")
Back to Journals » Infection and Drug Resistance » Volume 18
New Directions in Helicobacter pylori Urease Inhibitors: Focusing on Nickel Ions Transfer and Auxiliary Protein Interactions During Urease Maturation
Authors Chen X, Gou L, Wang Y, Yang J, Dong Y, Xie B, Zhang D
Received 4 February 2025
Accepted for publication 29 April 2025
Published 17 June 2025 Volume 2025:18 Pages 3037—3053
DOI https://doi.org/10.2147/IDR.S519194
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Sandip Patil
Xinglan Chen, Lingzhu Gou, Yuanyuan Wang, Jinxia Yang, Yeze Dong, Baoyuan Xie, Dekui Zhang
Key Laboratory of Digestive Diseases of Gansu Province, The Second Hospital of Lanzhou University, Lanzhou, 730030, People’s Republic of China
Correspondence: Dekui Zhang, Email [email protected]
Abstract: Helicobacter pylori infection is closely linked to digestive diseases such as inflammation, ulceration, gastric cancer, and mucosa-associated lymphoid tissue lymphoma. Current treatment relies on antibiotic combinations, but antibiotic resistance increasingly undermines eradication efforts. Urease, a metalloenzyme secreted by H. pylori, is crucial for bacterial colonization. Traditional urease inhibitors target either the active site or mimic the substrate, but the buried active site poses a challenge for effective inhibition. Moreover, current inhibitors often have non-negligible side effects. Recent research highlights complex interactions during urease maturation, involving auxiliary proteins and nickel ion transfer. These studies suggest that auxiliary proteins involved in metalloenzyme maturation, as well as the process of nickel ion delivery, could be novel targets for inhibitors. This review summarizes nickel ion delivery during urease maturation, interactions between auxiliary proteins, and the structure of the active site. It also categorizes and summarizes half inhibitory concentration (IC50) values of existing inhibitors. Finally, we discuss potential inhibitors’mechanisms of action, challenges, and future perspectives, aiming to provide new strategies for eradicating H. pylori infection.
Keywords: Helicobacter pylori, urease inhibitors, urease active site, nickel ions, auxiliary proteins
Introduction
Helicobacter pylori (H. pylori), which infects approximately 50% of the world’s population., is transmitted mainly in the digestive tract, and reciprocal transmission among family members is relatively common.1 After colonization, untreated H. pylori infection persists for the lifetime of the host, rarely resolves spontaneously, most people are usually asymptomatic.2 However, in some people, persistent infection causes chronic inflammation of the gastric mucosa, which in turn leads to the development of chronic non-atrophic gastritis, atrophic gastritis, intestinal metaplasia, and dysplasia. The characteristics of H. pylori, which is transmitted by the digestive tract, and its persistent colonisation, with a high recurrence rate after eradication treatment (5–10% in adults and children in developing countries), are the main reasons for the high prevalence of H. pylori infection.3 More than 75% of duodenal ulcers and 17% of gastric ulcers are associated with H. pylori infection. H. pylori has been classified as a human carcinogen because infection with H. pylori is the strongest known risk factor for gastric cancer, which is the second leading cause of cancer-related deaths worldwide.4 There is also evidence that H. pylori is associated with extragastric disorders, including iron deficiency anaemia, idiopathic thrombocytopenic purpura, and vitamin B12 deficiency.5
The Kyoto Global Consensus Report states that there is a consensus on the necessity of treating individuals with H. pylori gastritis for eradication unless there are competing considerations.6 Currently, medications used to eradicate H. pylori include proton pump inhibitors (PPIs), bismuth, and antibiotics.7 The first-line treatment of H. pylori infections consists of the use of proton pump inhibitors associated with two antibiotics (clarithromycin plus amoxicillin or metronidazole) for a period of 1–2 weeks.8
However, the tricky part is that the antibiotic resistance rate is increasing every year. Investigations have found that H. pylori resistance rates have risen gradually since 2013 or 2014 and remain at high levels. Furthermore, a survey of trends in antibiotic resistance from 2015 to 2020 found that total resistance increased by 2.8% per year relative to the previous year, with resistance to clarithromycin, metronidazole and levofloxacin increasing by 2.7%, 2.3% and 2.4% per year, respectively.9,10 In light of this increase in global antibiotic resistance, there is a need to rethink antibiotic stewardship strategies and develop new treatment guidelines.11 Consequently, there is an urgent need to develop new methods and drugs that can effectively eradicate H. pylori and a vaccine for H. pylori.12
Urease is the most highly secreted protein of H. pylori.13 It hydrolyses urea with high efficiency, creating suitable conditions for H. pylori to colonize extremely acidic environments. At the same time, as one of the most important virulence factors of H. pylori, urease can cause local tissue damage. Therefore, targeting urease for drug design, immunotherapy, and vaccine development may lead to major breakthroughs in eradicating H. pylori infection and preventing its spread.14,15 Additionally, according to the World Health Organization (WHO), urease is prevalent among numerous antibiotic-resistant pathogens, highlighting the importance of developing effective urease inhibitors.16 Traditional urease inhibitors typically target the active site or simulate the substrate to inhibit enzyme activity. However, owing to the deeply buried active site and strong substrate specificity of bacterial urease, the development efficiency of these traditional inhibitors is low.17 In addition, most of these compounds have serious side effects, toxicity and instability. For example, the only urease inhibitor currently approved for clinical use, acetohydroxamic acid (AHA), used in the treatment of recurrent urinary catheter obstruction and bladder stones, has shown side effects such as haemolytic anaemia and teratogenicity and has been reduced in clinical use and withdrawn from the general market.18,19 Therefore, there is great promise in the development of new effective urease inhibitors for the treatment of infections associated with ammonia-producing bacteria, such as H. pylori in the digestive system and Proteus in the urinary system.20
In fact, some of the antibiotics that have been routinely used at present also have urease inhibitory effect, and their structure can be modified to become stronger urease inhibitors. For example, the hydroxamic acid and hydrazine derivatives of levofloxacin exhibit unusual urease inhibitory activities.21 In addition, two novel series of nitroimidazoles, which are synthesized for the reaction of metronidazole and salicylic acid derivatives, have been shown to inhibit H. pylori urease, and molecular docking suggested that the possible mechanism is the inhibition of the active site of the urease. However, its specific mechanism of action and biological and toxicological studies are still unclear.22
Encouragingly, several cutting-edge compounds that can inhibit the maturation and activation of urease have been identified. These compounds offer new opportunities for designing more effective urease inhibitors and identifying new targets for antibacterial drug development.23 In view of these findings, this review comprehensively analyses the synthesis and activation mechanisms of urease and summarizes the existing and potential types of inhibitors to provide a more comprehensive and in-depth reference for future research.
