doi:10.1038/ncb3555
19. Trindade A, Kumar SR, Scehnet JS, et al. Overexpression of delta-like 4 induces arterialization and attenuates vessel formation in developing mouse embryos. Blood. 2008;112(5):1720–1729. doi:10.1182/blood-2007-09-112748
20. Li L, Liu Q, Shang T, et al. Aberrant activation of Notch1 signaling in glomerular endothelium induces albuminuria. Circ Res. 2021;128(5):602–618. doi:10.1161/CIRCRESAHA.120.316970
21. Sharaf SA, Kantoush NA, Ayoub DF, et al. Altered expression of WFS1 and NOTCH2 genes associated with diabetic nephropathy in T2DM patients. Diabet Res Clin Pract. 2018;140:304–313. doi:10.1016/j.diabres.2018.03.053
22. Sharma M, Magenheimer LK, Home T, et al. Inhibition of Notch pathway attenuates the progression of human immunodeficiency virus-associated nephropathy. Am J Physiol Renal Physiol. 2013;304(8):F1127–F1136. doi:10.1152/ajprenal.00475.2012
23. Murea M, Park JK, Sharma S, et al. Expression of Notch pathway proteins correlates with albuminuria, glomerulosclerosis, and renal function. Kidney Int. 2010;78(5):514–522. doi:10.1038/ki.2010.172
24. Niranjan T, Bielesz B, Gruenwald A, et al. The Notch pathway in podocytes plays a role in the development of glomerular disease. Nat Med. 2008;14(3):290–298. doi:10.1038/nm1731
25. Niranjan T, Murea M, Susztak K. The pathogenic role of Notch activation in podocytes. Nephron Exp Nephrol. 2009;111(4):e73–e79. doi:10.1159/000209207
26. Liu Y, Wang S, Jin G, et al. Network pharmacology-based study on the mechanism of ShenKang injection in diabetic kidney disease through Keap1/Nrf2/Ho-1 signaling pathway. Phytomedicine. 2023;118:154915. doi:10.1016/j.phymed.2023.154915
27. Chen F, Ge L, Jiang X, et al. Construction of the experimental rat model of gestational diabetes. PLoS One. 2022;17(9):e0273703. doi:10.1371/journal.pone.0273703
28. Mann JFE, Ørsted DD, Buse JB. Liraglutide and renal outcomes in type 2 diabetes. N Engl J Med. 2017;377(22):2197–2198. doi:10.1056/NEJMoa1616011
29. Ren H, Shao Y, Wu C, et al. Metformin alleviates oxidative stress and enhances autophagy in diabetic kidney disease via AMPK/SIRT1-FoxO1 pathway. Mol Cell Endocrinol. 2020;500:110628. doi:10.1016/j.mce.2019.110628
30. Li N, Zhao Y, Yue Y, Chen L, Yao Z, Niu W. Liraglutide ameliorates palmitate-induced endothelial dysfunction through activating AMPK and reversing leptin resistance. Biochem Biophys Res Commun. 2016;478(1):46–52. doi:10.1016/j.bbrc.2016.07.095
31. Ying W, Meiyan S, Wen C, et al. Liraglutide ameliorates oxidized LDL-induced endothelial dysfunction by GLP-1R-dependent downregulation of LOX-1-mediated oxidative stress and inflammation. Redox Rep. 2023;28(1):2218684. doi:10.1080/13510002.2023.2218684
32. McLean BA, Wong CK, Kabir MG, et al. Glucagon-like Peptide-1 receptor Tie2+ cells are essential for the cardioprotective actions of liraglutide in mice with experimental myocardial infarction. Mol Metab. 2022;66:101641. doi:10.1016/j.molmet.2022.101641
33. Li PC, Liu LF, Jou MJ, et al. The GLP-1 receptor agonists exendin-4 and liraglutide alleviate oxidative stress and cognitive and micturition deficits induced by middle cerebral artery occlusion in diabetic mice. BMC Neurosci. 2016;17(1):37. doi:10.1186/s12868-016-0272-9
34. Zhao Q, Xu H, Zhang L, et al. GLP-1 receptor agonist lixisenatide protects against high free fatty acids-induced oxidative stress and inflammatory response. Artif Cells Nanomed Biotechnol. 2021;49(1):544.
