")
Back to Journals » Infection and Drug Resistance » Volume 18
Mechanisms of Antimicrobial Resistance in Klebsiella: Advances in Detection Methods and Clinical Implications
Authors Li J , Shi Y, Song X, Yin X, Liu H
Received 27 November 2024
Accepted for publication 20 February 2025
Published 11 March 2025 Volume 2025:18 Pages 1339—1354
DOI https://doi.org/10.2147/IDR.S509016
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Sandip Patil
Jian Li, Yuwen Shi, Xuanli Song, Xiaoyu Yin, Hui Liu
Department of Bacteria Disease, Jinan Center for Disease Control and Prevention, Jinan, Shandong, People’s Republic of China
Correspondence: Hui Liu, Department of Bacteria Disease, Jinan Center for Disease Control and Prevention, Jinan, Shandong, People’s Republic of China, Tel +86-531-81278920, Email [email protected]
Abstract: Klebsiella spp. are ubiquitous gram-negative bacteria, commonly present in natural environments and as part of the human microbiota. Klebsiella is involved in the occurrence and development of many diseases, and effective antibiotics for it have attracted the attention of researchers. In recent years, its multi-drug resistance, particularly to carbapenems and β-lactam antibiotics, has posed significant challenges to clinical treatment. Therefore, a comprehensive understanding of the resistance mechanisms of Klebsiella, along with the enhancement of detection methods, is crucial for effectively controlling the spread of drug-resistant strains and guiding individualized clinical treatment. This paper systematically reviews the epidemiological characteristics, resistance mechanisms, detection methods, and treatment strategies of Klebsiella, aiming to provide new insights for the clinical management of this pathogen.
Keywords: Klebsiella, drug resistance, detection method, β-lactam, carbapenems
Introduction
Klebsiella pneumoniae (KP) is a gram-negative, non-motile, capsulated opportunistic bacterium belonging to the Enterobacteriaceae family.1 It is widely distributed in the environment, including soil, water, and plant surfaces, and is also commonly found on the skin, in the respiratory tract, and within the intestines of humans and animals.2 Klebsiella was originally identified by German pathologist Carl Friedländer, who first described it in 1882. During his research on lung tissue samples from patients with pneumonia, he discovered a Gram-negative, short rod-shaped bacterium. Due to its strong association with certain cases of pneumonia, this bacterium is also referred to as “Friedlander pneumococcus”. In 1896, the microbiology community named the genus Klebsiella in honor of Edwin Klebs, recognizing his significant contributions to medical microbiology. Klebs was a pioneer in the field of pathology and was among the first to apply bacteriology to the study of disease, and his research provided a theoretical foundation for the integration of bacterial taxonomy and pathology.3,4 Subsequently, as medical research advanced, K. pneumoniae was recognized for its high pathogenicity and diverse infection routes, leading to a range of serious infections, including community-acquired pneumonia, urinary tract infections, gastroenteritis, septicemia, and liver abscesses, particularly in immunocompromised individuals,5–8 this expanded understanding of its pathogenic scope suggested that K. pneumoniae is a pathogen with significant clinical importance.
In recent years, the widespread use of antibiotics has exacerbated the issue of Klebsiella resistance, and its antibiotic resistance has evolved from Multidrug Resistance (MDR) to Extensively Drug Resistance (XDR) and Pan-Drug Resistance (PDR).9 Notably, hypervirulent Klebsiella pneumoniae (hvKP) not only possesses heightened virulence factors but also demonstrates increased drug resistance, complicating clinical treatment.10 According to data from the Chinese Bacterial Resistance Surveillance (CHINET), the resistance rate of Klebsiella to carbapenem antibiotics, such as imipenem and meropenem, significantly increased between 2015 and 2019, although a decrease was observed in 2021.11 These fluctuations indicate that resistance surveillance and management measures for Klebsiella require optimization and enhancement. Countries have developed their own policies for managing antimicrobial resistance in accordance with the WHO action plan. China’s policy, known as the National Action Plan on Antimicrobial Resistance (2016–2020), includes several key initiatives: strengthening the surveillance of antimicrobial resistance, implementing a classification management system for antibiotics to standardize clinical use; restricting the use of growth-promoting antibiotics in aquaculture; and raising public awareness to reduce the self-purchase and use of antibiotics. In terms of medical institution management, the plan involves the formulation of an antibacterial drug prescription list and the implementation of special rectification actions to ensure the rational clinical use of antibiotics. Additionally, the management of resistance in the environment focuses on strengthening the monitoring and control of antibiotic pollution. Given that Klebsiella drug resistance poses a major threat to global public health, international health organizations and national centers for disease control have focused considerable attention on this issue. This paper reviews the classification, resistance mechanisms, virulence factors, and current treatment strategies for Klebsiella, aiming to provide valuable references for its prevention, control, and clinical application. By strengthening drug property monitoring, optimizing antibiotic usage, and developing new therapeutic methods, it is anticipated that the clinical challenges posed by Klebsiella can be more effectively addressed. This paper systematically reviews the epidemiological characteristics, resistance mechanisms, detection methods, and treatment strategies of Klebsiella, aiming to provide new insights for the clinical management of this pathogen.
Classification and Epidemiology of Klebsiella
Klebsiella pneumoniae (KP) can be classified into two pathogenic categories: classic Klebsiella pneumoniae (cKP) and hypervirulent Klebsiella pneumoniae (hvKP).12 cKP is a prevalent pathogen in hospital-acquired infections, ranking second in infection rates only to E. coli, with a clinical detection rate of 6.69%. This pathogen is commonly found in the mouth, skin, digestive tract mucosa, and on the surfaces of medical equipment. It poses a significant risk to individuals with compromised immune systems, such as those with diabetes or malignant tumors, leading to severe pneumonia, urinary tract infections, respiratory infections, and other complications.13,14 cKP exhibits high levels of drug resistance, particularly with respect to carbapenem antibiotics, where resistance rates exceed 10%.9,15 Since the 1990s, hvKP has garnered increased attention as a notable cause of community-acquired infections, with a case fatality rate ranging from 3% to 32%.16 Infections caused by hvKP are more frequently observed in otherwise healthy individuals and often result in invasive conditions, such as liver abscesses, endophthalmitis, and brain abscesses. Its high viscosity contributes to its propensity for causing such invasive infections. The identification criteria for hvKP include the wire drawing test,17 capsule serotyping (eg, K1, K2),18,19 and detection of virulence genes (eg rmpA/rmpA2). An hvKP infection is confirmed if two or more of these criteria are met.20,21 hvKP infections tend to develop rapidly, exhibit stronger transmission, and result in more severe disease, thereby becoming a focal point of clinical concern.22 Routine surveillance data revealed increasing rates of invasive extended-spectrum cephalosporin-resistant Klebsiella pneumoniae (ESCR-KP) in Switzerland, from 1.3% in 2004 to 8.5% in 2019.23 A global exploration of 13,178 KP strains revealed that 7.8% of them were CR-hvKP (carbapenem-resistant hypervirulent Klebsiella pneumoniae).24 In China, a significant study indicated that among the screened carbapenem-resistant Klebsiella pneumoniae (CR-KP), as much as 36% carried hypervirulence factors.25 This shows a significant risk to public health since these strains integrate resistance to potent antibiotics and increased pathogenicity.
Mechanism of Drug Resistance in Klebsiella
In recent years, the drug - resistance problem of Klebsiella has become increasingly severe. This is manifested in an increasing number of types of drugs that can be tolerated, and different species of Klebsiella exhibit species-specific resistance, for example, in the case of KP, genes like blaKPC and blaNDM confer resistance to β-lactams including carbapenems; mcr-1 leads to colistin resistance; In Klebsiella oxytoca, blaCTX-M causes resistance to multiple β-lactams, and aac(3)-II results in aminoglycoside resistance; Klebsiella granulomatis may have tet genes, making it resistant to tetracycline.1,26 An in - depth understanding of the drug - resistance mechanisms of Klebsiella is conducive to controlling its drug resistance.
The Production of Beta-Lactamase
β-lactam antibiotics encompass a range of compounds, including penicillin and its derivatives, cephalosporins, monoamide rings, carbapenems, and penem enzyme inhibitors. These antibiotics function by binding to penicillin-binding proteins (PBPs), which disrupt the synthesis of bacterial cell walls.27 However, Gram-negative bacteria can develop resistance by hydrolyzing these antibiotics through the production of ultra-broad spectrum beta-lactamases (ESBLs).28,29 ESBLs are primarily transmitted via plasmids and are capable of hydrolyzing cephalosporins, monoamides, and penicillin antibiotics. They can be inhibited by clavulanate, which is classified as group 2be in functional classification and class A in molecular classification.30 ZHANG et al found that 12.6% of the 230 strains of hvKP isolated were ESBL positive, with the blaCTX-M gene being the predominant genetic determinant. ESBL-positive strains carrying the blaCTX-M gene exhibit increased susceptibility in patients with a history of steroid therapy, neutropenia, or combination therapy.31 SHANKAR et al and Qiu et al reported that the hvKP strains producing ESBL were predominantly of the serotype K54, with sequence types ST29 and ST2318.32,33 Additionally, another study indicated that 56% of cKP strains and 17% of hvKP strains carried ESBLs.34
β-lactamase is a significant mechanism of bacterial drug resistance. Conventional β-lactamases can be categorized into chromosome-mediated and plasmid-mediated enzymes, and further classified based on their hydrolytic activity into penicillinases, cephalosporinases, broad-spectrum enzymes, and ultra-broad-spectrum enzymes.35 Ultra-broad-spectrum β-lactamases (ESBLs) represent a critical resistance mechanism in Klebsiella; these plasmid-mediated enzymes, composed of serine protein derivatives, are capable of hydrolyzing penicillins, broad-spectrum and ultra-broad-spectrum cephalosporins, as well as monocyclic β-lactam antibiotics. The genotypes of ESBLs include TEM, SHV, OXA, CTX-M, PER, VEB, GES, IBC, among others. Notably, TEM and SHV are the most prevalent genotypes, with significant regional variations observed in different countries. For instance, SHV and CTX-M types are predominant in Italy, while SHV types are more common in Canada.36 In China, CTX-M is the primary genotype, with CTX-M-3 and SHV-12 being dominant in Shanghai, whereas CTX-M types are predominant in Hangzhou. TEM-type ESBLs primarily derive from TEM-1, SHV-type ESBLs are predominantly found in K. pneumoniae, and CTX-M ESBLs exhibit resistance to ceftazidime while having a weaker hydrolytic ability against ceftaxime. The main drugs used to treat ESBL-producing bacteria are carbapenems and cephamycin antibiotics, with imipenem being the preferred choice due to its broad spectrum and high antibacterial activity.37 In recent years, cases of ESBL-producing hypervirulent Klebsiella pneumoniae (hvKP) infections have been reported globally, indicating a growing concern regarding resistance to ESBLs.
