")
Back to Journals » Infection and Drug Resistance » Volume 18
Molecular Characteristics and Phenotypical Analysis of Carbapenem-Resistant K. Pneumoniae in the Lüliang Region, Shanxi Province
Authors Wang H , Jia Z, Li X, Hao Y, Zhang J , Zhao X , Li H, Jin S
Received 21 December 2024
Accepted for publication 28 May 2025
Published 7 June 2025 Volume 2025:18 Pages 2911—2921
DOI https://doi.org/10.2147/IDR.S512203
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 6
Editor who approved publication: Dr Sandip Patil
Huiyuan Wang,1,2,* Zijiang Jia,2,* Xiaoyan Li,2 Yan Hao,1 Jinjing Zhang,2 Xinyi Zhao,1 Hui Li,1 Sifan Jin1
1Department of Pathogenic Biology, School of Fenyang College of Shanxi Medical University, Fenyang, Shanxi, People’s Republic of China; 2Department of Clinical Laboratory, Shanxi Province, Fenyang Hospital Affiliated with Shanxi Medical University, Fenyang, Shanxi, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Huiyuan Wang, Email [email protected]
Introduction: To explore the resistance characteristics and molecular features of carbapenem-resistant K. pneumoniae (CRKP) isolates prevalent in the Lüliang region, 81 CRKP isolates were collected from three hospitals in the Lüliang region, Shanxi Province.
Methods: The resistance of these CRKP isolates to 11 antibiotics was determined using the disk diffusion method, and antimicrobial resistance encoding genes and virulence genes were detected by PCR. The mucoid phenotype of the CRKP isolates was examined via the string test, and bacterial biofilm formation ability was measured using the crystal violet staining method.
Results: The resistance rates of the 81 CRKP isolates to the 11 antibiotics ranged from 62.96% to 100%, with a multidrug resistance rate of 83.95%. The resistance genes blaSHV, blaTEM and blaKPC were the most widely distributed, with a detection rate of 100%. Among the 81 CRKP isolates, 70 had the ability to form biofilms, and 58 presented highly mucoid phenotypes. The virulence genes rmpA2, peg-344, and fimH presented high carriage rates of 92.59%, 91.36%, and 88.89%, respectively. The carriage rate of IroB was low, at 20.99%. Among these genes, fimH, rmpA2, and iucA were associated with biofilm formation, while markD and fimH were associated with a highly mucoid phenotype, and the highly mucoid phenotype was strongly correlated with the biofilm formation ability.
Conclusion: This study revealed that the CRKP strains isolated in the Lüliang region of Shanxi Province were strongly resistant and that this resistance was related to virulence characteristics. Therefore, antibiotic management should be strengthened in clinical practice to control the prevalence of CRKP in this region.
Keywords: CRKP, resistance, virulence, biofilm, ucoid phenotypes
Introduction
Klebsiella pneumoniae (K. pneumoniae) is an opportunistic pathogen bacterium that is widely present in hospital environments and isolates of this bacterium have developed resistance to multiple antibiotics, causing severe community-associated infections and posing a significant threat to human health.1 K. pneumoniae often resides in the human intestine, and can cause infections in the respiratory system, urinary system, and skin and soft tissues, especially in patients with compromised immune systems.2,3 Currently, cephalosporins and carbapenems are commonly used to treat K. pneumoniae infections in clinical settings. However, the extensive use of these antibiotics has led to the emergence of multidrug-resistant and pan-resistant CRKP isolates. In particular, the detection rate of carbapenem-resistant KP has increased in recent years,4 making the prevention and treatment of the associated diseases more challenging.
The resistance mechanisms of CRKP are diverse, with the main mechanism being the production of carbapenemases,5 which can hydrolyze various carbapenem antibiotics, including meropenem, imipenem, and ertapenem. In 1997, the first CRKP strain was discovered in North America,6 and since then, reports of CRKP infections have increased globally.7 The spread of CRKP has become a severe problem, especially in some regions where the resistance rate of CRKP is very high, exceeding 50%.8 In China, according to the report of the National Antimicrobial Resistance Monitoring System (China Antimicrobial Resistance Surveillance System, CARSS), the prevalence and characteristics of CRKP vary across different regions. Therefore, understanding the phenotypic and molecular characteristics of CRKP isolates from specific regions is highly useful for guiding clinical drug use. According to an analysis by the CARSS, the resistance rate of CRKP has increased in recent years. However, there are few reports on the prevalence characteristics, resistance status, and virulence-related characteristics of CRKP in the Lüliang region of Shanxi Province. This study aims to provide effective theoretical and data support for the prevention and treatment of CRKP infections in this region by conducting research on the prevalence characteristics, phenotypic resistance, resistance genes, and virulence genes of CRKP in the Lüliang region of Shanxi Province.