Biological Characteristics of H. pylori Urease
Function of Urease
Once in the stomach, H. pylori neutralizes the hostile acidic environment through the activity of the bacterially encoded urease. The bacterial cell subsequently moves towards the gastric epithelium via flagellum-mediated motility.24 H. pylori adhesins further interact with host cell receptors, leading to successful colonization and persistent infection. After successful colonization, H. pylori produces several effector proteins/toxins that damage the host gastric mucosal tissue and alter the pattern of gastric hormone release, thereby affecting gastric physiology.25–27 It is thus clear that urease plays a key role in the initial stages of successful colonization by H. pylori and is the initial step in its survival mechanism. Thus, it may be possible to eradicate H. pylori colonization at its source by effectively inhibiting urease synthesis or activity.28
As one of the most abundantly expressed proteins in H. pylori, urease is a nickel metal enzyme that can produce ammonia by hydrolysing urea to survive in a highly acidic gastric environment,29,30 and additionally promotes flagellar swimming in the mucosal layer through urease-mediated hydrolysis of urea.31 Urease can also provide nutrients for bacteria by releasing host metabolites.25 At the same time, urease is also actively involved in the production of proton power to maintain the life activities of bacteria.32
In addition, H. pylori urease exhibits a range of biological activities independent of its catalytic function. It induces an inflammatory response in vivo, promotes platelet aggregation and angiogenesis, and more directly activates human neutrophils and endothelial cells, triggering the generation of reactive oxygen species (ROS).33–35 In addition, H. pylori urease may also be neurotoxic, and its proinflammatory activity and activation of the immune system together trigger pathological processes such as neuroinflammation and tau protein phosphorylation, which are key factors in the progression of neurodegenerative diseases.36 In summary, H. pylori urease is not only crucial for the survival and growth of H. pylori in the acidic gastric environment but also one of the key molecules involved in its pathogenicity.28
Urease Active Site
The active site of urease contains two nickel ions bridged by a carbamylated lysine residue and coordinated by one aspartate and four histidine residues.37 This structure not only provides the necessary metal ion coordination environment for urease but also ensures the catalytic activity and substrate specificity of urease through precise amino acid arrangement and coordination. In addition to the active site of urease that binds directly to the substrate, the nearby active flap is crucial for the catalytic activity of urease, especially the multiple cysteine residues containing sulfhydryl groups on the active flap.38 Superposition of the structures of uninhibited H. pylori urease and AHA-inhibited H. pylori urease shows that the active flap is a helix‒flip‒helix secondary structure formed by a fragment consisting of residues α313–α346, which regulates the open and closed states of the active site via conformational changes, opening the active site when the inhibitor binds and closing it when the inhibitor is not bound. Furthermore, although the amino acid sequence of the active flap region in H. pylori urease shows a high degree of conservation in the primary and secondary helical structures, there is a significant difference in the region of specific residues preceding the active flap (especially α308 to α312). This difference is not coincidental because the active site, which is the core of the enzyme-catalysed reaction, is highly structurally conserved. It has been suggested that these nonconserved amino acid residues play a crucial role in these mobile flaps and may be a key factor in regulating the flexibility and conformational changes of the mobile flap. This difference may have led to different dynamic behaviours of the urease-catalysed active flaps of H. pylori, Klebsiella aerogenes, and Bartonella, thus affecting the catalytic properties of the respective enzymes. This is the reason why H. pylori urease has an unusually low Km value compared with other urease-producing bacteria; under the physiological conditions of the human stomach, the enzyme has evolved to function efficiently even in the presence of urea concentrations as low as 1.7–3.4 mM, owing to its extremely low Km value.39–41
Urease Generation and Activation
Safe Entry of Nickel Metal Ions into Bacterial Cells
The gene cluster encoding H. pylori urease consists of seven adjacent gene segments, UreABIHEFG.42 The core structure of urease is a ((αβ)3)4 heterodimer of the β and α subunits encoded by the structural genes UreA and UreB, known as apo-urease.43 At this point, the enzyme is inactive, and a nickel ion must be embedded in the active centre of the enzyme for it to function.29 The urease active site is located in the α-subunit and is highly conserved, and there are two binding sites for nickel ions on one subunit, so an active urease molecule requires 24 nickel ions to be fully enzymatically active.44,45
Nickel ions are free in the blood, and nickel ions in the blood enter the bacterial outer membrane through the outer membrane proteins FecA3 and FrpB4, and under the control of the protein NikR, enter the bacteria through the cytoplasmic membrane protein channel NixA (Figure 1).46,47 NixA is a high-affinity Ni2+ transporter that can independently absorb Ni2+ and transfer it to the cytoplasm during urease synthesis.48 However, nickel ions are cytotoxic, and when nickel ions enter the cell and compete with other ions to bind to enzymes, they may replace those ions that are essential for enzyme function, such as magnesium ions (Mg2+); this displacement leads to the loss of enzyme function, thus affecting the normal metabolism and function of the cell.43 Nickel ions, therefore, cannot move freely to the metal active site, but the cells have evolved an interesting mechanism to transfer this toxic metal to the enzyme active site through the auxiliary proteins encoded by the auxiliary genes (UreEFGH) in the urease gene cluster.49 Specifically, these accessory proteins undergo meticulous conformational adjustments to assemble into specific protein complexes. Simultaneously, during the process of conformational change, nickel ions are efficiently and orderly transferred between these accessory proteins, thereby ensuring that toxic metal ions do not leak into the cytoplasm. Ultimately, nickel ions are safely and accurately directed to their active sites.37
 |
Figure 1 The nickel delivery along the urease-maturation pathway. Ni2+ enters bacterial cells to form hydrogenase. It’s then transferred via HypA-UreE interaction, initiating urease maturation. Ni2+-loaded UreE interacts with UreGFH, transferring Ni2+ to the dimerized UreG. Stimulated by bicarbonate, UreG releases Ni2+ to the urease α subunit, activating it. Urea enters via UreI, gets hydrolyzed by urease into NH4+, which diffuses through UreI for neutralization in the periplasm. |
These accessory proteins are required for nickel insertion into the active centre of the enzyme, and studies have confirmed that UreF, UreG and UreH knockout mutants have a complete loss of urease activity, whereas the residual activity of UreE knockout mutants is only 1% of that of the wild type. Thus, these individual auxiliary proteins and their complex forms play irreplaceable roles in inserting nickel ions into the active centre of the enzyme and in regulating urease activity.50
Auxiliary Proteins Deliver Nickel Ions to the Urease Active Site
Upon entering the bacterium, nickel ions are first involved in the synthesis of another nickel-containing enzyme, hydrogenase, which is the key enzyme that supplies energy to H. pylori. During the maturation pathway of hydrogenase, nickel ions are delivered to the hydrogenase coenzyme HypA, which then transfers nickel ions from HypA to UreE through the formation of the HypA/UreE2 complex and into the maturation pathway of urease.51 During the formation of urease, the individual auxiliary proteins play different roles. The primary function of UreE is to facilitate the transport of nickel ions to the downstream protein UreG, with the nickel ions bound to UreE serving as the sole nickel ion supply for the synthesis of urease.52 UreE exists in solution as a homodimer, and each UreE dimer can bind only one nickel ion. Studies have demonstrated that the presence of the auxiliary protein, UreE, enhances the activity of urease.53
The central player among the many auxiliary proteins of urease is UreG, a GTPase,54 and each UreG has two specific binding motifs, a conserved metal-binding Cys-Pro-His motif and a GTPase P-loop motif, which, when replaced, disrupts urease maturation.55–57 The monomeric form of UreG lacks GTPase activity, and its activation as a GTPase is contingent upon its dimerization.58 Specifically, the presence of nickel ions plays a crucial role in dictating the conformational state of UreG. In the absence of nickel, UreG remains in its monomeric form irrespective of the guanine nucleotide concentration.57
In the presence of nickel and GDP, UreG forms dimers, although these dimers may not be as stable as those formed in the presence of nickel and GTP. In fact, UreG forms stable dimers only in the presence of both GTP and nickel ions. When GDP is the only guanine nucleotide present, it binds to a specific ligand located between the two UreG monomers. These GDP-bound UreG dimers are then recruited by the homodimer UreF2H2, which strictly interacts with UreF via UreG to form the UreGFH complex to form the UreGFH complex, and the formed UreGFH complex awaits the injection of nickel ions.37,57,59
During nickel ion delivery, nickel ions released from the UreE dimer are not directly received by the monomeric form of UreG, but are taken up by the complex UreGFH. The key step in the transfer of nickel ions from UreE to UreG relies on the simultaneous involvement of GTP and Mg2+.
