")
Back to Journals » Research and Reports in Tropical Medicine » Volume 16
Novel Drug Targets for the Bradyzoite Form of Toxoplasma gondii
Authors Ortiz JO, Potter AK, Benmerzouga I
Received 11 February 2025
Accepted for publication 25 March 2025
Published 29 March 2025 Volume 2025:16 Pages 25—30
DOI https://doi.org/10.2147/RRTM.S431290
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Mario A. Rodríguez-Pérez
Justin Orlando Ortiz, Anna K Potter, Imaan Benmerzouga
Kiran C. Patel College of Osteopathic Medicine, Department of Foundational Sciences, Clearwater, FL, USA
Correspondence: Imaan Benmerzouga, Email [email protected]
Abstract: Toxoplasma gondii is a world-wide parasite, with an estimated prevalence of approximately 30%. Toxoplasmosis is a severe disease in the immunocompromised, but few symptoms are exhibited by patients with an intact immune system, making this parasite a worldwide burden. Currently, few drugs exist in treating acute toxoplasmosis and no drugs exist to eliminate the bradyzoite of T. gondii. Effective therapies against acute and chronic toxoplasmosis are urgently needed to reduce the burden of this disease. This review aims to give a summary of recent findings in the bradyzoite form of Toxoplasma gondii and the implication of these findings on drug development. A thorough search of PubMed and Google Scholar databases was used to identify studies within the past 10 years that illustrate targetable key elements in the differentiation and formation of the bradyzoite form of T. gondii.
Keywords: chronic toxoplasmosis, bradyzoite, toxoplasmosis, anti-toxoplasma agents, Toxoplasma gondii
Introduction
Toxoplasma gondii is a single-celled obligatory intracellular parasite of the Apicomplexa phylum and has infected up to 30% of the world population. However, this estimate varies and is dependent on the region, for instance, prevalence can be as high as 90% in Europe, Central America and South America.1 T. gondii is a uniquely opportunistic parasite that can infect any warm-blooded mammals, including humans. Within these intermediate hosts is where T. gondii undergoes its asexual life cycle, where the parasite alternates between two distinct forms, a rapidly growing disease-causing form called tachyzoite and a slow, dormmate, cyst creating form called bradyzoite. T. gondii has only one definitive host – all feline species, where the parasite will undergo sexual reproduction. T. gondii can be contracted through contaminated food or water from feline feces, undercooked meat containing cysts from animals and from infected mothers to the fetus through the placenta.2 Within the mammalian hosts, the asexual reproductive tachyzoite form is easily cleared by the host's immune system. However, upon detection of an immune response, the parasite converts to the dormmate bradyzoite form, which is protected within a cyst wall and converts to a chronic infection.3 In healthy individuals, T. gondii infection symptoms are similar to the flu, however in immunocompromised patients, the bradyzoite form can convert back to the rapidly proliferating, disease causing tachyzoite form triggering the sequelae of acute toxoplasmosis in the brain.3
In the United States, T. gondii infections leading to toxoplasmosis, rank second as the most common cause of death related to foodborne illness.4 The current treatments include combination therapies of pyrimethamine and sulfonamide antibiotics, which target the tachyzoite form. However, increases in price per tablet and clinically significant side effects have limited effective treatment of toxoplasmosis. The adverse events of these front-line therapies are collected by the FDA adverse events reporting system (FAERS), and a recent evaluation of these reports showed that anti-toxoplasma drugs were listed as the primary suspect drug leading to several adverse outcomes including death.5 Of note, these therapies are relatively ineffective against the cyst encapsulated bradyzoite form. The development of novel drugs to treat chronic T. gondii infection is imperative. Here, we review important findings made in the last 10 years that are promising treatments for the cyst form of T. gondii, bradyzoite.
