")
Back to Journals » Drug Design, Development and Therapy » Volume 19
Recent Update of Natural Compounds as HIF-1α Inhibitors in Colorectal Carcinoma
Authors Pandey P, Lakhanpal S, Mahmood D, Baldaniya L, Kang HN, Hwang S, Kang S, Choi M, Moon S, Pandey S, Chaudhary K, Khan F , Kim B
Received 10 December 2024
Accepted for publication 8 February 2025
Published 18 March 2025 Volume 2025:19 Pages 2017—2034
DOI https://doi.org/10.2147/DDDT.S511406
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Solomon Tadesse Zeleke
Pratibha Pandey,1,2 Sorabh Lakhanpal,3 Danish Mahmood,4 Lalji Baldaniya,5 Han Na Kang,6 Sungho Hwang,7 Sojin Kang,7 Min Choi,7 Seungjoon Moon,7 Shivam Pandey,8 Kamlesh Chaudhary,9 Fahad Khan,10 Bonglee Kim6,7
1Centre for Research Impact & Outcome, Chitkara University Institute of Engineering and Technology, Chitkara University, Rajpura, Punjab, 140401, India; 2Chitkara Centre for Research and Development, Chitkara University, Baddi, Himanchal Pradesh, 174103, India; 3School of Pharmaceutical Sciences, Lovely Professional University, Phagwara, Punjab, India; 4Department of Pharmacology and Toxicology, College of Pharmacy, Qassim University, Buraydah, 51452, Saudi Arabia; 5Marwadi University Research Center, Department of Pharmaceutical Sciences, Faculty of Health Sciences, Marwadi University, Rajkot, Gujarat, 360003, India; 6KM Convergence Research Division, Korea Institute of Oriental Medicine, Daejeon, Republic of Korea; 7Department of Pathology, College of Korean Medicine, Kyung Hee University, Hoegidong Dongdaemungu, Seoul, 05253, Republic of Korea; 8School of Applied and Life Sciences, Uttaranchal University, Dehradun, Uttarakhand, 248007, India; 9Department of Neurology, National Institute of Medical Sciences, NIMS University Rajasthan, Jaipur, Rajasthan, India; 10Center for Global Health Research, Saveetha Medical College, Saveetha Institute of Medical and Technical Sciences, Chennai, Tamil Nadu, India
Correspondence: Fahad Khan; Bonglee Kim, Email [email protected]; [email protected]
Abstract: Hypoxia-inducible factor (HIF)-1 is a transcription factor that regulates the expression of target genes associated with oxygen homeostasis under hypoxic conditions, thereby contributing to tumor development and progression. Accumulating evidence has demonstrated that HIF-1α mediates different biological processes, including tumor angiogenesis, metastasis, metabolism, and immune evasion. Thus, overexpression of HIF-1α is strongly associated with poor prognosis in cancer patients. Natural compounds are important sources of anticancer drugs and studies have emphasized the decisive role of these mediators in modulating HIF-1α. Therefore, the pharmacological targeting of HIF-1α has emerged as a novel cancer therapeutic approach in recent years. The novelty of this review is that it summarizes natural products targeting HIF-1α in colorectal cancer that have not been presented earlier. We studied research publications related to the HIF-1α domain in cancer from 2010 to 2024. However, our main focus was to identify a better targeted approach for colorectal carcinoma management. Our review described HIF-1α role in tumor progression, summarizes the natural compounds employed as HIF-1α inhibitors, and discusses their potential in the development of natural compounds as HIF-1α inhibitors for colorectal cancer treatment.
Keywords: HIF-1α, colorectal cancer, natural compounds, inhibitors, hypoxia
Introduction
A significant advancement in cancer treatment over the last decade has been the emergence of targeted therapies. Unlike traditional chemotherapy, which indiscriminately kills cells, targeted therapy focuses on blocking specific proteins and genes.1,2 Targeted therapy is an innovative and elective strategy that effectively extends the overall lifespan of patients with colorectal cancer (CRC). New drugs that target many key pathways and immunological checkpoints have appeared rapidly after the successful use of anti-angiogenesis agents (bevacizumab) and anti-EGFR agents (cetuximab). Targeted medicines can effectively impede the growth, specialization, and movement of cancer cells. Targeted medications can modify the TME, including nearby blood arteries and immune cells, to hinder tumor growth and enhance immune surveillance and attack.3,4 Tumor hypoxia is a prevalent medical condition that occurs when there is insufficient oxygen to reach tissues.5 This leads to changes in metabolism and an increased ability of cancer cells to spread to other parts of the body.6 Cancer cells have the ability to regulate their growth and metabolism in hypoxic environments by altering the tumor microenvironment (TME) in response to hypoxia.7,8 These hypoxic responses are predominantly affected by Hypoxia-inducible factor (HIFs) and their signaling pathways. Hypoxia, caused by insufficient oxygen delivery to tumor tissues, is a critical clinical state that leads to angiogenesis, cellular proliferation, and metastasis.9 Tumor cells exhibit increased ROS levels in hypoxic environments. ROS production initiates the degradation of cellular macromolecules including RNA, lipids, DNA, and proteins, ultimately resulting in cell death. However, cancer cells employ several strategies to evade apoptosis and ultimately persist in adverse environments.10 The primary survival characteristics of cancer cells involve an increased focus on metabolic reprogramming, angiogenesis, and metastasis. The regulation of these reactions is controlled by the HIF protein family, whose expression is modified in response to hypoxia.
HIF was initially discovered by Semenza et al, who reported the existence of a hypoxia response element in every HIFs-regulated target gene.11,12 HIF proteins have different alpha and beta subunits, which form a structure called a heterodimers. This protein has a basic helical loop. In addition to HIF-1, the HIF family encompasses two other members: HIF-2 and HIF-3. HIF-1α is a 120 kDa subunit that is sensitive to oxygen levels and is found in all tissues, whereas HIF-2α and HIF-3α have a more limited presence in certain organs. Both share a common functional domain called the basic helix-loop-helix (bHLH) motif, which is responsible for DNA-binding. Additionally, they possess PAS-A and PAS-B domains that facilitate heterodimerisation.13 Hypoxia triggers an imbalance in the production of pro- and anti-angiogenic factors, resulting in the accelerated and disordered development of blood vessels. HIF-1α controls a wide array of genes responsible for epithelial-to-mesenchymal transition. This process involves the disruption of the basement membrane in the surrounding tumor tissue, leading to invasion. Hypoxia facilitates the movement of cells from the initial tumor to a distant location where it forms a secondary tumor by controlling the activity of HIF-1α.14,15 Furthermore, modifications to these novel traits enhance the aggressiveness of cancer cells under hypoxic conditions. Consequently, they may serve as potential targets for cancer therapies. Although numerous reviews have summarized natural compounds as HIF-1α inhibitors in various human carcinomas but to the best part of our knowledge, no review has enlisted natural compounds as HIF-1α inhibitors in colorectal carcinoma. Such review would aid in developing more specific therapeutic approach for colorectal and colon cancer patient with minimal side effects. This review comprises of different sections including: (i) HIF-1α in colorectal cancer (ii) Natural compounds inhibiting HIF-1 α in colorectal cancer and (iii) Overcoming chemoresistance in colorectal carcinoma. Altogether, this review would provide a thorough understanding of the HIF-1α inhibitor in colorectal and colon cancer that has not been explained anywhere.
HIF-1α Expression in Colorectal Cancer
HIF-1α is essential for tumor growth, cellular energy production, and the spread of cancer to other body parts. HIF-1 activation is strongly influenced by the translocation of HIF-1α into the nucleus, a process that can be disrupted by the von Hippel-Lindau (VHL) protein.16 PHD2 uses O2 (as a substrate) and introduces hydroxyl functional groups to HIF-1α under normoxic conditions. This results in ubiquitination of HIF-1α after binding to the VHL protein, resulting in the breakdown of HIF-1α via the recruitment of ubiquitin ligases.17 Under hypoxic conditions, a decrease in hydroxylation results in a reduction in the synthesis of the VHL and HIF-1α complex, which in turn promotes the stable production of HIF-1α.18 Oxygen levels in animal cells are closely correlated with changes in gene expression, which in turn trigger rapid oxygenation and cellular stress response via activated HIFs. Along with its involvement in maintaining physiological cellular processes in the normal cardiovascular system and embryonic development, HIF-1α also participates in tumor angiogenesis, cancer progression, anti-apoptosis, and metastasis throughout tumor development in numerous carcinomas, including prostate, hepatocellular, lung, cervical, and gastrointestinal carcinomas.19 Mounting data indicate that the adaptation of cancerous cells to hypoxia is directly associated with poor prognosis and metastasis in numerous malignancies. Hyperoxic breathing can reverse hypoxia-induced carcinogenesis. Furthermore, significant evidence indicates the potential involvement of HIF-1α and HIF-2α in the regulation of HIF-1 transcription in the TME (under low oxygen concentrations).20 Colorectal cancer (CRC) ranks third in terms of its prevalence as a malignant tumor and second in terms of its contribution to cancer-related deaths worldwide. The prevalence of colorectal cancer has progressively increased owing to lifestyle modifications. Despite advancements in chemotherapy, the overall rate of positive responses to treatment remains unsatisfactory.21,22 The ineffectiveness of this method is ascribed to chemoresistance of malignant tumors. Similar to other solid tumors, colorectal cancer (CRC) undergoes hypoxia.23 The consensus is that the existence of hypoxia in the tumor microenvironment might promote tumor proliferation and metastasis, as well as increase their resistance to pharmaceutical interventions.24
HIF-1α plays a role in controlling the activity of over 70 specific genes involved in different aspects of tumor growth. Four specific types of genes are strongly associated with tumors. These genes are associated with glucose transport and glycolysis, angiogenesis, tumor growth, cell death, and tumor spread and invasion. They elicit a cascade of reactions in tissues and cells to acclimate to an oxygen-deprived atmosphere, facilitating the growth of blood vessels in tumors and enhancing the ability of the tumor to invade surrounding tissues, along with its resistance to radiation and chemotherapy.25,26 A significant discovery in understanding how hypoxia contributes to drug resistance is the identification of HIF-1α as a key activator of the multidrug resistance gene 1 (MDR1). This gene encodes for P-glycoprotein (P-gp), which functions as a drug efflux pump and reduces the concentration of drugs in cells.27–29 Therefore, it is crucial to discover innovative methods to inhibit HIF-1α activation and MDR1 expression in cancer cells exposed to low oxygen levels.