Specifically, under the synergistic effect of GTP and Mg2+, the two UreG subunits inside the UreGFH complex, which were originally in a separated state, will undergo a dimerisation reaction. Immediately thereafter, this dimerised UreG will detach from the UreGFH complex and, through the interaction between the Arg 101 of UreE and the Cys66 of UreG, tightly bind to another independent UreE dimer and together form a stable UreE2G2 complex. The formation of this complex is crucial for the advancement of the whole process, as it not only significantly enhances the binding capacity of the UreG dimer to GTP but also ensures that nickel ions can be efficiently and unidirectionally transferred from UreE to the UreG dimer.60 The detached UreFH, on the other hand, forms a preactivation complex together with the urease precursor and awaits the entry of nickel ions.
On the basis of the completion of the UreE2G2 complex, bicarbonate acts as a trigger for the hydrolysis of the nickel ion-bound UreG dimer with the participation of GTP, resulting in the redissociation of the UreG dimer into a monomeric form, as well as the conversion of GTP into GDP. During this process, nickel ions are precisely released and directly localised to the preactivation complex formed by the apourease and the UreFH complex.57
Upon the nickel ions ingress into the central active site of the urease enzyme, it is noteworthy that the nickel-binding site within the urease is intricately positioned deep within the enzyme’s structure, significantly removed from the nickel-binding site of UreG. To cross this spatial barrier, a narrow tunnel is naturally formed within the UreFH/apourease complex, serving as an efficient conduit for the precise migration of nickel ions from the UreG binding site to the active site of the urease, thereby markedly accelerating the activation process of the enzyme.61 Subsequently, following the successful activation of the urease, the UreI channel protein promptly responds and opens, ensuring unimpeded and rapid permeation of urea molecules into the cellular interior.62
UreI is a distinctive inner membrane protein that significantly enhances the influx efficiency of urea molecules by forming a proton-gated urea channel.63 This protein possesses the ability to pre-bind with apourese, forming a membrane complex, which facilitates the efficient assembly of the apourease apoprotein on the membrane surface. Upon successful activation of the urease in the presence of nickel ions, the UreI channel promptly responds by rapidly opening its gateway, allowing urea to freely and rapidly enter the bacterial urease system. Within this system, urea is efficiently converted into ammonia and carbon dioxide, a process that occurs swiftly within the cytoplasm, effectively achieving rapid neutralization of pH levels.50
In summary, the transport process of nickel ions is not only dependent on the involvement of individual accessory proteins but also requires intricate interactions between multiple accessory proteins. This includes the formation of complexes such as UreGFH, UreFH, UreE2G2 and the preactivation complex. These accessory proteins are interconnected and each is essential to the overall process. Any disruption of this intricate sequence may result in failure of urease formation and activation.37
Urea Hydrolysis by Activated Urease
The UreI protein is a proton-gated urea channel that closes at a neutral pH (approximately 7.0) and opens at an acidic pH (approximately 5.0). The UreI channel consists of six subunits, each containing six transmembrane helices that form a central channel with a diameter of approximately 95 Å and a height of approximately 45 Å. There is a double-layer stopper formed by ordered lipids in the centre of the channel, which jointly enhances the stability of the channel.62 The channel has two contraction sites, which are near the bacterial cytoplasm and periplasm. Conserved aromatic and aliphatic residues play key roles at these contraction sites. The first contraction site is composed of Leu 6, Phe 84 and Trp 149. The second contraction site is composed of Leu 13, Thr 87, Tyr 88, Leu 152 and Trp 153. These sites ensure the selective passage of urea molecules by limiting the size of the channel while preventing the entry of protons and avoiding acidification within the cell.
Under acidic conditions, the periplasmic loop structures of UreI channels undergo conformational changes and open the channels. Protonated residues such as histidines and carboxylic acids on these loops are essential for sensing ambient pH and regulating channel opening and closing. When the pH is reduced, the protonated residues sense the acidic environment, triggering a conformational change in the channel that allows urea to rapidly enter the bacterial cytosol. In the cytosol, urease decomposes urea into ammonia (NH3) and carbon dioxide (CO2), which are then returned to the periplasm, neutralizing the protons entering the bacterial periplasm and thereby maintaining the periplasmic pH at approximately 6.1 under extremely acidic conditions. This adaptive mechanism enables Helicobacter pylori to survive the highly acidic gastric environment by neutralizing stomach acid through ammonia production, while effectively preventing self-toxicity caused by excessive cytoplasmic alkalization.64,65
Classification of Urease Inhibitors