The Life Cycle of Toxoplasma gondii
T. gondii is able to proliferate through sexual and asexual replication mechanisms. During asexual reproduction, tachyzoite parasites transverses the intermediate host cells using a gliding motility system to arrive at the plasma membrane. This complex mechanism at the apical end of the parasite requires multiple proteins anchored to an actin myosin motor network. This network involves the parasites endomembrane system comprising micronemes and other proteins that recognize and attach to the host cell’s membrane Following invasion of a tachyzoite, the parasite forms a protective parasitophorous vacuole (PV) derived from the host plasma membrane, creating a physical separation of the parasite from the host that will protect T. gondii from immune responses, lysosomal fusion, and autophagy.6 Once in the PV, the tachyzoites actively divide through a process called endodyogeny—where daughter cells are formed within the mother cell. This process allows for the parasites to exponentially duplicate within hours. After multiple replications, the parasite will egress via rupture of the PV and host cell plasma membrane to liberate hundreds of tachyzoites. These tachyzoites can then infect other nearby host cells. Gradually, in response to the host cells immune system and limited essential nutrient supply, the tachyzoites will slow their replication rate to convert into the cyst encapsulated bradyzoite form in skeletal muscle, cardiac muscle and the brain.7 Cyst encapsulated bradyzoites can remain for years within the host and lie dormant until an opportunity arises to prompt reactivation.8 Additionally, among T. gondii exist different strain types, type I, II, and III, with each displaying differences in virulence and ability to form cysts. Type I are rapidly growing tachyzoites resulting in their high virulence. Type II and III readily form cysts and are responsible for chronic infection.9 Within the feline species, T. gondii will undergo sexual reproduction. The cycle begins once a feline consumes tissue cysts. Then, within the feline's gut, proteolytic enzymes from the stomach and small intestines degrade the cyst wall membrane that contains many bradyzoites. This will release the parasites and allow them to infect the intestinal epithelium of the feline, resulting in the start of sexual reproduction. After a series of morphological changes leading to the formation of macro and microgametes, which fuse to form the diploid oocyst, a thick impermeable wall is formed around the oocyte and is then eliminated with the cat feces. The oocysts are then exposed to ambient air and temperature, allowing sporulation and maturation stabilizing the oocysts for 18 months.8,10 Experimental evidence suggests that sexual reproduction occurs within the feline species because of the accumulation of linoleic acid resulting from the absence of the metabolizing enzyme Δ-6-desaturase in felines, as the inhibition of this enzyme in mice permitted sexual reproduction of T. gondii and the formation of infectious oocysts.10
Toxoplasmosis and Current Treatments
In the United States, only pyrimethamine in combination with sulfonamide antibiotics is approved for the treatment of toxoplasmosis as first-line therapy.11 Pyrimethamine is a dihydrofolate reductase (DHFR) inhibitor that decreases the levels of folic acid and will inhibit purine and pyrimidine synthesis. Pyrimethamine is given with sulfadiazine, to synergistically inhibit DNA synthesis and is supplemented with folinic acid to inhibit the myelosuppression side effects, which results from the lack of significant specificity of the current approved DHFR inhibitors. Additionally, pyrimethamine poses potential teratogenic effects limiting its use during pregnancy. Although effective, pyrimethamine is associated with potentially severe side effects including thrombocytopenia, hematologic neutropenia and Steven-Johnson syndrome.12,13 In addition to pyrimethamine-based regimens, trimethoprim-sulfamethoxazole (TMP-SMX), dapsone, atovaquone, azinomycin and clindamycin are used for various clinical manifestations of toxoplasmosis. Nevertheless, all available treatments are ineffective in eliminating the bradyzoite form of T. gondii. The bradyzoite is a long-lived form that also places individuals at risk of repeated reactivation. Very few early-stage results promise effective targeting of chronic toxoplasmosis including: 1) endochin-like quinolone-based compounds targeting the parasites oxidative phosphorylation and energy production, 2)Toxoplasma cathepsin-L (TgCPL), targeting an essential cysteine protease for parasite survival in the bradyzoite form and the 3) calcium-dependent protein kinase 1 (TgCDPK1), targeting a serine/threonine kinase that is essential for the parasite life cycle.11
New Promising Targeting of Bradyzoites
