")
Back to Journals » International Journal of Nanomedicine » Volume 19
Physicochemical Property Effects on Immune Modulating Polymeric Nanoparticles: Potential Applications in Spinal Cord Injury
Authors Kolpek DJ , Kim J, Mohammed H , Gensel JC, Park J
Received 25 September 2024
Accepted for publication 26 November 2024
Published 12 December 2024 Volume 2024:19 Pages 13357—13374
DOI https://doi.org/10.2147/IJN.S497859
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Eng San Thian
Daniel J Kolpek,1 Jaechang Kim,1 Hisham Mohammed,1 John C Gensel,2 Jonghyuck Park1,2
1Department of Pharmaceutical Sciences, College of Pharmacy, University of Kentucky, Lexington, KY, USA; 2Spinal Cord and Brain Injury Research Center, Department of Physiology, College of Medicine, University of Kentucky, Lexington, KY, USA
Correspondence: Jonghyuck Park, Pharmaceutical Sciences, College of Pharmacy, Spinal Cord and Brain Injury Research Center, College of Medicine, University of Kentucky, 789 S. Limestone St, Lexington, KY, 40506, USA, Tel +1 859 257 1850, Email [email protected]
Abstract: Nanoparticles (NPs) offer promising potential as therapeutic agents for inflammation-related diseases, owing to their capabilities in drug delivery and immune modulation. In preclinical studies focusing on spinal cord injury (SCI), polymeric NPs have demonstrated the ability to reprogram innate immune cells. This reprogramming results in redirecting immune cells away from the injury site, downregulating pro-inflammatory signaling, and promoting a regenerative environment post-injury. However, to fully understand the mechanisms driving these effects and maximize therapeutic efficacy, it is crucial to assess NP interactions with innate immune cells. This review examines how the physicochemical properties of polymeric NPs influence their modulation of the immune system. To achieve this, the review delves into the roles played by innate immune cells in SCI and investigates how various NP properties influence cellular interactions and subsequent immune modulation. Key NP properties such as size, surface charge, molecular weight, shape/morphology, surface functionalization, and polymer composition are thoroughly examined. Furthermore, the review establishes connections between these properties and their effects on the immunomodulatory functions of NPs. Ultimately, this review suggests that leveraging NPs and their physicochemical properties could serve as a promising therapeutic strategy for treating SCI and potentially other inflammatory diseases.
Keywords: nanomaterials, biodegradable carriers, particle engineering, biomaterial-based therapies, neuroinflammation, nerve tissue repair
Introduction
Spinal cord injury (SCI) results in loss of sensation, movement, and autonomic functions of the body below the level of injury.1,2 The time course of SCI can be divided into two phases: the primary injury and prolonged secondary injury.3,4 The primary injury is defined as the initial mechanical trauma to the spinal cord initiating various secondary physiological processes.4–6 The secondary physiological disruptions following SCI include the disturbance of the blood-spinal cord barrier, inflammation, glial scarring, demyelination, compromised axonal regeneration, and neuronal death. These disruptions lead to various functional disorders such as locomotor and somatosensory dysfunction.7–9
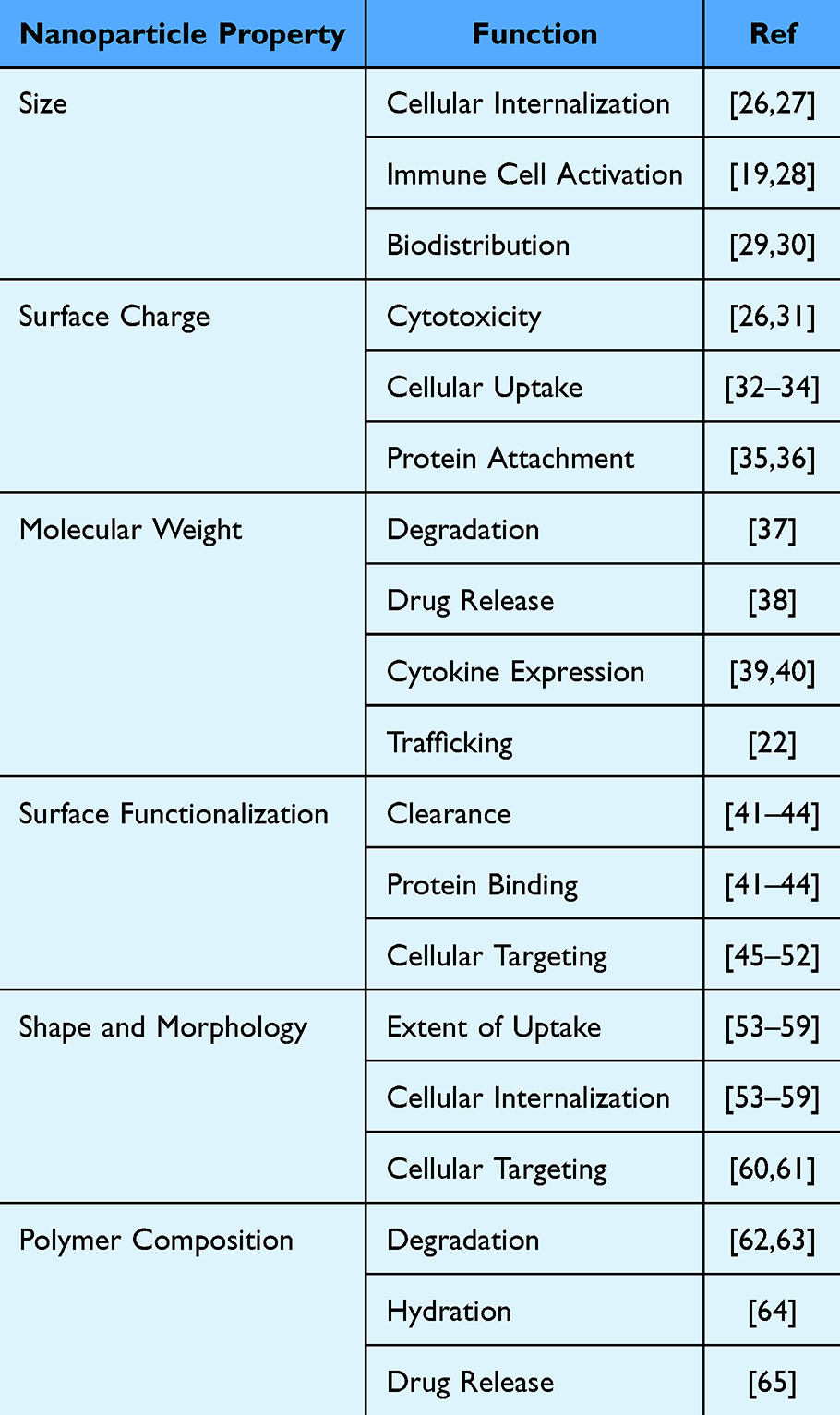 |
Table 1 Properties of Nanoparticles and Their Major Functions |
Currently, treatment options for SCI remain limited with no FDA approved therapeutics that restore neurological function.10 Previously, glucocorticoids such as methylprednisolone were used to treat SCI due to their anti-inflammatory effects. However, they have since been linked to many severe side effects, leading to a decline in their use and creating a gap between patient needs and effective treatment options.4 Several treatments for SCI are currently in preclinical development. These include rehabilitation,11 electrical stimulation of the motor cortex and spinal cord,12 transplantation of stem cells,13 and the use of antibodies or pharmacological agents to target various axon growth inhibitory molecules in the central nervous system (CNS).14–18
Recently, polymeric nanoparticles (NPs) have emerged as a promising therapeutic treatment for SCI. NPs are broadly classified as particles with dimensions on the nanometer scale and can vary in multiple characteristics, such as base material and physicochemical properties, which is a major focus of this review. Previously, NPs have been utilized for drug delivery owing to their ability to increase the safety and efficacy of various drugs.19 In addition to drug delivery, NPs have recently shown potential in modulating the immune response to SCI. Studies have demonstrated the ability of polymeric NPs to target the innate immune response after SCI, and a few preclinical studies have shown beneficial effects such as behavioral recovery and axonal regrowth in rodent models of SCI.20–22 The effects of NPs are influenced by their physicochemical properties, including size, surface charge, polymer molecular weight, surface functionalization, shape/morphology, and polymer composition (Table 1).23–25 These NP properties are key design features and have been shown to influence interactions with immune cells, resulting in varied therapeutic outcomes.23–25 In this review, we will investigate multiple physicochemical properties and their effects. Understanding the relationship between these properties and the ability of NPs to target specific outcomes in SCI enables the design of customized NP treatments. This tailored approach offers a promising therapeutic option for individuals with SCI.
Pathophysiology of SCI and Therapeutic Implications
The disruption of the blood-spinal cord barrier (BSCB) after initial traumatic SCI results in an inflammatory response, causing the invasion of circulating innate immune cells such as neutrophils, monocytes, and macrophages and the release of various inhibitory factors including pro-inflammatory factors at the injury site.66
The inflammatory response after SCI consists of both adaptive and innate immune responses. Several cell types and inflammatory cytokines are involved in mediating the inflammatory response to SCI. Hellenbrand et al, has described the chemokine cascade and cell infiltration patterns after SCI.66 During the early stages of injury, the resident microglia changes into pro-inflammatory cells that secrete cytokines and trigger peripheral immune cell infiltration.45,67 These changes contribute to myelin disruptions and fiber degeneration known as Wallerian degeneration (WD), leading to delayed sensorimotor dysfunction.3 Neutrophils are the first peripheral immune cell to arrive at the injury site within 4–6 hours and produce proteolytic and oxidative enzymes to prepare the area for repair.68 During 6–12 hours post injury, polymorphonuclear leukocytes are seen at the injury site and release reactive oxygen and nitrogen species (ROS and NOS).69 These ROS and NOS create widespread cell damage within the injury site, promoting an inflammatory and growth inhibiting environment, contributing to functional loss after SCI.70 From 24 hours to 7 days, T cells, monocytes, and dendritic cells are recruited to the injury site. Beck et al, using flow cytometry in rats with SCI demonstrated that neutrophils peak after 1 day, macrophages and microglia peak after 7 days, and T cells peak after 9 days.71 Following infiltration, both B and T cells persist indefinitely within the injury site and continue to increase in number for at least 9 weeks.45,72–74 They continue to promote cytokine/chemokine imbalance within the injury site, promoting inflammation and immune cell activation.72,75 Additionally, they remain within the injury site following repair of the BSCB, suggesting that B and T cells are reactivated locally within the injury site, leading to long injury residence times.73
From 7 days to 2 weeks, there is proliferation of oligodendrocyte progenitor cells resulting in improper remyelination.76 Infiltration of fibroblast-like cells and the formation of the fibrotic tissue matrix in mice occurs at 2 weeks.77 After 2 weeks, astrocytes form the glial scar around the lesion expressing glial fibrillary acidic protein (GFAP).78 Additionally, pericytes enter the lesion with blood vessel sprouts and form the stromal component of the scar tissue.79 The scar itself inhibits multiple repair processes, including axonal regeneration, sprouting, conduction, and remyelination, preventing the reconnection of axons above and below the level of injury and contributing to paralysis.80 The final glial scar has infiltrating immune cells in the center, microglia interacting with pericytes around the edges, and astrocytes surrounding the periphery (Figure 1). Collectively, both the innate and adaptive immune response along with changes to the intraspinal environment in response to injury lead to limited endogenous repair and ongoing secondary injury.