Since urease is essential for the colonization of H. pylori, urease inhibitors have been researched and developed, and many natural products and synthetic or semisynthetic compounds have been studied and classified as urease inhibitors. The inhibitory activity of urease inhibitors was expressed as half inhibitory concentration (IC50). The ammonia production concentration of urease inhibitors was usually measured by Berthelot’s method and compared with that of standard urease inhibitors (AHA or thiourea). The IC50 values were calculated according to the experimental design formula. In conclusion, a smaller IC50 implies better urease inhibitor activity.66,67
This review categorizes these compounds into three primary groups based on their enzymatic inhibition mechanisms. It briefly summarizes the half-maximal inhibitory concentrations (IC50 values) of representative inhibitors belonging to the first and second categories, while providing a detailed elaboration on the third category. 1) Substrate-like inhibitors (Table 1): Structurally similar to substrates, these compounds bind to amino acid residues and/or Ni²+ in the enzyme’s active site in a mode analogous to urea (eg, thiourea and urea derivatives); 2) Mechanism-based inhibitors (Table 2): Although structurally distinct from substrates, these inhibitors directly bind to the active site or critical surrounding residues. Based on binding sites, they are further classified into: 2.1) Metal-chelating compounds (eg, hydroxamic acids), which complex with nickel ions; 2.2) Active site residue-targeting compounds (eg, metal complexes and imidazoles); 2.3) Sulfhydryl group-binding compounds (eg, quinones and phenols);68,69 3) Urease maturation inhibitors (Table 3): These inhibitors disrupt urease activity by interfering with its maturation process.8,23,70
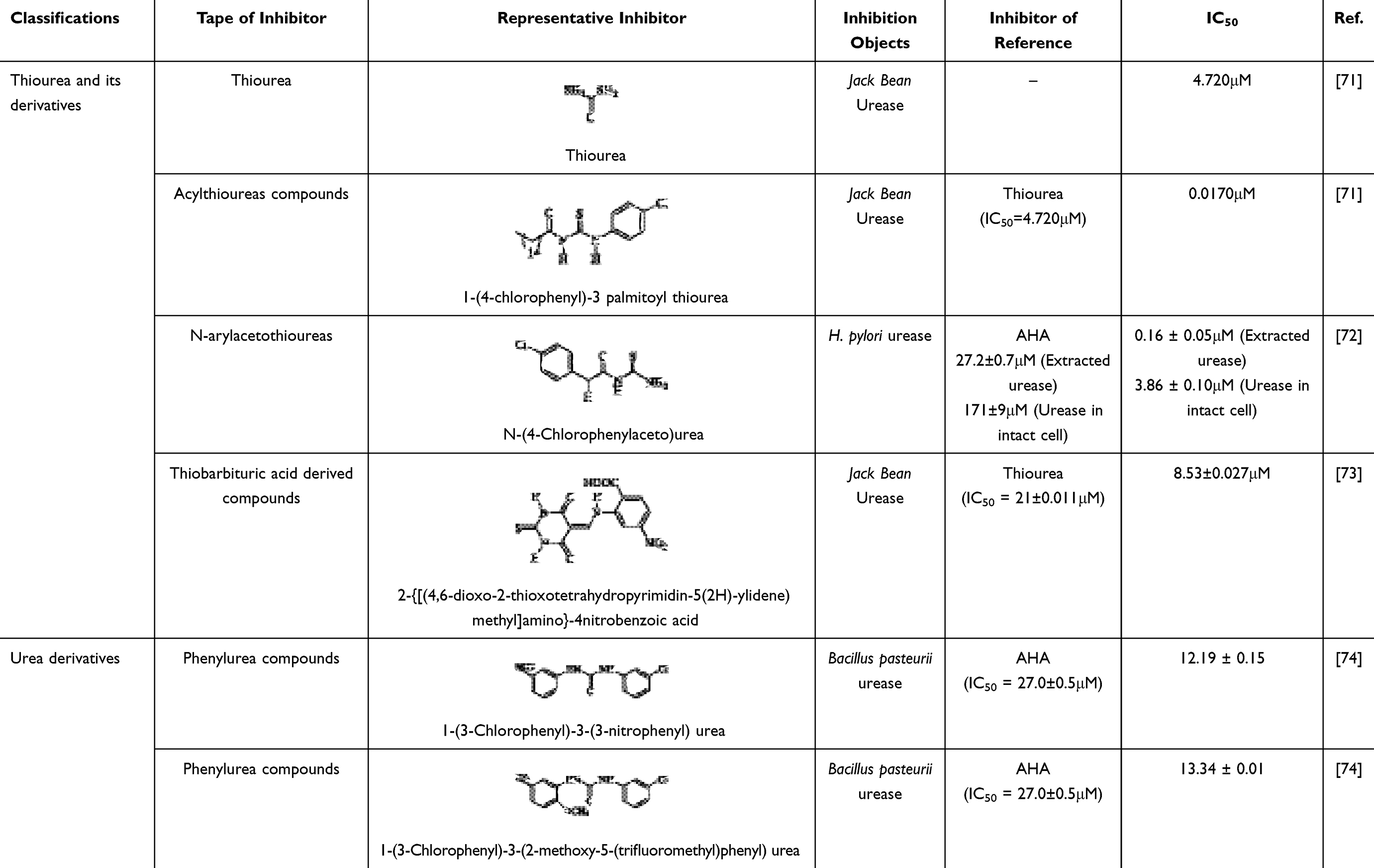 |
Table 1 Substrate Analogs Urease Inhibitors |
 |
Table 2 Mechanistic Urease Inhibitors |
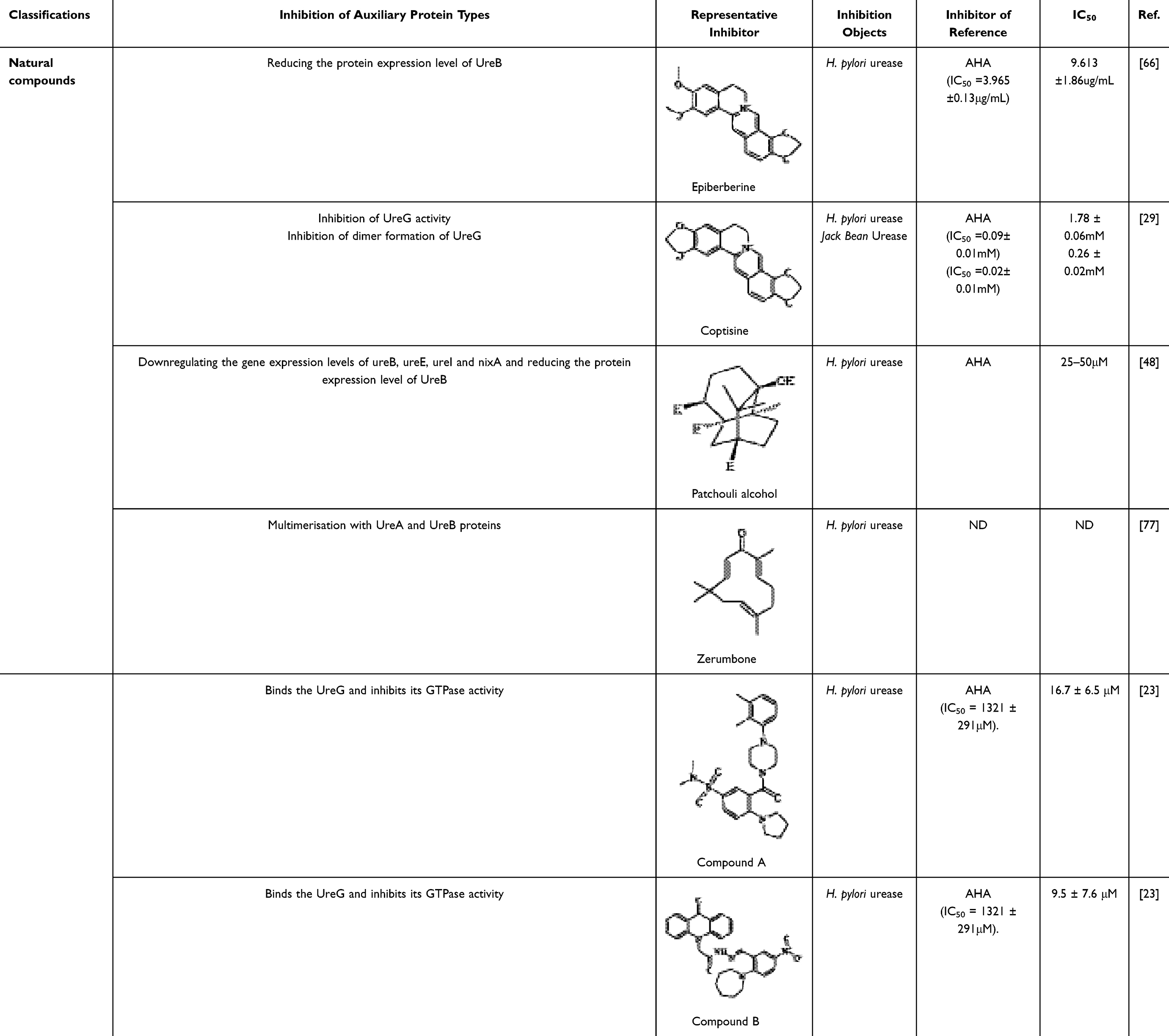 |
Table 3 Inhibitors of Urease Maturation |
Inhibitors of Urease Maturation
Most of the current inhibitors that target the urease maturation process are natural compounds, most of which come from herbs with thousands of years of experience in use. Rhizoma Coptidis (derived from the rhizome of Coptis chinensis Franch., officially recorded in the Chinese Pharmacopoeia) is one of the most commonly used herbs for the treatment of gastrointestinal disorders. Its functional components are mainly alkaloids, including berberine, palmatine, epiberberine and jatrorrhizine, some of which significantly inhibit urease activity.78 The study elucidates that following berberine treatment, the expression levels of both UreB mRNA and UreB protein exhibit a concentration-dependent downregulation. Furthermore, the results of molecular docking analysis indicate that berberine is capable of binding to the amino acid residues located at the active site of urease. This suggests that berberine not only inhibits the formation of urease but also directly targets its active site, thereby exerting a synergistic inhibitory effect.66
Meanwhile, another alkaloidal extract derived from Rhizoma coptidis, coptisine, interferes with the urease maturation process by inhibiting the activity of the urease auxiliary protein UreG and hindering the formation of UreG dimers. In addition, safranine promotes the dissociation of nickel ions from UreG dimers, thereby inhibiting urease production. More interestingly, safranine also binds to sulfhydryl groups on the active flap around the urease active centre, further synergistically inhibiting urease activity. Together, these findings reveal the multiple mechanisms of action of flavonoids in the inhibition of urease.29
Patchouli, a traditional herbal remedy in Asian countries for the treatment of gastrointestinal disorders such as gastritis and ulcers, boasts a historical pedigree spanning several centuries. Within this herbal medicine, Patchouli alcohol (PA), a tricyclic sesquiterpene compound, has demonstrated potent eradication capabilities against H. pylori in vitro, alongside significant inhibitory effects on urease activity. More specifically, PA achieves this by downregulating the expression levels of crucial genes, including UreB, ureE, UreI, and NixA (the nickel transporter protein), which subsequently reduces the abundance of the UreB protein. This mechanism effectively suppresses the expression of urease-related proteins and genes, thereby blocking the synthesis of urease.48
Zerumbone is a major constituent of the tropical zingiberaceous plant Zingiber zerumbet (also known as shampoo ginger). Rhizomes, which contain large amounts of zerumbone, were used as anti-inflammatory folk medicines in Southeast Asia as early as the early second century.79 Gingerone has a wide range of pharmacological effects, including anti-inflammatory, antibacterial and anticancer effects, and in natural systems, it can be used as a starting material for the diversification of the biosynthesis of other natural products.80 Studies have shown that gingerone has the ability to inhibit the urease activity of H. pylori. Although it does not have a direct effect on gene transcription and protein expression of UreA and UreB, gingerone is able to induce dimerisation, trimerisation or tetramerisation of UreA and UreB proteins, which in turn has an effect on the structure of deacylated urease. Since deacylated urease plays an important role in the maturation and activation of urease, this effect of gingerone indirectly affects the activity level of H. pylori urease.77