Plasmid Mediated AmpC Enzyme
Plasmid-mediated resistance is a primary mechanism through which bacteria acquire antibiotic resistance. This resistance propagates among bacterial populations via plasmids—circular DNA molecules that carry resistance genes, thereby facilitating the rapid dissemination and long-term survival of resistant strains. Plasmids can spread among bacteria through various mechanisms, including conjugation, transformation, transduction, integrons, and transposons. Several factors contribute to the ability of plasmid-mediated resistance to promote the swift spread and persistence of drug-resistant strains: efficient transmission, strong adaptability, genetic stability, multi-resistance, and environmental transmission. The plasmids associated with K. pneumoniae drug resistance mainly include IncFII, IncL, IncX3, IncC and ColKP3. In addition, insertion sequences such as IS26 and transposon like Tn1546 also play important roles in the dissemination of K. pneumoniae drug resistance.38–40
Cephalosporinase (AmpC) is a significant β-lactamase produced by gram-negative bacilli, which can be mediated by both chromosomes and plasmids.41 The plasmid-mediated expression of the AmpC enzyme in bacteria can result in resistance to first- to third-generation cephalosporins, cephalomycin, aztreonam, and other antibiotics; moreover, β-lactamase inhibitors are not effective against this enzyme.42 AmpC enzymes are classified as Group 1 in the functional classification and Class C in the molecular classification. They can hydrolyze third-generation cephalosporins, cephalomycin (with the exception of ACC-1), and monocyclic β-lactam antibiotics, yet they remain sensitive to fourth-generation cephalosporins and carbapenems.
Resistance in K. pneumoniae is frequently linked to multiple germplasm and mobile genetic elements capable of carrying and transmitting antibiotic resistance genes. AmpC enzymes are commonly found in bacteria that do not possess the inducible chromosomal genes encoding these enzymes, including Escherichia coli, K. pneumoniae, Klebsiella acidogenes, and Enterobacter aerogenes. The expression of the AmpC gene is regulated by various genes. AmpD belongs to the ampC gene cluster located on chromosomes. This cluster is often associated with induced drug resistance. AmpR, a promoter region about 100bp upstream of the ampC gene, controls the expression of the AmpC gene. AmpG, which is located in other regions of the chromosome, is responsible for transporting peptidoglycan fragments.43 Research has demonstrated that certain strains, such as DHA-1- producing Klebsiella pneumoniae, exhibit reduced sensitivity to carbapenem drugs following the loss of outer membrane proteins.44 Since the initial discovery of plasmid-mediated AmpC enzymes, such as MIR-1, in 1988, over 40 different genotypes have been reported. Studies indicate that plasmid-mediated enzymatic hydrolysis by AmpC enzymes has a broader spectrum, contributing to increased bacterial resistance to cefepime and other antibiotics.45 Furthermore, plasmids encoding AmpC enzymes often coexist with other antibiotic resistance genes, including those conferring resistance to aminoglycosides, chloramphenicol, and sulfonamides, reflecting a pattern of multidrug resistance. Some bacteria are capable of simultaneously producing extended-spectrum β-lactamases (ESBLs) and plasmid-mediated AmpC enzymes, a phenomenon referred to as “super-broad spectrum β-lactamase”.46 This type of enzyme exhibits a wide range of degradation substrates and enhanced hydrolysis efficiency, which has significant epidemiological implications.
Carbapenemases
Carbapenems are atypical β-lactam antibiotics characterized by their broad antibacterial spectrum and potent antibacterial activity. Due to their stability against β-lactamases and low toxicity, carbapenems have emerged as one of the primary antibacterial agents for treating severe bacterial infections.19 The production of Klebsiella pneumoniae carbapenemase (KPC) represents the principal mechanism by which K. pneumoniae develops resistance to carbapenem antibiotics.47 These enzymes not only confer resistance to a wide array of β-lactam antibiotics but also exhibit resistance to β-lactam inhibitors.
Carbapenemase refers to a class of β-lactamases capable of hydrolyzing imipenem or meropenem. According to the Ambler classification (Table 1), it is primarily divided into Class A, Class B (which includes metallo-β-lactamases, or MBLs), and Class D enzymes. The most significant enzyme in Class A is KPC, which was first identified in North Carolina in 1996 and named KPC.48 KPC-positive Klebsiella pneumoniae (KP) was first detected in China in 2004 in a 75-year-old ICU patient in Zhejiang Province. KPC-2 is the most prevalent carbapenemase type in China.49 Class B metalloenzymes include NDM-1, IMP, VIM, among others, while Class D is primarily represented by OXA-48 enzymes. Studies have shown that resistance of carbapenemase - producing KP to carbapenem antibiotics has increased over the years, which is primarily because carbapenemases are generally plasmid - encoded enzymes, and the IncL/M plasmid is related to plasmids that carry the KPC-type carbapenemase gene (such as bla_KPC), which confers resistance to carbapenem antibiotics, and the types of plasmids involved in the propagation of the bla_KPC gene typically include the widely distributed IncF, IncI2, IncN, IncA/C, and pKpQIL-like plasmids, especially important in spreading bla_KPC in K. pneumoniae.38,39 According to the 2017 CHINET Bacterial Resistance Surveillance data, the resistance rates of KP to imipenem and meropenem rose from 3.0% and 2.9% in 2005 to 20.9% and 24.0% in 2017, respectively. Carbapenemase-producing strains exhibit resistance not only to carbapenem antibiotics, but also to penicillins, cephalosporins, and monobactams.
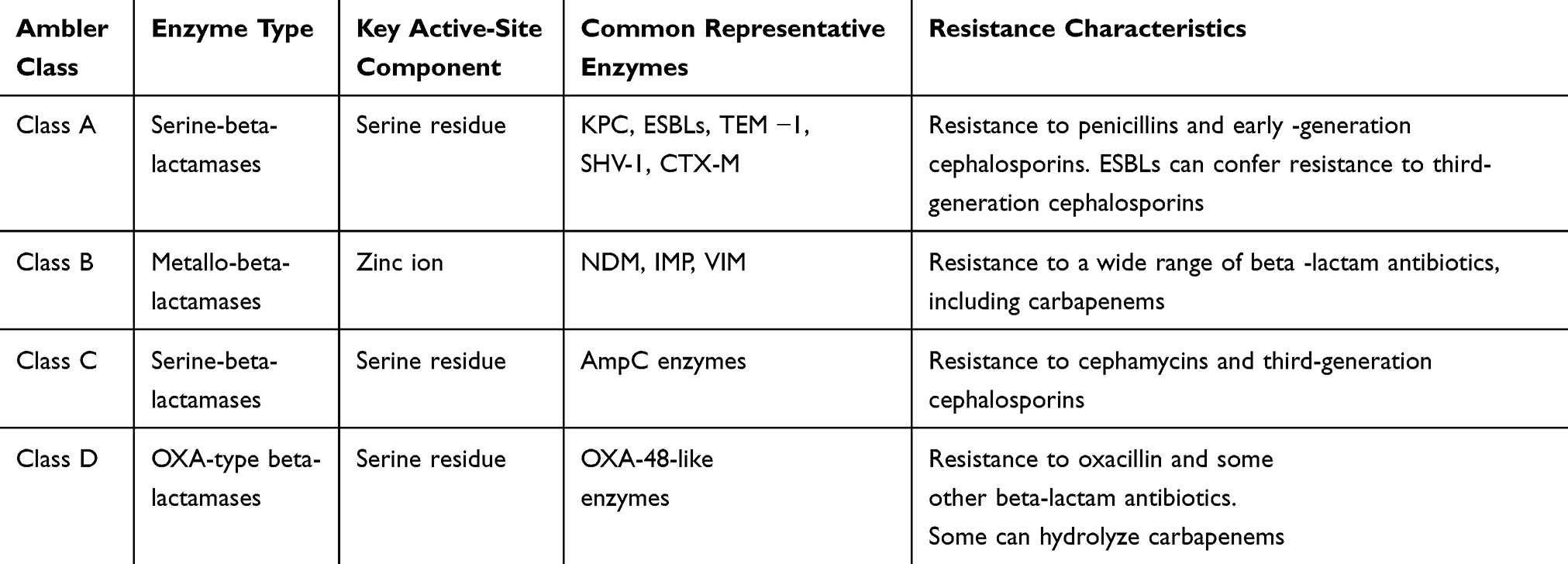 |
Table 1 Ambler Classification of Beta-Lactamases |
Beta-Lactamase Resistant to Enzyme Inhibitors
Enzyme-resistant beta-lactamases (IRBLs) are classified as Class A in molecular classification and group 2br in functional classification. These enzymes primarily arise from 1–3 mutations in the TEM-1, TEM-2, and SHV-1 genes at specific amino acid sites. Other variants include SHV-10, SHV-49, OXY-2, and OXA-type β-lactamases. The amino acid replacement sites of IRBLs are commonly found at positions 69, 130, 244, 275, and 276 of TEM-1 and TEM-2. Mutations at these sites can significantly decrease the affinity for β-lactam antibiotics and alter the interaction with enzyme inhibitors, particularly at sites 69 and 244, which are located at or near the active site of TEM enzymes. Such mutations can confer resistance to enzyme inhibitors.50,51 IRBLs can readily induce resistance in Klebsiella pneumoniae to amoxicillin, ticarcillin, and various enzyme inhibitors (such as clavulanate and sulbactam); however, they remain sensitive to narrow-spectrum cephalosporins, 7-alpha-methylcephalosporins, and oxyiminocephalosporins.