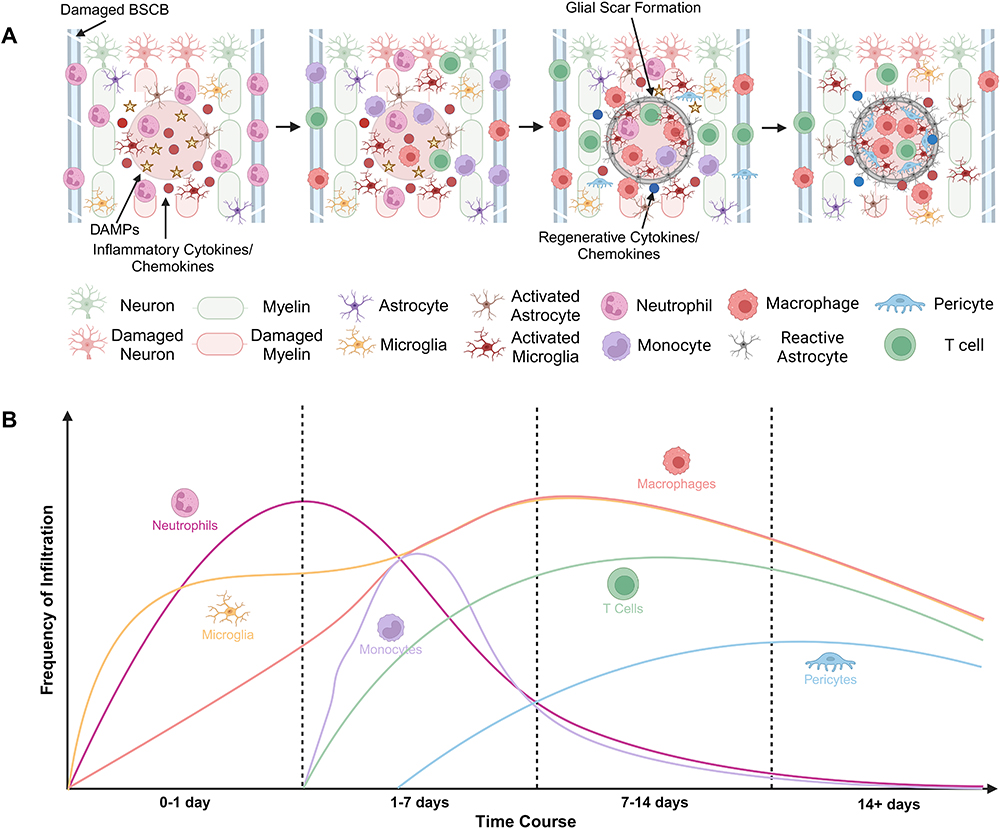 |
Figure 1 (A) Schematic depicting time course of SCI and infiltration of immune cells. (B) Graph illustrating the infiltration frequency of various immune cell populations over the time course of SCI. Created in BioRender. Kolpek, D (2024) https://BioRender.com/w82h824. |
Given the complex pathophysiology of SCI and the limited availability of treatments, NP technology may offer minimally invasive and multimodal therapeutic tools to address both the neuroinflammatory responses and multifaceted immune dysfunction associated with SCI. Various NP types have been studied, each with unique mechanisms and benefits. Anti-inflammatory drug loaded NPs have shown potential in manipulating the immune response to SCI. One study using chitosan/poly-(γ-glutamic acid) based NPs loaded with the non-steroidal anti-inflammatory drug diclofenac demonstrated inhibition of prostaglandin E2 synthesis and IL-6 production in lipopolysaccharide (LPS)-treated human macrophages, leading to downregulation of inflammatory signaling.81 Despite their anti-inflammatory effects, these NPs failed to significantly impact pro-regenerative macrophage markers like IL-10, a key factor in promoting regeneration after SCI. In contrast, studies using antioxidant loaded NPs have been able to show both a reduction in inflammation and promotion of regeneration by minimizing oxidative stress. One study using curcumin-loaded poly(lactic-co-glycolic acid) (PLGA) NPs released from a gelatin/alginate scaffold in the spinal cord injury site of rat models found increased functional recovery and axonal regrowth following injury.82 In a separate study, curcumin-loaded lipid NPs were shown to regulate the TLR4/2-NF-κB pathway in mice models for sepsis, which plays a major role in the immune response to SCI.83 Additional antioxidants such as catechin from green tea have also demonstrated anti-inflammatory effects when delivered via NPs to mouse macrophages.84
Despite the potential for anti-inflammatory drug and antioxidant-loaded NPs, other approaches have focused on using NPs to both directly and indirectly modulate the immune response to SCI without the delivery of additional compounds, providing a less invasive method and minimizing the potential for off-target effects. For example, we previously used intravenously injected PLGA NPs to reprogram the immune response after SCI in mice by interacting with circulating innate immune cells such as macrophages, monocytes, and neutrophils in the blood.20 Systemically administered NPs can selectively target and reprogram immune cells, particularly cell populations with scavenger receptors such as Macrophage Receptor with Collagenous Structure (MARCO)-positive subsets.20,85 These NP-associated cells are sequestered in the spleen and undergo apoptotic change, thereby delaying the infiltration into the injury site. This limits intraspinal cell accumulation and decreases pathogenic inflammation.20 Therefore, NPs indirectly attenuate the immune pathology in the inflamed area after injury. Generally, systemically administered NPs are taken up by immune cells in the vasculature and accumulate in the lysosome.86 The PLGA-based NPs undergo lysosomal biodegradation, generating glycolic and lactic acids that are metabolized in the Krebs cycle and eliminated as carbon dioxide and water.87 These byproducts effectively attenuate inflammatory responses by limiting NF-kB nuclear localization and modifying effector proteins, resulting in reduced expression of multiple inflammatory factors after injury.88 Published data demonstrate the accumulation of NP-positive cells at the injury site, which enables the upregulation of multiple pro-regenerative associated genes (RAGs) that create a more permissive environment after injury.20,21 Thus, this approach excludes invasive strategies such as surgical procedures and localized drug administration. In addition, the NP-mediated approach has more effective therapeutic strategies relative to antibody- and/or clodronate-encapsulated liposome-mediated approaches which can only employ a single factor or deplete entire systemic immune cell populations.89,90 Overall, NPs can serve as minimally invasive acute care of SCI, providing spinal tissue protection upon injury by reprogramming inflammatory immune cells, in turn reducing the development of an inhibitory environment and enhancing neurological outcomes after SCI.
As foreign substances, injected NPs rapidly engage the innate immune system, triggering tailored immune responses depending on their physicochemical properties. This underscores the potential of using engineered NP properties to modulate innate pro-inflammatory cell phenotypes, including inflammatory monocytes, neutrophils, and macrophages.20 The physicochemical properties of NPs greatly influence the interactions between NPs and the innate immune system, particularly the downstream signaling for immunomodulatory events.25,26,91 Here we summarize the effects of physicochemical properties of NPs on immunomodulation and apply them to SCI.
Impact of Nanoparticle Size on Innate Immune Cells
The size of NP can significantly impact interactions with innate immune cells, influencing cellular internalization, immune cell activation, and biodistribution.27,92,93 Five distinct uptake pathways have been shown to play a role in the internalization of NPs. These include clathrin-mediated endocytosis (CME), fast endophilin-mediated endocytosis (FEME), clathrin-independent carrier (CLIC)/glycosylphosphatidylinositol-anchored protein enriched early endocytic compartment (GEEC) endocytosis, macropinocytosis, and phagocytosis. Furthermore, these pathways can all be subclassified based on their clathrin and dynamin dependencies (Figure 2).86 Previously, caveolar-dependent endocytosis was considered to be significant. However, it has been reported that while caveolae can be present on the plasma membrane, few nanocarriers rely on them for uptake. This discrepancy is due to the insufficient specificity of inhibitors targeting endocytic pathways.28,86
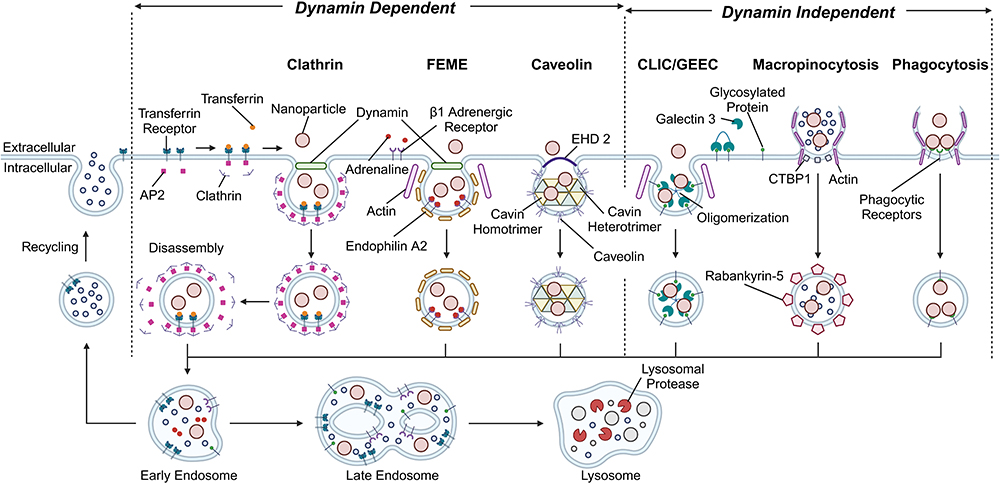 |
Figure 2 Primary uptake mechanisms used for NP internalization in cells. Created in BioRender. Kolpek, D (2024) https://BioRender.com/o86q647. |
The route of cellular uptake used among innate immune cells to internalize polymer-based NPs has been shown to vary widely with size. One study using 100 nm poly(methyl methacrylate) (PMMA) based NPs found LPS-activated microglia to selectively uptake NPs through CME.29 A similar study using mouse macrophages showed that smaller 40 nm polystyrene (PS) NPs were taken up by a combination of CME and macropinocytosis. In contrast, only phagocytosis and macropinocytosis were used to take up larger 600 nm NPs.94 The overall pattern indicates that immune cells employ a mix of CME, FEME, CLIC/GEEC, and macropinocytosis to internalize smaller NPs (<200 nm), whereas phagocytosis and macropinocytosis are utilized for larger NPs (>200 nm).86
The processes of CME, FEME, and phagocytosis are specific pathways and are well controlled through receptors.86 However, CLIC/GLEEC and macropinocytosis are non-specific pathways that internalize both NPs and other fluids into the cell.30,86 Certain findings indicate that this variation in endocytic specificity may result in diverse immune responses. One study by Park et al found PLGA-based NPs with a diameter of 500 nm can effectively reprogram innate immune cells through association with scavenger receptors that rely on dynamin-dependent endocytosis and macropinocytosis.20,95 Furthermore, these NPs facilitated the differentiation of macrophages towards an anti-inflammatory phenotype following SCI in mice models.20 In contrast, another study using smaller polypyrrole (PPy) based NPs ranging in size from 20 nm to 100 nm implicated the mid-sized 60 nm NPs to induce the highest production of ROS and costimulatory markers in macrophages.96 While variations in specificity of the endocytic uptake route likely contribute to these results,86 the exact mechanism remains unknown and requires further investigation before conclusions can be made.