HIF-1α in tumor cells not only stimulates angiogenesis but also facilitates the acquisition of energy through the reconfiguration of cellular metabolic pathways, with glycolysis being a crucial mechanism.30 Initially, HIF-1α activates lactate dehydrogenase A (LDHA) and glucose transporters to facilitate the transformation of the metabolic pathway from oxidative to non-oxidative carbon metabolism as well as ATP-producing pathways (glycolysis).31 Furthermore, the PDK protein, which is produced by the pyruvate dehydrogenase kinase-1 (PDK1) gene, acts as a target of HIF-1α. This hinders the synthesis of acetyl-CoA, obstructs the tricarboxylic acid cycle (TCA), and reduces the ingestion of oxygen.32 Cells deficient in HIF-1α were found to have decreased ATP synthesis in low-oxygen settings, resulting in increased ROS production and the facilitation of programmed cell death. Furthermore, activated HIF-1α affects the pentose phosphate pathway by converting the intermediate products of glycolysis into 5-phosphoribose, which is a crucial substance used in the production of nucleotides.33,34 These findings indicate that HIF-1α enhances cell survival under low-oxygen conditions by reorganizing the cellular metabolic pathways. This reorganization is necessary to transform glucose metabolism into DNA and RNA, which are crucial for the survival and proliferation of tumor cells in low-oxygen environments.35
HIF-1α additionally enhances the growth of cancer cells by controlling associated factors. For instance, HIF-1α stimulates the synthesis of substances, including transforming factor-2 (TGF-2) and insulin-like growth factor-2 (IGF-2), which further stimulate the MAPK/PI3K signaling pathways by binding to specific receptors. This leads to cell hyperplasia, which enhances HIF-1α activation and accelerates the transcriptional activity of HIF-1α-induced genes that are crucial in tumors. HIF-1α stabilizes p53 by preventing p53 from being broken down and preventing p53 from moving out of the nucleus. This leads to the activation of various genes involved in programmed cell death, and promotes apoptosis.36 Transcriptional initiation is a crucial regulatory process that affects HIF-1α mRNA expression. The activation of the HIF-1α gene locus is triggered by the interaction between transcription factors, such as STAT3 and NF-κB, and their promoter regions [Figure 1]. Owing to the excessive expression of HIF-1α in various types of human malignancies and their metastases, targeting the HIF-1 pathway shows potential as a viable strategy for cancer treatment.37 Researchers argue that inhibiting the hypoxic response within solid tumors increases their vulnerability to radiotherapy and chemotherapy, leading to better patient outcomes.
 |
Figure 1 The role of HIF1 alpha in drug resistance mechanisms of colorectal cancer. Under hypoxia, HIF-1α accumulates leading to modulated metabolic pathways and angiogenesis induction which further potentiate cancer initiation and metastasis. |
Natural Compounds Inhibiting HIF-1 α in Colorectal Cancer
Several natural substances are under consideration for the research and development of novel, secure, and dependable chemicals for cancer prevention of cancer.38 There is substantial circumstantial evidence suggesting that consuming relatively large amounts of fruits and plant-based foods is associated with a decreased risk of colorectal cancer (CRC). Previous reports have indicated that these natural compounds can affect gene expression and the resulting products via flexible modulatory signaling pathways. This enables them to regulate various cellular processes, including migration, survival, the cell cycle, differentiation, and apoptosis.39–41 Furthermore, there is a higher occurrence of excessive production of pro-inflammatory transcription factors and cytokines, which are significant contributors to the risk of colitis and development of CRC. This overproduction is particularly observed in the colon due to repeated inflammation.42 However, the anti-inflammatory properties of various phytochemicals have been shown to inhibit this process, thereby preventing CRC onset. Researchers have discovered several methods to primarily downregulate HIF-1α, including inhibition of HIF-1α/p300 connections, reduction of HIF-1α mRNA production, and decrease in HIF-1α protein stability. Various natural medications and TCM prescriptions effectively reduce HIF-1α levels, leading to a therapeutic impact in modern research.43,44 Therefore, certain compounds reduced HIF-1 transcription by (i) preventing the phosphorylation of HIF-1α, which affects its movement from the cytoplasm to the nucleus, (ii) inhibiting the interaction between Hsp90 and HIF-1α, (iii) interfering with the mitochondrial ETC, (iv)inhibiting protein synthesis, and (v) destabilizing proteins [Figure 2]. Multiple mechanisms are associated with the downregulation of HIF-1 transcriptional activity by some flavonoids such as apigenin, chrysin, and 4′-O-methylalpinumisoflavone. Consequently, angiogenesis is suppressed, leading to an increased susceptibility of tumor cells to radiation and chemotherapy. The phosphorylation of HIF-1α is regulated by the p42/p44 MAPK and PI3K/Akt signaling pathways. Some flavonoids such as quercetin, luteolin, and kaempferol can interfere with the MAPK pathway. This interference leads to a decrease in the nuclear concentration of HIF-1α, subsequently affecting its transcriptional activity.45–48 Thus, different classes of natural compounds targeting HIF-1α in colorectal carcinoma are summarized in the subsequent section in more detail [Figure 3].
 |
Figure 2 Natural compounds targeting HIF-1α in colorectal cancer via inhibiting the mRNA expression, protein synthesis, protein stabilization, DNA binding and transactivation of HIF-1α in colorectal cancer. |
 |
Figure 3 Representation of chemical structure of various natural compounds as HIF-1α modulators. |
Polyphenols Targeting HIF-1α
The term polyphenol lacks a precise definition; however, it is generally accepted that polyphenols are naturally occurring compounds with hydroxyl groups (one or more) attached to the aromatic rings. Polyphenols can be classified into four main categories: phenolic acids, flavonoids, stilbenes, and lignans. The effect of flavonoids on HIF-1α activation varies significantly depending on the specific methods through which flavonoids disrupted the HIF-1 pathway.49 Flavonoids function as iron chelators because of their keto and hydroxyl functional groups. Flavonoids, such as (-)-epicatechin-3-gallate, quercetin, fisetin, baicalein, galangin, and luteolin, bind to iron inside cells, which then affects the function of iron-dependent enzymes such as PHDs. Therefore, PHD activity is hindered, resulting in HIF-1α stabilization under normoxic conditions. Flavonoids imitate hypoxia and trigger adaptive cellular responses. However, flavonoids disrupt the HIF-1 pathway regardless of their ability to chelate iron. Vanillic acid (a dietary phenolic compound) suppresses HIF-1α expression in HCT116 cells via inhibition of the MEK/ERK and mTOR cellular pathways, and has emerged as an efficient HIF-1α inhibitor in colon cancer.49
Quercetin
The anticancer potential of quercetin has been exploited to address hypoxia. Pharmacological processes have been explored by focusing on AMP-activated protein kinase (AMPK).50 Quercetin effectively reduced tumor growth in both vivo and vitro models of HCT116 cancer xenografts by drastically decreasing AMPK activation. Quercetin significantly increased the occurrence of programmed cell death (apoptosis) in HCT116 cancer cells exposed to low-oxygen conditions (hypoxia) compared to those exposed to normal oxygen levels (normoxia). This effect is strongly linked to suppression of hypoxia-induced AMPK activity.51 Quercetin decreases HIF-1α activation, which is a transcription factor responsible for the adaptive cellular response to low oxygen levels.52 Quercetin significantly reduced angiogenesis generated by HT-29 cells in Human umbilical vein endothelial cells (HUVECs). Quercetin also suppresses the expression of VEGF-A, NF-κB, and p65. Furthermore, quercetin effectively suppressed the expression and movement of VEGFR-2 in human vein endothelial cells (HUVECs) when they were exposed to high concentrations of quercetin-treated HT-29 cells. Collectively, quercetin inhibited the formation of new blood vessels (angiogenesis) by blocking VEGF-A activity through the NF-κB signaling pathway in HT-29 cells. Additionally, it decreased the expression and movement of VEGFR-2 in HUVECs.53
Luteolin
Luteolin, a flavonoid, exhibited significant cytotoxicity against both HCT116 and MDA-MB231 cell lines. It induces apoptosis (cell death) owing to injury (necrosis), and this outcome is not influenced by the activation of HIF-1α. Luteolin induced autophagy and inhibited HIF-1α transcriptional activity in these cells, which was accompanied by a reduction in the number of protein markers linked to stem cell traits and invasion as well as a decrease in the ability of cells to migrate. Thus, luteolin has emerged as a promising therapeutic agent for hypoxic cancers.54 Luteolin has a notable ability causes cell death in HCT116 and MDA-MB231 cells. This was achieved by triggering apoptotic and necrotic cell death. Importantly, the activation of HIF-1α does not affect this effect. Luteolin has been utilized as an effective therapeutic drug to target hypoxic malignancies.55 Compound 7f, a derivative of the 5,6,7-Trimethoxy flavonoid salicylate, efficiently reduced the expression of glycolysis-related proteins (PKM2, HIF-1α, and PFKM) and tumor angiogenesis-related protein (VEGF).56