Gene Mutation and Horizontal Gene Transfer
The Role and Change of Membrane Porins
The extracellular lipid layers of Gram-negative bacteria, such as KP, contain numerous pores formed by various microporins, including OmpK35, OmpK36, and OmpK37. These pores serve as water-soluble diffusion channels that are non-specific and facilitate the crossing of the cell membrane.52 Some beta-lactamases can penetrate the bacterial interior through these channels, thereby exerting their effects. However, alterations or the absence of microporous proteins can impede the penetration of antibacterial drugs into the bacterial cell, leading to drug resistance. OmpK35 is identified as the primary membrane porin associated with bacterial drug resistance, while OmpK37 is typically expressed at low levels or not at all. Changes in porin structure enhance bacterial resistance, and this resistance is more pronounced when inactivated enzymes coexist with altered porins, compared to the effects of inactivated enzymes alone. Webster et al demonstrated that the absence of OmpK35 and OmpK36 can result in meropenem resistance in KPN.53 Conversely, Yang et al indicated that while the loss of membrane porins elevates the minimum inhibitory concentration (MIC) of carbapenem antibiotics, it does not necessarily confer imipenem resistance.54
Carbapenems represent the “last line of defense” among beta-lactam antibiotics and are utilized to treat Gram-negative bacterial infections that exhibit resistance to other antibiotic classes. However, inappropriate or excessive use of these agents can accelerate the emergence and dissemination of resistance. Many Gram-negative bacteria, such as Pseudomonas aeruginosa and Acinetobacter baumannii, inherently possess low levels of beta-lactamase genes, including AmpC. Under the selective pressure of carbapenems, these genes may undergo mutations that enhance their efficacy, leading to the production of ultra-broad spectrum β-lactamases or carbapenemases. Carbapenemase production is the primary mechanism by which bacteria develop resistance to carbapenem antibiotics. The genes encoding these enzymes, including bla_NDM-1, bla_KPC, and bla_OXA-48, are typically found on plasmids, facilitating their rapid spread through horizontal gene transfer.
Additionally, bacteria may diminish the permeability of carbapenems by down-regulating the expression or mutating porins. This mechanism often co-occurs with carbapenemase production, thereby further augmenting drug resistance. Mutations in porin genes may manifest as point mutations, small fragment deletions, whole gene deletions, or insertions. The most prevalent mutation involves insertions that disrupt porin synthesis, leading to reduced or absent porin levels.52,55 The combination of porin deletion and β-lactamase activity can contribute to carbapenem resistance. Research has shown that K. pneumoniae strains with a deletion of OmpK36 expression, coupled with the production of SHV-2 extended-spectrum beta-lactamases (ESBLs), exhibit resistance to ketomycin, β-lactams, enzyme inhibitors, and carbapenems.55
Efflux Pump and Gene Mutation
The efflux pump is a significant mechanism of resistance in KP.56,57 This organism possesses an energy-dependent protein efflux system located in its inner membrane, which actively expels drugs that infiltrate the bacteria. When specific regulatory mechanisms within the bacteria are altered, leading to enhanced expression of the active efflux system, the concentration of drugs within the bacteria may become insufficient to exert an antibacterial effect, thereby resulting in drug resistance.
The OqxAB efflux pump in KP is a key part of the multidrug - resistance mechanism. It reduces the intracellular concentration of β - lactam antibiotics, boosting the efficiency of β - lactam enzymes in degrading these drugs. When bacteria have both the OqxAB pump and β - lactam resistance genes, their resistance to β - lactam antibiotics rises significantly. This is often seen when the OqxAB pump works with down - regulated or mutated membrane porin expression, thereby reducing antibiotic penetration. OqxAB efflux pumps often co - exist on the same plasmid as other resistance genes like bla_CTX - M, bla_KPC, and aac(6’) - Ib - cr. This not only lowers antibiotic levels by efflux but also heightens resistance through other means such as enzymatic degradation.56 Furthermore, the OqxAB efflux pump can decrease quinolone concentrations by expelling them, thus acting synergistically with mutations in DNA cyclotase. Such synergistic mechanisms are particularly prevalent in drug-resistant strains of Salmonella and K. pneumoniae.
The expression of the OqxAB efflux pump is often upregulated during biofilm formation, which further enhances bacterial tolerance to antibiotics. The interplay between the biofilm’s barrier function and the efflux pump complicates the treatment of bacterial infections.58 Additionally, OqxAB efflux pumps may share substrates or energy sources with other toxic secretion systems, thereby bolstering bacterial viability. For instance, efflux pumps can diminish bacterial sensitivity to environmental toxins, such as bile salts, and enhance their capacity for colonization within the host. Due to its non-specific nature, this system encompasses a broad range of efflux substrates, including beta-lactams, quinolones, macrolides, and other antibacterial agents. Consequently, multiple active efflux systems often coexist within the same bacterial strain, contributing to resistance against various antibacterial drugs with distinct structures.
Research has identified AcrAB-TolC as the primary efflux system in bacteria, which is prevalent in KP and is a major contributor to its multidrug resistance.59 Furthermore, studies indicate that mutations in the negative regulatory ramR gene, along with overexpression of the positive regulatory ramA gene—both of which are associated with the expression of the AcrAB efflux pump—can lead to increased expression of the AcrAB efflux pump, thereby enhancing the resistance of beta-lactam producing bacteria to beta-lactam antibiotics.60,61 Additionally, the OqxAB efflux pump gene was initially believed to reside on the K. pneumoniae chromosome; however, it has since been recognized as a plasmid-mediated quinolone resistance gene capable of transferring between different bacterial species.
Topoisomerase and Gene Variation
In KP, the structural conformation of the parC gene, which encodes topoisomerase IV, and the gyrA gene, which encodes DNA gyrase, is susceptible to alteration when the targets of these two enzymes are mutated.62 This alteration leads to the destabilization of the enzyme-DNA complex with antibiotics, ultimately resulting in quinolone resistance in KPN. Variation in the gyrA gene plays a predominant role in target gene variation, with common loci of variation including Ser83 and Asp87 of gyrA, as well as Ser80 and Glu84 of parC. Additionally, the plasmid-mediated drug resistance gene qnr specifically interacts with topoisomerase IV by encoding a protein that diminishes the efficacy of quinolones, thereby contributing to bacterial drug resistance.
Mechanism of Polymyxin Resistance
In polymyxin antibiotics, the primary mechanism of KP resistance is attributed to chromosomal mutations that facilitate target modification via the lipopolysaccharide modification system.63 In strains possessing this system, alterations in the lipopolysaccharide structure result in a reduction of anionic charge, effectively impeding the binding of polymyxins. The typical modifications observed in lipopolysaccharides primarily stem from mutations in core genes, which lead to lipid A maturation, lipid A neutralization, and reactions involving phosphoethanolamine or palmitolipid binding.64 Furthermore, the enhanced activity of lipopolysaccharide-modifying gene regulators, along with other mutations, can directly cause an abnormal increase in pmrB expression or a loss of activity in mgrB, both of which may contribute to polymyxin resistance in these strains. See Table 2 for details.
 |
Table 2 Klebsiella Specific Drug Resistance Phenotypes/Genotypes |
Klebsiella Resistance Detection Method
Klebsiella resistance detection is an important link in clinical microbiology. It is often used to assess the susceptibility of the bacteria to different antibiotics, thereby helping to guide clinical treatment. The following are several commonly used detection methods (Table 3):
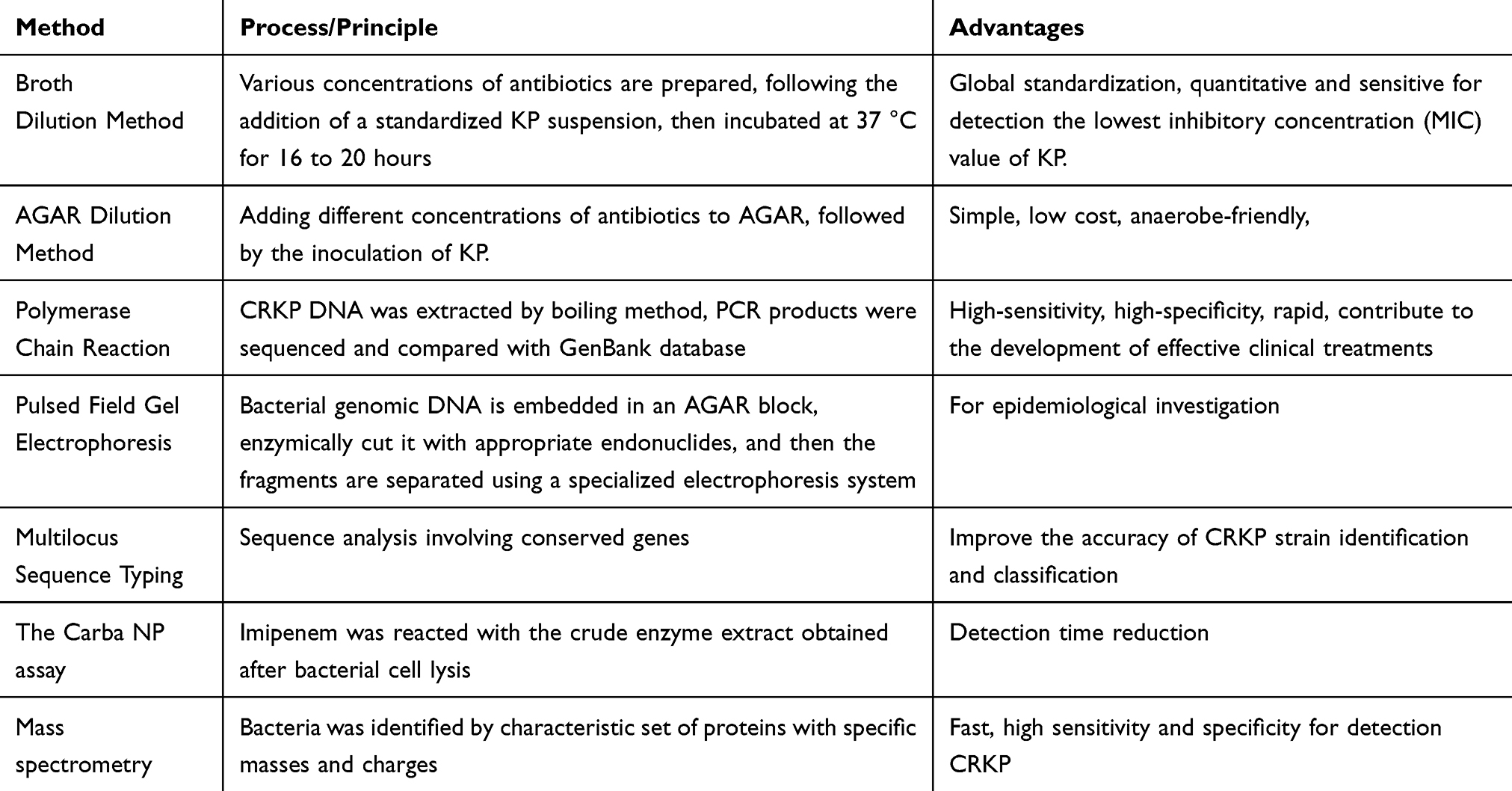 |
Table 3 Klebsiella Resistance Detection Method |
Traditional Drug Susceptibility Test
The traditional testing methods for KP primarily include the disk diffusion method, agar dilution method, broth dilution method, and concentration gradient method, among other classical approaches. These methods typically require a detection time of 16 to 24 hours and are commonly employed for the phenotypic screening of clinical drug-resistant strains. Their advantages include convenience, sensitivity, reproducibility, and low cost. However, these classical methods are limited to the phenotypic detection of bacterial resistance and do not allow for the identification of drug-resistance genes. The disk diffusion method, also known as the Kirby-Bauer (K-B) antibiotic test, involves placing a paper disc containing a specific concentration of antimicrobial agents on an MHA agar plate inoculated with KP.65 The antimicrobial agents diffuse into the surrounding medium, creating one dimension, while the bacteria on the surface of the agar plate grow, produce resistant enzymes, and spread into the medium, forming a second dimension; hence, it is referred to as a two-dimensional test.66 The plates are incubated at 37°C for 16 to 18 hours. Following incubation, the diameter of the antibacterial zone surrounding the paper disc is measured, allowing for the determination of bacterial sensitivity to the drug based on the size of this zone.