Currently, there exist numerous therapeutic regimens for the eradication of H. pylori infections in clinical practice, with bismuth-containing compounds serving as a pivotal component in most of these schemes. Research has demonstrated that colloidal bismuth citrate possesses a unique capability: it effectively disrupts the transitional UreG dimer formed in the presence of a GTPase activator element, resulting in the loss of GTPase activity of this dimer, without interfering with the stable Ni²+/GTP binding state of UreG. Crucially, colloidal bismuth citrate precisely targets UreG, inhibiting the formation of the 2E-2G complex, and thereby efficiently blocking the production of urease. Based on these findings, research suggests that UreG holds promise as an alternative novel target for the development of potent urease inhibitors.23
Building upon this theoretical foundation, researchers have screened two compounds, A and B (Table 3), from the PubChem database, which target UreG. These two compounds exhibit remarkable urease inhibitory activity, with half-maximal inhibitory concentrations (IC50) against clinically isolated strains of Helicobacter pylori as low as 16.7 ± 6.5 µM and 9.5 ± 7.6µM, respectively, providing invaluable leads for future drug development endeavors.23
Conclusion and Future Prospects
H. pylori has a large infection base in the population, and the resistance rate to conventional antibiotic treatment is increasing annually, especially for clarithromycin-resistant H. pylori strains;81 even the WHO lists clarithromycin-resistant H. pylori as a strain in urgent need of the development of a new drug,82 researchers around the world are trying to solve this problem in different ways.
Urease has a long history of research, and a variety of inhibitors have been developed and modified for its active site.83 However, the majority of previous research has predominantly focused on the active sites of existing urease enzymes. If the synthesis and activation of urease could be preemptively inhibited, the suppression of its activity would yield significantly greater efficacy with less effort.84 Some studies have demonstrated that some inhibitors of urease maturation have higher activity than AHA. Furthermore, our summary revealed that the majority of potential inhibitors capable of disrupting the urease maturation process are predominantly derived from natural sources. In recent times, plant-based natural products have garnered significant attention due to their innate origin, non-toxic or low toxicity profiles, chemical stability, and exceptional bioavailability.85 This suggests that there is unlimited potential to focus on the urease maturation process to design and develop safe and less costly urease inhibitors.
Although some natural products that inhibit urease maturation have shown good effects in inhibiting urease activity, their clinical application is still mainly for the treatment of other gastrointestinal diseases, and the treatment of Helicobacter pylori infection is not universal. The possible reasons are as follows:
Insufficient Clinical Trial Data
Although the efficacy of berberine triple therapy (consisting of berberine 500 mg, amoxicillin 1000 mg, and vonoprazan 20 mg) has been demonstrated to be comparable to that of vonoprazan quadruple therapy (vonoprazan 20 mg, amoxicillin 1000 mg, clarithromycin 500 mg, and colloidal bismuth tartrate 220 mg) and rabeprazole quadruple therapy (rabeprazole 10 mg, amoxicillin 1000 mg, clarithromycin 500 mg, and colloidal bismuth tartrate 220 mg) in some single-centre, small-sample clinical trials. Moreover, it has been observed that berberine triple therapy is well tolerated, safe and economically advantageous, while avoiding the necessity of bismuth and its associated adverse effects.86,87 However, it must be acknowledged that there is still a lack of robust large-scale multicentre clinical trials to further validate these findings.
Development of Multitarget Inhibitors
Inhibitors with a single target may face the problem of drug resistance. Therefore, future studies could explore the development of multitarget inhibitors, such as the simultaneous inhibition of the urease active centre, the active flap around the urease active centre, and the urease maturation process (eg, epiberberine, coptisine), to improve therapeutic efficacy and reduce the occurrence of drug resistance.
Drug Delivery
Urease is found both intracellularly and extracellularly, but the main function of urease is carried out intracellularly.88 Finding a means of delivering urease inhibitors precisely to the site of H. pylori colonization is an important research direction.
In conclusion, the maturation and activation of urease are extremely complex processes that require the accurate delivery of nickel ions to its active centre through the interaction of accessory proteins. Any problems with interactions are likely to affect their maturity. Each accessory protein has its own unique function. However, only some of these proteins can be inhibited, and only a few of these inhibited proteins have been identified as targets of potent urease inhibitors. In addition, few studies have investigated the necessary auxiliary nickel ion transport protein interactions to develop of urease inhibitor compounds. Therefore, the development of urease inhibitors could focus on the maturation of urease, leading to the development of more accurate, more effective and safer urease inhibitors, new strategies for the eradication of H. pylori infection, and significant progress in the field of drug development. We look forward to identifying urease inhibitors that could promote a new, more effective H. pylori eradication programme and clinical treatment of urease-related diseases, bringing new breakthroughs and hope.