In addition to cellular uptake route, particle size also acts as a major determinant for where NPs accumulate within the body and whether they reach the targeted cell population. One study comparing 50 nm and 500 nm PS NPs found higher agglomeration of larger (500 nm) NPs in the liver.97 This result was linked with protein attachment to the NP surface, commonly referred to as the protein corona, and subsequent opsonization. The protein corona has been shown to influence multiple biodistribution factors including NP circulation time, clearance, and ability to cross biological barriers depending on the attached proteins.98 These principles were demonstrated by a study using pegylated poly methoxypolyethyleneglycol cyanoacrylate-co-n-hexadecyl cyanoacrylate (PEG-PHDCA) NPs with varied size which found a strong correlation between NP size and protein absorption. Additionally, the results showed that smaller NPs with less protein absorption took twice as long to clear compared to larger NPs with greater protein absorption. Furthermore, the uptake percentage by murine macrophages was reported to decrease with increased NP size.31 Overall, larger NPs tend to have faster clearance due to hepatic filtration and opsonization following protein attachment, while smaller NPs remain in the blood longer and can extravasate into permeable tissues, increasing their uptake.99
The Effects of Nanoparticle Surface Charge on Immune Interactions, Cytotoxicity, and Therapeutic Implications
Surface charge of polymeric NPs can vary widely, ranging from negative (below −10 mV), to neutral (−10 mV to +10 mV), to positive (above +10 mV).100 Variations in charge are often accomplished through surface attachment of different surfactants during the fabrication process. Some commonly used surfactants include poly(ethylene-alt-maleic anhydride) (PEMA), poly(vinyl alcohol) (PVA), and Tween.20,67,101,102 Surface charge has been shown to affect immune cell interactions through alterations in cytotoxicity, cellular uptake, and protein attachment/opsonization. Each of these factors can greatly influence the overall immune response and resulting treatment efficacy, thus highlighting the need for a better understanding of the effects of NP surface charge on immune modulation.
NP cytotoxicity has been shown to differ significantly among treatments with varied surface charge. It is well established that positively charged NPs exhibit acute systemic toxicity and nonspecific stimulation of the immune system.91 One study using human neutrophils found that cationic NPs interact with the cell membrane to induce membrane disruption, calcium influx, oxidative stress, and cell death.32 The exact mechanism behind these effects remains unclear, but one potential cause is the activation of pattern-recognition receptors (PRRs) in immune cells via the cationic polymer base of the fabricated NPs.33 PRRs are well known for their role in the inflammatory cascade following SCI. Specifically, the toll-like receptor (TLR) family of PRRs facilitate the immune response through signaling mechanisms in microglia, astrocytes, macrophages, neurons, and oligodendrocytes.34 The activation of these cells via TLRs has been shown to inhibit neuronal regeneration, promote growth cone collapse, trigger neuronal cell death, and promote inflammatory signaling.34 Despite their cytotoxicity, positively charged polymeric NPs have shown promise in pulmonary immunization and gene delivery.103,104 However, modifications are necessary to minimize cytotoxicity for these cationic particles to progress beyond in vitro studies. In contrast, both neutral and negatively charged polymeric NPs have shown minimal toxicity and promising immunomodulatory properties, such as downregulation of pro-inflammatory signaling and upregulation of pro-regenerative cytokines.20,101 These effects are attributed to the inhibition of the LPS-TLR4 cascade by negatively charged NPs, which leads to a decrease in the inflammatory response by macrophages following SCI.20,34 Overall, cationic polymeric NPs exhibit higher risk due to their greater toxicity through TLR signaling activation compared to neutral and negatively charged alternatives, which have shown minimal toxicity with promising immunomodulatory potential.
The route of cellular internalization and extent of cellular uptake also heavily depends on the surface charge of the NP. Positively charged NPs exhibit increased interaction with the negatively charged cell membrane. These electrostatic interactions lead to increased uptake, which has been demonstrated in both macrophages and dendritic cells.105,106 Studies have also indicated a relationship between surface charge and cellular internalization pathways, with one finding positively charged particles to be taken up by clathrin-mediated endocytosis, while caveolin-mediated endocytosis was the preferred endocytic route for neutral and negatively charged NPs.107 How these differences influence the downstream effects of the NPs remains unknown and must be further studied for improved NP-based therapies.
Protein attachment to the surface of the NPs and the formation of the protein corona have also been shown to be impacted by surface charge.98 Neutral NPs without a strong surface charge have been shown to absorb fewer proteins when compared to NPs with a strong charge.35 Positively charged NPs tend to accumulate a thicker coating of plasma proteins compared to negatively charged NPs due to the abundance of negatively charged proteins in circulation.36,108 Beyond the extent of protein absorption, surface charge has been shown to influence the types of proteins absorbed, with basic proteins preferring negatively charged NPs and acidic proteins preferring positively charged NPs.109 The increased protein attachment has also been shown to generate a greater rate of opsonization, leading to faster clearance for charged NPs compared to treatments involving more neutral NPs.37 Recent research has achieved success by utilizing these effects to selectively enhance the attachment of specific proteins. One study using PS NPs with varied charge showed little variation in concentration of the most abundant corona proteins but found an increased presence of less abundant clusterin proteins in the negatively charged NP group.38 Protein attachment due to surface charge has also been shown to influence the immune response to NPs. One study using silica NPs with varied surface chemistries in human whole blood found negatively charged NPs with hydrocarbon groups to bind albumin and increase pro-inflammatory signaling in macrophages.39 Negative NPs functionalized with acid groups showed a similar inflammatory response due to complement and lower apoprotein binding. On the other hand, positively charged NPs functionalized with amine groups showed increased levels of anti-inflammatory marker expression.39 These results highlight the value of manipulating the surface charge of NPs to create a more targeted treatment and underscore the need for additional research in the area.
The Role of Polymer Molecular Weight in the Immune Response
Many polymer bases used for NP fabrication come in a variety of molecular weights (MW). Some of the most prevalent examples include chitosan, PLGA, and polyethylene glycol (PEG).40 MW has been shown to influence multiple factors, including degradation rate, hydration, and drug release rates.41,42,110 However, the role of polymer MW and its effects on immune modulation remains largely unexplored in the context of SCI.
Multiple studies using varied polymer MWs have demonstrated a clear relationship between MW and the resulting immune response. One study examining the effect of varied hyaluronic acid (HA) MW found high MW HA to decrease both nitric oxide (NO) production and gene expression of classical pro-inflammatory macrophage markers in LPS-activated RAW 264.7 macrophages, while medium MW HA increased the expression of these markers. Despite these contrasting pro-inflammatory responses, both high and medium MW increased anti-inflammatory gene expression.43 Another study examining the immunomodulatory effects of polysaccharides isolated from Chlorella ellipsoidea found lower MW polysaccharides to reduce the production of NO in RAW 264.7 macrophages. Additionally, the lower MW polymers did not activate the pro-inflammatory NF-κB, JNK, and p38 pathways.44 These diverse results for different polymers highlight how the effects of MW can vary greatly between polymer bases.
The role of MW in polymer-based therapeutics has also been examined in the context of various inflammation-mediated disease states. A study by Casey et al examined how varied MW in PLGA-based NPs can affect TLR activation, which plays a major role in sepsis. Results showed that high MW NPs downregulated the expression of surface markers necessary for TLR activation in mouse bone marrow-derived macrophages, while low MW NPs had little to no effect. Additionally, high MW PLGA-based NPs showed greater suppression of pro-inflammatory cytokine production compared to their low MW counterparts.101 MW has also been studied in the context of experimental autoimmune encephalomyelitis (EAE) and cancer. Saito et al used low and high molecular weights of PLGA-based NPs to test their role in immune cell trafficking and therapeutic effect on EAE. Intravenous PLGA injections into EAE mice showed that high molecular weight PLGA had reduced clinical scores compared to controls and was associated with neutrophils accumulated in the spleen.23 This research suggests that high molecular weight PLGA redirects trafficking of immune cells and reduces inflammation. It also highlights the ability of MW to influence the resulting immune modulation following NP treatments.
Overall, these studies clearly indicate that MW plays an important role in the immune modulation of polymer-based therapeutics and underscores the need for more research on the effects of varied polymeric NP MW on immune modulation after SCI. However, as demonstrated by the varied results of these studies, each polymer must be examined individually to establish a relationship between MW and downstream immune response.