Hispidulin
Hispidulin (a naturally occurring flavonoid) inhibits hypoxia-induced EMT (epithelial-mesenchymal transition) in human colorectal cancer cells, which is necessary for cancer metastasis.57 Hispidulin administration effectively reversed the EMT-related phenotype, including hypoxia-induced cell migration, morphological alterations, and E-cadherin downregulation. This impact was partially achieved by the suppression of HIF-1α (at both the mRNA and protein levels) through the regulation of the PTEN/PI3K/Akt pathway. Xie et al (2015) found that hispidulin effectively inhibits hypoxia-induced EMT in human colon cancer cells.57
Isorhamnetin
Isorhamnetin is a significant monomethoxyflavonol present in the G. biloba leaf extract. Isorhamnetin strongly inhibited CoCl2- and HIF-1α- induced activity of the hypoxic response element reporter gene, as well as HIF-1α-dependent gene transcription, including carbonic anhydrase-IX, glucose transporter 1, pyruvate dehydrogenase kinase 1, and lactate dehydrogenase A, in HCT116 and HT29 cells. The antioxidant properties of isorhamnetin were verified by measuring the ROS generation using CoCl2 or H2O2. In HEK293 cells, the overexpression of HIF-1α was consistently reduced by N-acetyl-l-cysteine and isorhamnetin. Isorhamnetin hinders the buildup of HIF-1α through the inhibition of ROS, thereby enhancing its effectiveness in preventing metastasis.58
Piceatannol
Piceatannol is a naturally occurring polyphenol with anticancer and anti-inflammatory activities.59 PIC–BSA nanoparticles (where PIC was encapsulated within bovine serum albumin) showed greater efficacy in reducing the expression of nuclear HIF-1α and p65 in colon cancer cells than free PIC. A notable decrease in inflammation caused by chemical colitis in mice treated with PIC–BSA NPs. In addition, the mouse model of colorectal cancer (colitis-associated) showed a notable decrease in both the number and size of colon tumors when treated with PIC–BSA NPs compared to free PIC. These findings suggested that the therapeutic potential of PIC is enhanced when they are formed as PIC-BSA NPs. These findings have paved the way for the development of a novel range of secure and efficient treatments for individuals with cancer.60
Other Phenolic Compounds
Hypoxia-induced medication resistance poses a significant challenge for the advancement of effective cancer treatments.61 Wogonin (O-methylated flavone) effectively counteracted the hypoxic resistance of HCT116 cells by reducing glycolysis and HIF-1α activity. This was achieved by suppression of the PI3K/Akt pathway. Thus, wogonin is a potential candidate for the development of new medicines to reverse multidrug resistance (MDR). The mechanism of reversal is likely to be inhibition of HIF-1α production by blocking the PI3K/Akt signaling pathway.62 Oleuropein (OL) is the predominant phenolic component found in the fruits of olive trees. The growth of HT-29 cells is restricted, and programmed cell death is triggered by OL through the activation of the p53 pathway, which modifies the response of HIF-1α to low oxygen levels.63
Calebin A (CA), a natural polyphenol extracted from turmeric, is known for its ability to inhibit cancer growth by regulating the NF-κB pathway. CA partially hindered the migration of HCT-116 cells by suppressing HIF-1α, a crucial factor for the survival of CRC cells. It also suppresses HIF-1α activation and metastatic biomarker expression initiated by the tumor microenvironment while promoting apoptosis through caspase-3 activation.64 Parthenolide (PT) can be extracted from Mexican Indian medicinal herbs and exhibits anti-inflammatory characteristics. Parthenolide inhibits HIF-1α signalling and hypoxia-induced EMT in colorectal cells PT suppresses growth in colorectal cancer xenograft models and modulates NF-κB, HIF-1α, and EMT-specific markers in tissue samples. The results showed that PT blocked HIF‑1α signaling and prevented hypoxia-induced EMT. This suggests a new molecular mechanism by which HIF‑1α contributes to cancer metastasis.65
Traditional Chinese Medicine Targeting HIF-1α
Several malignant tumors, such as colorectal cancer, develop functioning blood arteries to facilitate tumor growth and spread to other body parts. Vasculogenic mimicry (VM) is the capacity of a vastly invasive tumor to connect with one another and create vasculature and is linked to a negative cancer prognosis.66 TCM has also been used in cancer treatment.67 Jianpi Jiedu decoction (JPJD), a long-standing traditional Chinese medicinal formula, has been used for several decades to treat colorectal cancer. JPJD substantially inhibited the viability and proliferation of colorectal cancer cell lines. Flow cytometry analysis revealed that JPJD induced apoptosis in HCT116 cells. Moreover, JPJD efficiently restricted the movement of tumor cells, their invasion, and new blood vessel formation by blocking the mTOR/HIF-1α/VEGF signaling pathway. JPJD significantly inhibited HCT116 tumor growth (in athymic nude mice) when tested in vivo. Additionally, it reduces CD34 and VEGF levels and downregulates the mTOR/HIF-1α/VEGF pathway, as reported by Peng et al in 2018.68
Zong et al (2020) found that Astragalus Atractylodes mixture (AAM), a TCM, effectively inhibited the migration and formation of vascular mimicry (VM) in colorectal cancer (HCT-116 and LoVo) cells under hypoxic conditions.69 This inhibition was achieved by the suppression of the ROS/HIF-1α/MMP2 pathway. These findings suggest the therapeutic potential of AAM in the prevention of VM in human colorectal cancer cells.69 Imperatorin triggered a significant HIF-1α protein reduction in accumulation produced by hypoxia. Imperatorin inhibited the expression of HIF-1 target genes in response to hypoxia. Flow cytometric analysis revealed that imperatorin causes G1 phase arrest. Imperatorin reduced the production of HIF-1α protein in human colon cancer cells (HCT116) by reducing the activity of the mTOR/p70S6K/4E-BP1 and MAPK pathways. This study suggested that imperatorin is an efficient inhibitor of HIF-1α in colon cancer cells.70
Evodiamine (Evo), (quinolone alkaloid) is derived from the traditional herbal remedy, Evodia rutaecarpa. Evo caused a noticeable decrease in HIF-1α activity (in vivo and vitro). Introducing HIF-1α from an external source can weaken the inhibitory effect of Evo on LoVo cell proliferation Conversely, reducing the levels of HIF-1α enhances this inhibitory effect to a significant extent. The anticancer effect of Evo in colon cancer is partially achieved by reducing the production of HIF-1α, which is triggered by deactivation of the PI3K/Akt signaling pathway by reducing IGF-1 expression in colon cancer cells.71
Ursolic acid (UA) is a pentacyclic triterpenic acid present in various plants, including Chinese medicinal herbs.72 Cancer cells are expected to exhibit increased levels of HIF-1α when exposed to low oxygen levels (hypoxic stress) or treated with chemotherapy. Normal cells rarely encounter hypoxic stress and, hence, maintain normal HIF-1α levels. UA effectively inhibited MDR1 expression by decreasing hypoxia-induced HIF-1α build-up. This discovery offers limited clarification on how UA may counteract the resistance of colon cancer cells to chemotherapy in low-oxygen environments.73,74 HIF-1α is a prominent transcription factor that plays a significant role in regulating various tumor characteristics, including the growth, spread, and formation of new blood vessels.
Alkaloid Targeting HIF-1α
Matrine
Matrine (an alkaloid) is derived from the roots of TCM (Sophora flavescens Ait. Matrine) has been shown to exhibit specific cytotoxicity towards cancerous cells.75 Matrine counteracted the Warburg effect by limiting lactate formation and glucose uptake. Matrine effectively reduced HIF-1α mRNA and protein expression Matrine significantly decreased the expression levels of LDHA GLUT1, and HK2 which are downstream targets of HIF-1α and are involved in the regulation of glucose metabolism. Furthermore, the suppressive effect of matrine was notably diminished when HIF-1α was depleted or artificially elevated in the colon cancer cells. Matrine has the potential to be used as a therapeutic drug for colon cancer by specifically targeting the HIF-1α-mediated Warburg effect, as suggested by Hong et al in 2019.76
Berberine
Hyperactivated glucose absorption and glycolytic metabolism are the characteristic features of cancer.77 Berberine, a naturally occurring alkaloid, exhibits selective anticancer effects in tumors and has been demonstrated to enhances glucose uptake in metabolic organs and cells. Berberine effectively slowed the growth of colon cancer (HCT116 and KM12C) cells. Berberine treatment significantly suppressed the protein production HIF-1α that played a crucial role in the abnormal glucose metabolism in colon cancer cell lines.77 However, no significant effect on HIF-1α mRNA expression was observed. Regulation occurs through protein synthesis rather than through protein stability. The effect of berberine was not affected by obstructing the degradation of HIF-1α protein using the hypoxia mimic desferrioxamine (DFX) or proteasome inhibitor MG132. Furthermore, berberine suppresses mTOR signaling, which has been previously shown to influence HIF-1α protein synthesis. These findings demonstrated that berberine effectively suppressed excessive glucose metabolism in colon cancer cells by inhibiting HIF-1α protein synthesis, which is dependent on the mTOR pathway. This study provides a theoretical foundation for its potential use in colon cancer treatment.