Acknowledgments
The authors appreciate all individuals and supporters for their participation in this study.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This work was supported by the Innovation Driving Power Project of Gansu Association for Science and Technology (GXH20240328-25), Major projects of Joint Scientific Research Fund of Gansu Province (23JRRA1501).
Disclosure
The authors have declared that no conflict of interest exists.
References
1. Zhou XZ, Lyu NH, Zhu HY, et al. Large-scale, national, family-based epidemiological study on Helicobacter pylori infection in China: the time to change practice for related disease prevention. Gut. 2023;72(5):855–869. doi:10.1136/gutjnl-2022-328965
2. Humans IWG on the E of CR to. Infection with Helicobacter pylori. In: Schistosomes, Liver Flukes and Helicobacter Pylori. International Agency for Research on Cancer; 1994. Available from: https://www.ncbi.nlm.nih.gov/books/NBK487794/.
3. Malfertheiner P, Camargo MC, El-Omar E, et al. Helicobacter pylori infection. Nat Rev Dis Primer. 2023;9(1):1–24. doi:10.1038/s41572-023-00431-8
4. Wroblewski LE, Peek RM, Wilson KT. Helicobacter pylori and gastric cancer: factors that modulate disease risk. Clin Microbiol Rev. 2010;23(4):713–739. doi:10.1128/cmr.00011-10
5. Robinson K, Atherton JC. The spectrum of Helicobacter -mediated diseases. Annu Rev Pathol Mech Dis. 2021;16(1):123–144. doi:10.1146/annurev-pathol-032520-024949
6. Sugano K, Tack J, Kuipers EJ, et al. Kyoto global consensus report on Helicobacter pylori gastritis. Gut. 2015;64(9):1353–1367. doi:10.1136/gutjnl-2015-309252
7. Fallone CA, Chiba N, van Zanten SV, et al. The Toronto consensus for the treatment of Helicobacter pylori infection in adults. Gastroenterology. 2016;151(1):51. doi:10.1053/j.gastro.2016.04.006
8. Fiori-Duarte AT, Rodrigues RP, Kitagawa RR, et al. Insights into the design of inhibitors of the urease enzyme - a major target for the treatment of Helicobacter pylori infections. Curr Med Chem. 2020;27(23):3967–3982. doi:10.2174/0929867326666190301143549
9. Malfertheiner P, Megraud F, O’Morain CA, et al. Management of Helicobacter pylori infection—the Maastricht V/Florence consensus report. Gut. 2017;66(1):6–30. doi:10.1136/gutjnl-2016-312288
10. Shu X, Ye D, Hu C, et al. Alarming antibiotics resistance of Helicobacter pylori from children in Southeast China over 6 years. Sci Rep. 2022;12:17754. doi:10.1038/s41598-022-21661-y
11. Lee YC, Dore MP, Graham DY. Diagnosis and treatment of Helicobacter pylori infection. Annu Rev Med. 2022;73:183–195. doi:10.1146/annurev-med-042220-020814
12. Zawahir S, Czinn SJ, Nedrud JG, et al. Vaccinating against Helicobacter pylori in the developing world. Gut Microbes. 2013;4(6):568–576. doi:10.4161/gmic.27093
13. Weeks DL, Eskandari S, Scott DR, et al. A H+-gated urea channel: the link between Helicobacter pylori urease and gastric colonization. Science. 2000;287(5452):482–485. doi:10.1126/science.287.5452.482
14. Rego YF, Queiroz MP, Brito TO, et al. A review on the development of urease inhibitors as antimicrobial agents against pathogenic bacteria. J Adv Res. 2018;13:69–100. doi:10.1016/j.jare.2018.05.003
15. Kafarski P, Talma M. Recent advances in design of new urease inhibitors: a review. J Adv Res. 2018;13:101–112. doi:10.1016/j.jare.2018.01.007
16. Oliyaei N, Hashempur MH, Zomorodian K, et al. An overview of potential algae-derived bioactive compounds against urease-positive microorganisms. Algal Res. 2024;77:103372. doi:10.1016/j.algal.2023.103372
17. Zhang X, He Y, Xiong Z, et al. Chelerythrine chloride: a potential Rumen microbial urease inhibitor screened by targeting UreG. Int J Mol Sci. 2021;22(15):8212. doi:10.3390/ijms22158212
18. Hassan STS, Žemlička M. Plant-derived urease inhibitors as alternative chemotherapeutic agents. Arch Pharm. 2016;349(7):507–522. doi:10.1002/ardp.201500019
19. Heylen RA, Cusick N, White T, et al. Rational design and in vitro testing of new urease inhibitors to prevent urinary catheter blockage. RSC Med Chem. 2024;15(10):3597. doi:10.1039/d4md00378k
20. Follmer C. Ureases as a target for the treatment of gastric and urinary infections. J Clin Pathol. 2010;63(5):424–430. doi:10.1136/jcp.2009.072595
21. Abdullah MAA, Abuo-Rahma GEDAA, Abdelhafez ESMN, et al. Design, synthesis, molecular docking, anti-Proteus mirabilis and urease inhibition of new fluoroquinolone carboxylic acid derivatives. Bioorganic Chem. 2017;70:1–11. doi:10.1016/j.bioorg.2016.11.002
22. Mao WJ, Lv PC, Shi L, et al. Synthesis, molecular docking and biological evaluation of metronidazole derivatives as potent Helicobacter pylori urease inhibitors. Bioorg Med Chem. 2009;17(21):7531–7536. doi:10.1016/j.bmc.2009.09.018
23. Yang X, Koohi-Moghadam M, Wang R, et al. Metallochaperone UreG serves as a new target for design of urease inhibitor: a novel strategy for development of antimicrobials. PLoS Biol. 2018;16(1):e2003887. doi:10.1371/journal.pbio.2003887
24. Suerbaum S, Coombs N, Patel L, et al. Identification of antimotilins, novel inhibitors of Helicobacter pylori flagellar motility that inhibit stomach colonization in a mouse model. mBio. 2022;13(2):e03755–21. doi:10.1128/mbio.03755-21
25. Sharndama HC, Mba IE. Helicobacter pylori: an up-to-date overview on the virulence and pathogenesis mechanisms. Braz J Microbiol Publ Braz Soc Microbiol. 2022;53(1):33–50. doi:10.1007/s42770-021-00675-0
26. Shiota S, Suzuki R, Yamaoka Y. The significance of virulence factors in Helicobacter pylori. J Dig Dis. 2013;14(7):341–349. doi:10.1111/1751-2980.12054
27. Huang Y, Wang QL, Cheng DD, et al. Adhesion and invasion of gastric mucosa Epithelial cells by Helicobacter pylori. Front Cell Infect Microbiol. 2016;6:6. doi:10.3389/fcimb.2016.00159
28. Baj J, Forma A, Sitarz M, et al. Helicobacter pylori virulence factors—mechanisms of bacterial pathogenicity in the gastric microenvironment. Cells. 2021;10(1):27. doi:10.3390/cells10010027
29. Li C, Huang P, Wong K, et al. Coptisine-induced inhibition of helicobacter pylori: elucidation of specific mechanisms by probing urease active site and its maturation process. J Enzyme Inhib Med Chem. 2018;33(1):1362–1375. doi:10.1080/14756366.2018.1501044
