")
Back to Journals » International Journal of Nanomedicine » Volume 20
Research Advances and Application Progress on miRNAs in Exosomes Derived From M2 Macrophage for Tissue Injury Repairing
Authors Zhu Z, Zhang X, Lin X, Wang Y , Han C , Wang S
Received 5 December 2024
Accepted for publication 23 January 2025
Published 4 February 2025 Volume 2025:20 Pages 1543—1560
DOI https://doi.org/10.2147/IJN.S508781
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 4
Editor who approved publication: Professor Eng San Thian
Zhikang Zhu,1,* Xinge Zhang,1,* Xuran Lin,1 Yuechen Wang,1 Chunmao Han,2,3 Shoujie Wang1,4
1Department of Plastic Surgery, The Fourth Affiliated Hospital of School of Medicine, and International School of Medicine, Zhejiang University, Yiwu, Zhejiang, People’s Republic of China; 2Department of Burns & Wound Care Center, The Second Affiliated Hospital, School of Medicine, Zhejiang University, Hangzhou, Zhejiang, People’s Republic of China; 3Key Laboratory of The Diagnosis and Treatment of Severe Trauma and Burn of Zhejiang Province, Hangzhou, Zhejiang, People’s Republic of China; 4Department of Plastic Surgery, The First Affiliated Hospital, School of Medicine, Zhejiang University, Hangzhou, Zhejiang, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Shoujie Wang; Chunmao Han, Email [email protected]; [email protected]
Abstract: Tissue injury repair is a multifaceted and dynamic process characterized by complex interactions among various immune cells, with M2 macrophages assuming a crucial role. Exosomes derived from M2-type macrophages (M2-Exos) significantly influence the injury repair process through intercellular communication mediated by enriched microRNAs (miRNAs). This review aims to elucidate the biological processes underlying exosome formation, the synthesis and function of miRNAs, and the diverse methodologies employed for exosome extraction. Furthermore, we provide a comprehensive summary of the established multifarious functions and mechanisms of M2-Exos miRNAs in tissue injury repair across different systems, while also exploring their potential applications in disease prevention, diagnosis, and clinical practice. Despite the challenges encountered, the therapeutic use of M2-Exos in clinical contexts appears promising, prompting research efforts to focus on improving the efficiency of exosome extraction and application, as well as ensuring the safety of their clinical utilization.
Keywords: M2 macrophages, exosomes, miRNAs, tissue repair, drug delivery systems, engineered exosome
Introduction
Tissue injury, which may arise from various causes such as infections, autoimmune disorders, mechanical trauma, or toxic agents, triggers a complex and dynamic reparative process. In the acute phase following an injury to tissue or organs, there is a rapid recruitment of neutrophils, monocytes (precursors to macrophages), and other immune cells to the affected area. These immune cells play a dual role; they are involved in the clearance of necrotic tissue and the elimination of pathogens, while also actively participating in the subsequent repair of tissue.1 Monocytes and macrophages are pivotal components of the human immune system, significantly influencing tissue repair and regeneration.2,3 They secrete matrix metalloproteinases (MMPs) that degrade the basement membrane and produce chemokines, which facilitate the migration of inflammation-associated cells to the injury site. Additionally, these cells engage in the phagocytosis of cellular debris and pathogenic microorganisms, aiding in the removal of damaged cells.4 The recruitment and activation of macrophages at the injury site are essential; any limitation in this process can lead to an extended inflammatory phase, thereby delaying the repair of injuries. Moreover, the phenotypic transformation of macrophages is crucial for regulating cell proliferation and regeneration, and any disruption in this transformation may result in incomplete tissue regeneration.5,6 Consequently, the orchestrated recruitment and activation of macrophages, along with their functional transformations at various stages, are imperative for the effective repair of tissue injury.7
Macrophages play a crucial role in the regulation of tissue repair following injury by secreting signaling molecules that facilitate communication with adjacent cells. Among the various macrophage subtypes, M2-type macrophages are particularly proficient in enhancing tissue repair processes. Their communication with surrounding cells is usually mediated through exosome-dependent mechanisms, which are significantly influenced by microRNAs (miRNAs) contained within these exosomes.8–10 These miRNAs are derived from diverse genomic loci and undergo post-transcriptional processing and modification within the cell, ultimately maturing into single-stranded RNA molecules approximately 22 nucleotides in length. The mature miRNAs are associated with RNA-induced silencing complexes (RISCs), which act as regulatory entities by modulating the degradation or translation of target message RNAs (mRNAs).11 The miRNAs derived from M2-type macrophage exosomes (M2-Exos) are closely linked to tissue injury and are involved in various biological processes, including cell proliferation, granulation tissue formation, angiogenesis, and inflammation regulation. Through the regulation of these activities, M2-Exos contributes to the therapeutic management of respiratory, neurological, circulatory, systemic, and chronic inflammatory diseases.12–15 Therefore, M2-Exos and their associated miRNAs provide a robust theoretical framework for the synergistic application of exosome nanotechnology and macrophage immunotherapy.13,16,17
Considering the crucial regulatory function of miRNAs in M2-Exos concerning tissue repair mechanisms and their prospective clinical implications, we have undertaken an extensive examination and literature review focused on miRNAs in M2-Exos. This review aims to clarify existing challenges while providing insights into the utilization of intracellular miRNAs within M2-Exos, as well as identifying potential directions for future research in this field (Figure 1).
 |
Figure 1 A scheme of miRNAs in exosomes derived from M2 macrophage for tissue injury repairing. |
Biogenesis and Function of M2-Exos
Exosomes have gained significant attention in the field of research as crucial mediators of communication between M2 macrophages and their surrounding microenvironment. These nanoscale extracellular vesicles are derived from the endomembrane system. Their formation involves a process characterized by the double invagination of the plasma membrane, which leads to the development of intracellular multivesicular bodies (MVBs). Within these MVBs are intraluminal vesicles (ILVs), which are subsequently processed into exosomes with diameters ranging from 40 to 160 nm. The fate of MVBs can vary; they may either merge with lysosomes or autophagosomes for degradation, or they may fuse with the plasma membrane, facilitating the release of exosomes into the extracellular space via a process known as exocytosis (Figure 2).18 The process of exosome release plays a crucial role in cellular communication and the transmission of biological signals. Numerous regulatory factors involved in the synthesis and secretion of exosomes have been identified, such as the Endosomal Sorting Complex Required for Transport (ESCRT), Rab proteins (GTPases), Sytenin1, and tumor susceptibility genes (TSG101). However, the precise functions of these factors have yet to be elucidated.19
 |
Figure 2 A visual illustration depicting the process of exosome formation:1) The first invagination of the plasma membrane forms a cup-shaped structure that includes cell surface proteins and soluble proteins associated with the extracellular environment through the formation of early sorting endosomes (ESEs).20,21 2) Subsequently, a double-layered MVB is formed through the inward invagination of the plasma membrane of the ESE. The ESE matures into a late-sorting endosome (LSE), which also includes intraluminal vesicles (ILVs; future exosomes) that are about 100 nm in diameter. 3) Finally, the MVB may fuse with lysosomes or autophagosomes and undergo degradation, or it may fuse with the plasma membrane to release the stored ILVs as exosomes.19,20,22 |
The established mechanisms by which exosomes interact with their target cells encompass three primary processes: membrane fusion, direct receptor interaction, and internalization.23 Variations in these mechanisms and the pathways involved in the uptake of exosomes by recipient cells, along with the metabolic state of these cells and the chemotactic specificity of exosomes for particular cell types, may result in alterations to the composition of exosomes. Such variations complicate the investigation of exosomal functions in intercellular communication.22,24,25 Current research predominantly emphasizes the induction of phenotypic changes and the biological alterations in recipient cells. Most experimental investigations of exosomes are conducted using exosomes derived from in vitro cultures; however, it remains uncertain whether these in vitro-generated exosomes maintain the same properties as those produced in vivo when utilized in functional studies.26–28 Additionally, discrepancies in the conditions under which exosomes are extracted may lead to differential gene expression within the exosomes, potentially resulting in variations in their regulatory functions.29,30 Therefore, further elucidation of the pathways through which exosomes exert their effects on recipient cells is warranted. It is also essential to explore whether the regulatory functions of exosomes cultured in vitro are physiologically comparable to those derived from in vivo sources.