Other Natural Compounds Targeting HIF-1α (Sulforaphane, Epigallocatechin Gallate, Salternamide A, Verbascoside, Ilexgenin A, Desferrioxamine, Worenine, Sesamin, Genipin, Brusatol, Panaxadiol)
Another study examined the effect of sulforaphane, a naturally occurring substance often present in broccoli,78 on the expression of HIF-1α in HCT116 (colon cancer) and AGS (gastric cancer) cells. Sulforaphane suppressed HIF-1α protein expression in HCT116 and AGS cells when exposed to low-oxygen conditions. Furthermore, sulforaphane administration suppresses VEGF expression in HCT116 cells under hypoxic conditions.79 Inhibition of HIF-1α protein expression by sulforaphane has been linked to the instability of the protein itself and avoidance of the activation of genes targeted by HIF-1α. Sulforaphane additionally suppressed the movement of cells generated by hypoxia. These studies indicate that sulforaphane can hinder the growth of blood vessels and movement of human colon cancer cells by suppressing the production of HIF-1α and VEGF under low-oxygen conditions. Sulforaphane inhibits the latent-to-hindered advancement of colon cancer and blood vessel formation in human cancer cells by suppressing the production of HIF-1α and VEGF.80 Epigallocatechin gallate (green tea) inhibited the growth of colorectal cancer cells by blocking the activation of the VEGF/VEGFR pathway. This effect is achieved by suppressing the expression of HIF-1α and other important growth factors. Therefore, it may have potential applications in the prevention and treatment of colorectal cancer.81
Salternamide A (SA) is A newly discovered compound obtained from the halophilic Streptomyces sp. It has been shown to have strong cytotoxic properties in numerous human cancer cells. SA effectively suppressed hypoxia-induced buildup of HIF-1α in various types of human cancer cells, with the inhibition dependent on both time and concentration. Furthermore, SA inhibited the activation of the HIF-1α signaling pathway under hypoxic conditions. Moreover, SA triggers cell death in colorectal cancer cells by promoting G2/M (cell cycle) phase arrest and apoptosis.82 SA, a small-molecule HIF-1α inhibitor derived from natural marine products, has been recognized as a promising candidate for the development of anticancer medicines.
Colorectal cancer cells (specifically HT29 cells) show an increase in the expression of Rac-1 and HIF-1α, which are crucial for the initiation and advancement of cell movement, new blood vessel formation, and cancer spread to other body parts. A study by Seyfi et al (2018) found that verbascoside demonstrated notable anti-metastatic and anti-invasion effects by inhibiting Zeb-1, Rac-1, and HIF-1α signaling pathways. These findings suggest that verbascoside is a promising candidate to combat the metastatic characteristics of colon cancer.83 Ilexgenin A (IA), the primary biologically active component of Ilex hainanensis Merr., exhibits hypolipidemic properties. It has the potential to decrease the levels of triglycerides (TG) and expression of ACC, HIF1α, FASN, and SREBP-1 in colon (HT 29 and HCT 116) cancer cells. Desferrioxamine (DFX) was shown to weaken the inhibitory effect of IA on SREBP-1, indicating that HIF1α plays a role in the regulation of SREBP-1 by RA. The study conducted by Zhang et al (2019) found that IA inhibited the early development of colon cancer in mice treated with AOM/DSS. Additionally, IA partially alters lipid metabolism via the involvement of HIF1α/SREBP-1.83
HIF-1α is essential for cancer metastasis. Inhibition of CRC cell migration was observed when baicalein administration decreased HIF-1α expression. However, this effect was reversed by overexpression of TLR4. Baicalein directly interacts with TLR4 and inhibits the TLR4/HIF-1α/VEGF signaling pathway in CRC.84 In the Warburg effect, cancerous cells preferentially utilize glycolysis over oxidative phosphorylation to metabolize glucose, even when oxygen is available, resulting in the production of lactate as an end-product. The metabolic shift to aerobic glycolysis promotes tumor cell proliferation by allowing cancer cells to survive and redirect metabolic intermediates from energy production to biosynthesis. This phenomenon arises because of the ability of certain transcription factors, such as HIF-1α, to alter the expression of various isoforms of glycolytic enzymes in cancerous cells. Worenine, extracted from Coptis chinensis, was found to possess anticancer properties in SW620 and HCT116 cells. In vitro, worenine effectively suppressed cell proliferation, growth phase (cell cycle) arrest, and the Warburg effect in colorectal cancer cells by specifically targeting HIF-1α. According to Ji et al (2021), Worenine has the ability to reverse the Warburg effect and hinder the proliferation of colon cancer cells by exerting a negative regulatory effect on HIF-1α.85
Sesamin (sesame seeds) effectively suppressed the formation of new blood vessels in colorectal cancer under low-oxygen conditions in a dose-dependent manner, as observed in laboratory experiments. Furthermore, ingestion of sesamin in nude mice significantly inhibited the development of new blood vessels in matrigel plugs containing CRC cells. Sesamin decreases VEGFA expression, hindering new blood vessel formation in hypoxia-induced colorectal cancer produced by hypoxia.86 Furthermore, sesamin prevented the phosphorylation of IκBα, thereby preventing the activation of NF-κB p65 and the transcription of HIF-1α under hypoxic conditions. These findings demonstrated that sesamin effectively suppressed new blood vessel formation in colorectal, driven by low oxygen levels through the NF-κB/HIF-1α/VEGFA signaling pathway. These findings serve as a theoretical and experimental foundation for utilizing sesamin in CRC prevention and treatment, as suggested by Huang et al (2022).87 Genipin inhibits HIF-1α buildup in different cancer cells, such as HCT116, under low-oxygen conditions by regulating the breakdown of proteins. Genipin inhibits VEGF expression and the invasion of colon cancer cells by obstructing the extracellular signal-regulated kinase signaling pathway.88 Brusatol (Bru, a quassinoid), derived from Brucea Esters, suppresses the HIF-1 signaling pathway and inhibits HIF-1 transactivation activity and the production of its target genes, including LDHA, VEGF, HK2, and GLUT1, under hypoxic conditions. Furthermore, Bru reduced glucose intake under low-oxygen conditions by inhibiting the HIF-1α signaling pathway. Subsequent research has indicated that the suppressive effect of Bru on the HIF-1 signaling pathway could be ascribed to its ability to enhance the degradation of HIF-1α.89 Remarkably, ROS levels within the cells and mitochondria were reduced by BRU therapy, suggesting the involvement of mitochondrial ROS regulation in the mechanism of action of Bru. The collective findings present strong evidence supporting the control of HIF-1α by Bru, and indicate its potential as a therapeutic approach for colon cancer.90
Another study examined whether l-carnosine targets HIF-1α in HCT-116 cells. HIF-1α is highly expressed in several human cancers and is the primary factor responsible for drug and radiation resistance in solid tumors. l-carnosine was found to lower the protein level of HIF-1α, thereby reducing its stability and inhibiting HIF-1 transcriptional activity. Furthermore, L-carnosine plays a role in the UPS, which aids HIF-1α degradation of.91 Panaxadiol inhibited hypoxia-induced HIF-1α production by acting on the PI3K and mitogen-activated protein kinase pathways. Panaxadiol hindered the activation of STAT3 via JAK1, JAK2, and Src pathways. Furthermore, prior administration of panaxadiol enhanced the efficacy of cytotoxic T lymphocytes and restored their ability to eliminate tumor cells in a co-culture system including T cells and tumor cells. Immunoprecipitation demonstrated that panaxadiol hindered the production of PD-L1 by obstructing the interaction between HIF-1α and STAT3. This study demonstrates the antitumor properties of panaxadiol and provides valuable information on the potential use of PD-L1 inhibition in cancer treatment.92 Table 1 summarizes all reported phytocompounds that target HIF-1α in both colorectal and colon cancer cells.
