")
Back to Journals » Drug Design, Development and Therapy » Volume 19
Tempol Mitigates Methotrexate-Induced Osteotoxicity via Oxidative Stress Modulation and MAPK Pathway Inhibition
Authors Selimli F, Taş Reyhanioğlu M, Haskan AC, Altun MS, Mete S , Kaplan HM
Received 4 December 2024
Accepted for publication 17 February 2025
Published 28 February 2025 Volume 2025:19 Pages 1441—1449
DOI https://doi.org/10.2147/DDDT.S510206
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Tamer Ibrahim
Fariz Selimli,1 Meryem Taş Reyhanioğlu,1 Ahmet can Haskan,1 Muhammed Said Altun,1 Soner Mete,2 Halil Mahir Kaplan3
1Department of Oral and Maxillofacial Surgery, Faculty of Dentistry, Mustafa Kemal University, Hatay, Turkey; 2Department of Medical Services and Techniques, Medical Promotion and Marketing Program, Medical Vocational Higher Services School, Nevsehir Haci Bektas University, Nevşehir, Turkey; 3Deparment of Medicinal Pharmacology, Faculty of Medicine, Cukurova University, Adana, Turkey
Correspondence: Fariz Selimli, Department of Oral and Maxillofacial Surgery, Faculty of Dentistry, Mustafa Kemal University, Hatay, 01330, Turkey, Tel +905456744444, Email [email protected]
Purpose: Osteotoxicity, a common consequence of Methotrexate (MTX) therapy, significantly compromises bone health by inducing oxidative stress and disrupting bone remodeling. This study examines the protective effects of Tempol, a nitroxide compound with antioxidant properties, against MTX-induced osteotoxicity.
Methods: Osteocyte-like MLO-Y4 cells were cultured and treated with Tempol and MTX to evaluate changes in apoptotic mediators, MAPK signaling pathways, and oxidative stress parameters.
Results: MTX treatment significantly increased caspase-3 activity and Bax expression while decreasing Bcl-2 levels, thereby creating a pro-apoptotic environment. It also activated stress-related pathways by elevating JNK and ERK activities. Conversely, Tempol effectively countered these effects by restoring the balance of apoptotic mediators, downregulating MAPK activation, and enhancing Total Antioxidant Status (TAS). Additionally, Tempol reduced Total Oxidant Status (TOS) and improved the activities of superoxide dismutase (SOD) and glutathione peroxidase (GPx).
Conclusion: These findings highlight Tempol’s potential to mitigate oxidative stress and apoptosis linked to MTX therapy, supporting its use as an adjunctive treatment to protect bone health in patients undergoing MTX therapy. Emphasizing Tempol’s clinical implications as a protective agent reinforces the urgency for further research into its long-term effects on cellular viability and bone integrity in the context of chemotherapy.
Keywords: tempol, osteotoxicity, methotrexate, oxidative stress and MAPK
Introduction
Osteotoxicity, also known as skeletal toxicity, is defined as any detrimental effect experienced by skeletal tissue during its development and maintenance. This condition occurs as a result of acute or chronic exposure to exogenous substances, which will henceforth be referred to as osteotoxic compounds or osteotoxicants. These substances may be found in the environment or may be ingested through food or introduced through injury. Osteotoxicity can result in morphological deformities and impairments in skeletal processes such as mineralization, remodeling, and morphogenesis.1
Methotrexate (MTX) is known to cause significant bone defects, primarily manifested as reduced bone mineral density (BMD) and an increased likelihood of bone fractures. This is largely due to MTX’s adverse effects on osteoblasts, the cells responsible for bone formation. The drug can impair the proliferation and function of these cells, leading to decreased bone formation and an imbalance in the normal bone remodeling process. Additionally, MTX may promote osteoclastogenesis, resulting in enhanced bone resorption. Consequently, this disruption in the balance between bone formation and resorption contributes to lower BMD, weakening the bones and making them more susceptible to fractures. Given these risks, it is crucial for patients undergoing MTX therapy to be monitored for changes in bone health and to consider preventive measures that could help maintain bone density and overall skeletal integrity.1,2
Recent studies have investigated the role of oxidative stress in mediating the osteotoxic effects of Methotrexate, revealing that excess reactive oxygen species (ROS) can trigger cell apoptosis and inflammatory pathways.1 The increase in reactive oxygen species (ROS), along with altered levels of pro-inflammatory mediators and microRNAs (miRNAs), enhances osteoclastogenesis and reduces osteoblastogenesis through mechanisms associated with the activation of MAPK and transcription factors.3 The Mitogen-Activated Protein Kinase (MAPK) signaling pathway has been implicated in this process, as it is activated in response to oxidative stress and contributes to cellular responses that may exacerbate bone damage.4–6
Tempol (TPL) is a nitroxyl antioxidant with superoxide dismutase (SOD) mimetic activity, allowing it to effectively protect cells by crossing biological membranes. TPL safeguards tissues against the harmful effects of oxidative stress by preventing the formation of hydroxyl radicals.7 In addition to its antioxidant activity at low to moderate concentrations, TPL exhibits pro-oxidant effects at higher concentrations. These elevated levels increase the production of reactive oxygen species (ROS) within cells, thereby inhibiting the growth of cancer cells.8 This mechanism involves promoting apoptosis and halting the cell cycle in cancer cells, suggesting that TPL’s pro-oxidant activity may lead to the development of new potential strategies in cancer therapy. However, given the dual effects of TPL providing both protective and potentially detrimental outcomes it must be carefully evaluated in clinical applications. Consequently, TPL continues to be an important area of research in the management of oxidative stress-related diseases and cancer treatment.