Exosomal miRNAs Biology of Macrophages
The synthesis and maturation of miRNAs is a complex, multistep process, as illustrated in Figure 3. 31,32 Following their synthesis and maturation, miRNAs perform their regulatory functions through interactions with mRNA. A single miRNA can modulate the expression of multiple target mRNAs, while a single mRNA may be subject to regulation by various miRNAs.33 miRNAs can engage in several other mechanisms of action: 1) Inhibition of translation: miRNAs primarily exert their effects post-transcriptionally by base pairing with the 3′ untranslated region (3′ UTR) of mRNAs. miRNAs can impede translation at the initiation phase by targeting the cap recognition process or by obstructing the assembly of the ribosomal 80S complex, as well as at the post-initiation stage.34 In a non-classical mechanism, after binding to mRNAs, miRNAs may also attach to the 5′ UTR of mRNAs to inhibit translation.35 2) Under specific conditions, miRNAs may enhance translation rather than inhibit it. Research by Vasudevan et al indicates that miRNAs suppress translation in proliferating cells but promote it in quiescent cells that are arrested at the G0/G1 checkpoint.36 Furthermore, Orom et al demonstrated that miR-10a interacts with the 5′ UTRs of various mRNAs encoding ribosomal proteins, thereby activating their translation.34,36,37 3) Deadenylation and decay of mRNAs: miRNAs can destabilize target mRNAs through mechanisms such as deadenylation, subsequent decapping, and 5′-3′ nucleic acid exonuclease activity.34 Notably, some mature miRNAs have been identified in the nucleus, where they can induce the degradation of nuclear mRNAs.38 4) Activation of transcription: In addition to repressing the expression of target genes, certain miRNAs can activate the transcription of target genes following their binding to mRNAs.29
 |
Figure 3 The formation of miRNA and its main mechanism of action: First, in the nucleus, RNA polymerase II catalyzes the formation of double-stranded RNAs with a stem-loop structure called primary miRNAs (pri-miRNAs). Subsequently, the pri-miRNAs are cleaved into 60–70 nucleotide precursor miRNAs (pre-miRNAs) by RNA polymerase III (RNase III), Drosha and its cofactor, DGCR8, are then, transferred to the cytoplasm via the transport protein Exportin 5. In the cytoplasm, the pre-miRNAs are sheared and processed by another RNase III, known as Dicer to generate a segment of complementary miRNA duplex. Finally, with the help of the Argonaute protein, the miRNA duplex removes the non-functional coding single strand and dissociates into a functionally active “leading strand”, which forms a miRNA-induced silencing complex (miRISC) with RISC protein. The RISC-coated miRNA is complementarily paired with the 3′-UTR of the target mRNA through its “seed region” (2–8 nucleotides at the 5′ end). This binding leads to the inhibition of gene expression or promotion of mRNA degradation, resulting in the regulation of the expression of target genes. 31,39,40 |
While the specific mechanisms by which miRNAs regulate gene transcription or translation through classical or non-classical pathways are not yet fully elucidated, these processes highlight the intricate nature and importance of miRNAs in the functional regulation of diverse biological processes.
Isolation Methods of M2-Exos
Exosomes play a crucial role in intercellular communication, and their regulatory functions are of considerable importance, leading to an annual increase in research focused on these entities. However, current extraction methods do not consistently yield the desired levels of purity and concentration. To address the challenges of reducing extraction costs and enhancing the purity of isolated exosomes to fulfill research requirements, there is a growing diversification and targeting of extraction techniques. In this context, we provide a comprehensive summary of the most recent methodologies for the extraction of macrophage-derived exosomes, as detailed in Table 1.
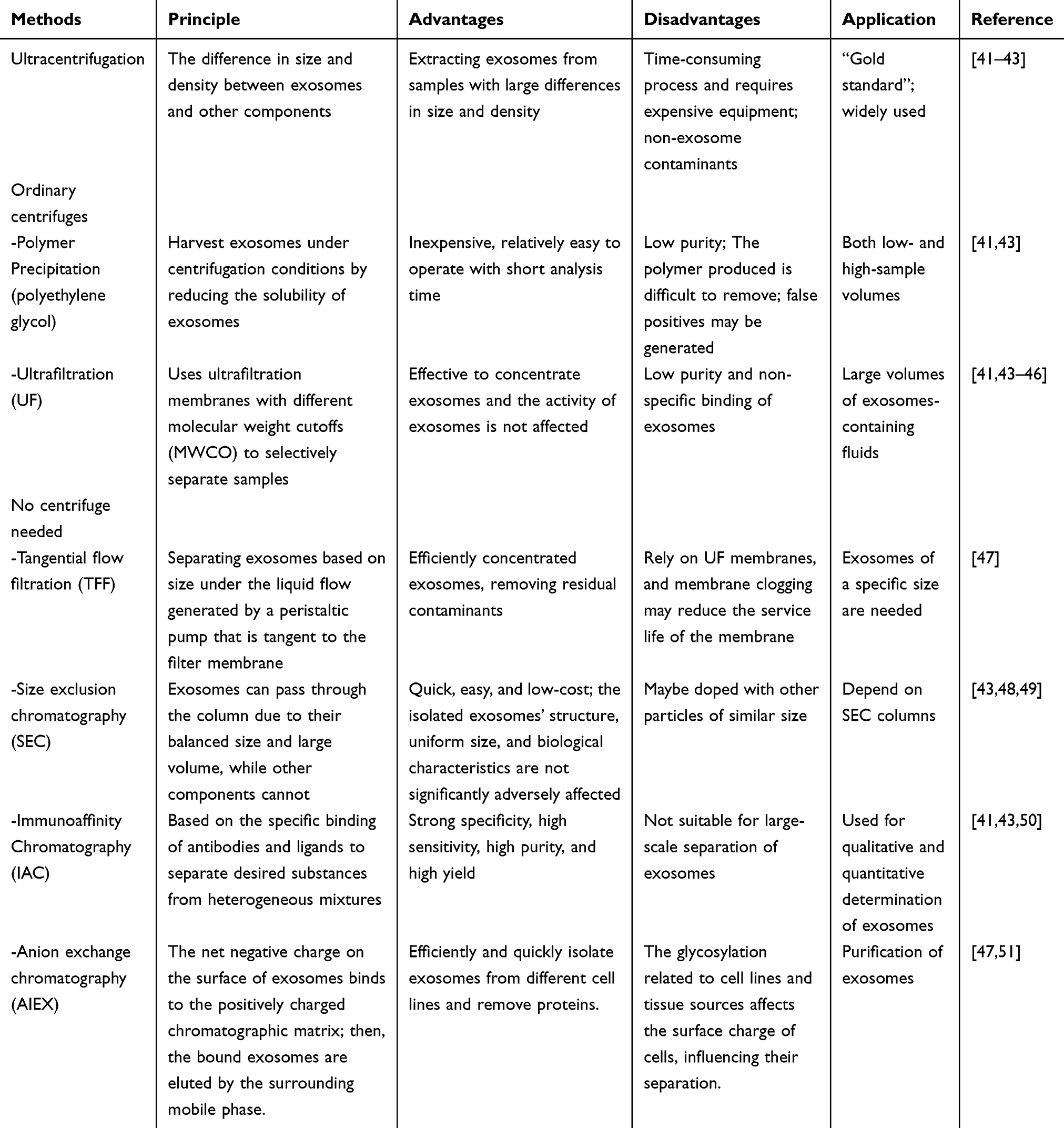 |
Table 1 Methods Used to Extract Macrophage-Derived Exosomes |
Ultracentrifugation is the predominant technique employed for the extraction of macrophage-derived exosomes and is regarded as the “gold standard” for this purpose.52,53 The methodology is predicated on the differential size and density of exosomes compared to other cellular components, rendering it particularly effective for isolating exosomes from samples characterized by significant variations in size and density.41 The extraction process encompasses several sequential steps. Initially, the cell supernatant undergoes centrifugation at low speeds (500–10,000 g) at 4 °C, followed by filtration to eliminate debris from the samples. Subsequently, exosomes present in the culture medium or tissue fluid are precipitated through ultracentrifugation. The morphological features of the isolated exosomes are then evaluated using transmission electron microscopy (TEM), while their sizes are quantified via nanoparticle tracking analysis (NTA).41,48 This method is recognized for its ability to yield exosomes of high purity and activity. However, it is important to note that the process is labor-intensive and necessitates costly equipment. Furthermore, various factors, including centrifugation duration and force, can significantly influence the extraction efficiency and purity of the target exosomes.54
Given the labor-intensive and equipment-dependent characteristics of ultracentrifugation, researchers have devised alternative methods for exosome extraction that can be executed using standard centrifuges or even in the absence of centrifugation. For instance, conventional centrifugation techniques such as polymer precipitation and ultrafiltration can effectively isolate and purify exosomes.55 Additionally, methods such as Tangential Flow Filtration (TFF), Size Exclusion Chromatography (SEC), and Anion Exchange Chromatography (AEC) do not necessitate the use of a centrifuge.41,56,57 However, the purity and recovery rates of exosomes obtained through these methods tend to be suboptimal, which may hinder subsequent functional analyses.