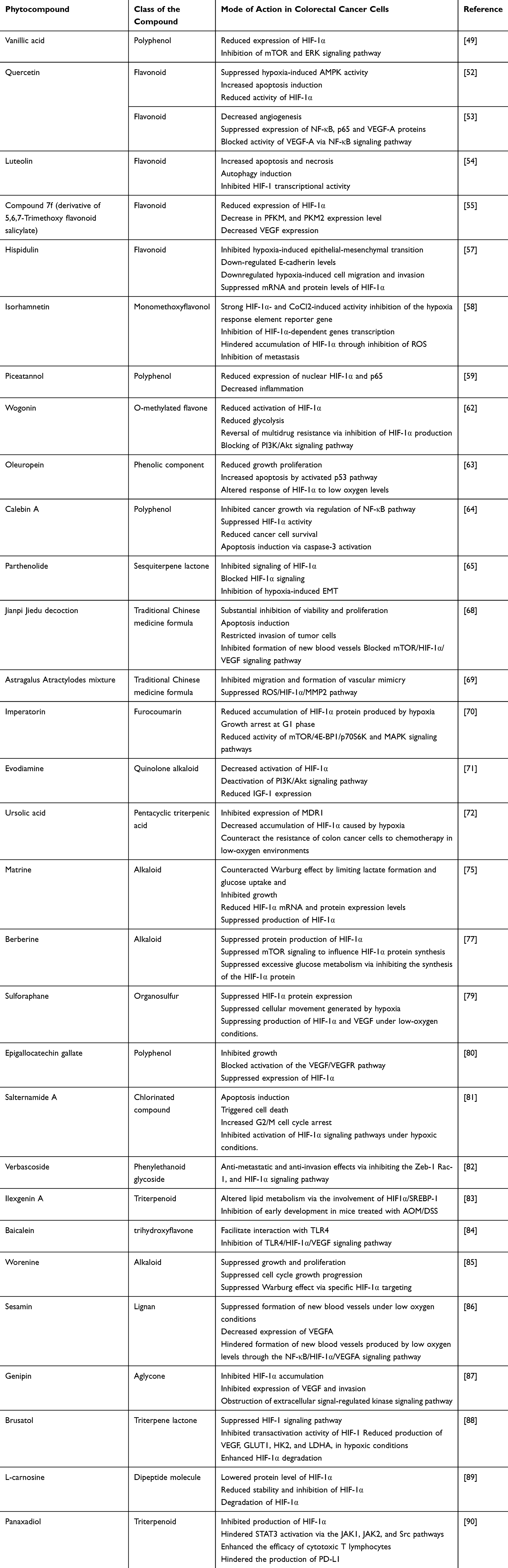 |
Table 1 Phytocompounds Targeting HIF-1α in Both Colorectal and Colon Cancerous Cells |
Overcoming Chemoresistance in Colorectal Carcinoma
Hypoxia during carcinogenesis promotes aggressive tumor development and resistance to radiation and chemotherapy and is associated with poor prognosis. The effectiveness of chemotherapy in cancer treatment is limited by drug resistance caused by the hypoxia-induced stabilization of HIF-1α. HIF-1α overexpression has been detected in a wide range of human carcinomas, including colon and colorectal cancer.91 Therefore, focusing on HIF-1α is a promising approach for overcoming chemoresistance and improving the efficacy of chemotherapy for CRC. The frequent emergence of resistance to the chemotherapeutic medication 5-fluorouracil (5-FU) poses challenges for the treatment of advanced colorectal cancer. Resveratrol uses β1-integrin receptors to overcome 5-FU chemoresistance in CRC cells. The β1-integrin/HIF-1α signaling pathway plays a crucial role in enhancing the sensitivity of CRC cells to 5-FU and overcoming resistance to chemotherapy. Resveratrol activates this pathway, highlighting its potential as a supportive treatment for CRC. Sanguisorba officinalis DiYu (DY) has traditionally been used in China for cancer therapeutics because it has the potential to enhance the immune system and reduce hematological toxicity. DY hinders the growth and movement of 5-FU-resistant cells and enhances their responsiveness to 5-FU. These results suggest that DY is a promising medication for therapeutic management and a supplementary therapy for drug-resistant CRC.92
Dihydromyricetin, a flavonoid compound derived from Hovenia dulcis (Japanese raisin tree), suppressed the expression of MRP2 and its promoter activity by inhibiting the NF-κB-Nrf2 signaling pathway in HCT-8/VCR and HCT116/L-OHP cells. This contributes to the reversal of L-OHP/VCR resistance in colorectal cancerous cells.93 Curcumin, a lipid-soluble polyphenol, effectively inhibits cell proliferation, increases cell apoptosis, blocks G0/G1 (growth phase) arrest, and reduces TET1 (a DNA demethylase) activity and NKD2 (a negative regulator of Wnt signaling). These findings suggest an anti-resistance effect of curcumin on 5-FU-resistant HCT116 cells via modulation of the TET1-NKD2-Wnt signaling pathway, thereby inhibiting the progression of epithelial-mesenchymal transition (EMT).94 Fan et al discovered that curcumin could prevent the advancement of tumors and reverse multidrug resistance in HCT-8/5-FU cells by inhibiting the heat shock protein-27 and P-gp. Resveratrol, a naturally occurring polyphenol, increased the sensitivity of HCT116/5-FU and SW480/5-FU cells to 5-FU. This occurs by slowing the process of epithelial-mesenchymal transition (EMT), which involves strengthening the connections between cells and reducing the NF-κB pathway.95 Resveratrol reversed resistance to L-OHP by enhancing drug accumulation, blocking the NF-κB/MDR1 pathway, and reducing the transcriptional activity of cAMP-responsive regions (in L-OHP-resistant) HCT116 cells.96 β-Elemene (sesquiterpene molecule), derived from the Chinese plant Curcumae rhizoma, effectively suppresses cell growth and overcomes the resistance of HCT116 cells to 5-FU by triggering cell cycle arrest and autophagy in a cyclin D3-dependent manner.97,98 Atractylenolide II, a single compound derived from the traditional Chinese medicinal plant Atractylodes macrocephala, can suppress tumor growth and enhance the sensitivity of Lovo and SW480 cell lines to the chemotherapy drugs 5-FU, mitomycin, and Adriamycin.98
Emodin, a naturally occurring compound derived from anthraquinone, counteracts 5-FU resistance in SW480/5-FU cells. It inhibits cell proliferation and migration, promotes cell death, and suppresses the PI3K/Akt pathway.99 Tanshinones are diterpene quinones extracted from the roots of Salvia miltiorrhiza Bunge. It possesses antioxidation, anticancer, anti-inflammation, and other pharmacological properties. Additionally, they have been shown to reverse chemoresistance. Dihydrotanshinone, Tanshinone IIA, and cryptotanshinone are three constituents of tanshinone that partially counteract chemoresistance in CRC. Tanshinone IIA can overcome L-OHP resistance in the SW480/L-OHP cell line by inhibiting the ERK/Akt signaling pathway. The application of Tanshinone IIA in combination with L-OHP led to a considerable decrease in the levels of Bcl-2, p-ERK, and p-Akt while increasing Bax and active caspase 3 levels.100 Cryptotanshinone and dihydrotanshinone inhibited the proliferation of MDR-resistant SW620/Ad300 cells by triggering p53-independent cytotoxicity and autophagic cell death and.101 Hypericin, an organic compound belonging to the anthraquinone class, is extensively studied in photodynamic therapy. Hypericin-based photodynamic therapy has the potential to restore sensitivity to L-OHP in CRC-resistant cells, specifically the HCT-8/L-OHP and HCT116/L-OHP cell lines. This restoration is achieved by reducing drug efflux (downregulating MRP2 levels), inhibiting GSH-related detoxification, and suppressing nucleotide excision repair (NER)-mediated DNA repair.93
Evodiamine, an alkaloid, can impede cell proliferation and trigger programmed cell death as well as reduce ABCG2-mediated multidrug resistance in HCT116/L-OHP cells by blocking the p50/p65 NF-κB pathway.102 Ginsenoside Rh2 prevents the growth and movement of cells, triggers cell death, and reduces the expression of genes associated with drug resistance, including glutathione S-transferase (GST), MRP1, lung resistance-related protein (LRP), and MDR1. This reversal of chemoresistance has been observed in HCT-8/5-FU and LoVo/5-FU cell lines.120 β-Sitosterol, a phytosterol, can counteract L-OHP multidrug resistance in HCT116/L-OHP cells by suppressing breast cancer resistance protein (BCRP/ABCG2). This is achieved by disrupting the interaction between murine double minute 2 (MDM2), an enzyme that attaches ubiquitin molecules to proteins, and p53.103 Thus, ubiquitination and degradation of p53 are inhibited, leading to p53 activation and increased apoptosis. Salvianolic acid B (phenolic acid) was obtained from the dried roots and rhizomes of Salvia miltiorrhiza Bge. (Labiatae) counteracted multidrug resistance (MDR) in HCT-8/VCR cells by increasing reactive oxygen species levels. Elevated ROS levels lead to apoptosis and reduced P-gp expression, thereby enhancing the sensitivity of drug-resistant cancer cells to chemotherapy.104
Photodynamic therapy (PDT) causes death of tumor cells via ROS generation. However, its effectiveness is hindered by hypoxia, one of the primary factors restricting its efficiency. Bufalin (BU) improves the effectiveness of mTHPC-mediated PDT for CRC. This was achieved through controlled release of BU, which suppressed HIF-1α and decreased VEGF-induced angiogenesis by targeting the SRC-3/HIF-1α pathway. As a result, a powerful PDT effect was observed against CRC. Bufalin enhances the effectiveness of photodynamic therapy in treating colorectal cancer by specifically targeting the SRC-3/HIF-1α pathway. The co-delivery of mTHPC and BU using nanoparticles effectively increased the therapeutic efficacy of PDT by suppressing the SRC-3/HIF-1α pathway in CRC. This study presents a successful approach to address the problem of tumor resistance caused by hypoxia and to overcome the obstacles associated with PDT treatment.104–106 Solid tumors are responsible for high morbidity globally. Tumors of solid origin often have hypoxic microenvironments that can be treated with immunotherapy, chemotherapy, and radiation. However, the effectiveness of molecular-targeted therapies in cancer treatment is low. HIF-1α inhibitors have been used in various clinical trials to treat solid tumors. Regrettably, there are currently no licensed medications available for the treatment of cancer patients that directly inhibit HIFs, either because of safety concerns or inadequate therapeutic efficacy. Thus, to pursue a specific approach for targeting HIF-1α, it would be worthwhile to investigate other natural chemicals to increase the likelihood of successful clinical trials for improved anticancer treatments. Additional research involving the use of both natural and synthetic substances in combination could offer a different approach for studying therapies that target HIF-1α.
Conclusion and Future Perspective
HIF-1α is upregulated in different tumor types in the absence of oxygen, leading to activation of gene transcription associated with tumor advancement. These genes are associated with the formation of new blood vessels, metabolic changes, cancer cell spread, and immune evasion. Various HIF-1α inhibitors have been developed to effectively treat cancer by targeting their fundamental role in promoting carcinogenesis under hypoxic conditions. The investigation of HIF-1α targeting in cancer therapy is a crucial and evolving area of study with the capacity to profoundly change current therapeutic approaches. Continued research and intervention in HIF-1 cancer-causing pathways are crucial for advancing colorectal cancer treatment and enhancing patient outcomes; thus, directing efforts towards HIF-1α in colorectal cancer treatment shows great potential as a therapeutic approach. This study discusses the discovery of several HIF-1α inhibitors derived from natural products that have shown promising antitumor properties in colorectal cancer. In general, natural products offer new molecules that can be used to develop HIF-1α inhibitors. However, additional structural modifications are necessary to generate medications with a greater potential. In summary, natural compounds show significant promise for further development as innovative and effective HIF-1α inhibitors and anticancer medicines by boosting their feasibility and minimizing their toxicity. Synergistic association between phytochemicals and synthetic HIF-1α inhibitors could prove to be effective in managing colorectal carcinoma. Thus, studies on the synergistic mechanism of action of both synthetic and natural drugs may create a unique avenue for HIF-1α targeting anti- colorectal cancer therapeutics. Further research is required to investigate the pharmacological and pharmacokinetic properties of natural compounds targeting HIF-1α and their associated mechanisms, along with modification of these molecules.