30. Yang H, Huang X, Zhang X, et al. AI-2 induces urease expression through downregulation of orphan response regulator HP1021 in Helicobacter pylori. Front Med. 2022;9:790994. doi:10.3389/fmed.2022.790994
31. Bansil R, Constantino MA, Su-Arcaro C, et al. Motility of different gastric Helicobacter spp. Microorganisms. 2023;11(3):634. doi:10.3390/microorganisms11030634
32. Meyer-Rosberg K, Scott DR, Rex D, et al. The effect of environmental pH on the proton motive force of Helicobacter pylori. Gastroenterology. 1996;111(4):886–900. doi:10.1016/s0016-5085(96)70056-2
33. Scopel-Guerra A, Olivera-Severo D, Staniscuaski F, et al. The impact of Helicobacter pylori urease upon platelets and consequent contributions to inflammation. Front Microbiol. 2017;8:8. doi:10.3389/fmicb.2017.02447
34. Olivera-Severo D, Uberti AF, Marques MS, et al. A new role for Helicobacter pylori urease: contributions to angiogenesis. Front Microbiol. 2017:8. doi:10.3389/fmicb.2017.01883
35. de Jesus Souza M, de Moraes JA, Da Silva VN, et al. Helicobacter pylori urease induces pro-inflammatory effects and differentiation of human endothelial cells: cellular and molecular mechanism. Helicobacter. 2019;24(3):e12573. doi:10.1111/hel.12573
36. Uberti AF, Callai-Silva N, Grahl MVC, et al. Helicobacter pylori urease: potential contributions to alzheimer’s disease. Int J Mol Sci. 2022;23(6):3091. doi:10.3390/ijms23063091
37. Tsang KL, Wong KB. Moving nickel along the hydrogenase–urease maturation pathway. Metallomics. 2022;14(5):mfac003. doi:10.1093/mtomcs/mfac003
38. Yu XD, Zheng RB, Xie JH, et al. Biological evaluation and molecular docking of baicalin and scutellarin as Helicobacter pylori urease inhibitors. J Ethnopharmacol. 2015;162:69–78. doi:10.1016/j.jep.2014.12.041
39. Loharch S, Berlicki Ł. Rational development of bacterial ureases inhibitors. Chem Rec. 2022;22(8):e202200026. doi:10.1002/tcr.202200026
40. Ha NC, Oh ST, Sung JY, et al. Supramolecular assembly and acid resistance of Helicobacter pylori urease. Nat Struct Biol. 2001;8(6):505–509. doi:10.1038/88563
41. Amtul Z, Siddiqui RA, Choudhary MI, et al. Chemistry and mechanism of urease inhibition. Curr Med Chem. 2002;9(14):1323–1348. doi:10.2174/0929867023369853
42. Mobley HLT. Urease. In: Mobley HL, Mendz GL, Hazell SL, editors. Helicobacter Pylori: Physiology and Genetics. ASM Press; 2001. Available from: http://www.ncbi.nlm.nih.gov/books/NBK2417/.
43. Farrugia MA, Macomber L, Hausinger RP. Biosynthesis of the urease metallocenter *. J Biol Chem. 2013;288(19):13178–13185. doi:10.1074/jbc.R112.446526
44. Nim YS, Wong KB. The maturation pathway of nickel urease. Inorganics. 2019;7(7):85. doi:10.3390/inorganics7070085
45. De Reuse H, Vinella D, Cavazza C. Common themes and unique proteins for the uptake and trafficking of nickel, a metal essential for the virulence of Helicobacter pylori. Front Cell Infect Microbiol. 2013;3:72334. doi:10.3389/fcimb.2013.00094
46. Ernst FD, Stoof J, Horrevoets WM, et al. NikR mediates nickel-responsive transcriptional repression of the Helicobacter pylori outer membrane proteins FecA3 (HP1400) and FrpB4 (HP1512). Infect Immun. 2006;74(12):6821–6828. doi:10.1128/iai.01196-06
47. Camporesi G, Minzoni A, Morasso L, et al. Nickel import and export in the human pathogen Helicobacter pylori, perspectives from molecular modelling. Metallomics. 2021;13(12):mfab066. doi:10.1093/mtomcs/mfab066
48. Lian D, Xu Y, Deng Q, et al. Effect of patchouli alcohol on macrophage mediated Helicobacter pylori digestion based on intracellular urease inhibition. Phytomedicine. 2019;65:153097. doi:10.1016/j.phymed.2019.153097
49. Alfano M, Cavazza C. Structure, function, and biosynthesis of nickel-dependent enzymes. Protein Sci. 2020;29(5):1071–1089. doi:10.1002/pro.3836
50. Voland P, Weeks DL, Marcus EA, et al. Interactions among the seven Helicobacter pylori proteins encoded by the urease gene cluster. Am J Physiol-Gastrointest Liver Physiol. 2003;284(1):G96–G106. doi:10.1152/ajpgi.00160.2002
51. Xinming Y, Hongyan L, Tianfan C, et al. Nickel translocation between metallochaperones HypA and UreE in Helicobacter pylori. Met Integr Biometal Sci. 2014;6(9). doi:10.1039/c4mt00134f
52. Yuen MH, Fong YH, Nim YS, et al. Structural insights into how GTP-dependent conformational changes in a metallochaperone UreG facilitate urease maturation. Proc Natl Acad Sci. 2017;114(51):E10890–E10898. doi:10.1073/pnas.1712658114
53. Benoit S, Maier RJ. Dependence of Helicobacter pylori Urease activity on the nickel-sequestering ability of the UreE accessory protein. J Bacteriol. 2003;185(16):4787–4795. doi:10.1128/jb.185.16.4787-4795.2003
54. Pierro A, Etienne E, Gerbaud G, et al. Nickel and GTP modulate Helicobacter pylori UreG structural flexibility. Biomolecules. 2020;10(7):1062. doi:10.3390/biom10071062
55. Nalini M, Ste ́phane B, Robert JM. Roles of conserved nucleotide-binding domains in accessory proteins, HypB and UreG, in the maturation of nickel-enzymes required for efficient Helicobacter pylori colonization. Microb Pathog. 2003;35(5). doi:10.1016/s0882-4010(03)00151-7
56. Boer JL, Quiroz-Valenzuela S, Anderson KL, et al. Mutagenesis of Klebsiella aerogenes UreG to probe nickel binding and interactions with other urease-related proteins. Biochemistry. 2010;49(28):5859–5869. doi:10.1021/bi1004987
57. Fong YH, Wong HC, Yuen MH, et al. Structure of UreG/UreF/UreH complex reveals how urease accessory proteins facilitate maturation of Helicobacter pylori urease. PLOS Biol. 2013;11(10):e1001678. doi:10.1371/journal.pbio.1001678
58. Raphael G, Simon M, Katja G, et al. It takes two to tango: regulation of G proteins by dimerization. Nat Rev Mol Cell Biol. 2009;10(6). doi:10.1038/nrm2689
59. Hang Fong Y, Wong HC, Chuck CP, et al. Assembly of preactivation complex for urease maturation in Helicobacter pylori: CRYSTAL STRUCTURE OF UreF-UreH PROTEIN COMPLEX. J Biol Chem. 2011;286(50):43241–43249. doi:10.1074/jbc.M111.296830