In response to these challenges, recent technological advancements have led to the development of numerous commercial kits designed to streamline the exosome extraction process. These kits facilitate the rapid acquisition of significant quantities of exosomes while preserving their structural integrity, thereby offering considerable potential for various applications. Nonetheless, these kits frequently do not achieve optimal separation efficiency, and their high cost, coupled with the inadequate purity of the extracted exosomes, presents notable limitations. To address these shortcomings, employing a combination of different exosome extraction techniques or integrating multiple methods tailored to specific research objectives may enhance both the purity and yield of exosomes.41
M2-Exos miRNAs in Diagnosis and Prognosis of Tissue Injury
Exosomes are present in a variety of tissue and body fluids, including blood, urine, and bile,58 and their surface proteins, combined with their inherent stability, render them promising candidates for diagnostic and prognostic biomarkers in numerous injury-related diseases.59–61 Concurrently, macrophages, which play pivotal roles in various tissue injury diseases and exhibit complex regulatory functions, release exosomes that may be more valuable for diagnostic and preventive applications than exosomes derived from other sources (Figure 4). The miRNA within exosomes derived from macrophages is influenced by their phenotype, with exosomes from M2 macrophages potentially exhibiting higher levels of specific miRNAs compared to those from M1 macrophages.62 Furthermore, it has been proposed that the detection of exosomes can facilitate longitudinal sampling to monitor disease progression and may be incorporated into minimally invasive liquid biopsy methodologies in clinical settings.18,63 This is particularly relevant as macrophage-derived exosomes represent a substantial fraction of blood-borne exosomes, and analyzing their content and dynamic release provides a novel strategy for tracking disease progression.64
 |
Figure 4 M2-Exos for disease prevention and diagnosis. |
Current detection methodologies primarily focus on capturing the complex extracellular and intracellular “molecular cargo” and conducting multiparameter diagnostic assays. Historically, exosomes have been utilized mainly as diagnostic tools for diseases associated with cardiovascular and central nervous system (CNS) injuries. However, recent investigations have broadened the focus to encompass injuries affecting other tissues and organs, such as the liver, kidneys, and lungs.48,65,66 In a review, Yang et al highlighted the role of miRNAs in exosomes from various cell types, including hepatic macrophages, where these miRNAs significantly contribute to the inhibition of fibrosis, inflammatory responses, and lipogenesis, thereby facilitating the diagnosis of non-alcoholic fatty liver disease (NAFLD). They further posited that miRNAs in macrophage-derived exosomes hold considerable promise for the diagnosis and prognosis of NAFLD.67 Nevertheless, it is important to note that macrophage-derived exosomes may differ based on the characteristics of their parent cells. Therefore, identifying the specific exosomes secreted by distinct macrophage phenotypes may enhance efforts in the prevention, diagnosis, and treatment of related diseases.64
M2-Exosomal miRNAs for Tissue Injury Repairing
Numerous studies on M2-Exos miRNAs have demonstrated their ability to either promote or hinder tissue injury repair in both cellular and animal models. In this section, we present a detailed overview of the known functions of miRNAs in M2-Exos and the mechanisms that regulate these functions in relation to tissue repair, systematically arranged based on the anatomical structures of the human body (Figure 5).
 |
Figure 5 The regulatory role of M2-Exos miRNAs in tissue injury repair: (A) Repairing for respiratory system injury. (B) Repairing for central nervous system injury. (C) Repairing for circulatory system injury. (D) Repairing for urinary system injury. (E) Repairing for skin injury. (F) Repairing for locomotor system injury. |
Repairing for Respiratory System Injury
Pulmonary Fibrosis Therapy
Pulmonary fibrosis (PF) is a permanent lung condition marked by the growth of fibroblasts, significant accumulation of extracellular matrix (ECM), inflammation, and damage to lung tissue structure. Research by Yao et al indicated that miRNA-328 from M2-Exos can stimulate the growth of lung interstitial fibroblasts and enhance the expression of proliferation and differentiation markers, such as collagen, by inhibiting FAM13A. Increased collagen levels lead to the release of inflammatory factors from macrophages, worsening fibrosis after tissue injury and accelerating the progression of PF.68 However, Guiot et al reported that macrophage-derived exosomes can alleviate the progression of fibrosis in alveolar epithelial cells and lung fibroblasts by delivering miR-142-3p, which inhibits transforming growth factor beta receptor 1 (TGFβ-R1).69,70 Conversely, Guiot et al found that exosomes derived from macrophages can slow down fibrosis in alveolar epithelial cells and lung fibroblasts by delivering miR-142-3p, which blocks the transforming growth factor beta receptor 1 (TGFβ-R1).69 Considering the opposing roles of miRNAs within M2-Exos, we propose that miRNA-328, which promotes PF progression, may serve as a future diagnostic biomarker for PF, whereas miR-142-3p, which inhibits PF progression, could be used as a therapeutic miRNA in clinical settings.
Asthma Therapy
Many studies have identified the therapeutic significance of miRNAs of M2-Exos in the progression of asthma. Li et al discovered that miR-370 from M2-Exos can reduce the growth of mouse airway smooth muscle cells (ASMCs) and airway remodeling by decreasing the levels of fibroblast growth factor 1 (FGF1) and the MAPK/STAT1 signaling pathway, thereby mitigating the progression of asthma.71 In a similar study, Tang et al demonstrated that a powder made from Chinese medicine, specifically scorpion and centipede, encourages M2 polarization and boosts the release of M2-Exos. This process helps to reduce inflammation in asthmatic mice by delivering miR-30b-5p, which lowers the levels of NLRP3, caspase-1, and IL-1β, as well as mitochondrial swelling, contributing to asthma treatment.72 Consequently, miR-370 and miR-30b-5p found in M2-Exos can help reduce asthma symptoms by preventing fibrosis in airway epithelial cells and inhibiting pyroptosis in these cells, respectively, which offered a more effective and innovative approach to treating the progression of asthma.
Repairing for Central Nervous System Injury
Cerebral Ischemia-Reperfusion Therapy
Following cerebral ischemia-reperfusion injury, intracranial macrophages serve as the initial defense against tissue injury. As the disease progresses, intracerebral macrophages change from the M2 type to the M1 type, which worsens the injury to brain tissue. Luckily, in the central nervous system, microglia polarize to the M2 phenotype and exert neuroprotective effects during cerebral ischemic injury.73 Furthermore, M2 microglia not only promote angiogenesis but also attenuate brain damage.74 Song et al showed that miR-124 in M2 microglia-derived exosomes would enter the damaged neurons and downregulate the expression of ubiquitin-specific protease 14 (USP14), thereby attenuating ischemic brain injury and promoting the survival of neurons.75 Additionally, Zhang et al found that microglial exosomes inhibit the expression of Notch by transporting miR-137, which reduces neuronal apoptosis and attenuates ischemia-reperfusion brain injury.76 Thus, the protective effect of M2-Exos on neurons and the reduction of brain damage, mediated by miRNAs, offer new therapeutic solutions for cerebral ischemia-reperfusion injury.
Multiple Sclerosis Therapy
Multiple sclerosis (MS) is a central nervous system (CNS) demyelinating disease of unknown origin, marked by damage to myelin, difficulties in repair, loss of neurons, and worsening neurological symptoms. The lesion areas contain numerous oligodendrocyte precursor cells (OPCs) as well as activated resident microglia and infiltrating macrophages. Exosomes derived from activated microglia carry miR-615-5p to OPCs, where they bind to the myelin regulatory factor (MYRF), an important transcription factor for myelin production, and hinder the maturation of OPCs. Reducing the expression of miR-615-5p in microglia can slow the progression of the disease, enhance the maturation of oligodendrocytes (OLGs), and encourage the regeneration of myelin.77 Consequently, targeting miR-615-5p in exosomes derived from microglia could be a viable approach for treating multiple sclerosis (MS) by enhancing myelin regeneration and acting as a diagnostic marker for the disease. While research on miRNA in M2-Exos related to MS is still scarce, we are optimistic that the delivery and therapeutic properties of exosomes indicate a strong potential for future use.
Repairing for Circulatory System Injury
Myocardial Injury Therapy
Myocardial ischemic injury typically manifests as either ischemic necrosis or temporary dysfunction of heart muscle cells, accompanied by the infiltration of macrophages. In a study using a rabbit model of myocardial ischemia, Dai et al discovered that miR-148 delivered via M2-Exos reduced the levels of thioredoxin-interacting protein (TXNIP) and inhibited the activation of the TLR4/NF-κB/NLRP3 pathway, thereby protecting heart tissue from damage caused by ischemia.78 In cases of acute myocardial infarction (AMI), Long et al found that M2-Exos delivered miR-1271-5p, which downregulated the expression of SOX6, a direct target, leading to a decrease in hypoxia-induced cell death and lessening cardiac injury in AMI.79 Additionally, Wei et al demonstrated that M2-Exos containing miR-145-5p prevented the death of heart muscle cells caused by hypoxia-reoxygenation (H/R) by lowering TLR4 expression.80 Regarding post-sepsis myocardial injury, Sun et al reported that miR-24-3p within M2-Exos can impart cardioprotection by decreasing the expression of Tnfsf10.81 Recent research highlights that the regenerative ability of adult cardiomyocytes is very limited. However, miRNAs in M2-Exos can safeguard cardiac muscle by minimizing cardiomyocyte injury, inhibiting apoptosis and pyroptosis in these cells, and curbing fibroblast proliferation, which presents opportunities for developing biologically-based therapies to repair myocardial damage.
Atherosclerosis Therapy
Current therapeutic strategies for AS aim to decrease inflammation and prevent plaque rupture by modulating the M2 macrophage polarization in the arterial wall.82,83 The exosomes isolated from the supernatant of bone marrow-derived macrophages (BMDMs) polarize into M2-type, containing anti-inflammatory microRNA-99a/146b/378a, inhibit inflammation by targeting NF-κB and TNF-α signaling and also promoted M2 polarization in recipient macrophages.84 Cheng et al demonstrated that overexpressing miR-221-3p in M2-Exos can attenuate the injury-induced inflammatory response and apoptosis of endothelial cells.85 Additionally, Xie et al developed P-M2EV (platelet membrane-modified M2EV), which contains miR-99a-5p, the most highly expressed miRNA, capable of targeting the mRNA of Homeobox A1 (HOXA1).86 This targeting effectively suppresses the formation of foam cells in vitro and successfully inhibits the progression of AS. This targeting effectively inhibits foam cell formation in vitro and slows the progression of AS. To summarize, the miRNAs from M2-Exos have potential therapeutic benefits for mitigating AS and preventing its advancement through various mechanisms, including reducing inflammation in arterial plaques, decreasing the inflammatory response and apoptosis in injured endothelial cells, and inhibiting foam cell formation.