Materials and Methods
Strain Source
In this study, 81 CRKP isolates were collected from three hospitals (Fenyang Hospital of Shanxi Province, Fenyang City People’s Hospital, and Fenyang City Maternal and Child Health Care Hospital) in the Lüliang region of Shanxi Province (Figure 1) between December 2022 and December 2023. The clinical information of the patients, including symptoms and strain sources, was collected on the basis of the patient’s medical record number. These data were obtained with the consent of the hospital. The CRKP isolates were obtained according to the “National Clinical Laboratory Procedures”. The isolates were sequentially labeled KP1-KP81. The control strains used for the clinical susceptibility test were Escherichia coli ATCC25922 and K. pneumoniae ATCC-BAA-1705H and ATCC-BAA-1706, which were used as positive and negative control strains for the mCIM test, respectively.
 |
Figure 1 The source locations of the 81 carbapenem-resistant K. pneumoniae isolates. Notes: The red dots represent the hospital where the samples were collected. |
Figure 1. The location of the hospital where the samples were collected. The red dots represent the areas where the samples were collected.
Carbapenemase Phenotype Confirmation
The modified carbapenem inactivation method (mCIM) recommended by the CLSI20209 was used to detect carbapenemase in the 81 clinical CRKP isolates. The specific method was as follows: A full loop of the test colony was inoculated into 2 mL of tryptic soy broth (TSB, Beijing Solarbio Science & Technology China) and vortexed for 15s to mix. A 10 μg meropenem disk was placed into the tube with sterile forceps, fully immersed in the bacterial suspension, sealed, and incubated at 35 ± 2 °C for 4 hours. Escherichia coli ATCC 25922 was prepared as a 0.5 McFarland bacterial suspension, inoculated onto Mueller-Hinton agar (MHA, Beijing Solarbio Science & Technology China) solid medium, and dried for 5 minutes. The meropenem disk was removed from the tube with sterile forceps, squeezed dry, and placed on the previously prepared MHA medium. The plate was incubated at 35 ± 2 °C for 18–24 hours, and the inhibition zone diameter was measured with a caliper. The results were interpreted according to the carbapenemase confirmation criteria of the CLSI 2020 version: (1) colonies with an inhibition zone diameter of 6–15 mm or scattered colonies within a 16–18 mm inhibition zone were considered carbapenemase positive. (2) colonies with an inhibition zone diameter ≥ 19 mm were considered carbapenemase negative. (3) The two scenarios can be classified as indeterminate for carbapenemase production: an inhibition zone diameter of 16–18 mm or ≥19 mm combined with scattered colonies within the inhibition zone.
Determination of the Antibiotic Resistance Phenotype of Carbapenem-Resistant K. Pneumoniae
The susceptibility of the 81 CRKP isolates to 11 antibiotics was determined via the disk diffusion method recommended by the CLSI. These antibiotics included ceftazidime (CAZ, 30 μg), cefoperazone (CFP, 30 μg), ceftriaxone (CRO, 30 μg), imipenem (IPN, 10 μg), meropenem (MEM, 10 μg), ciprofloxacin (CIP, 5 μg), levofloxacin (LVF, 5 μg), amikacin (AK, 30 μg), gentamicin (CN, 30 μg), trimethoprim-sulfamethoxazole (SXT, 25 μg), and ampicillin (AMP, 25 μg). The susceptibility results were interpreted according to the CLSI 2020 standards. CRKP isolates resistant to three or more classes of antibiotics were considered multidrug resistant.10
Examination of the Mucoid Phenotype of Carbapenem-Resistant K. Pneumoniae
The mucoid phenotype of the CRKP isolates was examined via the string test. The specific procedure was as follows: CRKP isolates were transferred to Columbia blood agar plates and incubated at 37 °C for 16–20 hours. A loopful of the colony was lifted with a loop, and if a mucous filament longer than 0.5 cm formed, the strain was considered hypermucoviscous (HM); otherwise, it was considered non-hypermucoviscous (NO HM).11
Determination of the Biofilm Formation Ability of Carbapenem-Resistant K. Pneumoniae
The biofilm formation ability of K. pneumoniae in vitro was measured via the crystal violet staining method in a 96-well plate, as referenced in the literature.1 The specific operation steps were as follows: A standard 96-well microtiter plate was prepared, with 200 μL of tryptic soy broth added to each well. A single colony of the fresh isolate was inoculated into the broth medium and grown with shaking 120 r/min at 37 °C for 16–18 hours. Then, 2 μL of the bacterial suspension was added to 200 μL of broth medium, with uninoculated broth medium serving as the blank control. The culture was incubated at 37 °C for 48 hours. After incubation, the optical density (OD1) at 570 nm was measured using a microplate reader, with the blank control representing the OD10. The medium was gently aspirated, and the wells were washed three times with sterile physiological saline to remove extracellular planktonic bacteria. The plate was then dried in an oven at 60 °C for 20 minutes. Two hundred microliters of crystal violet solution was added to each well, and the samples were stained for 20 minutes. After staining, the crystal violet solution was aspirated, and the wells were washed four times with sterile physiological saline to remove the dye from the well walls, followed by drying the 96-well plate in a sterile hood for 30 minutes. To each well, 200 μL of 95% ethanol was added, the mixture was gently shaken for 30 minutes at 60 r/min, and the optical density (OD2) at 570 nm was measured, with the blank control as the OD20. The biofilm formation ability was calculated using the formula B = (OD2 - OD20) / (OD1 - OD10).Strains with B < 0.1 were considered nonadherent; strains with B ≥ 0.1 were considered adherent; strains with 0.1 < B ≤ 1.0 were considered moderately adherent; and strains with B > 1.0 were considered strongly adherent.