Surface Functionalization of Nanoparticles
NPs can modulate immune responses through interaction with immune cells. Surface functionalization of NPs by attachment of polymers or functional groups can enhance this interaction by controlling the surface properties.111 PEGylation is a commonly used surface modification method to avoid clearance mechanisms and increase the circulation time of NPs. Conjugation of PEG to the NP surface hinders non-specific interactions with proteins through PEG’s hydrophilic and steric repulsive effects. This reduces the opsonization of particles by the reticuloendothelial system.46–49 For example, PEGylated polycyanoacrylate NPs have been shown to increase uptake in neural tissues by improving blood circulation compared to non-PEGylated NPs.50 In addition, conjugated PEG affects immune cell internalization. Microglia internalization was shown to vary depending on the amount of mPEG chemically bound to nanogel. This suggests that by tuning the nanogel coating, uptake from microglia can be changed.51
Conjugating small molecules or peptides to the surface can target specific cells, including immune cells and neural tissues, and increase the therapeutic effects.52,112 Wu et al employed apamin, a honeybee-derived peptide known for its penetration and distribution to the CNS, and developed apamin-conjugated PEG-distearoylphosphatidyl-ethanolamine (PEG-DSPE) micelles. These apamin-conjugated micelles enhanced drug accumulation in the spinal cord, and showed high efficacy in tissue repair after SCI.113 Surnar et al engineered NPs by incorporating the triphenylphosphonium (TPP) cation that accumulates in the mitochondria of cells that are actively involved in respiration. NPs made with PLGA-b-PEG functionalized with TPP demonstrated the ability to accumulate in the brain and provide neuroprotection.114 Gao et al developed NPs with curcumin, which can reduce the infiltration of inflammatory macrophages and induce macrophage polarization. The NPs were fabricated by a co-loading approach combining retinoic acid and curcumin with bovine serum albumin. This NP was shown to polarize macrophages, inhibit the release of inflammatory mediators, promote functional neuron regeneration, and improve motor function.53 Hua et al developed a nanocarrier as a combination treatment for SCI. This ROS-responsive nanocarrier mPEG-b-Lys-BECI-TCO can bind CD44 on the surface of human umbilical cord mesenchymal stem cells (Huc-MSCs) via Tz-A6 peptide. A combination of MSCs, NPs, and encapsulated Ferrostatin-1 promoted functional recovery in mice with SCI.54 Wang et al used apocynin (APO), which has the ability to reduce secondary damage, and synthesized an amphipathic block copolymer (Allyloxypolyethyleneglycol-b-poly [2-(((4-acetyl-2-methoxyphenoxy)carbonyl)oxy)ethyl methacrylate], PAPO). They also introduced a scar tissue-homing peptide (cysteine-alanine-glutamine-lysine, CAQK) into PAPO. The NP fabricated using these polymers effectively alleviated secondary damage and led to sustained recovery of motor function after SCI through lesion tissue-specific delivery and sustained release of APO.55 Kalashnikova et al conjugated miRNA to PLGA and formulated the NP with the excipient spermidine or polyethyleneimine. They used miRNA-129-5p, which has the ability to regulate microglia activation and checked the synergistic immunomodulatory effects between PLGA-based NPs and miRNA. These NPs exhibited sustained release of miRNAs, polarized activated microglia toward a more regenerative phenotype, and increased the production of pro-regenerative factors.67 Moreover, conjugation of cell adhesion peptides can affect the function and regulation of immune cells,56,57 and conjugation of ligand that binds to Fas, a cell surface receptor that contributes to the regulation of T lymphocytes, can also control the immune response.58
The Impact of Nanoparticle Geometry on Phagocytosis and Cellular Internalization in Innate Cells
Phagocytosis, a key component of innate immunity, involves antigen-presenting cells (APCs), such as macrophages internalizing antigen particles. The shape of NPs has a substantial impact on their cellular uptake. However, the role of NP geometry and its effects on immune modulation remains largely unexplored in the context of SCI.
In early investigations utilizing PS particles of varying sizes and shapes, NP shape was shown to exert a greater influence on absorption compared to its size. Specifically, the local shape of the particle at the point of initial contact with the macrophage was shown to play a crucial role in determining the initiation of phagocytosis. This is because the specific shapes of local particles have a direct impact on the actin structure, which is essential for the internalization process.59
Particles with a high aspect ratio (AR), such as elongated particles, show reduced macrophage-induced phagocytosis compared to spherical particles. Thus, worm-like particles characterized by very high ARs (>20) demonstrate minimal phagocytosis compared to conventional spherical particles of the same volume.60 Similarly, Geng et al found that longer worm-like micellar particles, known as filo micelles, exhibit reduced phagocytosis.61 Furthermore, other studies have indicated that high AR shapes contribute to increased NP residence time and decreased cellular internalization. The evaluation of human cell internalization of NPs with the same volume but different ARs showed that non-spherical particles had lower cellular absorption compared to spherical particles. This can be attributed to the average curvature radius of the particles adsorbed by the cell.62 In macrophages, the internalization of PS particles with three different shapes was studied utilizing spheres, oblate ellipsoids, and prolate ellipsoids. Spherical particles and oblate ellipsoids showed a greater degree of internalization than prolate ellipsoids. Phagocytosis is an actin-based process, so the extent of actin remodeling, an energy-intensive process, is related to internalization. Therefore, the shape that requires more actin remodeling induces slower internalization. The difference between oblate and prolate ellipsoids is likely due to differences in AR.64 In a study comparing nanodisk and nanosphere using PS, nanodisk showed lower permeability in human cell membranes.65
In contrast, some studies have shown different results. Using NPs larger than 100 nm, Gratton et al determined that rod-shaped particles showed the highest absorption, followed by spheres, cylinders, and cubes. Using particles made from cationic, cross-linked PEG hydrogel, rod-shaped high AR NPs (AR 3) were shown to internalize into human cells faster than symmetrical (AR 1) particles.63 In a study by Kolhar et al using equal volumes of PS nanospheres and nanorods, nanorods with targeting peptides showed higher endothelial specificity and lower nonspecific accumulation compared to nanospheres.115 In addition, when investigating the impact of particle AR on phagocytosis by different human immune cells, it was observed that neutrophils, unlike macrophages, exhibited a preference for internalizing rod-shaped particles over spherical ones. These differences may be due to differences in phagocytic mechanisms, such as increased motility of neutrophils and the dispensation of phosphorylation during phagocytosis.116 The variation in internalization tendencies across different cells provides a valuable opportunity to customize the shape of NPs according to the target cell, offering a means for precise and effective design.
Polymeric Materials of Nanoparticles for Immunomodulation in Spinal Cord Injury
The composition of polymer base used in NPs has demonstrated its influence on various factors, including degradation rate, hydration, and drug release rates.117–119 In the context of SCI, various polymeric materials have been developed and employed.120,121 Among these, some polymeric materials have immunomodulatory effects, like the previously mentioned PLGA.
Poly-Beta-Hydroxybutyrate (PHB)
PHB is a high molecular weight biopolyester that exists in the cytoplasm of many bacteria. Due to its biodegradability and non-toxicity, it has been widely used as a biomaterial for many years and has also been approved by the Food and Drug Administration (FDA).122 One study using a PHB scaffold showed the promotion of growth and differentiation of neural stem cells and supported marked axonal regeneration within the graft following SCI.123,124 In addition, PHB has shown immunomodulatory effects. For example, a study using a Poly(hydroxybutyrate-co-hydroxyvalerate)/polylactic acid/collagen (PHBV/PLA/Col) membrane reduced the infiltration of CD86+ macrophages to the lesion site.125 Moreover, PHB is slowly decomposed in vivo to produce β-hydroxybutyric acid (BHB), which was shown to attenuate microglial and glial activation in SCI mice while suppressing SCI-induced NLRP3 inflammasome.126 In another study, BHB attenuated neuroinflammation after SCI by downregulating the NLRP3 inflammasome and shifting the activation state of macrophages/microglia from an anti-regenerative to pro-regenerative phenotype.127 Although research on PHB-based nanoparticles for SCI is limited, their demonstrated immunomodulatory effect suggests that PHB holds potential as a material for NP-based treatment of SCI.
Polysialic Acid (PSA)
PSA is an endogenous carbohydrate polymer composed of α-2,8-linked N-acetyl neuraminic acid units that frequently attach to neural cell adhesion molecules (NCAM).128 PSA acts on cell-cell and cell-extracellular matrix interactions and plays an important role in axon regeneration, nervous system development, and plasticity.129–131 Because of these properties, PSA is being studied for SCI treatment and applied to nanotechnology. One study using PSA-based minocycline-loaded nanodrug delivery systems showed anti-inflammatory and neuroprotective activities both in vitro and in vivo. These NPs also protected the neurons and myelin sheaths from damage, and promoted neural regeneration, thereby improving the locomotor function of SCI rats.132 In another study, a polycaprolactone (PCL)/PSA hybrid nanofiber scaffold encapsulating methylprednisolone was implanted in the lesion area of rat SCI models. This reduced proinflammatory cytokine secretion by suppressing the activity of immune cells, promoted axonal growth, and improved functional recovery after SCI.133 Additionally, evidence of the role of PSA in immunomodulation has increased in recent years. Studies have shown that PSA decreases the activation of microglia and macrophages and attenuates the neuroinflammatory response by interacting with sialic acid-binding immunoglobulin-like lectins (Siglec). These results demonstrate the therapeutic potential of PSA as an immune modulating NP material for treatment of SCI.134
Chitosan
Chitosan is a linear polysaccharide, produced mainly from the exoskeleton of crustaceans. It has gained attention in biomedical applications due to its beneficial properties including biodegradability and low immunogenicity.135 Chitosan can be a promising material in treatment and management of inflammatory diseases. This is due to its anti-inflammatory activity by inhibiting activation of NF-kB and caspase-3 and lowering secretion of pro-inflammatory cytokines such as TNF-a and IL-6, and expression of iNOS.136,137 It also has proven effects on sealing nerve cell membranes and inhibiting neurotoxicity.138,139 Chitosan can be utilized as a material for NPs. One study used chitosan-based NPs to treat acrolein-mediated cell injury. The NPs effectively reduced damage to membrane integrity, secondary oxidative stress, and lipid peroxidation.140 Furthermore, chitosan-based NPs have shown the ability to restore nerve impulse transmission following SCI in guinea pigs.141 Glycol chitosan (GC), a derivative of chitosan, is recognized as a natural neuroprotective compound,142 and was employed to overcome the issue of chitosan’s poor solubility in neutral pH aqueous solutions.143 Ferulic acid-modified glycol chitosan NPs showed prolonged circulation time and effectively delivered both chitosan and ferulic acid to the damaged area. It significantly preserved axons and myelin while reducing inflammatory response at the lesion site, leading to improved locomotor recovery.144 Overall, chitosan shows great promise as a potential therapeutic tool for NP treatment of SCI.
Hyaluronic Acid (HA)
HA is a normal constituent of the extracellular matrix surrounding the basement membrane of multiple endogenous cell types and has many beneficial properties including non-toxicity, biodegradability, biocompatibility, and non-immunogenicity.145 Due to these properties, HA has great potential for use in immune modulating NPs. The HA receptor, CD44, is expressed on the majority of immune cells, thereby increasing interactions between NPs and CD44, and augmenting NP absorption into cells.146,147 Furthermore, the high molecular weight of HA exerts an anti-inflammatory effect through its interaction with immune cells. It diminishes proinflammatory cytokines and enhances the expression of anti-inflammatory genes, such as Arg1, IL10, and MRC1.43,148,149 Additionally, it has the ability to modify astrocyte proliferation and migration.150 Based on these characteristics, HA-based NPs present a promising therapeutic approach to regulate inflammatory responses following SCI.
Conclusion
In conclusion, NPs have demonstrated significant potential as therapeutic agents in SCI due to their immunomodulatory properties. In this review, we have detailed the pathophysiology of SCI, the role of inflammation, and the involvement of innate immune cells, while detailing how the physicochemical properties of polymer NPs influence their interactions with these cells. Key NP characteristics such as size, surface charge, molecular weight, shape, surface functionalization, and polymer composition all play crucial roles in modulating the immune response to SCI (Table 2).
 |
Table 2 Representative Papers Exhibiting Physicochemical Property Effects on Immune Modulation |
NP size acts as a major determinant of the cellular uptake route. These uptake routes include CME, FEME, CLIC/GLEEC, macropinocytosis, and phagocytosis.86 Each route varies in specificity, leading to variations in immune cell interactions and responses.20,86 Additionally, NP size greatly affects the formation of the protein corona. This can lead to variations in NP accumulation, uptake, and clearance, all of which are important factors for drug delivery.98 Overall, NP size plays a critical role in determining uptake pathways, cellular interactions, and protein corona formation, highlighting its importance in the development of NP treatments for SCI and other therapeutic applications.