The mechanism of conversion from a tachyzoite to a bradyzoite was recently shown to be controlled by a master protein known as Bradyzoite Formation Deficient (TgBFD1), and most recently TgBFD2. TgBFD1 or TbBFD2 are indispensable for differentiation and cyst formation in mice. Additionally, TgBFD1 was tested as a vaccine candidate and demonstrated immune responses with the potential for protection. In mice, however, the protection was significant against type II strain and not type I strain. The translational upregulation of TgBFD1 and TgBFD2 during T. gondii differentiation is regulated by the T. gondii translation initiation factor TgeIF1.2. However, the deletion of the network required for differentiation, including TgBFD1 did not prevent recrudescent disease. These results collectively support the development of targeted therapies regulating the stage-specific TgBFD1, which when combined with additional cytocidal therapies as a promising avenue for targeting the bradyzoite form of T. gondii.14–18
Another important feature of bradyzoites is the accumulation of amylopectin granules (AGs). AGs are unique at multiple levels, from their residence in the cytoplasm to their hybrid oligosaccharide metabolism, with a plant-like kinase activity, an alpha-glucan water dikinase TgGWD and an animal-like phosphatase activity, the glucan phosphatase TgLaforin. These granules are thought to be the site of energy storage that is necessary for the bradyzoite persistence and reactivation. Further characterization of TgLaforin depicted its important role in parasite growth and survival during the asexual life cycle. Specifically, the knockout of TgLaforin from tachyzoite resulted in attenuated virulence, reduced cyst burden and a milder illness when defective cysts were introduced to mice. The independent deletion of TgGWD and the deletion of the enzyme responsible for amylopectin synthesis, a UDP-sugar pyrophosphorylase gUSP, resulted in reduced replication of tachyzoites and reduced virulence in mice, suggesting a role for AGs in tachyzoite fitness. Additionally, inhibitors against TgLaforin have identified lead compounds that are specific to its catalytic activity. Therefore, combining the inhibition of a bradyzoite-specific phenomenon (AG accumulation) along with cytocidal therapies, is an additional attractive therapeutic approach against T. gondii.19–21
The cyst wall is characteristic of the bradyzoite stage of T. gondii and is associated with chronic infection. Natural products and plant extracts have become a potential alternative and are safe options for therapies as current treatment options are limited and risk harmful side effects. A recent study has shown that ginger extract can significantly reduce cyst counts in the brains of chronically infected mice comparable to standard treatments. In addition, the study showed that ginger extract reduced infection-induced edema and inflammation and protection against necrosis or apoptosis associated with chronic infections. Piceatamol is another natural plant product found in grapes that showed promising anti-toxoplasma activity in culture and in mice. A screening of fungal natural products identified 49 fungal metabolites with promising anti-toxoplasma activity and favorable pharmacological profiles that are yet to be tested in mice.22–24
Proteomic analysis of purified cyst wall identified important cyst wall (CST) proteins involved in T. gondii virulence, as deletion of TgCST1 or TgCST2 from parasites resulted in reduced cyst burden in mice. Transcriptome analysis of the bradyzoite identified one hundred and two putative bradyzoite secreted effectors that mostly remain uncharacterized. Several cyst wall proteins were characterized, and no significant phenotype was observed from knockout studies, except for TgMCP3, which was shown to be important for cyst size in mice but not cyst number or survival rates in mice. Another protein, achitinase-like protein 1 TgCLP1 was found to be important for the conversion from bradyzoite to tachyzoite but not from tachyzoite to bradyzoite, the former being a critical process for reactivation. While TgCLP1 is not a promising drug target for acute toxoplasmosis, a greater understanding of its role in reactivated disease is an attractive therapeutic intervention to prevent reactivated disease.25–27
T. gondii invasion requires the formation of a molecular bridge between the host cell membrane and the parasite known as the moving junction, a complex created by the assembly of apical membrane antigen proteins (AMA) secreted by the microneme organelle, and rhoptry neck partners (RONs), secreted by the rhoptry organelle. Recently, it was shown that bradyzoites invade host cells using the moving junction created by AMA2 and AMA4, of which are not detectable at the moving junction of tachyzoites. AMA4 is vital for cyst burden during the chronic infection stage but dispensable for the acute stage. Immunization challenge of mice with AMA4-RON2L1 complex protected the mice from chronic toxoplasmosis. This study indicated that AMA-RON complexes are potential candidates for vaccine development for chronic toxoplasmosis.28 Other molecular targets that have been shown to be effective against both tachyzoite and bradyzoite include the class of bicyclic azetidines that target phenylalanine tRNA synthetase TgPheRS. The lead compound halted the growth of tachyzoites, other apicomplexan parasites and appropriate bradyzoite formation and viability of ex-vivo cysts, prolonged the survival of mice infected with Toxoplasma and protected immunocompromised mice from toxoplasmosis.29 Additionally, the recent characterization of the T. gondii isolate Tg68 reveals important in-vitro characteristics that will allow a better understanding of bradyzoite biology aiding the discovery of druggable targets30 (Figure 1 and Table 1).