Detection of Resistance and Virulence Genes
Bacterial genomic DNA was extracted via the boiling method.12 Using this DNA as a template, PCR (Applied Biosystem America) was used to amplify the resistance and virulence genes of CRKP. The resistance genes included carbapenemase genes (blaKPC, blaNDM, blaOXA-48, blaIMP), ESBL enzyme-encoding genes (blaCTX-M, blaTEM, blaSHV), quinolone resistance genes (qnrB, oqxA), and aminoglycoside resistance genes (aac). The virulence genes included adhesion genes (fimH, markD), mucoid phenotype-related genes (rmpA, rmpA2, magA, IroB, peg-344), and iron carrier genes (iucA). The PCR primer information for the resistance and virulence genes is shown in Table 1.
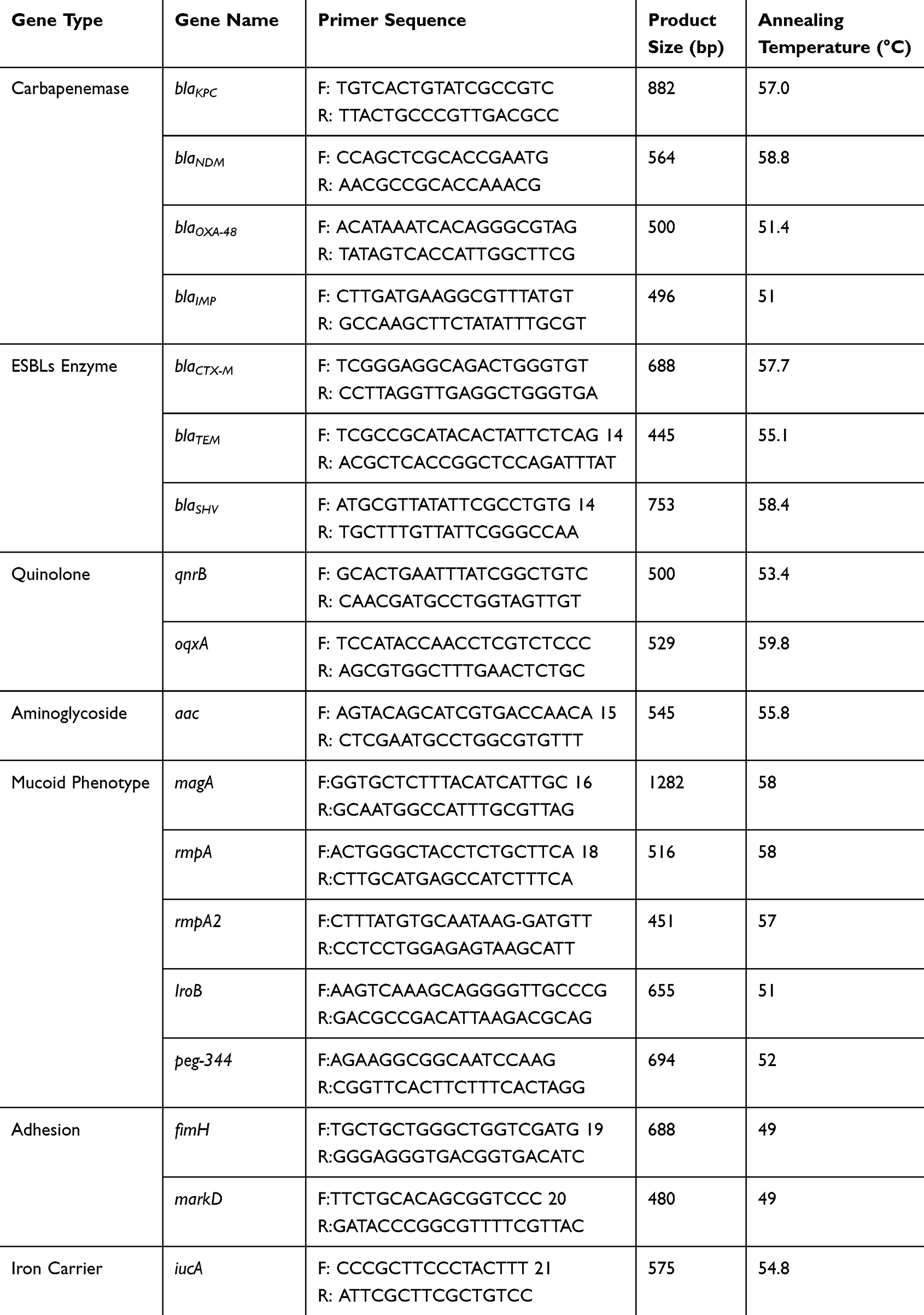 |
Table 1 PCR Primer Sequences for Resistance and Virulence Genes |
Statistical Analysis
GraphPad Prism software version 9.0 was used to conduct all the statistical comparisons. A nonparametric t-test and one-way ANOVA were used to compare the different groups. A Tukey post-hoc test was used to determine pairwise differences where appropriate. p-value of 0.05 or less was considered to be indicative of statistical significance. Pearson correlation analysis was used to analyze the correlations between phenotypic and molecular characteristics.
Results
Clinical Distribution Characteristics of the 81 CRKP Isolates