Similar to size, the surface charge of NPs can influence cellular uptake and protein corona formation. Variations in charge can lead to changes in interaction with the cell membrane, causing differences in extent of uptake and cellular uptake pathway.105–107 Also, certain types of proteins can be attracted to the surface of the NP depending on the surface charge.109 This leads to changes in the protein corona composition and further downstream effects that impact the immune response to the NP.39 In addition, the toxicity of NPs can be affected by their charge, particularly with positively charged NPs exhibiting increased cytotoxicity due to nonspecific immune system stimulation.91 These factors must be considered when designing a safe and efficacious NP treatment.
Polymer MW remains an understudied factor in NP treatment of SCI despite multiple studies demonstrating its relevance in other inflammatory diseases. In sepsis, NPs with varied MW showed differences in TLR activation.101 In EAE, MW influenced the trafficking of the innate immune cells and clinical scores of the experimental mice.23 Given the similarities in inflammatory responses between SCI and these diseases, additional research on the effects of NP MW in the context of SCI is warranted.
Surface functionalization of NPs has shown strong potential for manipulating multiple factors including clearance, cellular interactions, and immune response. Surface conjugation with factors such as PEG can decrease nonspecific interactions with cells and opsonization, leading to greater circulation time.46–49 Additionally, multiple studies have demonstrated the ability of a surface conjugated peptide to traffic NPs to specific target sites such as the spinal cord and influence the downstream immune response following cell interaction.52,112 These studies highlight the value of surface functionalization and underscore the need for its utilization when developing effective NP treatments.
NP geometry is another critical factor in cellular uptake and immune modulation. Studies have identified the AR of a NP to be a major determinant of cellular uptake and absorption with certain immune cells preferring low AR or high AR NPs.60,63 This allows for the opportunity to engineer NPs targeted to specific immune cell populations. While the effects of NP geometry in SCI remain largely unexplored, investigating how different ARs influence spinal cord-infiltrating immune cells could lead to targeted treatments with improved therapeutic outcomes.
The polymer composition used when fabricating NPs considerably impacts the downstream effects of the NPs when considering cellular interactions in the context of SCI. The non-toxic and biodegradable PHB has been shown to modulate the immune response through changes in cellular trafficking and NLRP3 inflammasome activation.125,126 By utilizing its ability to interact with NCAM and influence cellular interactions, PSA has shown therapeutic potential as a drug delivery carrier and immunomodulator in SCI.128–133 Chitosan has the ability to inhibit multiple pro-inflammatory signaling pathways, and has been shown to minimize neurotoxicity, making it a promising NP material for SCI.136–139 Lastly, HA has the ability to modulate the immune response through CD44 interactions and has shown anti-inflammatory effects.146–149 By leveraging the characteristics of these various polymer bases, targeted NP treatments can be developed for both SCI and other inflammatory diseases.
Understanding these physicochemical properties and their specific interactions with immune cells is crucial for the engineering and design of targeted NPs that can be optimized for modulating the immune response to neurological diseases. However, despite a large base of knowledge connecting NP properties with immune modulation, studies applying this knowledge to SCI and related inflammatory diseases remain scarce.
Polymeric NPs offer significant clinical potential for immunomodulation in SCI. Using their physicochemical properties, NPs can be designed to target specific immune cell populations such as macrophages, microglia, and neutrophils.20,60,63,101 Through these interactions, the NPs can reduce the inflammatory response to SCI and polarize immune cells towards a more regenerative phenotype, allowing for greater recovery following SCI.20,101 Additionally, polymeric NPs have the capacity to act as drug carriers to the spinal cord injury site, where a controlled release of anti-inflammatory drugs, antioxidants, or genetic material can be facilitated.67,81,82 Through NP delivery, the degradation of these therapeutics can be minimized compared to more traditional pharmaceutical interventions.19 Another advantage polymeric NPs have is their minimal toxicity. Many polymeric NPs are composed of biodegradable components that are broken down to endogenous chemicals within the body, thus minimizing toxicity and off-target effects seen with other drugs.87,122,128,145 However, despite these benefits, polymeric NPs still face challenges such as potential long-term effects and efficacy in human subjects.19 These shortcomings must be addressed before polymeric NPs can be applied in a clinical setting for treatment of SCI.
Future research should focus on applying these findings to the development and design of NPs specifically tailored for SCI, with the goal of targeting immune responses that maximize therapeutic outcomes. Further investigation is needed to explore how specific NP designs can target multiple facets of the immune response including different immune cell populations, distinct inflammatory pathways, and improved functional recovery in SCI patients. By advancing this research, there is an opportunity to refine current therapeutic strategies for SCI and extend the application of NPs to a broader range of inflammatory and neurodegenerative diseases.
Funding
This work was supported by National Institutes of Health through R01NS136272 and R01NS116068, the National Center for Advancing Translational Sciences UL1 TR001998, the UKY Bioelectronics and Nanomedicine Research Center, and the Center for Pharmaceutical Research and Innovation (CPRI, NIH P20 GM130456). The content is solely the responsibility of the authors and does not necessarily represent the official views of the NIH.
Disclosure
Dr John Gensel reports a patent 11660269 issued to John C. Gensel. The authors report no other conflicts of interest in this work.
References
1. Ahuja CS, Wilson JR, Nori S, et al. Traumatic spinal cord injury. Nat Rev Dis Primers. 2017;3:17018. doi:10.1038/nrdp.2017.18
2. Park J, Lim E, Back S, Na H, Park Y, Sun K. Nerve regeneration following spinal cord injury using matrix metalloproteinase-sensitive, hyaluronic acid-based biomimetic hydrogel scaffold containing brain-derived neurotrophic factor. J Biomed Mater Res A. 2010;93(3):1091–1099. doi:10.1002/jbm.a.32519
3. Hausmann ON. Post-traumatic inflammation following spinal cord injury. Spinal Cord. 2003;41(7):369–378. doi:10.1038/sj.sc.3101483
4. Sterner RC, Sterner RM. Immune response following traumatic spinal cord injury: pathophysiology and therapies. Front Immunol. 2022;13:1084101. doi:10.3389/fimmu.2022.1084101
5. Park J, Zheng L, Marquis A, et al. Neuroprotective role of hydralazine in rat spinal cord injury-attenuation of acrolein-mediated damage. J Neurochem. 2014;129(2):339–349. doi:10.1111/jnc.12628
6. Page JC, Park J, Chen Z, Cao P, Shi R. Parallel evaluation of two potassium channel blockers in restoring conduction in mechanical spinal cord injury in rat. J Neurotrauma. 2018;35(9):1057–1068. doi:10.1089/neu.2017.5297
7. Pinelli F, Pizzetti F, Veneruso V, et al. Biomaterial-mediated factor delivery for spinal cord injury treatment. Biomedicines. 2022;10(7):1673. doi:10.3390/biomedicines10071673
8. Due MR, Park J, Zheng L, et al. Acrolein involvement in sensory and behavioral hypersensitivity following spinal cord injury in the rat. J Neurochem. 2014;128(5):776–786. doi:10.1111/jnc.12500
9. Park J, Zheng L, Acosta G, et al. Acrolein contributes to TRPA1 up-regulation in peripheral and central sensory hypersensitivity following spinal cord injury. J Neurochem. 2015;135(5):987–997. doi:10.1111/jnc.13352
10. Failli V, Kleitman N, Lammertse DP, et al. Experimental treatments for spinal cord injury: what you should know. Top Spinal Cord Inj Rehabil. 2021;27(2):50–74. doi:10.46292/sci2702-50
11. Hollis ER 2nd, Ishiko N, Yu T, et al. Ryk controls remapping of motor cortex during functional recovery after spinal cord injury. Nat Neurosci. 2016;19(5):697–705. doi:10.1038/nn.4282
12. Pal A, Park H, Ramamurthy A, et al. Spinal cord associative plasticity improves forelimb sensorimotor function after cervical injury. Brain. 2022;145(12):4531–4544. doi:10.1093/brain/awac235
13. Kumamaru H, Lu P, Rosenzweig ES, Kadoya K, Tuszynski MH. Regenerating corticospinal axons innervate phenotypically appropriate neurons within neural stem cell grafts. Cell Rep. 2019;26(9):2329–2339e4. doi:10.1016/j.celrep.2019.01.099
14. Park J, Farmer M, Casson C, Kalashnikova I, Kolpek D. Therapeutic potential of combinative shRNA-encoded lentivirus-mediated gene silencing to accelerate somatosensory recovery after spinal cord trauma. Neurotherapeutics. 2023;20(2):564–577. doi:10.1007/s13311-022-01331-7
15. Park J, Decker JT, Smith DR, Cummings BJ, Anderson AJ, Shea LD. Reducing inflammation through delivery of lentivirus encoding for anti-inflammatory cytokines attenuates neuropathic pain after spinal cord injury. J Control Release. 2018;290:88–101. doi:10.1016/j.jconrel.2018.10.003
16. Park J, Decker JT, Margul DJ, et al. Local Immunomodulation with anti-inflammatory cytokine-encoding lentivirus enhances functional recovery after spinal cord injury. Mol Ther. 2018;26(7):1756–1770. doi:10.1016/j.ymthe.2018.04.022
17. Stewart AN, Kumari R, Bailey WM, et al. PTEN knockout using retrogradely transported AAVs restores locomotor abilities in both acute and chronic spinal cord injury. bioRxiv. 2023;368:114502. 10.1101/2023.04.17.537179.