The broth dilution method is a traditional technique for detecting bacterial resistance. In this method, various concentrations of antibiotics are added to broth containing KP, which is then incubated at 37 °C for 16 to 20 hours following the addition of a standardized bacterial suspension. The minimum inhibitory concentration (MIC) is determined by observing bacterial growth. The microbroth dilution method is currently recognized as the international reference method for drug susceptibility testing, as established by the Clinical and Laboratory Standards Institute (CLSI). Its primary advantages include the global standardization of antimicrobial susceptibility tests, the straightforward process of inoculation and interpretation, and the ability to test multiple antimicrobial agents simultaneously against a single strain. However, a notable disadvantage is the significant variability in MIC values, which can result from technical discrepancies; thus, it requires operators to possess high experimental skills and a strong awareness of aseptic techniques. Conversely, the broth macro dilution method is an easily standardized and reliable reference method, offering substantial value and significance for interpreting the MIC values of drugs against specific bacterial strains in research and testing contexts.
The AGAR dilution method involves adding different concentrations of antibiotics to AGAR, followed by the inoculation of KP. After 48 hours of culture, colony-forming units (CFU) are counted.66,67 Additionally, KP can be inoculated onto chromogenic AGAR medium, which allows for faster detection and screening of relevant drug-resistant bacteria, as sensitive strains exhibit a color response within 18 to 24 hours. Such methods are well-suited for multi-bacterial cultures and can effectively distinguish between strains and species. The AGAR dilution method is advantageous due to its simplicity, low cost, and the stability and reliability of the minimum inhibitory concentration (MIC) values. Epsilometer testing (E-test) employs a series of pre-prepared antibacterial drugs with a continuous exponential increase in concentration dilution on one side, while the other side features a plastic strip marked with MIC values on the concentration scale.66 This strip is placed on an MHA AGAR plate inoculated with KP, and the MIC values on the strip are read after the culture period. The stable antibiotic concentration gradient on the E-test strip renders this method simple, sensitive, and accurate, allowing for precise reading of specific MIC values.
Application of Molecular Biology Techniques
Polymerase Chain Reaction (PCR) is a highly efficient and sensitive molecular biological technique used for amplifying specific DNA fragments.68 In this study, CRKP DNA was extracted using the boiling method, followed by the application of specific primers to amplify gene fragments associated with drug resistance, including TEM, SHV, CTX-M, PER, VEB, DHA, MIR/ACT, KPC, IMP, VIM, SPM, GIM, and NDM-1. Subsequently, the PCR products underwent DNA sequencing and were compared against the GenBank database to identify specific resistance genotypes. This process provides a precise molecular basis for identifying drug resistance in bacteria, which can aid in the development of effective clinical treatments. Furthermore, through sequence analysis and the judicious design of PCR primers, the distribution and evolution of drug-resistant genes can be further investigated, enhancing the ability to monitor and control multi-drug-resistant strains.
Pulsed Field Gel Electrophoresis (PFGE) is a high-resolution molecular typing technique commonly employed for the analysis of bacterial genomes, particularly for the separation of large DNA fragments ranging from 50 kb to 10 Mb.69 The fundamental principle involves embedding bacterial genomic DNA in an agar block, enzymatically cleaving it with appropriate endonucleases, and subsequently separating the fragments using a specialized electrophoresis system. In contrast to conventional gel electrophoresis, PFGE enhances the separation of large DNA fragments by alternating the direction of the electric field and varying the pulse time. According to the typing criteria proposed by Tenover et al,70 PFGE maps can facilitate the typing analysis of strains. The specific criteria are as follows: if the PFGE maps of different strains exhibit the same number and size of bands, they are classified as belonging to the same type; if the differences in the map result in a change of 1 to 3 bands, this is regarded as a change in subtype. Conversely, if 4 to 6 bands differ in the map, the strains are considered to belong to different types. If seven or more bands differ, these strains are typically regarded as uncorrelated. PFGE is frequently utilized in epidemiological investigations, particularly in tracking outbreaks of pathogens and studying the relationships between different strains.71 For instance, in the context of bacterial food poisoning or nosocomial infection outbreaks, PFGE can aid in tracing the source of the infection and determining whether different pathogens originate from the same strain or from distinct strains. In studies of carbapenem-resistant Klebsiella pneumoniae (CRKP), PFGE is extensively employed to monitor the transmission pathways of various strains and to elucidate the genetic variation among these strains. The integration of PFGE with Multi-Locus Sequence Typing (MLST) can provide a comprehensive scientific foundation for the prevention and control of the spread of drug-resistant strains.
Multilocus Sequence Typing (MLST) is a primary method in bacterial molecular epidemiology that involves the sequence analysis of conserved genes.72 In the molecular epidemiological study of carbapenem-resistant CRKP, seven steward genes of K. pneumoniae (gapA, infB, mdh, pgi, phoE, rpoB, and tonB) were amplified and sequentially analyzed. The sequence results of each gene’s fixed region were then compared with sequences in the database to identify the Sequence Type (ST). The predominant genotypes identified were ST11 and ST15, which exhibited high frequency. Drug resistance genes encoded by ST11 include KPC-2, TEM-1, DHA-1, and SHV-11, among others. These genes are significant contributors to carbapenem resistance in K. pneumoniae, complicating the treatment of such bacterial infections. In addition to ST11 and ST15, other sequence types, such as ST395 and ST273, also hold important clinical and epidemiological significance. Different transmission pathways and drug resistance mechanisms may exist among these sequence types, highlighting the importance of studying them. Moreover, combining the MLST method with other molecular biology techniques, such as whole genome sequencing (WGS) and pulsed-field gel electrophoresis (PFGE), can enhance the accuracy of identification and classification of CRKP strains. This multi-technology approach not only aids in a comprehensive understanding of the genetic background and evolutionary relationships of resistant strains but also provides critical information for developing new diagnostic tools and treatment options. Overall, the application of MLST in the molecular epidemiology of CRKP offers robust technical support and a scientific foundation for addressing the challenges posed by drug resistance.
Emerging Rapid Detection Methods
The Carba NP assay utilizes a crude extract of enzymes obtained after bacterial cell lysis to react with imipenem.73 Carbapenemases can hydrolyze carbapenem antibacterial agents, leading to ring-opening and the subsequent production of hydrogen ions (H+). A specific concentration of H+ can alter the color of the phenol red indicator in the detection solution, allowing for the determination of carbapenemase production based on the observed color change. The types of carbapenemase-producing strains are further identified using various enzyme inhibitors. This detection method is characterized by its simplicity, rapidity, sensitivity, and high specificity, particularly for the detection of KPC and metalloenzymes; however, it may yield false negative results for OXA-48 carbapenemase.74 Commercially available kits can significantly reduce the testing time from 2 hours to 15 minutes.75
Mass spectrometry (MALDI-TOF) employs laser desorption ionization technology to swiftly identify carbapenem-resistant gram-negative bacteria by detecting specific protein signals.76 The detection time ranges from 30 minutes to 2 hours, and this technology has been implemented in numerous hospitals to rapidly screen for pathogens associated with nosocomial infections, facilitating timely treatment interventions. The enzyme-linked immunosorbent assay (ELISA) detects carbapenemase within minutes through an antigen-monoclonal antibody reaction, achieving sensitivity and specificity rates exceeding 90%.55 This method enables the simultaneous detection of KPC, NDM, VIM, IMP, and OXA-48 carbapenemases. Although the ELISA method is straightforward and easy to interpret, it is associated with a high cost, which may limit its accessibility for early and rapid diagnosis and treatment of high-risk patients.