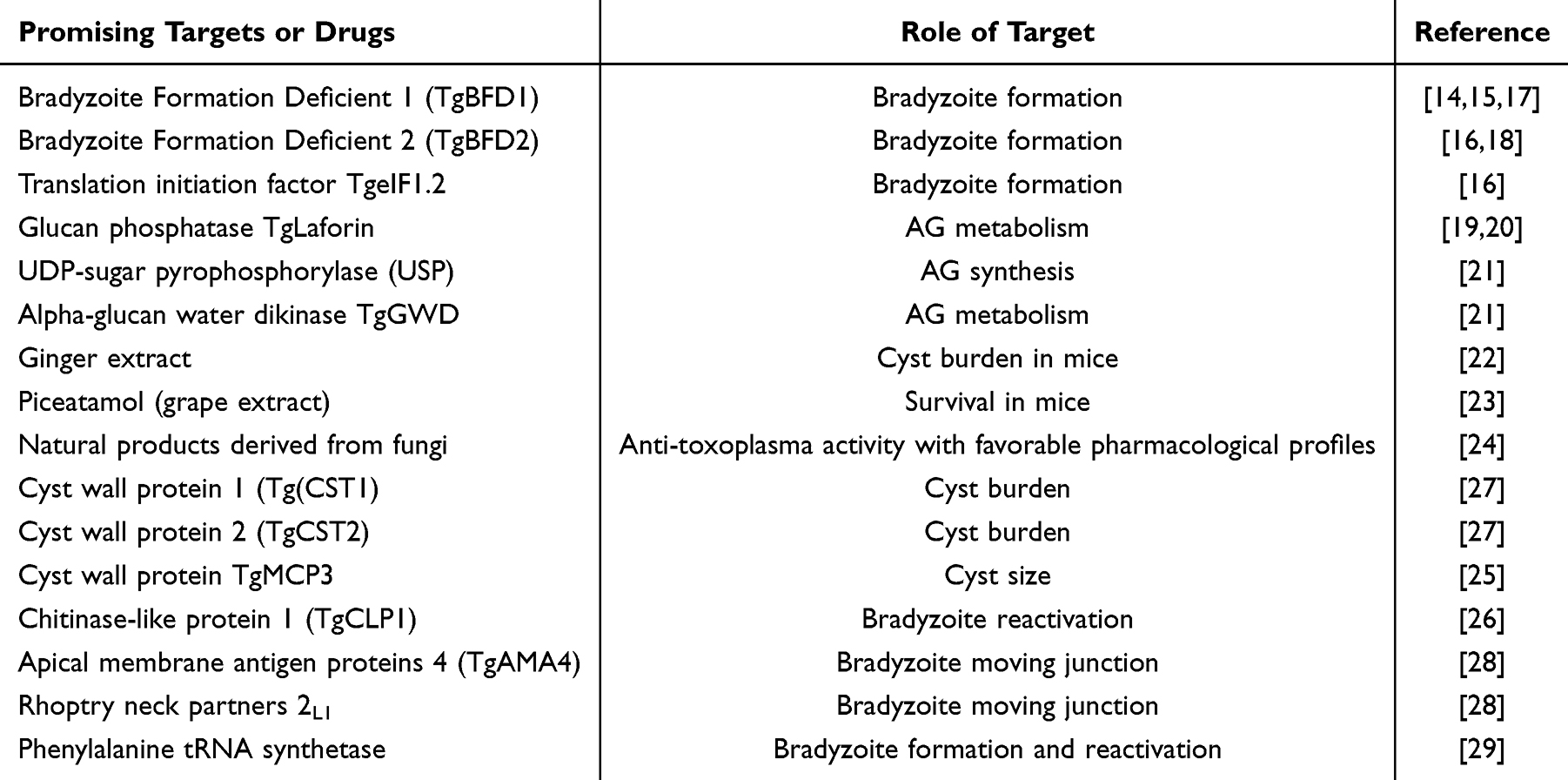 |
Table 1 Summary of Promising Targets and Drugs Against Bradyzoites |
 |
Figure 1 Promising druggable elements of bradyzoite biology. |
Conclusion
Toxoplasma gondii is a successful opportunistic parasite, which utilizes vehicles such as humans to reach its definitive host, the feline species. To achieve this, T. gondii developed effective means for this propagation. Some of these mechanisms are well understood, but others remain to be fully elucidated. This review focused on summarizing new findings in the past 10 years that are unique to T. gondii bradyzoite biology and thus making them attractive druggable targets or vaccine candidates. The intermediate host T. gondii takes shape of two forms, the fast-replicating tachyzoite and the dormant slow-replicating bradyzoite form. There are many unique aspects to T. gondii biology, and effective therapeutic targets have yet to emerge that could be as effective as the current first-line treatments that come with unfavorable side effects. By exploring the intricate mechanisms underlying the parasite’s survival and replication within the host, key molecular components have been identified that play crucial roles in the parasite’s life cycle. These components present promising opportunities for targeted drug interventions.