18. Park J. Immunomodulatory strategies for spinal cord injury. Biomed J Sci Tech Res. 2022;45(3):36467–36470. doi:10.26717/bjstr.2022.45.007202
19. Mitchell MJ, Billingsley MM, Haley RM, Wechsler ME, Peppas NA, Langer R. Engineering precision nanoparticles for drug delivery. Nat Rev Drug Discov. 2021;20(2):101–124. doi:10.1038/s41573-020-0090-8
20. Park J, Zhang Y, Saito E, et al. Intravascular innate immune cells reprogrammed via intravenous nanoparticles to promote functional recovery after spinal cord injury. Proc Natl Acad Sci U S A. 2019;116(30):14947–14954. doi:10.1073/pnas.1820276116
21. Jeong SJ, Cooper JG, Ifergan I, et al. Intravenous immune-modifying nanoparticles as a therapy for spinal cord injury in mice. Neurobiol Dis. 2017;108:73–82. doi:10.1016/j.nbd.2017.08.006
22. Saunders MN, Griffin KV, Kalashnikova I, et al. Biodegradable nanoparticles targeting circulating immune cells reduce central and peripheral sensitization to alleviate neuropathic pain following spinal cord injury. Pain. 2023;165:92–101. doi:10.1097/j.pain.0000000000002989
23. Saito E, Kuo R, Pearson RM, et al. Designing drug-free biodegradable nanoparticles to modulate inflammatory monocytes and neutrophils for ameliorating inflammation. J Control Release. 2019;300:185–196. doi:10.1016/j.jconrel.2019.02.025
24. Saito E, Kuo R, Kramer KR, et al. Design of biodegradable nanoparticles to modulate phenotypes of antigen-presenting cells for antigen-specific treatment of autoimmune disease. Biomaterials. 2019;222:119432. doi:10.1016/j.biomaterials.2019.119432
25. Getts DR, Shea LD, Miller SD, King NJ. Harnessing nanoparticles for immune modulation. Trends Immunol. 2015;36(7):419–427. doi:10.1016/j.it.2015.05.007
26. Liu D, Chen J, Jiang T, et al. Biodegradable spheres protect traumatically injured spinal cord by alleviating the glutamate-induced excitotoxicity. Adv Mater. 2018;30(14):e1706032. doi:10.1002/adma.201706032
27. Petrarca C, Clemente E, Amato V, et al. Engineered metal based nanoparticles and innate immunity. Clin Mol Allergy. 2015;13(1):13. doi:10.1186/s12948-015-0020-1
28. Parton RG. Caveolae: structure, function, and relationship to disease. Annu Rev Cell Dev Biol. 2018;34:111–136. doi:10.1146/annurev-cellbio-100617-062737
29. Papa S, Ferrari R, De Paola M, et al. Polymeric nanoparticle system to target activated microglia/macrophages in spinal cord injury. J Control Release. 2014;174:15–26. doi:10.1016/j.jconrel.2013.11.001
30. Pricop DAM. Endocytosis and exocytosis of gold nanoparticles. Romanian J Biophys. 2015;25(1):63–71.
31. Fang C, Shi B, Pei YY, Hong MH, Wu J, Chen HZ. In vivo tumor targeting of tumor necrosis factor-alpha-loaded stealth nanoparticles: effect of MePEG molecular weight and particle size. Eur J Pharm Sci. 2006;27(1):27–36. doi:10.1016/j.ejps.2005.08.002
32. Hwang TL, Aljuffali IA, Lin CF, Chang YT, Fang JY. Cationic additives in nanosystems activate cytotoxicity and inflammatory response of human neutrophils: lipid nanoparticles versus polymeric nanoparticles. Int J Nanomed. 2015;10:371–385. doi:10.2147/IJN.S73017
33. Chen H, Li P, Yin Y, et al. The promotion of type 1 T helper cell responses to cationic polymers in vivo via toll-like receptor-4 mediated IL-12 secretion. Biomaterials. 2010;31(32):8172–8180. doi:10.1016/j.biomaterials.2010.07.056
34. Kigerl KA, de Rivero Vaccari JP, Dietrich WD, Popovich PG, Keane RW. Pattern recognition receptors and central nervous system repair. Exp Neurol. 2014;258:5–16. doi:10.1016/j.expneurol.2014.01.001
35. Aggarwal P, Hall JB, McLeland CB, Dobrovolskaia MA, McNeil SE. Nanoparticle interaction with plasma proteins as it relates to particle biodistribution, biocompatibility and therapeutic efficacy. Adv Drug Deliv Rev. 2009;61(6):428–437. doi:10.1016/j.addr.2009.03.009
36. Almalik A, Benabdelkamel H, Masood A, et al. Hyaluronic acid coated chitosan nanoparticles reduced the immunogenicity of the formed protein corona. Sci Rep. 2017;7(1):10542. doi:10.1038/s41598-017-10836-7
37. Owens DE 3rd, Peppas NA. Opsonization, biodistribution, and pharmacokinetics of polymeric nanoparticles. Int J Pharm. 2006;307(1):93–102. doi:10.1016/j.ijpharm.2005.10.010
38. Gessner A, Waicz R, Lieske A, Paulke B, Mader K, Muller RH. Nanoparticles with decreasing surface hydrophobicities: influence on plasma protein adsorption. Int J Pharm. 2000;196(2):245–249. doi:10.1016/s0378-5173(99)00432-9
39. González-García LE, MacGregor MN, Visalakshan RM, et al. Nanoparticles surface chemistry influence on protein corona composition and inflammatory responses. Nanomaterials. 2022;12(4):682. doi:10.3390/nano12040682
40. Zielińska A, Carreiró F, Oliveira AM, et al. Polymeric nanoparticles: production, characterization, toxicology and ecotoxicology. Molecules. 2020;25(16):3731. doi:10.3390/molecules25163731
41. Tracy MA, Ward KL, Firouzabadian L, et al. Factors affecting the degradation rate of poly(lactide-co-glycolide) microspheres in vivo and in vitro. Biomaterials. 1999;20(11):1057–1062. doi:10.1016/s0142-9612(99)00002-2
42. Patel RB, Carlson AN, Solorio L, Exner AA. Characterization of formulation parameters affecting low molecular weight drug release from in situ forming drug delivery systems. J Biomed Mater Res Part A. 2010;94A(2):476–484. doi:10.1002/jbm.a.32724
43. Lee BM, Park SJ, Noh I, Kim C-H. The effects of the molecular weights of hyaluronic acid on the immune responses. Biomater Res. 2021;25(1):1–13. doi:10.1186/s40824-021-00228-4
44. Qi J, Kim SM. Effects of the molecular weight and protein and sulfate content of Chlorella ellipsoidea polysaccharides on their immunomodulatory activity. Int J Biol Macromol. 2018;107:70–77. doi:10.1016/j.ijbiomac.2017.08.144
45. Kigerl KA, McGaughy VM, Popovich PG. Comparative analysis of lesion development and intraspinal inflammation in four strains of mice following spinal contusion injury. J Comp Neurol. 2006;494(4):578–594. doi:10.1002/cne.20827
46. Woodle MC. Surface-modified liposomes: assessment and characterization for increased stability and prolonged blood circulation. Chem Phys Lipids. 1993;64(1–3):249–262. doi:10.1016/0009-3084(93)90069-F
47. Allen T. The use of glycolipids and hydrophilic polymers in avoiding rapid uptake of liposomes by the mononuclear phagocyte system. Adv Drug Delivery Rev. 1994;13(3):285–309. doi:10.1016/0169-409X(94)90016-7
48. Senior JH. Fate and behavior of liposomes in vivo: a review of controlling factors. Crit Rev Ther Drug Carrier Syst. 1987;3(2):123–193.
49. Senior J, Delgado C, Fisher D, Tilcock C, Gregoriadis G. Influence of surface hydrophilicity of liposomes on their interaction with plasma protein and clearance from the circulation: studies with poly (ethylene glycol)-coated vesicles. Biochimica et Biophysica Acta (BBA)-Biomembranes. 1991;1062(1):77–82. doi:10.1016/0005-2736(91)90337-8
50. Calvo P, Gouritin B, Villarroya H, et al. Quantification and localization of PEGylated polycyanoacrylate nanoparticles in brain and spinal cord during experimental allergic encephalomyelitis in the rat. Eur J Neurosci. 2002;15(8):1317–1326. doi:10.1046/j.1460-9568.2002.01967.x
51. Mauri E, Veglianese P, Papa S, et al. Chemoselective functionalization of nanogels for microglia treatment. Eur Polym J. 2017;94:143–151. doi:10.1016/j.eurpolymj.2017.07.003
52. Cerqueira SR, Ayad NG, Lee JK. Neuroinflammation treatment via targeted delivery of nanoparticles. Front Cell Neurosci. 2020;14:576037. doi:10.3389/fncel.2020.576037
53. Gao X, Han Z, Huang C, et al. An anti-inflammatory and neuroprotective biomimetic nanoplatform for repairing spinal cord injury. Bioact Mater. 2022;18:569–582. doi:10.1016/j.bioactmat.2022.05.026
54. Hua R, Zhao C, Xu Z, et al. ROS-responsive nanoparticle delivery of ferroptosis inhibitor prodrug to facilitate mesenchymal stem cell-mediated spinal cord injury repair. Bioact Mater. 2024;38:438–454. doi:10.1016/j.bioactmat.2024.05.015
55. Wang J, Li D, Liang C, et al. Scar tissue-targeting polymer micelle for spinal cord injury treatment. Small. 2020;16(8):1906415. doi:10.1002/smll.201906415
56. Kao WJ, Liu Y. Utilizing biomimetic oligopeptides to probe fibronectin-integrin binding and signaling in regulating macrophage function in vitro and in vivo. Front Biosci. 2001;6:D992–D999. doi:10.2741/Kao
57. Moral ME, Siahaan TJ. Conjugates of cell adhesion peptides for therapeutics and diagnostics against cancer and autoimmune diseases. Curr Top Med Chem. 2017;17(32):3425–3443. doi:10.2174/1568026618666180118154514
58. Hume PS, Anseth KS. Inducing local T cell apoptosis with anti-Fas-functionalized polymeric coatings fabricated via surface-initiated photopolymerizations. Biomaterials. 2010;31(12):3166–3174. doi:10.1016/j.biomaterials.2010.01.035
59. Champion JA, Mitragotri S. Role of target geometry in phagocytosis. Proc Natl Acad Sci. 2006;103(13):4930–4934. doi:10.1073/pnas.0600997103
60. Champion JA, Mitragotri S. Shape induced inhibition of phagocytosis of polymer particles. Pharm Res. 2009;26:244–249. doi:10.1007/s11095-008-9626-z
61. Geng Y, Dalhaimer P, Cai S, et al. Soft filaments circulate longer than spherical particles–shape effects in flow and drug delivery. Nat Nanotechnol. 2007;2:249–255. doi:10.1038/nnano.2007.70
62. Florez L, Herrmann C, Cramer JM, et al. How shape influences uptake: interactions of anisotropic polymer nanoparticles and human mesenchymal stem cells. Small. 2012;8(14):2222–2230. doi:10.1002/smll.201102002
63. Gratton SE, Ropp PA, Pohlhaus PD, et al. The effect of particle design on cellular internalization pathways. Proc Natl Acad Sci. 2008;105(33):11613–11618. doi:10.1073/pnas.0801763105
64. Sharma G, Valenta DT, Altman Y, et al. Polymer particle shape independently influences binding and internalization by macrophages. J Control Release. 2010;147(3):408–412. doi:10.1016/j.jconrel.2010.07.116