Comparison and Evaluation of Detection Methods
In gene detection and phenotype assessment, various methods possess distinct characteristics that render them suitable for different application scenarios.77 As a phenotypic detection approach, the traditional drug sensitivity test is limited to detecting a small number of antibacterial drugs, and it is both time-consuming and labor-intensive, particularly for certain bacteria that necessitate specialized growth environments. The MILST method demonstrates exceptional performance in genetic analysis, enabling the detection of multiple genotypes (eg ST11, ST15). Its advantages include good repeatability, high resolution, stable results, and ease of standardization; however, its high cost and limited applicability are notable drawbacks. FOUNOU et al utilized whole-genome sequencing (WGS) to analyze an ultra-broad-spectrum β-lactamase-producing K. pneumoniae strain isolated from a South African patient.77 The whole-genome sequencing analysis confirmed the observed resistance, revealing that the strain harbored multiple beta-lactamase genes along with various other resistance genes. All K. pneumoniae isolates contained multiple multidrug-resistant (MDR) efflux pump genes, including CmeA, CmeB, MATE, MFS, MacA, MarcB, MarA, OML, RND, AcrB, and AcrAB. These MDR efflux pumps confer resistance to several antibiotic families, including tetracyclines, fluoroquinolones, macrolides, tigecyclines, and beta-lactam antibiotics.
Due to its strong specificity and high sensitivity, the PCR method is widely employed to amplify drug-resistant genes and determine genotypes, such as TEM, SHV, and CTX-M. Despite its complexity and high cost, researchers favor this technique because of its robust specificity, high sensitivity, and low purity requirements for specimens. The PFGE method is primarily utilized for homology analysis and is regarded as the “gold standard” in bacterial molecular biotyping; however, improvements in its stability and standardization are still necessary. The RT-PCR method is used to analyze transcription products and drug resistance mechanisms of genes, demonstrating high sensitivity and broad applicability, although it has limitations when processing certain biological samples.78
Both the double-disk method and the three-dimensional test are employed to detect lactamase. The double-disk method is straightforward yet sensitive, whereas the three-dimensional test, while more reliable, is also more complex. The Carba NP test is rapid and easy to interpret, allowing for the detection of various carbapenemase enzymes; however, some strains yield ineffective results and are unable to detect the chromosomally encoded OXA carbapenemase.
Integration of Resistance Mechanisms and Detection Methods
Influence of Drug Resistance Mechanisms on Detection Strategies
The known resistance mechanisms of KP primarily include β-lactam enzymes, such as extended-spectrum β-lactamases (ESBLs) and carbapenemases, which hydrolyze and inactivate β-lactam antibiotics.79 Additionally, alterations in membrane porins decrease drug entry channels into the cells, thereby reducing the intracellular concentration of the drugs. Enhanced efflux pumps further diminish drug effectiveness by expelling the drugs from the cell. Furthermore, genetic mutations in genes such as gyrA and parC lead to variations in the antibiotic target sites, preventing effective binding of the antibiotics.
Traditional methods for detecting bacterial resistance can provide phenotypic information; however, they require extended periods for culture and identification and are unable to detect resistance genes. In contrast, new technologies utilizing physical, chemical, and molecular biological methods offer shorter detection cycles and higher sensitivity, yet they cannot identify unknown resistance genes or mutated genes. High-throughput sequencing technology enhances the phenotypic detection capabilities of classical methods by accurately identifying bacterial drug resistance genes and discovering novel drug resistance genes and mutation sites.
PCR and RT-PCR assays were employed to detect specific resistance genes, including extended-spectrum beta-lactamases (ESBLs) and carbapenemase genes such as KPC, NDM, and VIM. The high specificity and sensitivity of PCR make it the preferred method for identifying drug-resistant genes.80 Pulsed-field gel electrophoresis (PFGE) is utilized for homology analysis, which helps determine the homology and transmission routes of K. pneumoniae. This analysis is crucial for tracing the source of infections in hospitals and for developing effective prevention and control measures. The Carba NP test is used to identify the phenotype of carbapenemases. Additionally, two-disc and three-dimensional tests are employed to detect AmpC and ESBL enzymes. Although the three-dimensional test yields more reliable results, it is more complex to perform. Conversely, while the two-disc method is easier to operate, it exhibits lower sensitivity.
Optimize Clinical Treatment Strategies Based on Test Results
Understanding the resistance mechanisms of K. pneumoniae is crucial for selecting appropriate detection methods that can facilitate the rapid and accurate diagnosis of drug-resistant strains, thereby providing essential guidance for clinical treatment. Initially, therapeutic drugs should be selected based on the results of drug sensitivity tests to prevent the misuse of antimicrobials, particularly broad-spectrum agents, which can accelerate the development of bacterial resistance. Techniques such as PCR and RT-PCR enable the prompt identification of specific resistance genes in strains, allowing for the avoidance of ineffective medications. The choice of antibiotics should be informed by the identified resistance mechanisms (eg extended-spectrum beta-lactamases (ESBLs), carbapenemases). For instance, in the case of ESBL-producing KP, carbapenem antibiotics may be appropriate, while for carbapenemase-producing strains, drugs such as tigecycline or polymyxins should be considered.81,82 Furthermore, homology analysis using pulsed-field gel electrophoresis (PFGE) and other methodologies can be employed to trace the transmission pathways of drug-resistant bacteria, necessitating isolation measures to curb the spread of nosocomial infections. Regular monitoring of drug resistance patterns in pathogenic bacteria during treatment is essential, allowing for timely adjustments to treatment plans to prevent the further dissemination of drug-resistant strains.
The combination drug strategy, which represents the latest approach to treating patients with carbapenem-resistant Enterobacteriaceae (CRE) infections, advocates for the use of combination therapy to prevent or delay the emergence of resistant bacteria during antimicrobial treatment.83 This strategy leverages the synergistic action between drugs to enhance therapeutic efficacy and mitigate the selection pressure for drug-resistant bacteria. In an experiment conducted by Ji et al84 the results of drug sensitivity tests indicated that the combination of cefepime with amoxicillin and clavulanate achieved a significant synergistic effect in vitro, thereby reducing the minimum inhibitory concentration (MIC) of the respective antibacterial agents when the pathogens exhibited high resistance to carbapenems and cephalosporins. Additionally, other studies have demonstrated that tigecycline and polymyxins also exhibit synergistic effects in vitro.85
Phage therapy can be classified into two categories: lytic phages and lysogenic phages, based on their distinct life cycles. Lytic phages, in particular, are promising candidates for clinical treatment.86 Research indicates that the lytic phage KPO1K2 effectively eradicates biofilms produced by K. pneumoniae, and when administered in conjunction with ciprofloxacin, it can significantly reduce the emergence of drug-resistant mutant strains.66 In Western countries, phage therapy has re-emerged as a viable alternative to antibiotics for managing complex infections.87,88 A notable case report documented the successful treatment of a patient with a prosthetic infection caused by KP.89 pneumoniae using personalized intravenous phage therapy, which ultimately prevented the need for amputation. Furthermore, bacteriophage PSD has the ability to degrade bacterial polysaccharides externally; however, its application in vivo has thus far been restricted to animal studies. Tu et al demonstrated that this enzyme can markedly enhance the survival rate of mice infected with high doses of K. pneumoniae by analyzing the molecular structure and mechanism of PSD within the K1 type capsule.90
The emergence of resistance to K. pneumoniae presents a significant challenge in clinical treatment. The substantial financial investment and lengthy research and development cycle associated with new antimicrobial drugs have resulted in the rate of drug resistance outpacing the development of new antibiotics.91 Currently, the predominant approach in the research and development of new antibacterial agents involves the extraction of natural products or the artificial synthesis of novel chemical structures targeting bacterial infections. Subsequent development is guided by the antibacterial efficacy of these compounds. Overcoming bacterial resistance is critical for drug development, and progress has been made in identifying potential metallo-beta-lactamase inhibitors, including phthalic acid, mitoxantrone, and sulfonyl-triazole analogues.92 Furthermore, some studies have suggested that amlodipine and pantoprazole may partially reverse the resistance of Acinetobacter baumannii to imipenem; however, further investigation is required to determine whether amlodipine and pantoprazole are effective as efflux pump inhibitors (EPIs) for KP.93
Antibody therapy, it has been mentioned in studies that by inhibiting anti-alphagal antibodies in human serum, it can increase the bactericidal ability against KP and multi-drug resistant strains.94 This suggests that by modulating the interaction of antibodies with bacteria, it may help to improve the effectiveness of treatment. Another study conducted the development of a monoclonal IgG3 antibody 8E3 against K. pneumoniae K2 capsular polysaccharide.95 This antibody is specific and is able to recognize and bind to the bacteria’s capsule polysaccharide, which may help neutralize the bacteria and prevent infection. Monoclonal antibodies that act directly on the K1 serotype or the LPS-O antigen portion have been successfully used to prevent and treat hvKP infection.96
Advantages and Challenges
Klebsiella, a prevalent pathogen, exhibits a variety of drug resistance mechanisms, including the production of β-lactamases, alterations in outer membrane proteins, and the activation of drug efflux pumps. These mechanisms provide substantial material for the study of microbial resistance. In terms of detection methods, molecular biology technologies have made significant advancements in recent years. The application of polymerase chain reaction (PCR), gene sequencing, and mass spectrometry has enhanced the sensitivity and accuracy of drug resistance gene detection. This not only improves the efficiency of clinical diagnosis but also provides a foundation for the formulation of individualized treatment plans. In clinical applications, accurate detection methods enable physicians to swiftly identify drug-resistant strains, allowing for the selection of appropriate antibiotics for treatment. This approach reduces the unnecessary use of broad-spectrum antibiotics and mitigates the risk of further dissemination of drug-resistant strains. Furthermore, understanding the resistance mechanisms of Klebsiella contributes to the development of new antimicrobial agents and treatment strategies, thereby advancing the fields of medicine and pharmacy.