Repairing for Urinary System Injury
Diabetic Nephropathy Therapy
Diabetic nephropathy (DN) is a frequent complication of diabetes mellitus, characterized by elevated glucose levels that can trigger pro-inflammatory M1-type macrophages, resulting in podocyte apoptosis due to the release of inflammatory mediators. Wang et al discovered that M2-Exos can reduce lipopolysaccharide (LPS)-induced podocyte apoptosis by modulating the miR-93-5p/TLR4 pathway.87 Similarly, Huang et al showed that miR-25-3p found in M2-Exos promotes cellular autophagy, which decreases DUSP1 expression and mitigates podocyte damage caused by high glucose (HG) levels.55 Zhuang et al reported that γ-aminobutyric acid (GABA) can reverse the polarization of M1/M2 macrophages in HG conditions, reducing podocyte injury through the miR-21a-5p-Tnpo1/miR-25-3p-ATXN3 signaling pathway in M2-Exos.88 The miRNAs present in M2-Exos are effective in minimizing podocyte damage and slowing the progression of DN. These results offer new approaches for the prevention and treatment of DN, although additional research is necessary to explore the underlying mechanisms and pathways.
Post-Traumatic Urethral Stricture Therapy
Post-traumatic urethral stricture poses a significant challenge for both patients and healthcare providers. Focusing on glutamine metabolism to curb the excessive activation of urethral fibroblasts (UFBs) offers a promising and effective method to avert urethral scarring and stricture. Research has indicated that miR-381 found in M2-Exos can diminish the formation of myofibroblasts in UFBs by inhibiting YAP/GLS1-dependent glutamine breakdown, thereby preventing urethral scarring and stricture.89 Therefore, this strategy may be a viable therapeutic option for preventing urethral stricture.
Repairing for Skin Injury
Research has demonstrated that M2-Exos have a strong ability to promote angiogenesis during the healing of skin injuries.90–92 Lyu et al found that M2-Exos can enhance the vascularization of endothelial cells by transferring miR-21, which suppresses PTEN expression in these cells and activates the AKT/mTOR signaling pathway.15 Additionally, the researchers utilized a mouse model with complete skin defects and discovered that M2-Exos significantly expedited the healing of traumatic wounds due to their remarkable angiogenic properties, making them a promising clinical treatment for skin injuries. However, there is still a significant lack of understanding regarding the miRNAs present in these exosomes and the regulatory mechanisms involved.
Repairing for Locomotor System Injury
Bone Injury Therapy
Approximately 90% of fractures generally heal well with timely treatment and rest, but a small number of patients may face complications such as nonunion or delayed healing.93 Osteoblast differentiation plays an important role in fracture healing, with macrophages serving as crucial mediators between the inflammatory response and bone regeneration. M2-Exos can promote tissue repair by reducing inflammation and promoting osteoblast differentiation.94,95 Research by Xiong et al revealed that miR-5106, which is prevalent in M2-Exos, is transferred to bone mesenchymal stem cells (BMSCs) and targets the SIK2 and SIK3 genes to facilitate osteoblast differentiation.96 Furthermore, the equilibrium between osteogenesis and lipogenesis in BMSCs is vital for bone formation. It has been reported that M2-Exos can enhance osteogenesis while inhibiting adipogenesis via the miR-690/IRS-1/TAZ pathway, presenting a promising therapeutic approach for bone loss conditions.97 Additionally, Zhang et al found that miRNA-26a-5p found in M2-Exos can also promote osteogenesis and suppress lipogenesis in BMSCs.98
Radiation therapy is a successful method for treating cancer, but it can harm the healthy tissues nearby. Bone tissue is particularly vulnerable to radiation. Bone marrow mesenchymal stem cells (BMMSCs) are also affected by radiation, which may link them to radiation-related bone damage. Huang et al demonstrated that miR-142-3p from M2-Exos can help restore the normal differentiation balance of irradiated BMMSCs by targeting TGF-β1. This discovery highlights a potential pathway for a cell-free strategy to address radiation-induced bone injuries.99
Rheumatoid arthritis (RA) is an autoimmune condition that leads to inflammation and pain in the joints, eventually resulting in bone damage. Pascual-García et al reviewed the role of miRNAs found in the exosomes of osteoclasts, which are derived from the blood’s monocyte-macrophage system, to inform treatment strategies for bone damage caused by RA.100 They found that blocking miR-23a, miR-29b, and miR-214 in osteoclast exosomes promotes the growth of osteoblasts, while increasing levels of miR-22-3p, miR-26a, miR-27a, miR-29a, miR-125b, and miR-146a can inhibit osteoblast activity. Additionally, they explored potential therapeutic targets for these exosomal miRNAs and conducted tests in mice and in vitro RA models. Their findings indicate that these approaches could be significant in restoring the balance of osteoclast and osteoblast differentiation, aiding in the treatment of bone damage associated with RA.
Articular Cartilage and Muscle Injuries Therapy
Osteoarthritis (OA) is a prevalent age-related condition that impacts the entire joint structure, including the articular cartilage and subchondral bone.101 Qian et al found that miR-26b-5p from M2-Exos targets the TLR3 signaling pathway in vitro, which enhances the polarization of M2 macrophages and inhibits the hypertrophy of chondrocytes induced by M1 macrophage-conditioned medium by targeting COL10A1.102 In an OA animal model, miR-26b-5p improved gait abnormalities and alleviated pain, while also reducing synovitis and cartilage degeneration, thereby slowing the progression of OA. Additionally, Zhou et al demonstrated that miR-501, which is abundant in M2-Exos, can promote the differentiation of C2C12 myoblasts by targeting YY1.103 Their results offer new insights into how M2 macrophages can enhance myogenesis and indicate that delivering miR-501 through M2-Exos could be a promising treatment for muscle injury-related conditions.
In summary, the miRNAs from M2-Exos can facilitate healing and hinder the advancement of damage in the respiratory, circulatory, urinary, central nervous, and motor systems, as well as other systemic organ injuries by influencing various pathways. Research into the mechanisms and therapeutic benefits of these miRNAs offers a promising approach for addressing related health issues and establishes a basis for utilizing M2-Exos in the treatment of different diseases. Nonetheless, more comprehensive research and confirmation of the mechanisms involved are necessary, along with improvements in the biosafety of their use.
Application Methods of M2-Exos in Tissue Injury Repair
Direct Exertion of M2-Exos in Tissue Injury Repair
Numerous significant therapeutic outcomes have been documented through the direct utilization of M2-Exos in the repair of tissue injuries (Figure 6).75,104 For instance, exosomes derived from macrophages can be administered at the site of tendon injuries to stimulate the intrinsic reparative capabilities of the tissues, thereby facilitating effective tendon repair.105 Additionally, M2-Exos has demonstrated anti-inflammatory properties by inhibiting the release of pro-inflammatory enzymes and cytokines, while also promoting angiogenesis and re-epithelialization in diabetic wounds, which accelerates the processes of wound healing and flap survival.14,106,107 Despite the significant therapeutic potential of M2-Exos, their advancement is impeded by challenges such as low yield and high production costs. Consequently, researchers typically pursue several strategies to enhance their application: 1) Modifying the culture conditions of exosome-producing cells to increase exosome secretion.108 2) Employing the most suitable extraction method tailored to the specific characteristics of the exosomes. 3) Enhancing therapeutic efficacy through pretreatment aimed at improving exosome uptake by the donor.109 4) Achieving synergistic effects by co-delivering additional drug components.110 5) The integration of multiple distinct exosomes may facilitate the attainment of synergistic therapeutic outcomes.111
 |
Figure 6 Application methods of M2-Exos in tissue injury repair. |
M2-Exos as Drug Delivery for Tissue Injury Repair
Under physiological conditions, extracellular vesicles, including exosomes, serve as mediators of intercellular communication, as they transport various bioactive components that can influence recipient cells.112,113 In the context of atherosclerosis (AS), macrophages transform into foam cells, leading to the release of substantial amounts of pro-inflammatory factors. Exosomes engineered from modified M2 macrophages, in conjunction with the FDA-approved compound 5-aminolevulinic acid hexyl ester hydrochloride (HAL), exhibit the anti-inflammatory properties characteristic of M2 macrophages, with HAL further augmenting their anti-inflammatory effects.114 However, the therapeutic applications of exosomes have often been limited in effectiveness. To address this issue, Li et al developed an integrated hydrogel system incorporating functionalized gold nanorods (AuNRs) and M2-derived exosomes. This system facilitates wound angiogenesis through the sustained release of M2 exosomes, while simultaneously scavenging reactive oxygen species, inhibiting inflammation, promoting angiogenesis, and exhibiting antimicrobial properties, thereby accelerating the wound healing process.107
Several factors underscore the potential of exosome-based drug delivery systems. Firstly, the exosomal membrane is composed of a relatively impermeable lipid bilayer that regulates and prolongs the release of encapsulated substances, including therapeutic agents. Moreover, the surface of exosomes is adorned with various adhesion proteins that exhibit low immunogenicity and cytotoxicity, particularly in the case of autologous exosomes. This characteristic not only facilitates their efficient cellular uptake and the transport of therapeutic drugs but also protects against immune rejection.115 Additionally, exosomes can enhance the stability of certain drug monomers by increasing their resistance to enzymatic degradation and reducing their solubility, thereby preventing rapid degradation in the bloodstream. Consequently, exosomes represent an ideal and distinctive platform for the encapsulation and delivery of drug molecules.