60. Yang X, Li H, Lai TP, et al. UreE-UreG complex facilitates nickel transfer and preactivates GTPase of UreG in Helicobacter pylori*♦. J Biol Chem. 2015;290(20):12474–12485. doi:10.1074/jbc.M114.632364
61. Nim YS, Fong IYH, Deme J, et al. Delivering a toxic metal to the active site of urease. Sci Adv. 2023;9(16):eadf7790. doi:10.1126/sciadv.adf7790
62. Strugatsky D, McNulty R, Munson K, et al. Structure of the proton-gated urea channel from the gastric pathogen Helicobacter pylori. Nature. 2013;493(7431):255–258. doi:10.1038/nature11684
63. Sachs G, Kraut JA, Wen Y, et al. Urea transport in bacteria: acid acclimation by gastric Helicobacter spp. J Membr Biol. 2006;212(2):71–82. doi:10.1007/s00232-006-0867-7
64. Cáceres-Delpiano J, Teneb J, Mansilla R, et al. Variations in periplasmic loop interactions determine the pH-dependent activity of the hexameric urea transporter UreI from Helicobacter pylori: a molecular dynamics study. BMC Struct Biol. 2015;15(1):1–12. doi:10.1186/s12900-015-0038-0
65. McNulty R, Ulmschneider JP, Luecke H, et al. Mechanisms of molecular transport through the urea channel of Helicobacter pylori. Nat Commun. 2013;4(1):2900. doi:10.1038/ncomms3900
66. Wu H, Xie X, Tang Q, et al. Epiberberine inhibits Helicobacter pylori and reduces host apoptosis and inflammatory damage by down-regulating urease expression. J Ethnopharmacol. 2024;318:117046. doi:10.1016/j.jep.2023.117046
67. Montazer MN, Asadi M, Moradkhani F, et al. Design, synthesis, and biological studies of the new cysteine-N-arylacetamide derivatives as a potent urease inhibitor. Naunyn Schmiedebergs Arch Pharmacol. 2024;397(1):305–315. doi:10.1007/s00210-023-02596-1
68. Mazzei L, Cianci M, Musiani F, et al. Inactivation of urease by 1,4-benzoquinone: chemistry at the protein surface. Dalton Trans. 2016;45(13):5455–5459. doi:10.1039/C6DT00652C
69. Mazzei L, Cianci M, Musiani F, et al. Inactivation of urease by catechol: kinetics and structure. J Inorg Biochem. 2017;166:182–189. doi:10.1016/j.jinorgbio.2016.11.016
70. Upadhyay LSB. Urease inhibitors: a review. Indian J Biotechnol. 2012;11:381–388.
71. Saeed A, ur-Rehman S, Channar PA, et al. Jack bean urease inhibitors, and antioxidant activity based on palmitic acid derived 1-acyl-3- arylthioureas: synthesis, kinetic mechanism and molecular docking studies. Drug Res. 2017;67(10):596–605. doi:10.1055/s-0043-113832
72. Li WY, Ni WW, Ye YX, et al. N-monoarylacetothioureas as potent urease inhibitors: synthesis, SAR, and biological evaluation. J Enzyme Inhib Med Chem. 2020;35(1):404–413. doi:10.1080/14756366.2019.1706503
73. Rauf A, Shahzad S, Bajda M, et al. Design and synthesis of new barbituric- and thiobarbituric acid derivatives as potent urease inhibitors: structure activity relationship and molecular modeling studies. Bioorg Med Chem. 2015;23(17):6049–6058. doi:10.1016/j.bmc.2015.05.038
74. Bano B, Khan KM, Lodhi A, et al. Synthesis, in vitro urease inhibitory activity, and molecular docking studies of thiourea and urea derivatives. Bioorganic Chem. 2018;80:129–144. doi:10.1016/j.bioorg.2018.06.007
75. Mazzei L, Cianci M, Ciurli S. Inhibition of urease by hydroquinones: a structural and kinetic study. Chem Weinh Bergstr Ger. 2022;28(64):e202201770. doi:10.1002/chem.202201770
76. Zhou JT, Li CL, Tan LH, et al. Inhibition of Helicobacter pylori and its associated urease by palmatine: investigation on the potential mechanism. PLoS One. 2017;12(1):e0168944. doi:10.1371/journal.pone.0168944
77. Woo HJ, Yang JY, Lee P, et al. Zerumbone inhibits Helicobacter pylori urease activity. Molecules. 2021;26(9):2663. doi:10.3390/molecules26092663
78. Meng FC, Wu ZF, Yin ZQ, et al. Coptidis rhizoma and its main bioactive components: recent advances in chemical investigation, quality evaluation and pharmacological activity. Chin Med. 2018;13(1):13. doi:10.1186/s13020-018-0171-3
79. Kalantari K, Moniri M, Boroumand Moghaddam A, et al. A review of the biomedical applications of Zerumbone and the techniques for its extraction from Ginger Rhizomes. Molecules. 2017;22(10):1645. doi:10.3390/molecules22101645
80. Biji M, Prabha B, Lankalapalli RS, et al. Transition metal/Lewis Acid catalyzed reactions of Zerumbone for diverse molecular Motifs. Chem Rec. 2021;21(12):3943–3953. doi:10.1002/tcr.202100206
81. Phan TN, Tran VH, Tran TNH, et al. Antimicrobial resistance in Helicobacter pylori: current situation and management strategy in Vietnam. J Infect Dev Ctries. 2015;9(6):609–613. doi:10.3855/jidc.6942
82. Tacconelli E, Carrara E, Savoldi A, et al. Discovery, research, and development of new antibiotics: the WHO priority list of antibiotic-resistant bacteria and tuberculosis. Lancet Infect Dis. 2018;18(3):318–327. doi:10.1016/S1473-3099(17)30753-3
83. Kosikowska P, Berlicki Ł. Urease inhibitors as potential drugs for gastric and urinary tract infections: a patent review. Expert Opin Ther Pat. 2011;21:945–957. doi:10.1517/13543776.2011.574615
84. Zhang X, Xiong Z, He Y, et al. Epiberberine: a potential rumen microbial urease inhibitor to reduce ammonia release screened by targeting UreG. Appl Microbiol Biotechnol. 2024;108(1):289. doi:10.1007/s00253-024-13131-4
85. Modolo LV, de Souza AX, Horta LP, et al. An overview on the potential of natural products as ureases inhibitors: a review. J Adv Res. 2015;6(1):35–44. doi:10.1016/j.jare.2014.09.001
86. Chen S, Shen W, Liu Y, et al. Efficacy and safety of triple therapy containing berberine, amoxicillin, and vonoprazan for Helicobacter pylori initial treatment: a randomized controlled trial. Chin Med J. 2023;136(14):1690–1698. doi:10.1097/CM9.0000000000002696
87. Chen XX, Chen YX, Bi HX, et al. Efficacy and safety of triple therapy containing berberine hydrochloride, amoxicillin, and rabeprazole in the eradication of Helicobacter pylori. J Dig Dis. 2022;23(10):568–576. doi:10.1111/1751-2980.13146
88. Scott DR, Weeks D, Hong C, et al. The role of internal urease in acid resistance of Helicobacter pylori. Gastroenterology. 1998;114(1):58–70. doi:10.1016/S0016-5085(98)70633-X
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms.php
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms.php
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.