Despite numerous advancements in research and practice, the application of Klebsiella resistance mechanisms and their detection methods continues to encounter several challenges. First, the diversity and complexity of drug-resistant strains complicate research efforts, as different strains may develop resistance through various mechanisms, thereby complicating detection and diagnosis. Second, while current detection methods exhibit sensitivity, considerations regarding cost and time remain critical in clinical applications, particularly in medical settings with limited resources. Striking a balance between accuracy and cost-effectiveness in detection is an urgent issue that requires resolution. Furthermore, as resistant strains evolve, new resistance mechanisms may emerge, necessitating continuous updates to detection methods and diagnostic criteria to address these evolving challenges. To effectively curb the spread of drug-resistant strains, it is essential to enhance multidisciplinary cooperation and raise awareness among both the public and medical personnel.
Conclusion and Prospect
The issue of multidrug resistance in K. pneumoniae presents a significant challenge to global public health and clinical treatment. This paper provides an in-depth exploration of the epidemiological characteristics and complex resistance mechanisms associated with KP, highlighting the crucial roles of drug sensitivity testing and PCR technology in resistance detection. Although current treatment strategies, such as combination therapies, phage therapy, novel antimicrobial agents, and antibody therapies, have shown promise in mitigating KP resistance to some extent, further research and clinical validation are essential. In the long term, as the efficacy of existing antibiotics diminishes, the development of new antibacterial agents becomes increasingly vital. This development should not only aim to overcome the limitations of traditional antibiotics but also consider their environmental and ecological impacts. Bacteriophage therapy, recognized for its targeted approach and minimal side effects, has demonstrated considerable potential for application. However, despite encouraging results from laboratory studies, its safety and efficacy in clinical settings require validation through extensive clinical trials. Antibody therapy has also revealed distinct advantages in combating bacterial infections. Future research should focus on enhancing the development of anti-KP specific antibodies and optimizing their production and application strategies to improve clinical outcomes. Addressing the issue of resistance in KP necessitates close collaboration among clinical medicine, microbiology, pharmacy, and public health, alongside policy support from governments and relevant institutions to facilitate research advancements and effective clinical applications.
Future research must focus on multiple intertwined aspects to comprehensively address the challenges posed by KP. Firstly, efforts should be dedicated to enhancing the development of anti - KP specific antibodies and refining their production and application strategies. This is crucial as it directly contributes to improving clinical outcomes. Simultaneously, combatting KP resistance demands a collaborative approach involving clinical medicine, microbiology, pharmacy, and public health. This multi - disciplinary cooperation, complemented by strong policy support from governments and relevant institutions, is essential for driving research progress and ensuring that new findings are effectively translated into clinical applications. In addition to antibody - related research and resistance management, future research will zero in on two other key areas. One is to conduct an in - depth and comprehensive analysis of the Klebsiella resistance genome through large - scale genomic studies. This endeavor will help unearth additional potential resistance mechanisms, which in turn will serve as the cornerstone for the development of innovative detection methods. The other area of focus is the advancement of new rapid detection technologies, especially those that can provide instant results at the bedside. By significantly reducing the time from sample collection to result delivery, these technologies will greatly enhance the timeliness of clinical decision - making, ultimately leading to more effective treatment strategies.
Acknowledgments
The authors sincerely appreciate the financial support from Jinan Municipal Health Commission through its Science and Technology Development Program (Grant No. 2023-public-14). This support has been crucial for the completion of both the K. pneumoniae-related research and this article.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Dong N, Yang X, Chan EW, Zhang R, Chen S. Klebsiella species: taxonomy, hypervirulence and multidrug resistance. EBioMedicine. 2022;79:103998. doi:10.1016/j.ebiom.2022.103998
2. Piperaki ET, Syrogiannopoulos GA, Tzouvelekis LS, Daikos GL. Klebsiella pneumoniae: virulence, biofilm and antimicrobial resistance. Pediatr Infect Dis J. 2017;36(10):1002–1005. doi:10.1097/inf.0000000000001675
3. Barr JG. Klebsiella: taxonomy, nomenclature, and communication. J Clin Pathol. 1977;30(10):943–944. doi:10.1136/jcp.30.10.943
4. Montgomerie JZ. Epidemiology of Klebsiella and hospital-associated infections. Rev Infect Dis. 1979;1(5):736–753. doi:10.1093/clinids/1.5.736
5. Navon-Venezia S, Kondratyeva K, Carattoli A. Klebsiella pneumoniae: a major worldwide source and shuttle for antibiotic resistance. FEMS Microbiol Rev. 2017;41(3):252–275. doi:10.1093/femsre/fux013
6. Podschun R, Ullmann U. Klebsiella spp. as nosocomial pathogens: epidemiology, taxonomy, typing methods, and pathogenicity factors. Clin Microbiol Rev. 1998;11(4):589–603. doi:10.1128/cmr.11.4.589
7. Ye M, Tu J, Jiang J, et al. Clinical and genomic analysis of liver abscess-causing Klebsiella pneumoniae identifies new liver abscess-associated virulence genes. Front Cell Infect Microbiol. 2016;6:165. doi:10.3389/fcimb.2016.00165
8. Li B, Zhao Y, Liu C, Chen Z, Zhou D. Molecular pathogenesis of Klebsiella pneumoniae. Future Microbiol. 2014;9(9):1071–1081. doi:10.2217/fmb.14.48
9. Owaid HA, Al-Ouqaili MTS. Molecular and bacteriological investigations for the co-existence CRISPR/Cas system and β-lactamases of types extended-spectrum and carbapenemases in multidrug, extensive drug and pandrug-resistant Klebsiella pneumoniae. Saudi J Biol Sci. 2024;31(7):104022. doi:10.1016/j.sjbs.2024.104022
10. Chen Y, Chen Y. Clinical challenges with hypervirulent Klebsiella pneumoniae (hvKP) in China. J Transl Int Med. 2021;9(2):71–75. doi:10.2478/jtim-2021-0004
11. Liang Y, Qiu L, Zheng X, Liu J. Trend in antimicrobial resistance of Staphylococcus aureus: results from the China antimicrobial surveillance network (CHINET) in the Last 15-Year-period reports (2005-2019). Infect Drug Resist. 2021;14:2179–2181. doi:10.2147/idr.S318005
12. Kochan TJ, Nozick SH, Valdes A, et al. Klebsiella pneumoniae clinical isolates with features of both multidrug-resistance and hypervirulence have unexpectedly low virulence. Nat Commun. 2023;14(1):7962. doi:10.1038/s41467-023-43802-1
13. Martin RM, Bachman MA. Colonization, infection, and the accessory genome of Klebsiella pneumoniae. Front Cell Infect Microbiol. 2018;8:4. doi:10.3389/fcimb.2018.00004
14. Chen Q, Wang M, Han M, Xu L, Zhang H. Molecular basis of Klebsiella pneumoniae colonization in host. Microb Pathog. 2023;177:106026. doi:10.1016/j.micpath.2023.106026
15. Hu F, Guo Y, Yang Y, et al. Resistance reported from China antimicrobial surveillance network (CHINET) in 2018. Eur J Clin Microbiol Infect Dis. 2019;38(12):2275–2281. doi:10.1007/s10096-019-03673-1
16. Keynan Y, Rubinstein E. The changing face of Klebsiella pneumoniae infections in the community. Int J Antimicrob Agents. 2007;30(5):385–389. doi:10.1016/j.ijantimicag.2007.06.019
17. Shi Q, Lan P, Huang D, et al. Diversity of virulence level phenotype of hypervirulent Klebsiella pneumoniae from different sequence type lineage. BMC Microbiol. 2018;18(1):94. doi:10.1186/s12866-018-1236-2
18. Han X, Yao J, He J, et al. Clinical and laboratory insights into the threat of hypervirulent Klebsiella pneumoniae. Int J Antimicrob Agents. 2024;64(3):107275. doi:10.1016/j.ijantimicag.2024.107275
19. Liao Y, Gong J, Yuan X, Wang X, Huang Y, Chen X. Virulence factors and carbapenem-resistance mechanisms in hypervirulent Klebsiella pneumoniae. Infect Drug Resist. 2024;17:1551–1559. doi:10.2147/IDR.S461903
20. Wu H, Li D, Zhou H, Sun Y, Guo L, Shen D. Bacteremia and other body site infection caused by hypervirulent and classic Klebsiella pneumoniae. Microb Pathog. 2017;104:254–262. doi:10.1016/j.micpath.2017.01.049
21. Zhang QB, Zhu P, Zhang S, et al. Hypervirulent Klebsiella pneumoniae detection methods: a minireview. Arch Microbiol. 2023;205(10):326. doi:10.1007/s00203-023-03665-y
22. Arcari G, Carattoli A. Global spread and evolutionary convergence of multidrug-resistant and hypervirulent Klebsiella pneumoniae high-risk clones. Pathog Glob Health. 2023;117(4):328–341. doi:10.1080/20477724.2022.2121362
23. Renggli L, Gasser M, Plüss-Suard C, Harbarth S, Kronenberg A. Temporal and structural patterns of extended-spectrum cephalosporin-resistant Klebsiella pneumoniae incidence in Swiss hospitals. J Hosp Infect. 2022;120:36–42. doi:10.1016/j.jhin.2021.11.006
24. Spadar A, Perdigao J, Campino S, Clark TG. Large-scale genomic analysis of global Klebsiella pneumoniae plasmids reveals multiple simultaneous clusters of carbapenem-resistant hypervirulent strains. Genome Med. 2023;15(1):3. doi:10.1186/s13073-023-01153-y
25. Liu C, Dong N, Chan EWC, Chen S, Zhang R. Molecular epidemiology of carbapenem-resistant Klebsiella pneumoniae in China, 2016-20. Lancet Infect Dis. 2022;22(2):167–168. doi:10.1016/S1473-3099(22)00009-3
26. Arakawa Y. Systematic research to overcome newly emerged multidrug-resistant bacteria. Microbiol Immunol. 2020;64(4):231–251. doi:10.1111/1348-0421.12781
27. Bertonha AF, Silva CCL, Shirakawa KT, Trindade DM, Dessen A. Penicillin-binding protein (PBP) inhibitor development: a 10-year chemical perspective. Exp Biol Med (Maywood). 2023;248(19):1657–1670. doi:10.1177/15353702231208407
28. Husna A, Rahman MM, Badruzzaman ATM, et al. Extended-spectrum β-Lactamases (ESBL): challenges and opportunities. Biomedicines. 2023;11(11):2937. doi:10.3390/biomedicines11112937
29. Tseng CH, Liu CW, Liu PY. Extended-spectrum beta-lactamases (ESBL) producing bacteria in animals. Antibiotics (Basel). 2023;12(4). doi:10.3390/antibiotics12040661
30. Livermore DM, Hope R, Mushtaq S, Warner M. Orthodox and unorthodox clavulanate combinations against extended-spectrum beta-lactamase producers. Clin Microbiol Infect. 2008;14 Suppl 1:189–193. doi:10.1111/j.1469-0691.2007.01858.x
31. Zhang Y, Zhao C, Wang Q, et al. High prevalence of hypervirulent Klebsiella pneumoniae infection in China: geographic distribution, clinical characteristics, and antimicrobial resistance. Antimicrob Agents Chemother. 2016;60(10):6115–6120. doi:10.1128/aac.01127-16
32. Shankar C, Santhanam S, Kumar M, Gupta V, Devanga Ragupathi NK, Veeraraghavan B. Draft genome sequence of an extended-spectrum-β-lactamase-positive hypervirulent Klebsiella pneumoniae strain with novel sequence type 2318 isolated from a neonate. Genome Announc. 2016;4(6). doi:10.1128/genomeA.01273-16
33. Qiu J, Wei D, Ma J, et al. Covert dissemination of pLVPK-like virulence plasmid in ST29-K54 Klebsiella pneumoniae: emergence of low virulence phenotype strains. Front Cell Infect Microbiol. 2023;13:1194133. doi:10.3389/fcimb.2023.1194133
34. Li W, Sun G, Yu Y, et al. Increasing occurrence of antimicrobial-resistant hypervirulent (hypermucoviscous) Klebsiella pneumoniae isolates in China. Clin Infect Dis. 2014;58(2):225–232. doi:10.1093/cid/cit675
35. Geleta D, Abebe G, Alemu B, Workneh N, Beyene G. Mechanisms of bacterial drug resistance with special emphasis on phenotypic and molecular characterization of extended spectrum beta-lactamase. New Microbiol. 2024;47(1):1–14.