Engineered M2-Exos in Tissue Injury Repair
Exosomes possess the capacity to elicit both adaptive and innate immune responses, rendering them advantageous for the development of therapeutic agents. Nonetheless, challenges persist in the utilization of exosomes, including inadequate delivery efficiency and suboptimal targeting, which impose certain constraints on their application. Various strategies exist that may enhance their therapeutic efficacy, particularly in the context of engineered exosomes.115–117
Genetic engineering techniques enable the modification of ligands on the exosomal membrane, thereby enhancing the specificity and prolonging the in vivo action of exosomes.54,118–120 Numerous studies have employed these techniques to create engineered exosomes with specific functionalities. For instance, Liu utilized CD47 and human antigen R (HuR) to alter the exosomal membrane, transforming it into an effective drug delivery vehicle.121 Additionally, the modulation of gene expression within exosomes through genetic engineering can influence their functional capabilities to achieve targeted therapeutic outcomes. Huang et al exemplified this by developing genetically engineered M2-Exos that were designed to silence casein kinase 2 interacting protein-1 (Ckip-1), which subsequently rescued the mineralization and cementogenesis suppressed by Pg, thus promoting the regeneration of cementum.122 To further improve the targeted delivery efficacy of exosomes, researchers have increasingly focused on integrating exosomes with synthetic nano-microspheres for targeted therapeutic applications. For example, Zeng created a dual-layer microneedle-based wound dressing system (MEs@PMN) that encapsulates micelles in the needle tip and polydopamine (PDA) nanoparticles in the backing layer, which collectively enhances pro-angiogenic effects by elevating the expression of CD31 and von Willebrand factor (vWF).123
Limitations and Future Perspectives
Current methodologies for the isolation of exosomes yield satisfactory levels of both quantity and purity; however, the absence of standardized protocols for their extraction and characterization poses significant challenges for subsequent applications.124 The properties of exosomes, such as size and surface charge, vary depending on the extraction technique employed, which subsequently influences the drug loading capacity and encapsulation efficiency for future drug delivery systems. For exosomes to be effectively utilized in clinical settings, it is imperative to develop standardized extraction and validation protocols. We advocate for researchers to take into account various factors that could enhance the efficacy of exosome-based therapies during their investigations. This includes the establishment of criteria for the selection of appropriate exosomes and the identification of optimal drug delivery timing to maximize therapeutic outcomes.
Additionally, there exists a substantial disparity between the actual therapeutic effectiveness and the levels required for clinical implementation.125 Numerous researchers have suggested strategies to enhance therapeutic efficacy through the synergistic systemic effects of multiple active components, as well as by modifying the exosomal membrane to improve the uptake rate by receptor cells. Furthermore, the integration of exosomes with nanoengineering techniques, such as the incorporation of iron oxide and nanogold, has augmented the translational capacity of exosomes in clinical settings, indicating substantial research potential within the realm of interdisciplinary medicine. We contend that utilizing exosomes as vehicles for therapeutic drug delivery presents considerable promise, particularly in terms of enabling controlled release and minimizing the frequency of drug administration.
Exosomes are usually kept at −80°C for experiments to be used later. However, research has shown that multiple freeze-thaw cycles can alter the size, structure, and function of exosomes, which can affect future studies. Thus, preserving the activity of the bilayer lipid membrane is a matter that needs more exploration. For clinical applications, ensuring their safety is crucial, as is confirming their stable activity (therapeutic effectiveness). If exosomes are to be utilized as treatments for humans, their safety must be guaranteed. In China, exosomes are currently in Phase III clinical trials involving human participants; if they successfully pass further evaluations, they could be approved as drugs. Nonetheless, a few reports have indicated that significant allergic reactions can occur after facial injections of a standard dose of exosomes. Such severe allergic reactions can pose risks to human health and even life, highlighting the need for thorough testing and protocols.
Conclusion
The process of repairing tissue damage is complex and dynamic, involving various cell types and molecular signals, with macrophages being particularly important. Exosomes have garnered significant research interest, both nationally and internationally, due to their wide range of miRNAs. This review examines the role of miRNAs from M2-Exos in diagnosing, preventing, and treating diseases related to tissue injury. Although there has been extensive research on miRNAs from M2 macrophages, challenges remain, especially in improving yield and therapeutic effectiveness. Current studies are focused on creating drug-loaded and engineered exosomes to enhance their efficacy, showing promising results. In conclusion, miRNAs found in M2-Exos hold great potential for aiding tissue repair and could serve as novel targets for disease diagnosis and treatment, offering significant therapeutic opportunities for the future.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This research was supported by the Zhejiang Provincial Natural Science Foundation of China under Grant No.LGF22H150002.
Disclosure
The authors declare that they have no competing interests.
References
1. Eming SA, Wynn TA, Martin P. Inflammation and metabolism in tissue repair and regeneration. Science. 2017;356(6342):1026–1030. doi:10.1126/science.aam7928
2. Wynn TA, Vannella KM. Macrophages in Tissue Repair, Regeneration, and Fibrosis. Immunity. 2016;44(3):450–462. doi:10.1016/j.immuni.2016.02.015
3. Shrivastava R, Shukla N. Attributes of alternatively activated (M2) macrophages. Life Sci. 2019;224:222–231. doi:10.1016/j.lfs.2019.03.062
4. Gordon S. Phagocytosis: an immunobiologic process. Immunity. 2016;44(3):463–475. doi:10.1016/j.immuni.2016.02.026
5. Witherel CE, Sao K, Brisson BK, et al. Regulation of extracellular matrix assembly and structure by hybrid M1/M2 macrophages. Biomaterials. 2021;269:120667. doi:10.1016/j.biomaterials.2021.120667
6. Locati M, Curtale G, Mantovani A. Diversity, mechanisms, and significance of macrophage plasticity. Annu Rev Pathol. 2020;15:123–147. doi:10.1146/annurev-pathmechdis-012418-012718
7. Tu YC, Wang YM, Yao LJ. Macrophage-targeting DNA nanomaterials: a future direction of biological therapy. Int J Nanomed. 2024;19:3641–3655. doi:10.2147/IJN.S459288
8. Yan W, Li T, Yin T, et al. M2 macrophage-derived exosomes promote the c-KIT phenotype of vascular smooth muscle cells during vascular tissue repair after intravascular stent implantation. Theranostics. 2020;10(23):10712–10728. doi:10.7150/thno.46143
9. Gao ZS, Zhang CJ, Xia N, et al. Berberine-loaded M2 macrophage-derived exosomes for spinal cord injury therapy. Acta Biomater. 2021;126:211–223. doi:10.1016/j.actbio.2021.03.018
10. Mazini L, Rochette L, Hamdan Y, Malka G. Skin immunomodulation during regeneration: emerging new targets. J Pers Med. 2021;11(2):85. doi:10.3390/jpm11020085
11. Fawcett S, Al Kassas R, MD I, Hughes AT, Ghali F, Ross K. A time to heal: microRNA and circadian dynamics in cutaneous wound repair. Clin Sci. 2022;136(8):579–597. doi:10.1042/CS20220011
12. Zhao W, He X, Liu R, Ruan Q. Accelerating corneal wound healing using exosome-mediated targeting of NF-κB c-Rel. Inflamm regen. 2023;43(1):6. doi:10.1186/s41232-023-00260-y
13. von Stade DP, Meyers M, Johnson J, et al. Exosome cell origin affects in vitro markers of tendon repair in ovine macrophages and tenocytes. Tissue Eng Part A. 2023;29(9–10):282–291. doi:10.1089/ten.tea.2022.0185
14. Teng L, Maqsood M, Zhu M, et al. Exosomes derived from human umbilical cord mesenchymal stem cells accelerate diabetic wound healing via promoting m2 macrophage polarization, angiogenesis, and collagen deposition. Int J Mol Sci. 2022;23(18):10421. doi:10.3390/ijms231810421
15. Lyu L, Cai Y, Zhang G, et al. Exosomes derived from M2 macrophages induce angiogenesis to promote wound healing. Front Mol Biosci. 2022;9:1008802. doi:10.3389/fmolb.2022.1008802
16. Hou YY, Liu YT, Liang S, et al. The novel target: exosoms derived from M2 macrophage. Int Rev Immunol. 2020;40:183–196.