The CRKP isolates collected in this study were distributed in 13 different departments, with 30 CRKP isolates from the ICU, accounting for 37.04%, followed by the respiratory department, with 15 CRKP isolates, accounting for 18.52% (15/81), and the neurosurgery and oncology departments, each with 7 CRKP isolates, accounting for 8.64%. The distribution of the 81 CRKP isolates in the different departments is shown in Table 2. These CRKP isolates were isolated mainly from sputum (67.9%, 55/81), blood (12.35%, 10/81), and urine (9.88%), with the specific distributions shown in Table 3.
 |
Table 2 Distribution of the 81 CRKP Strains Among Clinical Departments |
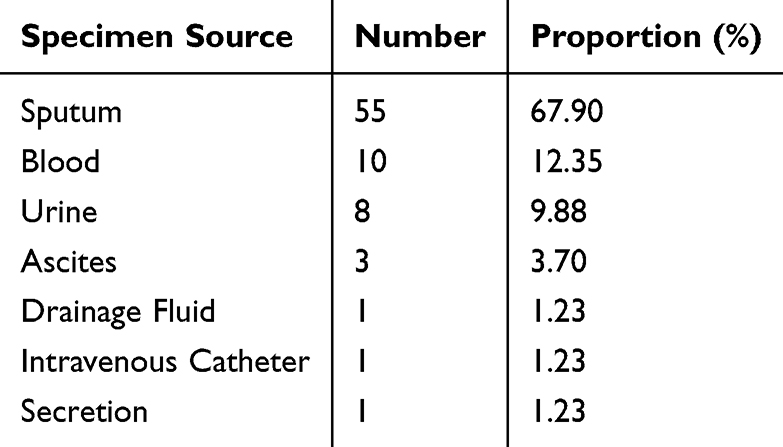 |
Table 3 Distribution of the 81 CRKP Strains Among Different Sample Types |
Results of Antimicrobial Susceptibility Testing for the 81 CRKP Isolates
We selected 11 drugs from 4 categories of antibiotics for antimicrobial susceptibility testing. All the CRKP isolates were resistant to imipenem, meropenem, ceftazidime, and ceftriaxone. The resistance rate for cefoperazone-sulbactam was 90.12%. The resistance rates for ciprofloxacin and levofloxacin were 97.53% and 96.30%, respectively. The resistance rates for amikacin and gentamicin were 77.78% and 82.72%, respectively. The resistance rate for sulfonamide drugs was relatively low, at 62.96%. The multidrug resistance rate of the 81 CRKP isolates was 83.95%. The specific results are shown in Table 4.