36. Bagattini M, Crivaro V, Di Popolo A, et al. Molecular epidemiology of extended-spectrum beta-lactamase-producing Klebsiella pneumoniae in a neonatal intensive care unit. J Antimicrob Chemother. 2006;57(5):979–982. doi:10.1093/jac/dkl077
37. Khademi F, Vaez H, Neyestani Z, Sahebkar A. Prevalence of ESBL-producing Enterobacter species resistant to carbapenems in Iran: a systematic review and meta-analysis. Int J Microbiol. 2022;2022:8367365. doi:10.1155/2022/8367365
38. Di Pilato V, Pollini S, Miriagou V, Rossolini GM, D’Andrea MM. Carbapenem-resistant Klebsiella pneumoniae: the role of plasmids in emergence, dissemination, and evolution of a major clinical challenge. Expert Rev Anti Infect Ther. 2024;22(1–3):25–43. doi:10.1080/14787210.2024.2305854
39. Dunn SJ, Connor C, McNally A. The evolution and transmission of multi-drug resistant Escherichia coli and Klebsiella pneumoniae: the complexity of clones and plasmids. Curr Opin Microbiol. 2019;51:51–56. doi:10.1016/j.mib.2019.06.004
40. Tseng CH, Huang YT, Mao YC, et al. Insight into the mechanisms of carbapenem resistance in Klebsiella pneumoniae: a Study on IS26 integrons, beta-lactamases, porin modifications, and plasmidome analysis. Antibiotics (Basel). 2023;12(4). doi:10.3390/antibiotics12040749
41. Coolen JPM, den Drijver EPM, Kluytmans J, et al. Development of an algorithm to discriminate between plasmid- and chromosomal-mediated AmpC β-lactamase production in Escherichia coli by elaborate phenotypic and genotypic characterization. J Antimicrob Chemother. 2019;74(12):3481–3488. doi:10.1093/jac/dkz362
42. Wang S, Yin B, Yu L, et al. Overexpression of AmpC promotes bacteriophage lysis of ampicillin-resistant Escherichia coli. Front Microbiol. 2019;10:2973. doi:10.3389/fmicb.2019.02973
43. Hanson ND, Sanders CC. Regulation of inducible AmpC beta-lactamase expression among Enterobacteriaceae. Curr Pharm Des. 1999;5(11):881–894. doi:10.2174/1381612805666230112191507
44. Wang XD, Cai JC, Zhou HW, Zhang R, Chen GX. Reduced susceptibility to carbapenems in Klebsiella pneumoniae clinical isolates associated with plasmid-mediated beta-lactamase production and OmpK36 porin deficiency. J Med Microbiol. 2009;58(Pt 9):1196–1202. doi:10.1099/jmm.0.008094-0
45. Pichardo C, Rodríguez-Martínez JM, Pachón-Ibañez ME, et al. Efficacy of cefepime and imipenem in experimental murine pneumonia caused by porin-deficient Klebsiella pneumoniae producing CMY-2 beta-Lactamase. Antimicrob Agents Chemother. 2005;49(8):3311–3316. doi:10.1128/aac.49.8.3311-3316.2005
46. Ting SMV, Ismail Z, Hanafiah A. Prevalence of AmpC beta-lactamase and extended spectrum beta-lactamase co-producer in Escherichia coli and Klebsiella species in a teaching hospital. Malays J Pathol. 2024;46(1):79–89.
47. Spaziante M, Oliva A, Ceccarelli G, Venditti M. What are the treatment options for resistant Klebsiella pneumoniae carbapenemase (KPC)-producing bacteria? Expert Opin Pharmacother. 2020;21(15):1781–1787. doi:10.1080/14656566.2020.1779221
48. Yigit H, Queenan AM, Anderson GJ, et al. Novel carbapenem-hydrolyzing beta-lactamase, KPC-1, from a carbapenem-resistant strain of Klebsiella pneumoniae. Antimicrob Agents Chemother. 2001;45(4):1151–1161. doi:10.1128/aac.45.4.1151-1161.2001
49. Xu Q, Lin H, Liu W, et al. Genomic characterization of Escherichia coli co-producing KPC-2 and NDM-5 carbapenemases isolated from intensive care unit in a Chinese hospital. Microb Drug Resist. 2024;30(1):27–36. doi:10.1089/mdr.2023.0050
50. Birgy A, Magnan M, Hobson CA, et al. Local and global protein interactions contribute to residue entrenchment in beta-lactamase TEM-1. Antibiotics (Basel). 2022;11(5). doi:10.3390/antibiotics11050652
51. Zhou G, Wang Q, Wang Y, et al. Outer membrane porins contribute to antimicrobial resistance in gram-negative bacteria. Microorganisms. 2023;11(7):1690. doi:10.3390/microorganisms11071690
52. de Moura VCN, Verma D, Everall I, et al. Increased virulence of outer membrane porin mutants of Mycobacterium abscessus. Front Microbiol. 2021;12:706207. doi:10.3389/fmicb.2021.706207
53. Webster DP, Gaulton T, Woodford N, et al. Emergence of carbapenem resistance due to porin loss in an extended-spectrum β-lactamase (ESBL)-producing Klebsiella pneumoniae strain during meropenem therapy. Int J Antimicrob Agents. 2010;36(6):575–576. doi:10.1016/j.ijantimicag.2010.08.003
54. Yang D, Guo Y, Zhang Z. Combined porin loss and extended spectrum beta-lactamase production is associated with an increasing imipenem minimal inhibitory concentration in clinical Klebsiella pneumoniae strains. Curr Microbiol. 2009;58(4):366–370. doi:10.1007/s00284-009-9364-4
55. Gaibani P, Bianco G, Amadesi S, Boattini M, Ambretti S, Costa C. Increased bla(KPC) Copy Number and OmpK35 and OmpK36 Porins disruption mediated resistance to imipenem/relebactam and meropenem/vaborbactam in a KPC-producing Klebsiella pneumoniae clinical isolate. Antimicrob Agents Chemother. 2022;66(5):e0019122. doi:10.1128/aac.00191-22
56. Hussein RA, Al-Kubaisy SH, Al-Ouqaili MTS. The influence of efflux pump, outer membrane permeability and beta-lactamase production on the resistance profile of multi, extensively and pandrug resistant Klebsiella pneumoniae. J Infect Public Health. 2024;17(11):102544. doi:10.1016/j.jiph.2024.102544
57. Abdi SN, Ghotaslou R, Ganbarov K, et al. Acinetobacter baumannii efflux pumps and antibiotic resistance. Infect Drug Resist. 2020;13:423–434. doi:10.2147/idr.S228089
58. Al-Ani NFI. Biofilm antimicrobial susceptibility pattern for selected antimicrobial agents against planktonic and sessile cells of clinical isolates of staphylococci using MICs, BICs and MBECs. Asian Journal of Pharmaceutics. 2018;12(4):S1375–S1383.