17. Lin CW, Hung CM, Chen WJ, et al. New horizons of macrophage immunomodulation in the healing of diabetic foot ulcers. Pharmaceutics. 2022;14(10):2065. doi:10.3390/pharmaceutics14102065
18. Mori MA, Ludwig RG, Garcia-Martin R, Brandão BB, Kahn CR. Extracellular miRNAs: from biomarkers to mediators of physiology and disease. Cell Metab. 2019;30(4):656–673. doi:10.1016/j.cmet.2019.07.011
19. Kalluri R, LeBleu VS. The biology, function, and biomedical applications of exosomes. Science. 2020;367(6478). doi:10.1126/science.aau6977
20. van Niel G, D’Angelo G, Raposo G. Shedding light on the cell biology of extracellular vesicles. Nat Rev mol Cell Biol. 2018;19(4):213–228. doi:10.1038/nrm.2017.125
21. Liu J, Ren L, Li S, et al. The biology, function, and applications of exosomes in cancer. Acta Pharmaceutica Sinica B. 2021;11(9):2783–2797. doi:10.1016/j.apsb.2021.01.001
22. Mathieu M, Martin-Jaular L, Lavieu G, Théry C. Specificities of secretion and uptake of exosomes and other extracellular vesicles for cell-to-cell communication. Nat Cell Biol. 2019;21(1):9–17. doi:10.1038/s41556-018-0250-9
23. David G, Zimmermann P. Heparanase Involvement in Exosome Formation. Adv Exp Med Biol. 2020;1221:285–307.
24. Kamerkar S, LeBleu VS, Sugimoto H, et al. Exosomes facilitate therapeutic targeting of oncogenic KRAS in pancreatic cancer. Nature. 2017;546(7659):498–503. doi:10.1038/nature22341
25. Song J, Song B, Yuan L, Yang G. Multiplexed strategies toward clinical translation of extracellular vesicles. Theranostics. 2022;12(15):6740–6761. doi:10.7150/thno.75899
26. Morishita M, Takahashi Y, Nishikawa M, Takakura Y. Pharmacokinetics of exosomes-an important factor for elucidating the biological roles of exosomes and for the development of exosome-based therapeutics. J Pharmaceut Sci. 2017;106(9):2265–2269. doi:10.1016/j.xphs.2017.02.030
27. Mendt M, Kamerkar S, Sugimoto H, et al. Generation and testing of clinical-grade exosomes for pancreatic cancer. JCI Insight. 2018;3(8). doi:10.1172/jci.insight.99263
28. Gangadaran P, Hong CM, Ahn BC. An update on in vivo imaging of extracellular vesicles as drug delivery vehicles. Front Pharmacol. 2018;9:169. doi:10.3389/fphar.2018.00169
29. Willms E, Cabañas C, Mäger I, Wood MJA, Vader P. Extracellular vesicle heterogeneity: subpopulations, isolation techniques, and diverse functions in cancer progression. Front Immunol. 2018;9:738. doi:10.3389/fimmu.2018.00738
30. Zhang H, Freitas D, Kim HS, et al. Identification of distinct nanoparticles and subsets of extracellular vesicles by asymmetric flow field-flow fractionation. Nat Cell Biol. 2018;20(3):332–343. doi:10.1038/s41556-018-0040-4
31. Medley JC, Panzade G, Zinovyeva AY. microRNA strand selection: unwinding the rules. Wiley Interdiscip Rev RNA. 2021;12(3):e1627. doi:10.1002/wrna.1627
32. Gerlach CV, Vaidya VS. MicroRNAs in injury and repair. Arch Toxicol. 2017;91(8):2781–2797. doi:10.1007/s00204-017-1974-1
33. Cai Y, Yu X, Hu S, Yu J. A brief review on the mechanisms of miRNA regulation. Genom Proteom Bioinform. 2009;7(4):147–154. doi:10.1016/S1672-0229(08)60044-3
34. Fabian MR, Sonenberg N, Filipowicz W. Regulation of mRNA translation and stability by microRNAs. Annu Rev Biochem. 2010;79:351–379. doi:10.1146/annurev-biochem-060308-103103
35. Du SW, Palczewski K. MicroRNA regulation of critical retinal pigment epithelial functions. Trends Neurosci. 2022;45(1):78–90. doi:10.1016/j.tins.2021.10.008
36. Vasudevan S, Steitz JA. AU-rich-element-mediated upregulation of translation by FXR1 and argonaute 2. Cell. 2007;128(6):1105–1118. doi:10.1016/j.cell.2007.01.038
37. Ørom UA, Nielsen FC, Lund AH. MicroRNA-10a binds the 5’UTR of ribosomal protein mRNAs and enhances their translation. Molecular Cell. 2008;30(4):460–471. doi:10.1016/j.molcel.2008.05.001
38. O’Brien J, Hayder H, Zayed Y, Peng C. Overview of MicroRNA biogenesis, mechanisms of actions, and circulation. Front Endocrinol. 2018;9:402. doi:10.3389/fendo.2018.00402
39. Jiang Y, Xu X, Xiao L, Wang L, Qiang S. The role of microRNA in the inflammatory response of wound healing. Front Immunol. 2022;13:852419. doi:10.3389/fimmu.2022.852419
40. Banerjee J, Chan YC, Sen CK. MicroRNAs in skin and wound healing. Physiol Genom. 2011;43(10):543–556. doi:10.1152/physiolgenomics.00157.2010
41. Zhang Y, Bi J, Huang J, Tang Y, Du S, Li P. Exosome: a review of its classification, isolation techniques, storage, diagnostic and targeted therapy applications. Int J Nanomed. 2020;15:6917–6934. doi:10.2147/IJN.S264498
42. Livshits MA, Khomyakova E, Evtushenko EG, et al. Isolation of exosomes by differential centrifugation: theoretical analysis of a commonly used protocol. Sci Rep. 2015;5:17319. doi:10.1038/srep17319
43. Coumans FAW, Brisson AR, Buzas EI, et al. Methodological guidelines to study extracellular vesicles. Cir Res. 2017;120(10):1632–1648. 1524-4571 (Electronic). doi:10.1161/CIRCRESAHA.117.309417
44. Moghadasi S, Elveny M, Rahman HS, et al. A paradigm shift in cell-free approach: the emerging role of MSCs-derived exosomes in regenerative medicine. J Transl Med. 2021;19(1):302. doi:10.1186/s12967-021-02980-6
45. Conlan RS, Pisano S, Oliveira MI, Ferrari M, Mendes Pinto I. Exosomes as reconfigurable therapeutic systems. Trends Mol Med. 2017;23(7):636–650. doi:10.1016/j.molmed.2017.05.003
46. Vergauwen G, Dhondt B, Van Deun J, et al. Confounding factors of ultrafiltration and protein analysis in extracellular vesicle research. Sci Rep. 2017;7(1):2704. doi:10.1038/s41598-017-02599-y
47. Haraszti RA, Miller R, Stoppato M, et al. Exosomes produced from 3D cultures of MSCs by tangential flow filtration show higher yield and improved activity. Mol Ther. 2018;26(12):2838–2847. doi:10.1016/j.ymthe.2018.09.015
48. Lin Y, Anderson JD, Rahnama LMA, Gu SV, Knowlton AA. Exosomes in disease and regeneration: biological functions, diagnostics, and beneficial effects. Am J Physiol Heart Circ Physiol. 2020;319(6):H1162–h1180. doi:10.1152/ajpheart.00075.2020
49. Böing AN, van der Pol E, Grootemaat AE, Coumans FA, Sturk A, Nieuwland R. Single-step isolation of extracellular vesicles by size-exclusion chromatography. J Extracell Vesicles. 2014;3:234380.