The chronic stage of T. gondii poses unique challenges due to the parasite’s ability to establish a latent infection within host tissues. Understanding the molecular mechanisms employed by the parasite during this stage has identified targets involved in the formation and maintenance of tissue cysts, reactivation of bradyzoite and successful invasion. By selectively targeting these pathways, it may be possible to eliminate dormant parasites and prevent the reactivation of the infection, thereby providing long-term therapeutic benefits for patients with chronic toxoplasmosis that are at risk of reactivated disease and potential consequences associated with dormancy in the brain.
Acknowledgments
We would like to acknowledge the Kiran C. Patel College of Osteopathic Medicine for providing resources to support this completion of this project.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Disclosure
The authors declare no conflicts of interest in this work.
References
1. Aguirre AA, Longcore T, Barbieri M, et al. The one health approach to Toxoplasmosis: epidemiology, control, and prevention strategies. Ecohealth. 2019;16(2):378–390. doi:10.1007/s10393-019-01405-7
2. Almeria S, Dubey JP. Foodborne transmission of Toxoplasma gondii infection in the last decade. An Overview Res Vet Sci. 2021;135:371–385. doi:10.1016/j.rvsc.2020.10.019
3. Basavaraju A. Toxoplasmosis in HIV infection: an overview. Trop Parasitol. 2016;6(2):129–135. doi:10.4103/2229-5070.190817
4. Furtado JM, Smith J, Belfort R, et al. Toxoplasmosis: a global threat. J Glob Infect Dis. 2011;3(3):281–284. doi:10.4103/0974-777X.83536
5. Shammaa AM, Powell TG, Benmerzouga I. Adverse outcomes associated with the treatment of Toxoplasma infections. Sci Rep. 2021;11(1):1035. doi:10.1038/s41598-020-80569-7
6. Clough B, Frickel EM. The Toxoplasma parasitophorous vacuole: an evolving host-parasite frontier. Trends Parasitol. 2017;33(6):473–488. doi:10.1016/j.pt.2017.02.007
7. Jeffers V, Tampaki Z, Kim K, et al. A latent ability to persist: differentiation in Toxoplasma gondii. Cell mol Life Sci. 2018;75(13):2355–2373. doi:10.1007/s00018-018-2808-x
8. Attias M, Teixeira DE, Benchimol M, et al. The life-cycle of Toxoplasma gondii reviewed using animations. Parasit Vectors. 2020;13(1):588. doi:10.1186/s13071-020-04445-z
9. Zhao XY, Ewald SE. The molecular biology and immune control of chronic Toxoplasma gondii infection. J Clin Invest. 2020;130(7):3370–3380. doi:10.1172/JCI136226
10. Martorelli Di Genova B, Wilson SK, Dubey JP, et al. Intestinal delta-6-desaturase activity determines host range for Toxoplasma sexual reproduction. PLoS Biol. 2019;17(8):e3000364. doi:10.1371/journal.pbio.3000364
11. Lapinskas PJ, Ben-Harari RR. Perspective on current and emerging drugs in the treatment of acute and chronic toxoplasmosis. Postgrad Med. 2019;131(8):589–596. doi:10.1080/00325481.2019.1655258
12. Ben-Harari RR, Goodwin E, Casoy J. Adverse event profile of pyrimethamine-based therapy in toxoplasmosis: a systematic review. Drugs R D. 2017;17(4):523–544. doi:10.1007/s40268-017-0206-8
13. Tari RM, Diallo A, Kouame E, et al. Assessment of the teratogenic effect of sulfadoxine-pyrimethamine on the chicken embryo. J Toxicol. 2022;2022:2995492. doi:10.1155/2022/2995492
14. Waldman BS, Schwarz D, Wadsworth MH, et al. Identification of a master regulator of differentiation in Toxoplasma. Cell. 2020;180(2):359–372e16. doi:10.1016/j.cell.2019.12.013