35. Chen SJ, Lv LL, Liu BC, et al. Crosstalk between tubular epithelial cells and glomerular endothelial cells in diabetic kidney disease. Cell Prolif. 2020;53(3):e12763. doi:10.1111/cpr.12763
36. Yang S, Lin C, Zhuo X, et al. Glucagon-like peptide-1 alleviates diabetic kidney disease through activation of autophagy by regulating AMP-activated protein kinase-mammalian target of rapamycin pathway. Am J Physiol Endocrinol Metab. 2020;319(6):E1019–E1030. doi:10.1152/ajpendo.00195.2019
37. Chen P, Shi X, Xu X, et al. Liraglutide ameliorates early renal injury by the activation of renal FoxO1 in a type 2 diabetic kidney disease rat model. Diabet Res Clin Pract. 2018;137:173–182. doi:10.1016/j.diabres.2017.09.006
38. Yamada S, Tanabe J, Ogura Y, et al. Renoprotective effect of GLP-1 receptor agonist, liraglutide, in early-phase diabetic kidney disease in spontaneously diabetic Torii fatty rats. Clin Exp Nephrol. 2021;25(4):365–375. doi:10.1007/s10157-020-02007-2
39. Liao TT, Zhao LB, Liu H, et al. Liraglutide protects from renal damage via Akt-mTOR pathway in rats with diabetic kidney disease. Eur Rev Med Pharmacol Sci. 2019;23(3 Suppl):117–125.
40. Jourde-Chiche N, Fakhouri F, Dou L, et al. Endothelium structure and function in kidney health and disease. Nat Rev Nephrol. 2019;15(2):87–108. doi:10.1038/s41581-018-0098-z
41. Fu J, Akat KM, Sun Z, et al. Single-cell RNA profiling of glomerular cells shows dynamic changes in experimental diabetic kidney disease. J Am Soc Nephrol. 2019;30(4):533–545. doi:10.1681/ASN.2018090896
42. Mahtal N, Lenoir O, Tharaux PL. Glomerular endothelial cell crosstalk with podocytes in diabetic kidney disease. Front Med. 2021;8:659013. doi:10.3389/fmed.2021.659013
43. Xiang E, Han B, Zhang Q, et al. Human umbilical cord-derived mesenchymal stem cells prevent the progression of early diabetic nephropathy through inhibiting inflammation and fibrosis. Stem Cell Res Ther. 2020;11(1):336. doi:10.1186/s13287-020-01852-y
44. Qi H, Casalena G, Shi S, et al. Glomerular endothelial mitochondrial dysfunction is essential and characteristic of diabetic kidney disease susceptibility. Diabetes. 2017;66(3):763–778. doi:10.2337/db16-0695
45. Papoutsi T, Luna-Zurita L, Prados B, Zaffran S, de la Pompa JL. Bmp2 and Notch cooperate to pattern the embryonic endocardium. Development. 2018;145(13):dev163378. doi:10.1242/dev.163378
46. Sweetwyne MT, Gruenwald A, Niranjan T, Nishinakamura R, Strobl LJ, Susztak K. Notch1 and Notch2 in podocytes play differential roles during diabetic nephropathy development. Diabetes. 2015;64(12):4099–4111. doi:10.2337/db15-0260
47. Elkhoely A. Liraglutide ameliorates gentamicin-induced acute kidney injury in rats via PGC-1α- mediated mitochondrial biogenesis: involvement of PKA/CREB and Notch/Hes-1 signaling pathways. Int Immunopharmacol. 2023;114:109578. doi:10.1016/j.intimp.2022.109578
48. Zhang LY, Wang Y, Yang YR, et al. MiR-135a regulates renal fibrosis in rats with diabetic kidney disease through the Notch pathway. Eur Rev Med Pharmacol Sci. 2020;24(4):1979–1987. doi:10.26355/eurrev_202002_20375
49. Yi L, Zhou X, Li T, et al. Notch1 signaling pathway promotes invasion, self-renewal and growth of glioma initiating cells via modulating chemokine system CXCL12/CXCR4. J Exp Clin Cancer Res. 2019;38(1):339. doi:10.1186/s13046-019-1319-4
 © 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms.php
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms.php
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.