50. Zhou X, Brown BA, Siegel AP, et al. Exosome-mediated crosstalk between keratinocytes and macrophages in cutaneous wound healing. ACS Nano. 2020;14(10):12732–12748. doi:10.1021/acsnano.0c03064
51. Heath N, Grant L, De Oliveira TM, et al. Rapid isolation and enrichment of extracellular vesicle preparations using anion exchange chromatography. Sci Rep. 2018;8(1):5730. doi:10.1038/s41598-018-24163-y
52. Prattichizzo F, Matacchione G, Giuliani A, et al. Extracellular vesicle-shuttled miRNAs: a critical appraisal of their potential as nano-diagnostics and nano-therapeutics in type 2 diabetes mellitus and its cardiovascular complications. Theranostics. 2021;11(3):1031–1045. doi:10.7150/thno.51605
53. Munagala R, Aqil F, Jeyabalan J, et al. Exosome-mediated delivery of RNA and DNA for gene therapy. Cancer Lett. 2021;505:58–72. doi:10.1016/j.canlet.2021.02.011
54. Fan MH, Pi JK, Zou CY, et al. Hydrogel-exosome system in tissue engineering: a promising therapeutic strategy. Bioact Mater 2024;38:1–30. doi:10.1016/j.bioactmat.2024.04.007
55. Huang H, Liu H, Tang J, et al. M2 macrophage-derived exosomal miR-25-3p improves high glucose-induced podocytes injury through activation autophagy via inhibiting DUSP1 expression. IUBMB Life. 2020;72(12):2651–2662. doi:10.1002/iub.2393
56. Visan KS, Lobb RJ, Ham S, et al. Comparative analysis of tangential flow filtration and ultracentrifugation, both combined with subsequent size exclusion chromatography, for the isolation of small extracellular vesicles. J Extracell Vesicles. 2022;11(9):e12266. doi:10.1002/jev2.12266
57. Seo N, Nakamura J, Kaneda T, et al. Distinguishing functional exosomes and other extracellular vesicles as a nucleic acid cargo by the anion-exchange method. J Extracell Vesicles. 2022;11(3):e12205. doi:10.1002/jev2.12205
58. Shirejini SZ, Inci F. The Yin and Yang of exosome isolation methods: conventional practice, microfluidics, and commercial kits. Biotechnol Adv 2022;54:107814. doi:10.1016/j.biotechadv.2021.107814
59. Xiao Y, Zhong J, Zhong B, et al. Exosomes as potential sources of biomarkers in colorectal cancer. Cancer Lett. 2020;476:13–22. doi:10.1016/j.canlet.2020.01.033
60. Ramanathan S, Shenoda BB, Lin Z, et al. Inflammation potentiates miR-939 expression and packaging into small extracellular vesicles. J Extracell Vesicles. 2019;8(1):1650595. doi:10.1080/20013078.2019.1650595
61. Lin B, Lei Y, Wang J, et al. Microfluidic-based exosome analysis for liquid biopsy. Small Methods. 2021;5(3):e2001131. doi:10.1002/smtd.202001131
62. Binenbaum Y, Fridman E, Yaari Z, et al. Transfer of miRNA in macrophage-derived exosomes induces drug resistance in pancreatic adenocarcinoma. Cancer Res. 2018;78(18):5287–5299. doi:10.1158/0008-5472.CAN-18-0124
63. Dong X, Lin Y, Li K, et al. Consensus statement on extracellular vesicles in liquid biopsy for advancing laboratory medicine. Clin Chem Lab Med. 2024;63(3):465–482. doi:10.1515/cclm-2024-0188
64. Zhang W, Zhou R, Liu X, et al. Key role of exosomes derived from M2 macrophages in maintaining cancer cell stemness (Review). Int J Oncol. 2023;63(5). doi:10.3892/ijo.2023.5574
65. Skliar M, Chernyshev VS. Imaging of extracellular vesicles by atomic force microscopy. J Visualized Exp. 2019;151. doi:10.3791/59254-v
66. Sluijter JPG, Davidson SM, Boulanger CM, et al. Extracellular vesicles in diagnostics and therapy of the ischaemic heart: position paper from the working group on cellular biology of the heart of the European society of cardiology. Cardiovasc Res. 2018;114(1):19–34. doi:10.1093/cvr/cvx211
67. Yang J, Tang X, Chen L, et al. Liver-derived exosomal miRNA in NAFLD: mechanisms of action, biomarkers, and therapeutic applications. Curr Med Chem 2024;31. doi:10.2174/0109298673276581231210170332
68. Yao MY, Zhang WH, Ma WT, Liu QH, Xing LH, Zhao GF. microRNA-328 in exosomes derived from M2 macrophages exerts a promotive effect on the progression of pulmonary fibrosis via FAM13A in a rat model. Exp Mol Med 2019;51(6):1–16. doi:10.1038/s12276-019-0255-x
69. Kishore A, Petrek M. Roles of macrophage polarization and macrophage-derived miRNAs in pulmonary fibrosis. Front Immunol. 2021;12:678457. doi:10.3389/fimmu.2021.678457
70. Guiot J, Cambier M, Boeckx A, et al. Macrophage-derived exosomes attenuate fibrosis in airway epithelial cells through delivery of antifibrotic miR-142-3p. Thorax. 2020;75(10):870–881. doi:10.1136/thoraxjnl-2019-214077
71. Li C, Deng C, Zhou T, et al. MicroRNA-370 carried by M2 macrophage-derived exosomes alleviates asthma progression through inhibiting the FGF1/MAPK/STAT1 axis. Int J Bio Sci. 2021;17(7):1795–1807. doi:10.7150/ijbs.59715
72. Tang B, Wu Y, Zhang Y, Cheng Y, Wu Y, Fang H. Scorpion and centipede alleviates severe asthma through M2 macrophage-derived exosomal miR-30b-5p. Aging. 2022;14(9):3921–3940. doi:10.18632/aging.204053
73. Xu X, Gao W, Li L, et al. Annexin A1 protects against cerebral ischemia-reperfusion injury by modulating microglia/macrophage polarization via FPR2/ALX-dependent AMPK-mTOR pathway. J Neuroinflamm. 2021;18(1):119. doi:10.1186/s12974-021-02174-3
74. Saeedi S, Israel S, Nagy C, Turecki G. The emerging role of exosomes in mental disorders. Transl Psych. 2019;9(1):122. doi:10.1038/s41398-019-0459-9
75. Song Y, Li Z, He T, et al. M2 microglia-derived exosomes protect the mouse brain from ischemia-reperfusion injury via exosomal miR-124. Theranostics. 2019;9(10):2910–2923. doi:10.7150/thno.30879
76. Zhang D, Cai G, Liu K, et al. Microglia exosomal miRNA-137 attenuates ischemic brain injury through targeting Notch1. Aging. 2021;13(3):4079–4095. doi:10.18632/aging.202373
77. Ji XY, Guo YX, Wang LB, et al. Microglia-derived exosomes modulate myelin regeneration via miR-615-5p/MYRF axis. J Neuroinflamm. 2024;21(1). 1742-2094 (Electronic). doi:10.1186/s12974-024-03019-5
78. Dai Y, Wang S, Chang S, et al. M2 macrophage-derived exosomes carry microRNA-148a to alleviate myocardial ischemia/reperfusion injury via inhibiting TXNIP and the TLR4/NF-κB/NLRP3 inflammasome signaling pathway. J Mol Cell Cardiol. 2020;142:65–79. doi:10.1016/j.yjmcc.2020.02.007
79. Long R, Gao L, Li Y, et al. M2 macrophage-derived exosomes carry miR-1271-5p to alleviate cardiac injury in acute myocardial infarction through down-regulating SOX6. Mol Immunol. 2021;136:26–35. doi:10.1016/j.molimm.2021.05.006
80. Wei L, Zhao D. M2 macrophage-derived exosomal miR-145-5p protects against the hypoxia/reoxygenation-induced pyroptosis of cardiomyocytes by inhibiting TLR4 expression. Ann Translat Med. 2022;10(24):1376. doi:10.21037/atm-22-6109
81. Sun X, Liu Y, Wang J, Zhang M, Wang M. Cardioprotection of M2 macrophages-derived exosomal microRNA-24-3p/Tnfsf10 axis against myocardial injury after sepsis. Mol Immunol. 2022;141:309–317. doi:10.1016/j.molimm.2021.11.003
82. Rahman K, Fisher EA. Insights from pre-clinical and clinical studies on the role of innate inflammation in atherosclerosis regression. Front Cardiovasc Med. 2018;5:32. doi:10.3389/fcvm.2018.00032
83. Bäck M, Yurdagul A Jr, Tabas I, Öörni K, Kovanen PT. Inflammation and its resolution in atherosclerosis: mediators and therapeutic opportunities. Nat Rev Cardiol. 2019;16(7):389–406. doi:10.1038/s41569-019-0169-2
84. Bouchareychas L, Duong P, Covarrubias S, et al. Macrophage exosomes resolve atherosclerosis by regulating hematopoiesis and inflammation via MicroRNA cargo. Cell Rep. 2020;32(2):107881. doi:10.1016/j.celrep.2020.107881
85. Cheng X, Zhou H, Zhou Y, Song C. M2 macrophage-derived exosomes inhibit apoptosis of HUVEC cell through regulating miR-221-3p expression. Biomed Res Int 2022;2022:1609244. doi:10.1155/2022/1609244
86. Xie L, Chen J, Hu H, et al. Engineered M2 macrophage-derived extracellular vesicles with platelet membrane fusion for targeted therapy of atherosclerosis. Bioact Mater 2024;35:447–460. doi:10.1016/j.bioactmat.2024.02.015
87. Wang Z, Sun W, Li R, Liu Y. miRNA-93-5p in exosomes derived from M2 macrophages improves lipopolysaccharide-induced podocyte apoptosis by targeting Toll-like receptor 4. Bioengineered. 2022;13(3):7683–7696. doi:10.1080/21655979.2021.2023794
88. Zhuang Y, Zheng H, Yang Y, Ni H. GABA alleviates high glucose-induced podocyte injury through dynamically altering the expression of macrophage M1/M2-derived exosomal miR-21a-5p/miR-25-3p. Biochem Biophys Res Commun 2022;618:38–45. doi:10.1016/j.bbrc.2022.06.019
89. Chen YH, Xu YC, Lin TT, et al. Exosomal MiR-381 from M2-polarized macrophages attenuates urethral fibroblasts activation through YAP/GLS1-regulated glutaminolysis. Inflamm Res. 2023;72(7):1359–1373. doi:10.1007/s00011-023-01735-x
90. Liu P, Xiong Y, Chen L, et al. Angiogenesis-based diabetic skin reconstruction through multifunctional hydrogel with sustained releasing of M2 macrophage-derived exosome. Chem Eng J. 2021;431:132413.