65. Zhang Y, Tekobo S, Tu Y, et al. Permission to enter cell by shape: nanodisk vs nanosphere. ACS Appl Mater Interfaces. 2012;4(8):4099–4105. doi:10.1021/am300840p
66. Hellenbrand DJ, Quinn CM, Piper ZJ, Morehouse CN, Fixel JA, Hanna AS. Inflammation after spinal cord injury: a review of the critical timeline of signaling cues and cellular infiltration. J Neuroinflammation. 2021;18(1):284. doi:10.1186/s12974-021-02337-2
67. Kalashnikova I, Cambell H, Kolpek D, Park J. Optimization and characterization of miRNA-129-5p-encapsulated poly (lactic-co-glycolic acid) nanoparticles to reprogram activated microglia. Nanoscale Adv. 2023;5(13):3439–3452. doi:10.1039/d3na00149k
68. Taoka Y, Okajima K, Uchiba M, et al. Role of neutrophils in spinal cord injury in the rat. Neuroscience. 1997;79(4):1177–1182. doi:10.1016/s0306-4522(97)00011-0
69. Stirling DP, Yong VW. Dynamics of the inflammatory response after murine spinal cord injury revealed by flow cytometry. J Neurosci Res. 2008;86(9):1944–1958. doi:10.1002/jnr.21659
70. Oyinbo C. Secondary injury mechanisms in traumatic spinal cord injury: a nugget of this multiply cascade. Acta neurobiologiae experimentalis. 2011;71:281–299. doi:10.55782/ane-2011-1848
71. Beck KD, Nguyen HX, Galvan MD, Salazar DL, Woodruff TM, Anderson AJ. Quantitative analysis of cellular inflammation after traumatic spinal cord injury: evidence for a multiphasic inflammatory response in the acute to chronic environment. Brain. 2010;133(Pt 2):433–447. doi:10.1093/brain/awp322
72. Ankeny DP, Popovich PG. Mechanisms and implications of adaptive immune responses after traumatic spinal cord injury. Neuroscience. 2009;158(3):1112–1121. doi:10.1016/j.neuroscience.2008.07.001
73. Ankeny DP, Lucin KM, Sanders VM, McGaughy VM, Popovich PG. Spinal cord injury triggers systemic autoimmunity: evidence for chronic B lymphocyte activation and lupus-like autoantibody synthesis. J Neurochem. 2006;99(4):1073–1087. doi:10.1111/j.1471-4159.2006.04147.x
74. Bilgen M, Dogan B, Narayana PA. In vivo assessment of blood-spinal cord barrier permeability: serial dynamic contrast enhanced MRI of spinal cord injury. Magnetic Resonance Imag. 2002;20(4):337–341. doi:10.1016/S0730-725X(02)00504-0
75. Babcock AA, Kuziel WA, Rivest S, Owens T. Chemokine expression by glial cells directs leukocytes to sites of axonal injury in the CNS. J Neurosci. 2003;23(21):7922. doi:10.1523/JNEUROSCI.23-21-07922.2003
76. Tripathi R, McTigue DM. Prominent oligodendrocyte genesis along the border of spinal contusion lesions. Glia. 2007;55(7):698–711. doi:10.1002/glia.20491
77. Sroga JM, Jones TB, Kigerl KA, McGaughy VM, Popovich PG. Rats and mice exhibit distinct inflammatory reactions after spinal cord injury. J Comp Neurol. 2003;462(2):223–240. doi:10.1002/cne.10736
78. Okada S, Hara M, Kobayakawa K, Matsumoto Y, Nakashima Y. Astrocyte reactivity and astrogliosis after spinal cord injury. Neurosci Res. 2018;126:39–43. doi:10.1016/j.neures.2017.10.004
79. Goritz C, Dias DO, Tomilin N, Barbacid M, Shupliakov O, Frisen J. A pericyte origin of spinal cord scar tissue. Science. 2011;333(6039):238–242. doi:10.1126/science.1203165
80. Alizadeh A, Dyck SM, Karimi-Abdolrezaee S. Traumatic spinal cord injury: an overview of pathophysiology, models and acute injury mechanisms. Front Neurol. 2019;10:282. doi:10.3389/fneur.2019.00282
81. Gonçalves RM, Pereira ACL, Pereira IO, Oliveira MJ, Barbosa MA. Macrophage response to chitosan/poly-(γ-glutamic acid) nanoparticles carrying an anti-inflammatory drug. J Mater Sci Mater Med. 2015;26(4):167. doi:10.1007/s10856-015-5496-1
82. Ai A, Hasanzadeh E, Safshekan F, et al. Enhanced spinal cord regeneration by gelatin/alginate hydrogel scaffolds containing human endometrial stem cells and curcumin-loaded PLGA nanoparticles in rat. Life Sci. 2023;330:122035. doi:10.1016/j.lfs.2023.122035
83. Wang J, Wang H, Zhu R, Liu Q, Fei J, Wang S. Anti-inflammatory activity of curcumin-loaded solid lipid nanoparticles in IL-1β transgenic mice subjected to the lipopolysaccharide-induced sepsis. Biomaterials. 2015;53:475–483. doi:10.1016/j.biomaterials.2015.02.116
84. Bae KH, Chan KH, Autoxidation-Resistant KM. ROS-scavenging, and anti-inflammatory micellar nanoparticles self-assembled from poly(acrylic acid)–green tea catechin conjugates. ACS Macro Lett. 2022;11(7):835–840. doi:10.1021/acsmacrolett.2c00239
85. Arredouani MS, Palecanda A, Koziel H, et al. MARCO is the major binding receptor for unopsonized particles and bacteria on human alveolar macrophages1. J Immunol. 2005;175(9):6058–6064. doi:10.4049/jimmunol.175.9.6058
86. Rennick JJ, Johnston APR, Parton RG. Key principles and methods for studying the endocytosis of biological and nanoparticle therapeutics. Nature Nanotechnol. 2021;16(3):266–276. doi:10.1038/s41565-021-00858-8
87. Houchin ML, Topp EM. Chemical degradation of peptides and proteins in PLGA: a review of reactions and mechanisms. J Pharm Sci. 2008;97(7):2395–2404. doi:10.1002/jps.21176
88. Caslin HL, Abebayehu D, Pinette JA, Ryan JJ. Lactate is a metabolic mediator that shapes immune cell fate and function. Front Physiol. 2021;12:688485. doi:10.3389/fphys.2021.688485
89. Arruebo M, Valladares M, González-Fernández Á. Antibody-conjugated nanoparticles for biomedical applications. J Nanomater. 2009;2009(1):439389. doi:10.1155/2009/439389
90. Ward NL, Loyd CM, Wolfram JA, Diaconu D, Michaels CM, McCormick TS. Depletion of antigen-presenting cells by clodronate liposomes reverses the psoriatic skin phenotype in KC-Tie2 mice. Br J Dermatol. 2011;164(4):750–758. doi:10.1111/j.1365-2133.2010.10129.x
91. Toy R, Roy K. Engineering nanoparticles to overcome barriers to immunotherapy. Bioeng Transl Med. 2016;1(1):47–62. doi:10.1002/btm2.10005
92. Vivier E, Malissen B. Innate and adaptive immunity: specificities and signaling hierarchies revisited. Nat Immunol. 2005;6(1):17–21. doi:10.1038/ni1153
93. Li X, Liu W, Sun L, et al. Effects of physicochemical properties of nanomaterials on their toxicity. J Biomed Mater Res Part A. 2015;103(7):2499–2507. doi:10.1002/jbm.a.35384
94. Kuhn DA, Vanhecke D, Michen B, et al. Different endocytotic uptake mechanisms for nanoparticles in epithelial cells and macrophages. Beilstein J Nanotechnol. 2014;5:1625–1636. doi:10.3762/bjnano.5.174
95. Kanno S, Hirano S, Sakamoto T, et al. Scavenger receptor MARCO contributes to cellular internalization of exosomes by dynamin-dependent endocytosis and macropinocytosis. Sci Rep. 2020;10(1):21795. doi:10.1038/s41598-020-78464-2
96. Kim SO WK, Jeong YS, Hong JY, Cho BR, Hahn JS, Jang J. Cytotoxicity of, and innate immune response to, size-controlled polypyrrole nanoparticles in mammalian cells. Biomaterials. 2011;32:2342–2350. doi:10.1016/j.biomaterials.2010.11.080
97. Nagayama S, Ogawara K, Fukuoka Y, Higaki K, Kimura T. Time-dependent changes in opsonin amount associated on nanoparticles alter their hepatic uptake characteristics. Int J Pharm. 2007;342(1–2):215–221. doi:10.1016/j.ijpharm.2007.04.036
98. Berrecoso G, Crecente-Campo J, Alonso MJ. Unveiling the pitfalls of the protein Corona of polymeric drug nanocarriers. Drug Delivery Transl Res. 2020;10(3):730–750. doi:10.1007/s13346-020-00745-0
99. Alexis F, Pridgen E, Molnar LK, Farokhzad OC. Factors affecting the clearance and biodistribution of polymeric nanoparticles. Mol Pharm. 2008;5(4):505–515. doi:10.1021/mp800051m
100. Clogston JD, Patri AK. Zeta potential measurement. Methods Mol Biol. 2011;697:63–70. doi:10.1007/978-1-60327-198-1_6
101. Casey LM, Kakade S, Decker JT, et al. Cargo-less nanoparticles program innate immune cell responses to toll-like receptor activation. Biomaterials. 2019;218:119333. doi:10.1016/j.biomaterials.2019.119333
102. Miyazawa T, Itaya M, Burdeos GC, Nakagawa K, Miyazawa T. A critical review of the use of surfactant-coated nanoparticles in nanomedicine and food nanotechnology. Int J Nanomed. 2021;16:3937–3999. doi:10.2147/IJN.S298606
103. Fromen CA, Robbins GR, Shen TW, Kai MP, Ting JPY, DeSimone JM. Controlled analysis of nanoparticle charge on mucosal and systemic antibody responses following pulmonary immunization. Proc Natl Acad Sci U S A. 2015;112(2):488–493. doi:10.1073/pnas.1422923112
104. Bonner DK, Zhao X, Buss H, Langer R, Hammond PT. Crosslinked linear polyethylenimine enhances delivery of DNA to the cytoplasm. J Control Release. 2013;167(1):101–107. doi:10.1016/j.jconrel.2012.09.004
105. Fytianos K, Chortarea S, Rodriguez-Lorenzo L, et al. Aerosol delivery of functionalized gold nanoparticles target and activate dendritic cells in a 3D lung cellular model. ACS Nano. 2017;11(1):375–383. doi:10.1021/acsnano.6b06061
106. Xu Y, Sherwood JA, Lackey KH, Qin Y, Bao Y. The responses of immune cells to iron oxide nanoparticles. J Appl Toxicol. 2016;36(4):543–553. doi:10.1002/jat.3282
107. Bhattacharjee S, Ershov D, Gucht J, et al. Surface charge-specific cytotoxicity and cellular uptake of tri-block copolymer nanoparticles. Nanotoxicology. 2013;7(1):71–84. doi:10.3109/17435390.2011.633714
108. Wang C-F, Mäkilä EM, Bonduelle C, et al. Functionalization of alkyne-terminated thermally hydrocarbonized porous silicon nanoparticles with targeting peptides and antifouling polymers: effect on the human plasma protein adsorption. ACS Appl Mater Interfaces. 2015;7(3):2006–2015. doi:10.1021/am507827n
109. Gossmann R, Fahrländer E, Hummel M, Mulac D, Brockmeyer J, Langer K. Comparative examination of adsorption of serum proteins on HSA- and PLGA-based nanoparticles using SDS–PAGE and LC–MS. Eur J Pharm Biopharm. 2015;93:80–87. doi:10.1016/j.ejpb.2015.03.021
110. Lim YW, Tan WS, Ho KL, et al. Challenges and complications of Poly(lactic-co-glycolic acid)-based long-acting drug product development. Pharmaceutics. 2022;14(3):614. doi:10.3390/pharmaceutics14030614
111. Jia Z, Li W. Nanosystems-enabled regenerative strategies for spinal cord Injury: recent advances and future prospects. Mater Des. 2023;2023:112617.