 |
Table 4 Resistance of the 81 K. Pneumoniae Strains to 11 Commonly Used Antibiotics |
Characteristics of the Resistance Genes Carried by the 81 CRKP Isolates
All 81 CRKP isolates prevalent in this region carried the blaSHV and blaTEM genes, and the carriage rate of the blaCTX-M gene was 55.56%. The number of CRKP isolates carrying all three resistance genes was 45, accounting for 55.56% (Table 4). The only gene detected for carbapenem resistance was blaKPC. Genes for quinolones resistance were also widely distributed in this sample, with detection rates of 53.09% for qnrB and 28.40% and 30.86% for oqxA and oqxB, respectively. The number of CRKP isolates carrying qnrB + oqxA was 13, the number of isolates carrying qnrB + oqxB was 15, and the isolates carrying all three resistance genes accounted for 16.05%. The number of CRKP isolates carrying the aminoglycoside resistance gene aac was 31, accounting for 38.27%. The frequency of antimicrobial resistance encoding genes carried by the 81 CRKP strains is shown in Table 5.
 |
Table 5 The Frequency of Antimicrobial Resistance Encoding Genes Carried by the 81 CRKP Strains |
Distribution of Virulence Phenotypes and Genotypes in the 81 CRKP Isolates
In terms of biofilm formation ability, 86.42% (70/81) of the CRKP isolates presented strong biofilm formation ability, whereas 71.60% (58/81) of the CRKP isolates presented a highly mucoid phenotype. The amplification of virulence genes revealed that the detection rates of the mucoid phenotype genes rmpA2 and peg-344 were the highest, accounting for 92.59% and 91.36%, respectively, followed by magA and rmpA, accounting for 82.72% and 83.95%, respectively. The carriage rate of IroB was low, with this gene detected in only 17 of the CRKP isolates detected. The genes encoding type I fimbriae (fimH) and type III fimbriae (markD) had high carriage rates of 88.89% and 65.43%, respectively; the gene encoding the iron siderophore aerobactin (iucA) had a detection rate of 83.95%. Table 6 shows the distribution of virulence phenotypes and genotypes among the 81 CRKP isolates.
 |
Table 6 Distribution of Virulence Phenotypes and Genotypes in the 81 CRKP Strains |
Correlation Analysis
On the basis of the Pearson correlation coefficient, we analyzed the correlations among the resistance phenotype, virulence phenotype, resistance genes, and virulence genes of the 81 CRKP isolates to determine whether there were potential connections among these factors. The correlation analysis results are shown. The formation of CRKP biofilms was significantly positively correlated with the levels of fimH (r = 0.689), rmpA2 (r = 0.490), iucA (r = 0.453), and HMV (r = 0.643). The highly mucoid phenotype of CRKP was significantly positively correlated with the levels of markD (r = 0.643), BF (r = 0.643), and fimH (r = 0.414). In the resistance analysis, the distribution of multidrug-resistant bacteria was strongly positively correlated with the resistance to cefoperazone/sulbactam (r = 0.757) and sulfamethoxazole (r = 0.570) and weakly negatively correlated with the resistance to ciprofloxacin and levofloxacin. Moreover, we found that multidrug resistance was correlated with the levels of resistance genes carried by bacteria, but the correlation coefficient R value was small. Multidrug resistance was negatively correlated with the formation of bacterial biofilms (r = −0.182). The CRKP phenotypes and genotypes were correlated to some extent, as shown in Figure 2.
 |
Figure 2 The correlation analysis among the resistance phenotype, virulence phenotype, resistance genes, and virulence genes of the 81 carbapenem-resistant K. pneumoniae isolates based on the Pearson correlation coefficient. |
Discussion
Clinical CRKP isolates have spread rapidly worldwide in recent years, posing a severe public health threat. However, there are few reports on the prevalence characteristics, resistance status, and virulence-related characteristics of CRKP in the Lüliang region of Shanxi Province. This is the first study providing effective theoretical and data support for the prevention and treatment of CRKP infections in this region. The number of clinically isolated K. pneumoniae strains has been increasing annually, but reports on the correlation between antimicrobial resistance phenotypes and molecular characteristics are relatively rare. In this study, 81 CRKP isolates were isolated from hospitals in Lüliang, Shanxi Province, from January 2021 to December 2023, and their resistance and virulence characteristics were studied. The susceptibility results revealed that the resistance rate of the 81 CRKP isolates was very high, with the resistance rate for sulfamethoxazole being 62.96% and the resistance rate for the other 10 antibiotics being greater than 80%. In addition, the MDR rate was 83.95%, indicating that the resistance rate of K. pneumoniae isolates prevalent in this region was high and significantly higher than that reported in the literature. Hu and colleagues recently reported that the prevalence of hypervirulent CRKP increased in prevalence from 28.2% in 2016 to 45.7% in 2020.13 We determined that the difference might be related to the molecular characteristics of K. pneumoniae prevalent in this region and the antibiotic use practices of the hospital, but further investigations of patient antibiotic use are still needed to determine the reasons for this difference.