91. Kim H, Wang SY, Kwak G, Yang Y, Kwon IC, Kim SH. Exosome-guided phenotypic switch of M1 to M2 macrophages for cutaneous wound healing. Adv Sci. 2019;6(20):1900513. doi:10.1002/advs.201900513
92. Li M, Wang T, Tian H, Wei G, Zhao L, Shi Y. Macrophage-derived exosomes accelerate wound healing through their anti-inflammation effects in a diabetic rat model. Artif Cells Nanomed Biotechnol 2019;47(1):3793–3803. doi:10.1080/21691401.2019.1669617
93. Hovius SE, de Jong T. Bone grafts for scaphoid nonunion: an overview. Hand Surg. 2015;20(2):222–227. doi:10.1142/S0218810415400043
94. Zhao X, Chen X, Deng Y, et al. A novel adhesive dual-sensitive hydrogel for sustained release of exosomes derived from M2 macrophages promotes repair of bone defects. Mater Today Bio. 2023;23:100840. doi:10.1016/j.mtbio.2023.100840
95. Shou J, Li S, Shi W, et al. 3WJ RNA nanoparticles-aptamer functionalized exosomes from M2 macrophages target BMSCs to promote the healing of bone fractures. Stem Cells Transl Med. 2023;12(11):758–774. doi:10.1093/stcltm/szad052
96. Xiong Y, Chen L, Yan C, et al. M2 Macrophagy-derived exosomal miRNA-5106 induces bone mesenchymal stem cells towards osteoblastic fate by targeting salt-inducible kinase 2 and 3. J Nanobiotechnol. 2020;18(1):66. doi:10.1186/s12951-020-00622-5
97. Li Z, Wang Y, Li S, Li Y. Exosomes derived from M2 macrophages facilitate osteogenesis and reduce adipogenesis of BMSCs. Front Endocrinol. 2021;12:680328. doi:10.3389/fendo.2021.680328
98. Bin-Bin Z, Da-Wa ZX, Chao L, et al. M2 macrophagy-derived exosomal miRNA-26a-5p induces osteogenic differentiation of bone mesenchymal stem cells. J Orthopaedic Surg Res. 2022;17(1):137. doi:10.1186/s13018-022-03029-0
99. Huang C, Zhao L, Xiao Y, et al. M2 macrophage-derived exosomes carry miR-142-3p to restore the differentiation balance of irradiated BMMSCs by targeting TGF-β1. Mol Cell Biochem. 2024;479(4):993–1010. doi:10.1007/s11010-023-04775-3
100. Pascual-García S, Martínez-Peinado P, Pujalte-Satorre C, et al. Exosomal osteoclast-derived miRNA in rheumatoid arthritis: from their pathogenesis in bone erosion to new therapeutic approaches. Int J Mol Sci. 2024;25(3):1506. doi:10.3390/ijms25031506
101. Dai J, Hu Z, Zeng F, et al. Osteoclast-derived exosomal miR-212-3p suppressed the anabolism and accelerated the catabolism of chondrocytes in osteoarthritis by targeting TGF-β1/Smad2 signaling. Arch Biochem Biophys 2024;751:109827. doi:10.1016/j.abb.2023.109827
102. Qian Y, Chu G, Zhang L, et al. M2 macrophage-derived exosomal miR-26b-5p regulates macrophage polarization and chondrocyte hypertrophy by targeting TLR3 and COL10A1 to alleviate osteoarthritis. J Nanobiotechnol. 2024;22(1):72. doi:10.1186/s12951-024-02336-4
103. Zhou M, Li B, Liu C, et al. M2 Macrophage-derived exosomal miR-501 contributes to pubococcygeal muscle regeneration. Int Immunopharmacol. 2021;101(Pt B):108223. doi:10.1016/j.intimp.2021.108223
104. Hou BR, Jiang C, Wang ZN, Ren HJ. Exosome-mediated crosstalk between microglia and neural stem cells in the repair of brain injury. Neural Regen Res. 2020;15(6):1023–1024. doi:10.4103/1673-5374.270302
105. Chamberlain CS, Clements AEB, Kink JA, et al. Extracellular vesicle-educated macrophages promote early achilles tendon healing. Stem Cells. 2019;37(5):652–662. doi:10.1002/stem.2988
106. Luo G, Zhou Z, Cao Z, et al. M2 macrophage-derived exosomes induce angiogenesis and increase skin flap survival through HIF1AN/HIF-1α/VEGFA control. Arch Biochem Biophys 2024;751:109822. doi:10.1016/j.abb.2023.109822
107. Li W, Wu S, Ren L, et al. Development of an antiswelling hydrogel system incorporating M2-exosomes and photothermal effect for diabetic wound healing. ACS Nano. 2023;17(21):22106–22120. doi:10.1021/acsnano.3c09220
108. Phan J, Kumar P, Hao D, Gao K, Farmer D, Wang A. Engineering mesenchymal stem cells to improve their exosome efficacy and yield for cell-free therapy. J Extracell Vesicles. 2018;7(1):1522236. doi:10.1080/20013078.2018.1522236
109. Zhou YK, Han CS, Zhu ZL, et al. M2 exosomes modified by hydrogen sulfide promoted bone regeneration by moesin mediated endocytosis. Bioact Mater 2024;31:192–205. doi:10.1016/j.bioactmat.2023.08.006
110. Chen L, Yu C, Xiong Y, et al. Multifunctional hydrogel enhances bone regeneration through sustained release of stromal cell-derived factor-1α and exosomes. Bioact Mater 2023;25:460–471. doi:10.1016/j.bioactmat.2022.07.030
111. Ma T, Chen S, Wang J, et al. Enhanced osteolysis targeted therapy through fusion of exosomes derived from M2 macrophages and bone marrow mesenchymal stem cells: modulating macrophage polarization. Small. 2023;20(7):e2303506. doi:10.1002/smll.202303506
112. Klyachko NL, Arzt CJ, Li SM, Gololobova OA, Batrakova EV. Extracellular Vesicle-Based Therapeutics: preclinical and Clinical Investigations. Pharmaceutics. 2020;12(12):1171. doi:10.3390/pharmaceutics12121171
113. Hu T, Wolfram J, Srivastava S. Extracellular vesicles in cancer detection: hopes and hypes. Trends Cancer. 2021;7(2):122–133. doi:10.1016/j.trecan.2020.09.003
114. Kuznetsova T, Prange KHM, Glass CK, de Winther MPJ. Transcriptional and epigenetic regulation of macrophages in atherosclerosis. Nat Rev Cardiol. 2020;17(4):216–228. doi:10.1038/s41569-019-0265-3
115. Yan C, Feng K, Bao B, et al. Biohybrid nanorobots carrying glycoengineered extracellular vesicles promote diabetic wound repair through dual-enhanced cell and tissue penetration. Adv Sci. 2024;11:e2404456. doi:10.1002/advs.202404456
116. Kwak G, Cheng J, Kim H, et al. Sustained exosome-guided macrophage polarization using hydrolytically degradable PEG hydrogels for cutaneous wound healing: identification of key proteins and MiRNAs, and sustained release formulation. Small. 2022;18(15):e2200060. doi:10.1002/smll.202200060
117. Sun B, Wu F, Wang X, et al. An optimally designed engineering exosome-reductive COF integrated nanoagent for synergistically enhanced diabetic fester wound healing. Small. 2022;18(26):e2200895. doi:10.1002/smll.202200895
118. Lu M, Xing H, Xun Z, et al. Functionalized extracellular vesicles as advanced therapeutic nanodelivery systems. Eur J Pharm Sci 2018;121:34–46. doi:10.1016/j.ejps.2018.05.001
119. Kim MS, Haney MJ, Zhao Y, et al. Engineering macrophage-derived exosomes for targeted paclitaxel delivery to pulmonary metastases: in vitro and in vivo evaluations. Nanomedicine. 2018;14(1):195–204. doi:10.1016/j.nano.2017.09.011
120. Zhang W, Yu ZL, Wu M, et al. Magnetic and folate functionalization enables rapid isolation and enhanced tumor-targeting of cell-derived microvesicles. ACS Nano. 2017;11(1):277–290. doi:10.1021/acsnano.6b05630
121. Liu S, Xiao X, Zhang L, et al. Reprogramming exosomes to escape from immune surveillance for mitochondrial protection in hepatic ischemia-reperfusion injury. Theranostics. 2024;14(1):116–132. doi:10.7150/thno.88061
122. Huang X, Deng Y, Xiao J, Wang H, Yang Q, Cao Z. Genetically engineered M2-like macrophage-derived exosomes for P. gingivalis-suppressed cementum regeneration: from mechanism to therapy. Bioact Mater 2024;32:473–487. doi:10.1016/j.bioactmat.2023.10.009
123. Zeng J, Sun Z, Zeng F, Gu C, Chen X. M2 macrophage-derived exosome-encapsulated microneedles with mild photothermal therapy for accelerated diabetic wound healing. Mater Today Bio. 2023;20:100649. doi:10.1016/j.mtbio.2023.100649
124. Li X, Corbett AL, Taatizadeh E, et al. Challenges and opportunities in exosome research—Perspectives from biology, engineering, and cancer therapy. APL Bioeng. 2019;3(1):011503. doi:10.1063/1.5087122
125. Meng W, He C, Hao Y, Wang L, Li L, Zhu G. Prospects and challenges of extracellular vesicle-based drug delivery system: considering cell source. Drug Delivery. 2020;27(1):585–598. doi:10.1080/10717544.2020.1748758
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms.php
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms.php
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Exosome-Derived miRNAs as Potential Biomarkers for Prostate Bone Metastasis
Lu Z, Hou J, Li X, Zhou J, Luo B, Liang S, Lo RK, Wong TM, Kuang GM
International Journal of General Medicine 2022, 15:5369-5383
Published Date: 1 June 2022