112. Yun X, Maximov VD, Yu J, Vertegel AA, Kindy MS, Kindy MS. Nanoparticles for targeted delivery of antioxidant enzymes to the brain after cerebral ischemia and reperfusion injury. J Cereb Blood Flow Metab. 2013;33(4):583–592. doi:10.1038/jcbfm.2012.209
113. Wu J, Jiang H, Bi Q, et al. Apamin-mediated actively targeted drug delivery for treatment of spinal cord injury: more than just a concept. Mol Pharmaceut. 2014;11(9):3210–3222. doi:10.1021/mp500393m
114. Surnar B, Basu U, Banik B, et al. Nanotechnology-mediated crossing of two impermeable membranes to modulate the stars of the neurovascular unit for neuroprotection. Proc Natl Acad Sci. 2018;115(52):E12333–E12342. doi:10.1073/pnas.1816429115
115. Kolhar P, Anselmo AC, Gupta V, et al. Using shape effects to target antibody-coated nanoparticles to lung and brain endothelium. Proc Natl Acad Sci U S A. 2013;110(26):10753–10758. doi:10.1073/pnas.1308345110
116. Safari H, Kelley WJ, Saito E, et al. Neutrophils preferentially phagocytose elongated particles—an opportunity for selective targeting in acute inflammatory diseases. Sci Adv. 2020;6(24):eaba1474. doi:10.1126/sciadv.aba1474
117. Makadia HK, Siegel SJ. Poly lactic-co-glycolic acid (PLGA) as biodegradable controlled drug delivery carrier. Polymers. 2011;3(3):1377–1397. doi:10.3390/polym3031377
118. Park TG. Degradation of poly(lactic-co-glycolic acid) microspheres: effect of copolymer composition. Biomaterials. 1995;16(15):1123–1130. doi:10.1016/0142-9612(95)93575-x
119. Mohammad AK, Reineke JJ. Quantitative detection of PLGA nanoparticle degradation in tissues following intravenous administration. Mol Pharm. 2013;10(6):2183–2189. doi:10.1021/mp300559v
120. Gunatillake PA, Adhikari R, Gadegaard N. Biodegradable synthetic polymers for tissue engineering. Eur Cell Mater. 2003;5(1):1–16. doi:10.22203/eCM.v005a01
121. Zhong Y, Bellamkonda RV. Biomaterials for the central nervous system. J Royal Soc Interface. 2008;5(26):957–975. doi:10.1098/rsif.2008.0071
122. Pulingam T, Appaturi JN, Parumasivam T, Ahmad A, Sudesh K. Biomedical applications of polyhydroxyalkanoate in tissue engineering. Polymers. 2022;14(11):2141. doi:10.3390/polym14112141
123. Xu X-Y, Li X-T, Peng S-W, et al. The behaviour of neural stem cells on polyhydroxyalkanoate nanofiber scaffolds. Biomaterials. 2010;31(14):3967–3975. doi:10.1016/j.biomaterials.2010.01.132
124. Novikova LN, Pettersson J, Brohlin M, Wiberg M, Novikov LN. Biodegradable poly-β-hydroxybutyrate scaffold seeded with Schwann cells to promote spinal cord repair. Biomaterials. 2008;29(9):1198–1206. doi:10.1016/j.biomaterials.2007.11.033
125. Zhao T, Xu K, Wu Q, et al. Duraplasty of PHBV/PLA/Col membranes promotes axonal regeneration by inhibiting NLRP3 complex and M1 macrophage polarization in rats with spinal cord injury. FASEB J. 2020;34(9):12147–12162. doi:10.1096/fj.202000190RR
126. Qian J, Zhu W, Lu M, Ni B, Yang J. D‐β‐hydroxybutyrate promotes functional recovery and relieves pain hypersensitivity in mice with spinal cord injury. Br J Pharmacol. 2017;174(13):1961–1971. doi:10.1111/bph.13788
127. Kong G, Liu J, Li R, et al. Ketone metabolite β-hydroxybutyrate ameliorates inflammation after spinal cord injury by inhibiting the NLRP3 inflammasome. Neurochem Res. 2021;46:213–229. doi:10.1007/s11064-020-03156-2
128. Gerardy-Schahn R, Delannoy P, von Itzstein M, Hildebrandt H, Dityatev A. Polysialic acid in brain development and synaptic plasticity. SialoGlyco Chem Biol I. 2015;2015:55–96.
129. Babu P. Glycans in regeneration. ACS Chem Biol. 2014;9(1):96–104. doi:10.1021/cb400784j
130. Schnaar RL, Gerardy-Schahn R, Hildebrandt H. Sialic acids in the brain: gangliosides and polysialic acid in nervous system development, stability, disease, and regeneration. Physiol Rev. 2014;94(2):461–518. doi:10.1152/physrev.00033.2013
131. Rutishauser U. Polysialic acid in the plasticity of the developing and adult vertebrate nervous system. Nat Rev Neurosci. 2008;9(1):26–35. doi:10.1038/nrn2285
132. Wang X-J, Peng C-H, Zhang S, et al. Polysialic-acid-based micelles promote neural regeneration in spinal cord injury therapy. Nano Lett. 2019;19(2):829–838. doi:10.1021/acs.nanolett.8b04020
133. Zhang S, Wang X-J, Li W-S, et al. Polycaprolactone/polysialic acid hybrid, multifunctional nanofiber scaffolds for treatment of spinal cord injury. Acta Biomater. 2018;77:15–27. doi:10.1016/j.actbio.2018.06.038
134. Gretenkort L, Thiesler H, Hildebrandt H. Neuroimmunomodulatory properties of polysialic acid. Glycoconjugate J. 2023;40(3):277–294. doi:10.1007/s10719-023-10120-z
135. Wada M, Nishimura Y, Watanabe Y, Takita T, Innami S. Accelerating effect of chitosan intake on urinary calcium excretion by rats. Biosci Biotechnol Biochem. 1997;61(7):1206–1208. doi:10.1271/bbb.61.1206
136. Kim M-S, Sung M-J, Seo S-B, Yoo S-J, Lim W-K, Kim H-M. Water-soluble chitosan inhibits the production of pro-inflammatory cytokine in human astrocytoma cells activated by amyloid β peptide and interleukin-1β. Neurosci lett. 2002;321(1–2):105–109. doi:10.1016/S0304-3940(02)00066-6
137. Khodagholi F, Eftekharzadeh B, Maghsoudi N, Rezaei PF. Chitosan prevents oxidative stress-induced amyloid [beta] formation and cytotoxicity in NT2 neurons: involvement of transcription factors Nrf2 and NF-[kappa] B. Mol Cell Biochem. 2010;337(1–2):39. doi:10.1007/s11010-009-0284-1
138. Cho Y, Shi R, Borgens RB. Chitosan produces potent neuroprotection and physiological recovery following traumatic spinal cord injury. J Exp Biol. 2010;213(9):1513–1520. doi:10.1242/jeb.035162
139. Zhou S, Yang Y, Gu X, Ding F. Chitooligosaccharides protect cultured hippocampal neurons against glutamate-induced neurotoxicity. Neurosci lett. 2008;444(3):270–274. doi:10.1016/j.neulet.2008.08.040
140. Cho Y, Shi R, Ben Borgens R. Chitosan nanoparticle-based neuronal membrane sealing and neuroprotection following acrolein-induced cell injury. J Biol Eng. 2010;4:1–11. doi:10.1186/1754-1611-4-2
141. Chen B, Bohnert D, Borgens RB, Cho Y. Pushing the science forward: chitosan nanoparticles and functional repair of CNS tissue after spinal cord injury. J Biol Eng. 2013;7(1):1–9. doi:10.1186/1754-1611-7-15
142. Pangestuti R, Kim S-K. Neuroprotective properties of chitosan and its derivatives. Mar Drugs. 2010;8(7):2117–2128. doi:10.3390/md8072117
143. Na JH, Lee S-Y, Lee S, et al. Effect of the stability and deformability of self-assembled glycol chitosan nanoparticles on tumor-targeting efficiency. J Control Release. 2012;163(1):2–9. doi:10.1016/j.jconrel.2012.07.028
144. Wu W, Lee S-Y, Wu X, et al. Neuroprotective ferulic acid (FA)–glycol chitosan (GC) nanoparticles for functional restoration of traumatically injured spinal cord. Biomaterials. 2014;35(7):2355–2364. doi:10.1016/j.biomaterials.2013.11.074
145. Lapcık L, Lapcık L, De SS, Demeester J, Chabrecek P. Hyaluronan: preparation, structure, properties, and applications. Chem Rev. 1998;98(8):2663–2684. doi:10.1021/cr941199z
146. Chen C, Sun X, Yang Q, Ma X. Hyaluronic acid-coated nanoparticles for the localized delivery of methylprednisolone to the injured spinal cord. J Nanomater. 2021;2021:1–9.
147. Gwak S-J, Jung JK, An SS, et al. Chitosan/TPP-hyaluronic acid nanoparticles: a new vehicle for gene delivery to the spinal cord. J biomater sci Poly ed. 2012;23(11):1437–1450. doi:10.1163/092050611X584090
148. Nakamura K, Yokohama S, Yoneda M, et al. High, but not low, molecular weight hyaluronan prevents T-cell-mediated liver injury by reducing proinflammatory cytokines in mice. J Gastroenterol. 2004;39:346–354. doi:10.1007/s00535-003-1301-x
149. Rayahin JE, Buhrman JS, Zhang Y, Koh TJ, Gemeinhart RA. High and low molecular weight hyaluronic acid differentially influence macrophage activation. ACS Biomater Sci Eng. 2015;1(7):481–493. doi:10.1021/acsbiomaterials.5b00181
150. Khaing ZZ, Milman BD, Vanscoy JE, Seidlits SK, Grill RJ, Schmidt CE. High molecular weight hyaluronic acid limits astrocyte activation and scar formation after spinal cord injury. J Neural Eng. 2011;8(4):046033. doi:10.1088/1741-2560/8/4/046033
 © 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms.php
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms.php
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.