Data Sharing Statement
Not applicable as this is a review article.
Acknowledgment
This research was supported by the Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Education (NRF-2020R1I1A2066868), National Research Foundation of Korea (NRF) grant funded by the Korean government (MSIT) (No. 2020R1A5A2019413), Basic Science Research Program through the National Research Foundation of Korea(NRF) grant funded by the Korean government(MSIT)(RS-2024-00350362), Basic Science Research Program through the National Research Foundation of Korea(NRF) grant funded by the Korean government(MSIT)(RS-2024-00350362), and National Research Foundation of Korea. Grant Number: 2021R1C1C2014229.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This research was supported by the Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Education (NRF-2020R1I1A2066868), National Research Foundation of Korea (NRF) grant funded by the Korean government (MSIT) (No. 2020R1A5A2019413), Basic Science Research Program through the National Research Foundation of Korea(NRF) grant funded by the Korean government(MSIT)(RS-2024-00350362), Basic Science Research Program through the National Research Foundation of Korea(NRF) grant funded by the Korean government(MSIT)(RS-2024-00350362), National Research Foundation of Korea. Grant Number: 2021R1C1C2014229, and the Starting growth Technological R&D Program (TIPS Program, (No. RS-2024-00507224)) funded by the Ministry of SMEs and Startups (MSS, Korea) in 2024.
Disclosure
The authors declare no conflicts of interest in this work.
References
1. Tsimberidou AM. Targeted therapy in cancer. Cancer Chemother Pharmacol. 2015;76(6):1113–1132. doi:10.1007/s00280-015-2861-1
2. Lee YT, Tan YJ, Oon CE. Molecular targeted therapy: treating cancer with specificity. Eur J Pharmacol. 2018;834:188–196. doi:10.1016/j.ejphar.2018.07.034
3. Xie YH, Chen YX, Fang JY. Comprehensive review of targeted therapy for colorectal cancer. Signal Transduction Tar. 2020;5(1):22.
4. Di Nicolantonio F, Vitiello PP, Marsoni S, et al. Precision oncology in metastatic colorectal cancer—from biology to medicine. Nat Rev Clin Oncol. 2021;18(8):506–525. doi:10.1038/s41571-021-00495-z
5. Sørensen BS, Horsman MR. Tumor hypoxia: impact on radiation therapy and molecular pathways. Front Oncol. 2020;10:562. doi:10.3389/fonc.2020.00562
6. Abou Khouzam R, Brodaczewska K, Filipiak A, et al. Tumor hypoxia regulates immune escape/invasion: influence on angiogenesis and potential impact of hypoxic biomarkers on cancer therapies. Front Immunol. 2021;11:613114. doi:10.3389/fimmu.2020.613114
7. Roy S, Kumaravel S, Sharma A, Duran CL, Bayless KJ, Chakraborty S. Hypoxic tumor microenvironment: implications for cancer therapy. Exp Biol Med. 2020;245(13):1073–1086. doi:10.1177/1535370220934038
8. Augustin RC, Delgoffe GM, Najjar YG. Characteristics of the tumor microenvironment that influence immune cell functions: hypoxia, oxidative stress, metabolic alterations. Cancers. 2020;12(12):3802. doi:10.3390/cancers12123802
9. King R, Hayes C, Donohoe CL, Dunne MR, Davern M, Donlon NE. Hypoxia and its impact on the tumour microenvironment of gastroesophageal cancers. World J Gastrointest Oncol. 2021;13(5):312. doi:10.4251/wjgo.v13.i5.312
10. Juan CA, Pérez de la Lastra JM, Plou FJ, Pérez-Lebeña E. The chemistry of reactive oxygen species (ROS) revisited: outlining their role in biological macromolecules (DNA, lipids and proteins) and induced pathologies. Int J mol Sci. 2021;22(9):4642. doi:10.3390/ijms22094642
11. Semenza GL. Hypoxia-inducible factor 1: regulator of mitochondrial metabolism and mediator of ischemic preconditioning. Biochim Biophys Acta mol Cell Res. 2011;1813(7):1263–1268. doi:10.1016/j.bbamcr.2010.08.006
12. Semenza GL. Expression of hypoxia-inducible factor 1: mechanisms and consequences. Biochem Pharmacol. 2000;59(1):47–53. doi:10.1016/S0006-2952(99)00292-0
13. Infantino V, Santarsiero A, Convertini P, Todisco S, Iacobazzi V. Cancer cell metabolism in hypoxia: role of HIF-1 as key regulator and therapeutic target. Int J mol Sci. 2021;22(11):5703. doi:10.3390/ijms22115703
14. Manuelli V, Pecorari C, Filomeni G, Zito E. Regulation of redox signaling in HIF‐1‐dependent tumor angiogenesis. FEBS J. 2022;289(18):5413–5425. doi:10.1111/febs.16110
15. Rashid M, Zadeh LR, Baradaran B, et al. Up-down regulation of HIF-1α in cancer progression. Gene. 2021;798:145796. doi:10.1016/j.gene.2021.145796
16. Tanimoto K, Makino Y, Pereira T, Poellinger L. Mechanism of regulation of the hypoxia‐inducible factor‐1α by the von Hippel‐Lindau tumor suppressor protein. EMBO J. 2000;19(16):4298–4309. doi:10.1093/emboj/19.16.4298
17. Pugh CW, Ratcliffe PJ. The von Hippel–Lindau tumor suppressor, hypoxia-inducible factor-1 (HIF-1) degradation, and cancer pathogenesis. In: Seminars in Cancer Biology. Vols. 13, No. 1. Academic Press; 2003:83–89.
18. Pezzuto A, Carico E. Role of HIF-1 in cancer progression: novel insights. A review. Curr Mol Med. 2018;18(6):343–351. doi:10.2174/1566524018666181109121849
19. Palazon A, Tyrakis PA, Macias D, et al. An HIF-1α/VEGF-A axis in cytotoxic T cells regulates tumor progression. Cancer Cell. 2017;32(5):669–683. doi:10.1016/j.ccell.2017.10.003
20. Hapke RY, Haake SM. Hypoxia-induced epithelial to mesenchymal transition in cancer. Cancer Lett. 2020;487:10–20. doi:10.1016/j.canlet.2020.05.012
21. Siegel RL, Wagle NS, Cercek A, Smith RA, Jemal A. Colorectal cancer statistics, 2023. CA Cancer J Clin. 2023;73(3):233–254. doi:10.3322/caac.21772
22. Sedlak JC, Yilmaz ÖH, Roper J. Metabolism and colorectal cancer. Annual Review of Pathology: Mechanisms of Disease. 2023;18(1):467–492. doi:10.1146/annurev-pathmechdis-031521-041113
23. Xu Y, Li Y, Yang X, et al. Low-temperature plasma-activated medium enhances the chemosensitivity of colorectal cancer cells by improving hypoxia. Am J Cancer Res. 2023;13(5):1985.