15. Tian X, Yang Z, Wan G, et al. Vaccination with recombinant Toxoplasma gondii bradyzoite-formation deficient 1 (rTgBFD1) antigen provides partial protective immunity against chronic T. gondii infection. Front Vet Sci. 2022;9:957479. doi:10.3389/fvets.2022.957479
16. Wang F, Holmes MJ, Hong HJ, et al. Translation initiation factor eIF1.2 promotes Toxoplasma stage conversion by regulating levels of key differentiation factors. Nat Commun. 2024;15(1):4385. doi:10.1038/s41467-024-48685-4
17. Sokol-Borrelli SL, Reilly SM, Holmes MJ, et al. A transcriptional network required for bradyzoite development in Toxoplasma gondii is dispensable for recrudescent disease. Nat Commun. 2023;14(1):6078. doi:10.1038/s41467-023-40948-w
18. Licon MH, Giuliano CJ, Chan AW, et al. A positive feedback loop controls Toxoplasma chronic differentiation. Nat Microbiol. 2023;8(5):889–904. doi:10.1038/s41564-023-01358-2
19. Murphy RD, Chen T, Lin J, et al. The Toxoplasma glucan phosphatase TgLaforin utilizes a distinct functional mechanism that can be exploited by therapeutic inhibitors. J Biol Chem. 2022;298(7):102089. doi:10.1016/j.jbc.2022.102089
20. Murphy RD, Troublefield CA, Miracle JS, et al. TgLaforin, a glucan phosphatase, reveals the dynamic role of storage polysaccharides in Toxoplasma gondii tachyzoites and bradyzoites. bioRxiv. 2023:2023–09.
21. Chen P, Lyu C, Wang Y, Pan M, Lin X, Shen B. Key roles of amylopectin synthesis and degradation enzymes in the establishment and reactivation of chronic toxoplasmosis. Animal Diseases. 2023;3(1):18.
22. El-Kady AM, Al-Megrin WAI, Abdel-Rahman IAM, et al. Ginger is a potential therapeutic for chronic toxoplasmosis. Pathogens. 2022;11(7):798. doi:10.3390/pathogens11070798
23. Jiang Y, Shi Y, Hu D, et al. The anti-Toxoplasma activity of the plant natural phenolic compound piceatannol. Front Vet Sci. 2022;9:972500. doi:10.3389/fvets.2022.972500
24. Jiang T, Godinez-Macias KP, Collins JE, et al. Identification of fungal natural products with potent inhibition in Toxoplasma gondii. Microbiol Spectr. 2024;12(4):e0414223. doi:10.1128/spectrum.04142-23
25. Tu V, Tomita T, Sugi T, et al. The Toxoplasma gondii cyst wall interactome. mBio. 2020;11(1). doi:10.1128/mBio.02699-19
26. Bando H, Murata Y, Han Y, et al. Toxoplasma gondii chitinase-like protein TgCLP1 regulates the parasite cyst burden. Front Cell Infect Microbiol. 2024;14:1359888. doi:10.3389/fcimb.2024.1359888
27. Tu V, Mayoral J, Sugi T, et al. Enrichment and proteomic characterization of the cyst wall from in vitro Toxoplasma gondii cysts. mBio. 2019;10(2). doi:10.1128/mBio.00469-19
28. Najm R, Ruivo MTG, Penarete-Vargas DM, et al. Invasion of Toxoplasma gondii bradyzoites: molecular dissection of the moving junction proteins and effective vaccination targets. Proc Natl Acad Sci U S A. 2023;120(5):e2219533120. doi:10.1073/pnas.2219533120
29. Radke JB, Melillo B, Mittal P, et al. Bicyclic azetidines target acute and chronic stages of Toxoplasma gondii by inhibiting parasite phenylalanyl t-RNA synthetase. Nat Commun. 2022;13(1):459. doi:10.1038/s41467-022-28108-y
30. Xia J, Fu Y, Huang W, Uddin T, Sibley LD. Constitutive upregulation of transcription factors underlies permissive bradyzoite differentiation in a natural isolate of Toxoplasma gondii. mBio. 2024;15(9):e0064124.
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms.php
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms.php
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.