Carbapenem antibiotics constitute the last line of defense in the treatment of infections caused by gram-negative bacteria that produce extended-spectrum β-lactamases, and the main mechanism of resistance is the production of carbapenemase, a type of β-lactamase that hydrolyzes various antibiotics, such as penicillins, cephalosporins, and monobactams.14 The 81 CRKP isolates collected in this study were mainly isolated from sputum (67.9%, 55/81), and the samples were obtained from 13 departments, indicating that CRKP is widely prevalent in hospitals in this region.A study of the resistance genotypes of the 81 CRKP isolates revealed that the main carbapenemase gene prevalent in this region was blaKPC, with no detection of blaNDM, blaIMP, or blaOXA-48, which is consistent with relevant reports in the literature.15 According to the data from the China CRE Network, KPC-type enzymes are the most common carbapenemase carried by K. pneumoniae.16
The emergence of multidrug-resistant bacteria poses a significant challenge in the clinical treatment of K. pneumoniae infections. Studying resistance mechanisms is highly important for reducing the mortality rate of multidrug-resistant bacterial infections, improving the spread of resistant bacteria, and delaying the occurrence of pandrug-resistant bacteria. This study revealed that the proportion of multidrug-resistant bacteria was as high as 83.95%, and the distribution of the resistance genes blaCTX-M, blaTEM, and aac were closely related to those of multidrug-resistant bacteria. These three types of genes are plasmid-mediated resistance genes,17 and 55.56% of the resistant bacteria carried all three genes, which may be crucial for the prevalence and transmission of multidrug-resistant bacteria in this region. Plasmid-mediated quinolone resistance (PMQR) genes are usually carried on plasmids with other antibiotic resistance genes, especially ESBL-encoding genes,18 and we speculate that the above mentioned prevalent resistance genes in this region are located on and transmitted via the same plasmid, but the specific mechanism needs further in-depth study.
The main virulence factors of K. pneumoniae include capsular polysaccharide (CPS), lipopolysaccharide (LPS), adhesins, and siderophores, and these four major virulence factors are also the main factors underlying the characteristic features of hypervirulent K. pneumoniae (hvKP).19 Many types of virulence factors associated with the virulence characteristics have been reported. In this study, 8 virulence genes, associated with mucoid phenotypes, adhesins, and siderophores were selected for amplification. The results revealed that the high mucosity of CRKP was strongly correlated with the levels of mainly rmpA2 (R = 0.474), markD (R = 0.643), and fimH (R = 0.414), which is consistent with previous research. Among these genes, rmpA regulates the expression of capsular polysaccharides and is closely related to the highly mucoid phenotype of K. pneumoniae,20,21 The gene iucA encodes the iron siderophore aerobactin, which is closely related to high virulence.22 This study revealed that the level of iucA was closely related to the expression of peq0344 (r = 0.505), rmpA2 (r = 0.557), rmpA (r = 0.482), magA (r = 0.286), and other virulence genes and was strongly correlated with the formation of biofilms (r = 0.453). In this study, the detection rates of the genes encoding type I and type III fimbriae, fimH and mrkD, were 88.89% and 65.43%, respectively. These genes are related to the formation of bacterial biofilms and have a high detection rate in Enterobacteriaceae, which is consistent with previous reports.23,24 A total of 86.42% of the CRKP isolates in this study were able to form biofilms, and type I and type III fimbriae are important factors for the colonization of K. pneumoniae. fimH encodes the FimH adhesin at the tip of type I fimbriae,25 which is closely related to urinary tract infections,26 and this study revealed that the biofilm formation ability was strongly correlated with the fimH level (r = 0.689); mrkD encodes the adhesive protein at the tip of type III fimbriae,27 which mediates the binding of K. pneumoniae with organ cells and lung tissue27 and is closely related to lung infections. Studies have shown that type III fimbriae play a major role in the formation of biofilms in vitro,28 and the biofilm formation ability in this study was also strongly positively correlated with the mrkD level (r = 0.413). Correlation analysis revealed that the rmpA2 level was strongly correlated with the magA and iutA levels (r values of 0.645 and 0.557, respectively), which may be related to these genes being located on the same pLVPK plasmid.29 Correlation analysis of the resistance and virulence characteristics of K. pneumoniae revealed that the resistance phenotype (including multidrug resistance) of the 81 CRKP isolates was negatively correlated with the distribution of virulence phenotypes and virulence genes (r < 0), which is consistent with previous reports.5
In summary, this study investigated the resistance and virulence characteristics of 81 CRKP strains isolated in Lüliang, Shanxi Province. Our data revealed that the resistance rates of CRKP in this region are and that multidrug-resistant highly mucoid CRKP strains are present in this region. Therefore, in clinical practice, the management of antibiotics should be strengthened to control the prevalence and transmission of CRKP in this region.