59. Jang S. AcrAB-TolC, a major efflux pump in Gram negative bacteria: toward understanding its operation mechanism. BMB Rep. 2023;56(6):326–334. doi:10.5483/BMBRep.2023-0070
60. Wan Nur Ismah WAK, Takebayashi Y, Findlay J, Heesom KJ, Avison MB. Impact of OqxR loss of function on the envelope proteome of Klebsiella pneumoniae and susceptibility to antimicrobials. J Antimicrob Chemother. 2018;73(11):2990–2996. doi:10.1093/jac/dky293
61. De Majumdar S, Yu J, Fookes M, et al. Elucidation of the RamA regulon in Klebsiella pneumoniae reveals a role in LPS regulation. PLoS Pathog. 2015;11(1):e1004627. doi:10.1371/journal.ppat.1004627
62. Abdelkreem RH, Yousuf AM, Elmekki MA, Elhassan MM. DNA gyrase and topoisomerase IV mutations and their effect on quinolones resistant proteus mirabilis among UTIs patients. Pak J Med Sci. 2020;36(6):1234–1240. doi:10.12669/pjms.36.6.2207
63. Macesic N, Nelson B, McConville TH, et al. Emergence of polymyxin resistance in clinical Klebsiella pneumoniae through diverse genetic adaptations: a genomic, retrospective cohort study. Clin Infect Dis. 2020;70(10):2084–2091. doi:10.1093/cid/ciz623
64. Nagy L, Urbán P, Makszin L, et al. The effect of mutation in lipopolysaccharide biosynthesis on bacterial fitness. Cells. 2022;11(20):3249. doi:10.3390/cells11203249
65. Yin D, Guo Y, Li M, et al. Performance of VITEK 2, E-test, Kirby-Bauer disk diffusion, and modified Kirby-Bauer disk diffusion compared to reference broth microdilution for testing tigecycline susceptibility of carbapenem-resistant K. pneumoniae and A. baumannii in a multicenter study in China. Eur J Clin Microbiol Infect Dis. 2021;40(6):1149–1154. doi:10.1007/s10096-020-04123-z
66. Kansak N, Arici N, Uzunoner Y, Adaleti R, Aksaray S, Gonullu N. Evaluation of broth disk elution method to determine colistin resistance in Klebsiella pneumoniae and Escherichia coli strains. Clin Lab. 2023;69(2). doi:10.7754/Clin.Lab.2022.221008
67. Pereira JV, Bari AK, Kokare R, Poojary A. Comparison of in vitro fosfomycin susceptibility testing methods with agar dilution for carbapenem resistant Klebsiella pneumoniae and Escherichiacoli. Indian J Med Microbiol. 2023;42:39–45. doi:10.1016/j.ijmmb.2023.01.005
68. Kee PS, Karunanathie H, Maggo SDS, Kennedy MA, Chua EW. Long-Range Polymerase Chain Reaction. Methods mol Biol. 2023;2967:181–192. doi:10.1007/978-1-0716-3358-8_15
69. Lagos-Susaeta D, Salazar O, Asenjo JA. openPFGE: an open source and low cost pulsed-field gel electrophoresis equipment. HardwareX. 2020;8:e00128. doi:10.1016/j.ohx.2020.e00128
70. Tenover FC, Arbeit RD, Goering RV, et al. Interpreting chromosomal DNA restriction patterns produced by pulsed-field gel electrophoresis: criteria for bacterial strain typing. J Clin Microbiol. 1995;33(9):2233–2239. doi:10.1128/jcm.33.9.2233-2239.1995
71. Park MN, Yeo SG, Park J, Jung Y, Hwang SM. Usefulness and limitations of PFGE diagnosis and nucleotide sequencing method in the analysis of food poisoning pathogens found in cooking employees. Int J mol Sci. 2024;25(7). doi:10.3390/ijms25074123
72. Gits-Muselli M, Campagne P, Desnos-Ollivier M, et al. Comparison of multilocus sequence typing (MLST) and microsatellite length polymorphism (MLP) for Pneumocystis jirovecii genotyping. Comput Struct Biotechnol J. 2020;18:2890–2896. doi:10.1016/j.csbj.2020.10.005
73. Nordmann P, Poirel L, Dortet L. Rapid detection of carbapenemase-producing Enterobacteriaceae. Emerg Infect Dis. 2012;18(9):1503–1507. doi:10.3201/eid1809.120355
74. Nordmann P, Naas T, Poirel L. Global spread of Carbapenemase-producing Enterobacteriaceae. Emerg Infect Dis. 2011;17(10):1791–1798. doi:10.3201/eid1710.110655
75. Mancini S, Kieffer N, Poirel L, Nordmann P. Evaluation of the RAPIDEC® CARBA NP and β-CARBA® tests for rapid detection of Carbapenemase-producing Enterobacteriaceae. Diagn Microbiol Infect Dis. 2017;88(4):293–297. doi:10.1016/j.diagmicrobio.2017.05.006
76. Yu J, Liu J, Li Y, et al. Rapid detection of carbapenemase activity of Enterobacteriaceae isolated from positive blood cultures by MALDI-TOF MS. Ann Clin Microbiol Antimicrob. 2018;17(1):22. doi:10.1186/s12941-018-0274-9
77. Founou RC, Founou LL, Allam M, Ismail A, Essack SY. Whole genome sequencing of extended spectrum β-lactamase (ESBL)-producing Klebsiella pneumoniae isolated from hospitalized patients in KwaZulu-Natal, South Africa. Sci Rep. 2019;9(1):6266. doi:10.1038/s41598-019-42672-2
78. Zhu H, Zhang H, Xu Y, Laššáková S, Korabečná M, Neužil P. PCR past, present and future. Biotechniques. 2020;69(4):317–325. doi:10.2144/btn-2020-0057
79. Logan LK, Weinstein RA. The epidemiology of carbapenem-resistant Enterobacteriaceae: the impact and evolution of a global menace. J Infect Dis. 2017;215(suppl_1):S28–s36. doi:10.1093/infdis/jiw282
80. Ferreira C, Otani S, Aarestrup FM, Manaia CM. Quantitative PCR versus metagenomics for monitoring antibiotic resistance genes: balancing high sensitivity and broad coverage. FEMS Microbes. 2023;4:xtad008. doi:10.1093/femsmc/xtad008
81. Bandick RG, Mousavi S, Bereswill S, Heimesaat MM. Review of therapeutic options for infections with carbapenem-resistant Klebsiella pneumoniae. Eur J Microbiol Immunol (Bp). 2020;10(3):115–124. doi:10.1556/1886.2020.00022
82. Zhao Y, Li C, Zhang J, et al. The in vitro activity of polymyxin B and tigecycline alone and combination with other antibiotics against carbapenem-resistant Enterobacter cloacae complex isolates, including high-risk clones. Ann Transl Med. 2019;7(23):779. doi:10.21037/atm.2019.11.33
83. Tamma PD, Cosgrove SE, Maragakis LL. Combination therapy for treatment of infections with gram-negative bacteria. Clin Microbiol Rev. 2012;25(3):450–470. doi:10.1128/cmr.05041-11
84. Ji S, Lv F, Du X, et al. Cefepime combined with amoxicillin/clavulanic acid: a new choice for the KPC-producing K. pneumoniae infection. Int J Infect Dis. 2015;38:108–114. doi:10.1016/j.ijid.2015.07.024
85. Okanda T, Matsumoto T. In vitro effect of an antimicrobial combination therapy without colistin and tigecycline for CPE and non-CPE. J Infect Chemother. 2020;26(4):322–330. doi:10.1016/j.jiac.2019.12.004
86. Alipour-Khezri E, Skurnik M, Zarrini G. Pseudomonas aeruginosa bacteriophages and their clinical applications. Viruses. 2024;16(7):1051. doi:10.3390/v16071051
87. Kortright KE, Chan BK, Koff JL, Turner PE. Phage therapy: a renewed approach to combat antibiotic-resistant bacteria. Cell Host Microbe. 2019;25(2):219–232. doi:10.1016/j.chom.2019.01.014
88. Gordillo Altamirano FL, Barr JJ. Phage therapy in the postantibiotic era. Clin Microbiol Rev. 2019;32(2). doi:10.1128/cmr.00066-18
89. Cano EJ, Caflisch KM, Bollyky PL, et al. Phage therapy for limb-threatening prosthetic knee Klebsiella pneumoniae infection: case report and in vitro characterization of anti-biofilm activity. Clin Infect Dis. 2021;73(1):e144–e151. doi:10.1093/cid/ciaa705
90. Tu IF, Lin TL, Yang FL, et al. Structural and biological insights into Klebsiella pneumoniae surface polysaccharide degradation by a bacteriophage K1 lyase: implications for clinical use. J Biomed Sci. 2022;29(1):9. doi:10.1186/s12929-022-00792-4
91. Årdal C, Balasegaram M, Laxminarayan R, et al. Antibiotic development - economic, regulatory and societal challenges. Nat Rev Microbiol. 2020;18(5):267–274. doi:10.1038/s41579-019-0293-3
92. Papp-Wallace KM, Endimiani A, Taracila MA, Bonomo RA. Carbapenems: past, present, and future. Antimicrob Agents Chemother. 2011;55(11):4943–4960. doi:10.1128/aac.00296-11
93. Abdulaal WH, Alhakamy NA, Asseri AH, et al. Redirecting pantoprazole as a metallo-beta-lactamase inhibitor in carbapenem-resistant Klebsiella pneumoniae. Front Pharmacol. 2024;15:1366459. doi:10.3389/fphar.2024.1366459
94. Olivera-Ardid S, Bello-Gil D, Perez-Cruz M, et al. Removal of natural anti-αGal antibodies elicits protective immunity against Gram-negative bacterial infections. Front Immunol. 2023;14:1232924. doi:10.3389/fimmu.2023.1232924
95. Wang-Lin SX, Olson R, Beanan JM, MacDonald U, Russo TA, Balthasar JP. Antibody dependent enhancement of Acinetobacter baumannii infection in a mouse pneumonia model. J Pharmacol Exp Ther. 2019;368(3):475–489. doi:10.1124/jpet.118.253617
96. Singh S, Wilksch JJ, Dunstan RA, et al. LPS O antigen plays a key role in Klebsiella pneumoniae capsule retention. Microbiol Spectr. 2022;10(4):e0151721. doi:10.1128/spectrum.01517-21
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms.php
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms.php
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Epidemiology, Drug Resistance, and Risk Factors for Mortality Among Hematopoietic Stem Cell Transplantation Recipients with Hospital-Acquired Klebsiella pneumoniae Infections: A Single-Center Retrospective Study from China
Liu YF, Liu Y, Chen X, Jia Y
Infection and Drug Resistance 2022, 15:5011-5021
Published Date: 30 August 2022
Clinical Characteristics, Drug Resistance, and Risk Factors for Death of Klebsiella pneumoniae Infection in Patients with Acute Pancreatitis: A Single-Center Retrospective Study from China
Jia Y, Liu Y, Huang Y, Wang J, Wang H, Tan S, Shi Y, Wang Q, Peng J
Infection and Drug Resistance 2023, 16:5039-5053
Published Date: 7 August 2023