24. Dzobo K, Senthebane DA, Dandara C. The tumor microenvironment in tumorigenesis and therapy resistance revisited. Cancers. 2023;15(2):376. doi:10.3390/cancers15020376
25. Yfantis A, Mylonis I, Chachami G, et al. Transcriptional response to hypoxia: the role of HIF-1-associated co-regulators. Cells. 2023;12(5):798. doi:10.3390/cells12050798
26. Zhao Y, Xing C, Deng Y, Ye C, Peng H. HIF-1α signaling: essential roles in tumorigenesis and implications in targeted therapies. Genes Dis. 2024;11(1):234–251. doi:10.1016/j.gendis.2023.02.039
27. Mazurakova A, Koklesova L, Csizmár SH, et al. Significance of flavonoids targeting PI3K/Akt/HIF-1α signaling pathway in therapy-resistant cancer cells–a potential contribution to the predictive, preventive, and personalized medicine. J Adv Res. 2024;55:103–118. doi:10.1016/j.jare.2023.02.015
28. Yong L, Tang S, Yu H, et al. The role of hypoxia-inducible factor-1 alpha in multidrug-resistant breast cancer. Front Oncol. 2022;12:964934. doi:10.3389/fonc.2022.964934
29. Lv Y, Zhao S, Han J, Zheng L, Yang Z, Zhao L. Hypoxia-inducible factor-1α induces multidrug resistance protein in colon cancer. Onco Targets Ther. 2015;1941–1948. doi:10.2147/OTT.S82835
30. Yang Y, Sun M, Wang L, Jiao B. HIFs, angiogenesis, and cancer. J Cell Biochem. 2013;114(5):967–974. doi:10.1002/jcb.24438
31. Hao LS, Liu Q, Tian C, et al. Correlation and expression analysis of hypoxia inducible factor 1α, glucose transporter 1 and lactate dehydrogenase 5 in human gastric cancer. Oncol Lett. 2019;18(2):1431–1441. doi:10.3892/ol.2019.10457
32. Jin F, Yang R, Wei Y, et al. HIF-1α-induced miR-23a∼ 27a∼ 24 cluster promotes colorectal cancer progression via reprogramming metabolism. Cancer Lett. 2019;440:211–222. doi:10.1016/j.canlet.2018.10.025
33. Dong S, Liang S, Cheng Z, et al. ROS/PI3K/Akt and Wnt/β-catenin signalings activate HIF-1α-induced metabolic reprogramming to impart 5-fluorouracil resistance in colorectal cancer. J Exp Clin Cancer Res. 2022;41(1):15. doi:10.1186/s13046-021-02229-6
34. Lee SH, Golinska M, Griffiths JR. HIF-1-independent mechanisms regulating metabolic adaptation in hypoxic cancer cells. Cells. 2021;10(9):2371. doi:10.3390/cells10092371
35. Li RL, He LY, Zhang Q, et al. HIF-1α is a potential molecular target for herbal medicine to treat diseases. Drug Des Devel Ther. 2020;Volume 14:4915–4949. doi:10.2147/DDDT.S274980
36. Koyasu S, Kobayashi M, Goto Y, Hiraoka M, Harada H. Regulatory mechanisms of hypoxia‐inducible factor 1 activity: two decades of knowledge. Cancer Sci. 2018;109(3):560–571. doi:10.1111/cas.13483
37. Huang M, Lu JJ, Ding J. Natural products in cancer therapy: past, present and future. Nat Prod Bioprospect. 2021;11(1):5–13. doi:10.1007/s13659-020-00293-7
38. Islam MR, Akash S, Rahman MM, et al. Colon cancer and colorectal cancer: prevention and treatment by potential natural products. Chem Biol Interact. 2022;368:110170. doi:10.1016/j.cbi.2022.110170
39. Lu L, Dong J, Liu Y, et al. New insights into natural products that target the gut microbiota: effects on the prevention and treatment of colorectal cancer. Front Pharmacol. 2022;13:964793. doi:10.3389/fphar.2022.964793
40. Petrocelli G, Farabegoli F, Valerii MC, Giovannini C, Sardo A, Spisni E. Molecules present in plant essential oils for prevention and treatment of colorectal cancer (CRC). Molecules. 2021;26(4):885. doi:10.3390/molecules26040885
41. An J, Liu Y, Wang Y, et al. The role of intestinal mucosal barrier in autoimmune disease: a potential target. Front Immunol. 2022;13:871713. doi:10.3389/fimmu.2022.871713
42. Schönberger T, Fandrey J, Prost-Fingerle K. Ways into understanding HIF inhibition. Cancers. 2021;13(1):159. doi:10.3390/cancers13010159
43. Ghosh R, Samanta P, Sarkar R, et al. Targeting HIF-1α by natural and synthetic compounds: a promising approach for anti-cancer therapeutics development. Molecules. 2022;27(16):5192. doi:10.3390/molecules27165192
44. Zhao Y, Huang H, Jia CH, et al. Apigenin increases radiosensitivity of glioma stem cells by attenuating HIF-1α-mediated glycolysis. Med Oncol. 2021;38(11):1–10. doi:10.1007/s12032-021-01586-8
45. Fu J, Zeng W, Chen M, et al. Apigenin suppresses tumor angiogenesis and growth via inhibiting HIF-1α expression in non-small cell lung carcinoma. Chem Biol Interact. 2022;361:109966. doi:10.1016/j.cbi.2022.109966
46. Yun BD, Son SW, Choi SY, Kuh HJ, Oh TJ, Park JK. Anti-cancer activity of phytochemicals targeting hypoxia-inducible factor-1 alpha. Int J mol Sci. 2021;22(18):9819. doi:10.3390/ijms22189819
47. Xu R, Wang F, Yang H, Wang Z. Action sites and clinical application of HIF-1α inhibitors. Molecules. 2022;27(11):3426. doi:10.3390/molecules27113426
48. Zhang Y, Liu K, Yan C, et al. Natural polyphenols for treatment of colorectal cancer. Molecules. 2022;27(24):8810. doi:10.3390/molecules27248810
49. Gong J, Zhou S, Yang S. Vanillic acid suppresses HIF-1α expression via inhibition of mTOR/p70S6K/4E-BP1 and Raf/MEK/ERK pathways in human colon cancer HCT116 cells. Int J mol Sci. 2019;20(3):465. doi:10.3390/ijms20030465
50. Lee DH, Lee YJ. Quercetin suppresses hypoxia‐induced accumulation of hypoxia‐inducible factor‐1α (HIF‐1α) through inhibiting protein synthesis. J Cell Biochem. 2008;105(2):546–553. doi:10.1002/jcb.21851
51. Darband SG, Kaviani M, Yousefi B, et al. Quercetin: a functional dietary flavonoid with potential chemo‐preventive properties in colorectal cancer. Cell Physiol. 2018;233(9):6544–6560. doi:10.1002/jcp.26595
52. Kim HS, Wannatung T, Lee S, et al. Quercetin enhances hypoxia-mediated apoptosis via direct inhibition of AMPK activity in HCT116 colon cancer. Apoptosis. 2012;17(9):938–949. doi:10.1007/s10495-012-0719-0
53. Uttarawichien T, Kamnerdnond C, Inwisai T, Suwannalert P, Sibmooh N, Payuhakrit W. Quercetin inhibits colorectal cancer cells induced-angiogenesis in both colorectal cancer cell and endothelial cell through downregulation of VEGF-A/VEGFR2. Sci Pharm. 2021;89(2):23. doi:10.3390/scipharm89020023
54. Monti E, Marras E, Prini P, Gariboldi MB. Luteolin impairs hypoxia adaptation and progression in human breast and colon cancer cells. Eur J Pharmacol. 2020;881:173210. doi:10.1016/j.ejphar.2020.173210
55. Liu R, Deng X, Peng Y, et al. Synthesis and biological evaluation of novel 5, 6, 7-trimethoxy flavonoid salicylate derivatives as potential anti-tumor agents. Bioorg. Chem. 2020;96:103652. doi:10.1016/j.bioorg.2020.103652
56. Sun Y, Li X, Cheng H, et al. Drug resistance and new therapies in gallbladder cancer. Drug Discov Ther. 2023;17(4):220–229. doi:10.5582/ddt.2023.01013
57. Xie J, Gao H, Peng J, et al. Hispidulin prevents hypoxia-induced epithelial-mesenchymal transition in human colon carcinoma cells. Am J Cancer Res. 2015;5(3):1047.
58. Bach A, Bender–Sigel J, Schrenk D, Flügel D, Kietzmann T. The antioxidant quercetin inhibits cellular proliferation via HIF-1-dependent induction of p21WAF. Antioxid Redox Sign. 2010;13(4):437–448. doi:10.1089/ars.2009.3000
59. Banik K, Ranaware AM, Harsha C, et al. Piceatannol: a natural stilbene for the prevention and treatment of cancer. Pharmacol Res. 2020;153:104635. doi:10.1016/j.phrs.2020.104635
60. Aljabali AA, Bakshi H, Hakkim LF, et al. Albumin nano-encapsulation of piceatannol enhances its anticancer potential in colon cancer via down regulation of nuclear p65 and HIF-1α. Cancers. 2020;12(1):113. doi:10.3390/cancers12010113
61. Sun X, Lv X, Yan Y, et al. Hypoxia-mediated cancer stem cell resistance and targeted therapy. Biomed Pharmacother. 2020;130:110623. doi:10.1016/j.biopha.2020.110623
62. Wang H, Zhao L, Zhu LT, et al. Wogonin reverses hypoxia resistance of human colon cancer HCT116 cells via downregulation of HIF‐1α and glycolysis, by inhibiting PI3K/Akt signaling pathway. Mol, Carcinog. 2014;53(S1):E107–E118. doi:10.1002/mc.22052
63. Cárdeno A, Sánchez-Hidalgo M, Rosillo MA, de la Lastra CA. Oleuropein, a secoiridoid derived from olive tree, inhibits the proliferation of human colorectal cancer cell through downregulation of HIF-1α. Nutr Cancer. 2013;65(1):147–156. doi:10.1080/01635581.2013.741758
64. Brockmueller A, Girisa S, Motallebi M, Kunnumakkara AB, Shakibaei M. Calebin A targets the HIF-1α/NF-κB pathway to suppress colorectal cancer cell migration. Front Pharmacol. 2023;14:1203436. doi:10.3389/fphar.2023.1203436
65. Kim SL, Park YR, Lee ST, Kim SW. Parthenolide suppresses hypoxia-inducible factor-1α signaling and hypoxia induced epithelial-mesenchymal transition in colorectal cancer. Int j Oncol. 2017;51(6):1809–1820. doi:10.3892/ijo.2017.4166
66. Luo Q, Wang J, Zhao W, et al. Vasculogenic mimicry in carcinogenesis and clinical applications. J Hematol Oncol. 2020;13(1):1–15. doi:10.1186/s13045-020-00858-6
67. Leng J, Lei L, Lei SF, Zhu Z, Ocampo A, Gany F. Use of traditional Chinese herbal medicine concurrently with conventional cancer treatment among Chinese cancer patients. J Immigr Minor Health. 2020;22(6):1240–1247. doi:10.1007/s10903-020-01017-2
68. Peng W, Zhang S, Zhang Z, et al. Jianpi Jiedu decoction, a traditional Chinese medicine formula, inhibits tumorigenesis, metastasis, and angiogenesis through the mTOR/HIF-1α/VEGF pathway. J Ethnopharmacol. 2018;224:140–148. doi:10.1016/j.jep.2018.05.039
69. Zong S, Tang Y, Li W, et al. A Chinese herbal formula suppresses colorectal cancer migration and vasculogenic mimicry through ROS/HIF-1 α/MMP2 pathway in hypoxic microenvironment. Front Pharmacol. 2020;11:705. doi:10.3389/fphar.2020.00705
70. Mi C, Ma J, Wang KS, et al. Imperatorin suppresses proliferation and angiogenesis of human colon cancer cells by targeting HIF-1α via the mTOR/p70S6K/4E-BP1 and MAPK pathways. J Ethnopharmacol. 2017;203:27–38. doi:10.1016/j.jep.2017.03.033
71. Huang J, Chen ZH, Ren CM, et al. Antiproliferation effect of evodiamine in human colon cancer cells is associated with IGF-1/HIF-1α downregulation. Oncol Rep. 2015;34(6):3203–3211. doi:10.3892/or.2015.4309
72. Shan JZ, Xuan YY, Zhang Q, Huang JJ. Ursolic acid sensitized colon cancer cells to chemotherapy under hypoxia by inhibiting MDR1 through HIF-1α. J Zhejiang Univ Sci B. 2016;17(9):672. doi:10.1631/jzus.B1600266
73. Rashid HU, Xu Y, Muhammad Y, Wang L, Jiang J. Ursolic acid sensitized colon cancer cells to chemotherapy under hypoxia by inhibiting MDR1 through HIF-1α. J Zhejiang Univ Sci B. 2019;17(9):672.