Data Sharing Statement
The datasets supporting the conclusions of this article are included within the article.
Ethics Approval and Consent to Participate
This research was carried out in accordance with the Declaration of Helsinki. The participants were informed of the study, including any benefits or risks involved, and voluntarily provided informed consent. The research was approved by the Ethics Committee of Fenyang College of Shanxi Medical University.
Funding
This research was supported by the Education Department Research Project of Shanxi Province (grant:2023L476).
Disclosure
The authors declare that they have no competing interests in this work.
References
1. Lin JY, Zhu ZC., Zhu JH. et al. Antibiotic heteroresistance in K. pneumoniae: Definition, detection methods, mechanisms, and combination therapy. Microbiol Res. 2024;283:127701. doi:10.1016/j.micres.2024.127701
2. Martin RM, Bachman MA. Colonization, infection, and the accessory genome of K. pneumoniae. Front Cell Infect Microbiol. 2018;8:4. doi:10.3389/fcimb.2018.00004
3. Santos SDAM, Machado SC, Neves JR. Virulence factors in k. Pneumoniae: a literature review. Indian J Microbiol. 2024;64(2):389–401. doi:10.1007/s12088-024-01247-0
4. Abu KH, Rana FA, Rania I, et al. The prevalence of carbapenem resistance gram negative pathogens in a tertiary teaching hospital in Jordan. BMC Infect Dis. 2023;23(1):634. doi:10.1186/s12879-023-08610-4
5. Hu F, Pan Y, Li H, et al. Carbapenem-resistant K. pneumoniae capsular types, antibiotic resistance and virulence factors in China: a longitudinal, multi-centre study. Nat Microbiol. 2024;9:814–829. doi:10.1038/s41564-024-01612-1
6. MacKenzie FM, Forbes KJ, Dorai-John T, Amyes SG, Gould IM. Emergence of a carbapenem-resistant K. pneumoniae. Lancet. 1997;350(9080):783. PMID: 9298003. doi:10.1016/s0140-6736(05)62567-6
7. Jun C, Dongmei Z, Xuejiao M, et al. Molecular epidemiology, risk factors, and outcomes of carbapenem-resistant K. pneumoniae infection in a tertiary hospital in eastern China: for a retrospective study conducted over 4 years. Front Microbiol. 2023;141223138.
8. Alhazmi AW. Review on the carbapenem resistance mechanisms of K. pneumoniae. J Pharm Res Int. 2021;352–357.
9. Clinical and Laboratory Standards Institute (CLSI). Performance Standards for Antimicrobial Susceptibility Testing: 30th Edition CLSI Supplement M100. PA: CLSI; 2020.
10. Magiorakos A, Srinivasan A, Carey R, et al. Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: an international expert proposal for interim standard definitions for acquired resistance. Clin Microbiol Infect. 2012;18(3):268–281. doi:10.1111/j.1469-0691.2011.03570.x
11. Fan Y, Baoguo D, Wei L, et al. High rate of multiresistant Klebsiella pneumoniae from human and animal origin. Infect Drug Resist. 2019;12272.
12. Ahmed OB, Dablool AS. Quality Improvement of the DNA extracted by boiling method in gram negative bacteria. Int J Bioassays. 2017;6(04):5347. doi:10.21746/ijbio.2017.04.004
13. Hu F, Pan Y, Li H, et al. Carbapenem-resistant Klebsiella pneumoniae capsular types, antibiotic resistance and virulence factors in China: a longitudinal, multi-centre study. Nat Microbiol. 2024;9(3):814–829. Epub 2024 Feb 29. PMID: 38424289; PMCID: PMC10914598. doi:10.1038/s41564-024-01612-1
14. Queenan MA, Bush K. Carbapenemases: the Versatile β-Lactamases. Clin Microbiol Rev. 2007;20(3):440–458. doi:10.1128/CMR.00001-07
15. Cuidan L, Xiaoyuan J, Tingting Y, et al. Genomic epidemiology of carbapenemase-producing Klebsiella pneumoniae in China. Genomic Proteomic Bioinformati. 2022;20(6):1154–1167. doi:10.1016/j.gpb.2022.02.005
16. Zhang Y, Wang Q, Yin Y, et al. Epidemiology of carbapenem-resistant Enterobacteriaceae infections: report from the China CRE network. Antimicrob Agents Chemother. 2018;62(2):e01882–17. PMID: 29203488; PMCID: PMC5786810. doi:10.1128/AAC.01882-17.
17. Mahboobeh MN, Mahdi MF, Sheila M. Prevalence and molecular characterization of plasmid-mediated extended-spectrum β-lactamase genes (balaTEM, blaCTX and blASHV) among urinary Escherichia coli clinical isolates in Mashhad, Iran. Iranian J Basic Medicalsci. 2012;15(3):833.
18. Yaxin G, Zexun L, Chenyang C, et al. Cunning plasmid fusion mediates antibiotic resistance genes represented by ESBLs encoding genes transfer in foodborne Salmonella. Inter J Food Microbiol. 2021;355109336.
19. Zhu J, Wang T, Chen L, Du H. Virulence Factors in Hypervirulent Klebsiella pneumoniae. Front Microbiol. 2021;12:642484. doi:10.3389/fmicb.2021.642484
20. Lai YC, Peng HL, HY C. RmpA2, an activator of capsule biosynthesis in K. pneumoniae CG43, regulates K2 cps gene expression at the transcriptional level. J Bacteriol. 2003;185(3):788–800. PMID: 12533454; PMCID: PMC142793. doi:10.1128/JB.185.3.788-800.2003
21. Li G, Sun S, Zhao ZY, Sun Y. The pathogenicity of rmpA or aerobactin-positive K. pneumoniae in infected mice. J Int Med Res. 2019;47(9):4344–4352. Epub 2019 Jul 21. PMID: 31328596; PMCID: PMC6753543. doi:10.1177/0300060519863544
22. Guoying W, Guo Z, Xiaoyu C, et al. The characteristic of virulence, biofilm and antibiotic resistance of K pneumoniae. Inter J Environ Res PublicHealth. 2020;17(17):6278. doi:10.3390/ijerph17176278
23. Liu C, Guo J. Hypervirulent K. pneumoniae (hypermucoviscous and aerobactin positive) infection over 6 years in the elderly in China: antimicrobial resistance patterns, molecular epidemiology and risk factor. Ann Clin Microbiol Antimicrob. 2019;18(1):4. PMID: 30665418; PMCID: PMC6341648. doi:10.1186/s12941-018-0302-9
24. Kline AK, Dodson WK, Caparon GM, et al. A tale of two pili: assembly and function of pili in bacteria. Trends Microbiol. 2010;18(5):224–232. doi:10.1016/j.tim.2010.03.002
25. Kulaç Ö, Başkan C, Koşar N. Klebsiella pneumoniae clinical isolates: extended spectrum β-lactamase production, biofilm formation, and virulence factors. Biologia. 2024;(prepublish):1–9.
26. Podschun R, Ullmann U. Klebsiella spp. as nosocomial pathogens: epidemiology, taxonomy, typing methods, and pathogenicity factors. Clin Microbiol Rev. 1998;11(4):589–603. PMID: 9767057; PMCID: PMC88898. doi:10.1128/CMR.11.4.589
27. Jagnow J, Clegg SK. pneumoniae MrkD-mediated biofilm formation on extracellular matrix- and collagen-coated surfaces. Microbiology. 2003;149(Pt 9):2397–2405. PMID: 12949165. doi:10.1099/mic.0.26434-0
28. Schroll C, Barken KB, Krogfelt KA, Struve C. Role of type 1 and type 3 fimbriae in K. pneumoniae biofilm formation. BMC Microbiol. 2010;10(1):179. PMID: 20573190; PMCID: PMC2911432. doi:10.1186/1471-2180-10-179
29. Tang H, Chiang M, Liou W, et al. Correlation between K. pneumoniae carrying pLVPK-derived loci and abscess formation. Off Public Europ Soc Clin Microbiol. 2010;29(6):689–698. doi:10.1007/s10096-010-0915-1
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms.php
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms.php
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
The Resistance and Virulence Characteristics of Salmonella Enteritidis Strain Isolated from Patients with Food Poisoning Based on the Whole-Genome Sequencing and Quantitative Proteomic Analysis
Xu B, Hou Z, Liu L, Yan R, Zhang J, Wei J, Du M, Xuan Y, Fan L, Li Z
Infection and Drug Resistance 2023, 16:6567-6586
Published Date: 6 October 2023
Molecular Characterization and Risk Factors of Carbapenem-Resistant Hypervirulent Klebsiella pneumoniae Isolated from Chinese Tertiary Hospital
Zhu R, Li J, Lian S, Li Y, Cai M, Cao Y, Xu X
Infection and Drug Resistance 2025, 18:83-92
Published Date: 6 January 2025