74. Nguyen HN, Ullevig SL, Short JD, Wang L, Ahn YJ, Asmis R. Ursolic acid and related analogues: triterpenoids with broad health benefits. Antioxidants. 2021;10(8):1161. doi:10.3390/antiox10081161
75. Hong X, Zhong L, Xie Y, et al. Matrine reverses the Warburg effect and suppresses colon cancer cell growth via negatively regulating HIF-1α. Front Pharmacol. 2019;10:478472. doi:10.3389/fphar.2019.01437
76. Yan L, Raj P, Yao W, Ying H. Glucose metabolism in pancreatic cancer. Cancers. 2019;11(10):1460. doi:10.3390/cancers11101460
77. Mao L, Chen Q, Gong K, et al. Berberine decelerates glucose metabolism via suppression of mTOR dependent HIF-1α protein synthesis in colon cancer cells. Oncol Rep. 2018;39(5):2436–2442. doi:10.3892/or.2018.6318
78. Nandini DB, Rao RS, Deepak BS, Reddy PB. Sulforaphane in broccoli: the green chemoprevention!! Role in cancer prevention and therapy. J Oral Maxillofac Pathol. 2020;24(2):405. doi:10.4103/jomfp.JOMFP_126_19
79. Kim DH, Sung B, Kang YJ, et al. Sulforaphane inhibits hypoxia-induced HIF-1α and VEGF expression and migration of human colon cancer cells. Int J Oncol. 2015;47(6):2226–2232. doi:10.3892/ijo.2015.3200
80. Pastorek M, Simko V, Takacova M, et al. Sulforaphane reduces molecular response to hypoxia in ovarian tumor cells independently of their resistance to chemotherapy. Int J Oncol. 2015;47(1):51–60. doi:10.3892/ijo.2015.2987
81. Bach DH, Kim SH, Hong JY, Park HJ, Oh DC, Lee SK. Salternamide A suppresses hypoxia-induced accumulation of HIF-1α and induces apoptosis in human colorectal cancer cells. Mar Drugs. 2015;13(11):6962–6976. doi:10.3390/md13116962
82. Seyfi D, Behzad SB, Nabiuni M, Parivar K, Tahmaseb M, Amini E. Verbascoside attenuates Rac-1 and HIF-1α signaling cascade in colorectal cancer cells. Anticancer Agents Med Chem. 2018;18(15):2149–2155. doi:10.2174/1871520618666180611112125
83. Zhang L, Qiao X, Chen M, et al. Ilexgenin A prevents early colonic carcinogenesis and reprogramed lipid metabolism through HIF-1α/SREBP-1. Phytomedicine. 2019;63:153011. doi:10.1016/j.phymed.2019.153011
84. Chen M, Zhong K, Tan J, et al. Baicalein is a novel TLR4‐targeting therapeutics agent that inhibits TLR4/HIF‐1α/VEGF signaling pathway in colorectal cancer. Clin Transl Med. 2021;11(11). doi:10.1002/ctm2.564.
85. Ji L, Shen W, Zhang F, et al. Worenine reverses the Warburg effect and inhibits colon cancer cell growth by negatively regulating HIF-1α. Cell Mol Biol Lett. 2021;26(1):19. doi:10.1186/s11658-021-00263-y
86. Huang Y, Liu Z, Li L, et al. Sesamin inhibits hypoxia-stimulated angiogenesis via the NF-κB p65/HIF-1α/VEGFA signaling pathway in human colorectal cancer. Food Funct. 2022;13(17):8989–8997. doi:10.1039/D2FO00345G
87. Lee SY, Kim HJ, Oh SC, Lee DH. Genipin inhibits the invasion and migration of colon cancer cells by the suppression of HIF-1α accumulation and VEGF expression. Food Chem Toxicol. 2018;116:70–76. doi:10.1016/j.fct.2018.04.005
88. Lu Y, Wang B, Shi Q, Wang X, Wang D, Zhu L. Brusatol inhibits HIF-1 signaling pathway and suppresses glucose uptake under hypoxic conditions in HCT116 cells. Sci Rep. 2016;6(1):39123. doi:10.1038/srep39123
89. Iovine B, Oliviero G, Garofalo M, et al. The anti-proliferative effect of L-carnosine correlates with a decreased expression of hypoxia inducible factor 1 alpha in human colon cancer cells. PLoS One. 2014;9(5):e96755. doi:10.1371/journal.pone.0096755
90. Wang Z, Li MY, Zhang ZH, et al. Panaxadiol inhibits programmed cell death-ligand 1 expression and tumour proliferation via hypoxia-inducible factor (HIF)-1α and STAT3 in human colon cancer cells. Pharmacol Res. 2020;155:104727. doi:10.1016/j.phrs.2020.104727
91. Nagaraju GP, Bramhachari PV, Raghu G, El-Rayes BF. Hypoxia inducible factor-1α: its role in colorectal carcinogenesis and metastasis. Cancer Lett. 2015;366(1):11–18. doi:10.1016/j.canlet.2015.06.005
92. Zhang W, Ou L, Peng C, et al. Sanguisorba officinalis L. enhances the 5-fluorouracil sensitivity and overcomes chemoresistance in 5-fluorouracil-resistant colorectal cancer cells via Ras/MEK/ERK and PI3K/Akt pathways. Heliyon. 2023;9(6).
93. Ma SC, Zhang JQ, Yan TH, et al. Novel strategies to reverse chemoresistance in colorectal cancer. Cancer Med. 2023;12(10):11073–11096. doi:10.1002/cam4.5594
94. Lu Y, Zhang R, Zhang X, Zhang B, Yao Q. Curcumin may reverse 5-fluorouracil resistance on colonic cancer cells by regulating TET1-NKD-Wnt signal pathway to inhibit the EMT progress. Biomed Pharmacother. 2020;129:110381. doi:10.1016/j.biopha.2020.110381
95. Fan YX, Abulimiti P, Zhang HL, Zhou YK, Zhu L. Mechanism of reversal of multidrug resistance by curcumin in human colorectal cancer cell line HCT-8/5-FU. Genet Mol Res. 2017;16(2):10. doi:10.4238/gmr16029414
96. Wang Z, Zhang L, Ni Z, et al. Resveratrol induces AMPK-dependent MDR1 inhibition in colorectal cancer HCT116/L-OHP cells by preventing activation of NF-κB signaling and suppressing cAMP-responsive element transcriptional activity. Tumor Biol. 2015;36(12):9499–9510. doi:10.1007/s13277-015-3636-3
97. Chen Y, Zhu Z, Chen J, et al. Terpenoids from Curcumae rhizoma: their anticancer effects and clinical uses on combination and versus drug therapies. Biomed Pharmacother. 2021;138:111350. doi:10.1016/j.biopha.2021.111350
98. Tan T, Li J, Luo R, et al. Recent advances in understanding the mechanisms of elemene in reversing drug resistance in tumor cells: a review. Molecules. 2021;26(19):5792. doi:10.3390/molecules26195792
99. Zhang D, Li X, Song D, et al. Atractylenolide III induces apoptosis by regulating the Bax/Bcl-2 signaling pathway in human colorectal cancer HCT-116 Cells in vitro and in vivo. Anti-Cancer Drugs. 2022;33(1):30–47. doi:10.1097/CAD.0000000000001136
100. Liu W, Qaed E, Zhu Y, et al. Research progress and new perspectives of anticancer effects of Emodin. Am J Chin Med. 2023;51(07):1751–1793. doi:10.1142/S0192415X23500787
101. Sui H, Zhao J, Zhou L, et al. Tanshinone IIA inhibits β-catenin/VEGF-mediated angiogenesis by targeting TGF-β1 in normoxic and HIF-1α in hypoxic microenvironments in human colorectal cancer. Cancer Lett. 2017;403:86–97. doi:10.1016/j.canlet.2017.05.013
102. Mühleisen L, Alev M, Unterweger H, et al. Analysis of hypericin-mediated effects and implications for targeted photodynamic therapy. Int J mol Sci. 2017;18(7):1388. doi:10.3390/ijms18071388
103. Bharathiraja P, Yadav P, Sajid A, Ambudkar SV, Prasad NR. Natural medicinal compounds target signal transduction pathways to overcome ABC drug efflux transporter-mediated multidrug resistance in cancer. Drug Resist Updat. 2023;71:101004.
104. Ni B, Song X, Shi B, et al. Research progress of ginseng in the treatment of gastrointestinal cancers. Front Pharmacol. 2022;13:1036498. doi:10.3389/fphar.2022.1036498
105. Shi M, Huang F, Deng C, Wang Y, Kai G. Bioactivities, biosynthesis and biotechnological production of phenolic acids in Salvia miltiorrhiza. Crit Rev Food Sci Nutr. 2019;59(6):953–964. doi:10.1080/10408398.2018.1474170
106. Yuan Z, Liu C, Sun Y, et al. Bufalin exacerbates photodynamic therapy of colorectal cancer by targeting SRC-3/HIF-1α pathway. Int J Pharm. 2022;624:122018. doi:10.1016/j.ijpharm.2022.122018
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms.php
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms.php
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.