TPL, a nitroxide compound with potent antioxidant properties, has emerged as a promising candidate for mitigating oxidative stress.9,10 By scavenging free radicals and reducing ROS levels, TPL may counteract the harmful effects of Methotrexate on bone cells.11 Moreover, the effect of Tempol on MAPK activity may offer a potential therapeutic avenue to maintain bone health during MTX treatment.
This study aims to explore the protective mechanisms of Tempol against Methotrexate-induced osteotoxicity, focusing on its capacity to diminish oxidative stress and modulate MAPK signaling pathways. Understanding these interactions is vital for developing strategies to enhance patient outcomes and preserve bone integrity in those undergoing Methotrexate therapy.
Materials and Methods
This research was conducted at the Department of Pharmacology, Cukurova University Faculty of Medicine, from January 1, 2023, to May 1, 2024.
Chemicals
The culture media utilized in this study were sourced from GIBCO BRL (Grand Island, NY, U.S.A)., while calf serum was obtained from HyClone Laboratories, Inc. (Logan, UT, U.S.A). Rat tail collagen type I was purchased from Becton Dickinson Laboratories (Bedford, MA, U.S.A). Reagents including Radioimmunoprecipitation Assay (RIPA) buffer, fetal bovine serum, bovine serum albumin, Phosphate-Buffered Saline (PBS), NaCl, TritonX-100, Ethylene Glycol Tetraacetic Acid Ethylene Glycol Tetraacetic Acid (EGTA), dithiothreitol, NaF, Tris–HCl, and Na3VO4 were all acquired from Sigma-Aldrich (St. Louis, MO, U.S.A). Furthermore, ELISA kits for analyzing bax, bcl-2, wee 1, AIF, gadd153, and grp78 were obtained from Shanghai Sunred Biological Technology Co., Ltd. Active ERK (p-ERK) and JNK (p-JNK) detection kits were sourced from My BioSource, Inc. (San Diego, U.S.A). SOD and GPX assay kits were from BioVision (USA), while Total Antioxidant Status (TAS) and Total oxidant Status (TOS) assays were purchased from Rel Assay Diagnostics Inc. (Gaziantep, Turkey). The Bradford dye reagent was obtained from Bio-Rad Laboratories, Inc. (California, U.S.A).
Cell Culture
The murine osteocyte-like cell line MLO-Y4 was acquired from Kerafast, Inc. Cells were cultured according to the methods established by Kato and Bonewald. MLO-Y4 cells were treated with MTX at a concentration of 10−5 M, both alone and in combination with Tempol at 100 μmol/L for a duration of 48 hours. Following treatment, the cells were homogenized for ELISA analysis.
Cell Homogenization
Cells treated with TPL, MTX and TPL+MTX for 48 h in six-well plates were taken into tubes with a volume of 15 mL. After the tubes were centrifuged at 2000 rpm at 4°C for 10 minutes, the liquid part was removed. After 5 mL of phosphate-buffered saline (PBS) was added to the tubes and centrifuged at 4°C at 2000 rpm for 10 minutes, the PBS was removed. 250 μL RIPA buffer (Radio-Immunoprecipitation Assay), 2.5 μL PMSF (Phenylmethylsulfonyl fluoride, 200 mm), 2.5 μL sodium vanadate (100 mm) and 2.5 μL protease inhibitor were added to the cells. Homogenates were obtained from cells using an ultrasonic homogenizer on ice. Homogenates were centrifuged at 10000 rpm for 10 minutes. Supernatants were taken, the remaining sediments (pellets) were discarded.
Total Protein Determination
The total protein concentration in the homogenized tissues was determined using the Bradford method. Bovine bovine serum albumin (1–100 μg/μL) was used as the standard for total protein analysis. The protein content (μg/μL) was calculated based on a standard curve established in Prism software.
ELISA (Enzyme-Linked Immunosorbent Assay) Tests
The ELISA technique was employed to measure caspase-3 activity and the expression levels of proteins involved in apoptotic pathways, including bax, bcl-2, wee 1, gadd153, and AIF. Additionally, active ERK (p-ERK) and JNK (p-JNK) were analyzed via ELISA. Assessments of TAS, TOS, SOD, and GPx were carried out following previously described methodologies.
Statistical Analysis
Data are expressed as means ± S.E.M., with “n” denoting the number of cell culture flasks utilized for each group. Comparisons between the groups were made using analysis of variance (ANOVA) with Bonferroni corrections applied for multiple comparisons, and p-values less than 0.05 were deemed statistically significant.
Results
In this study, we evaluated the mediators of apoptosis, which are crucial components in the process of programmed cell death.12 The activity of caspase-3 and the expression of specific proteins were measured to understand their role in this process. Both caspase-3 and the protein bax are known to promote apoptosis, whereas bcl-2 functions as a protective agent against cell death.13 Our results indicated that treatment with Methotrexate (MTX) led to a significant increase in caspase-3 activity (Figure 1, P<0.0001) and elevated bax expression (Figure 2, P<0.0001). Furthermore, we observed a notable decrease in bcl-2 expression (Figure 3, P<0.0001), leading to a disrupted bax/bcl-2 ratio that predisposes cells to apoptosis. MTX treatment also resulted in diminished levels of additional apoptotic mediators such as wee 1, grp78, gadd153, and AIF (Table 1). Fortunately, we found that treatment with Tempol significantly counteracted these adverse effects.
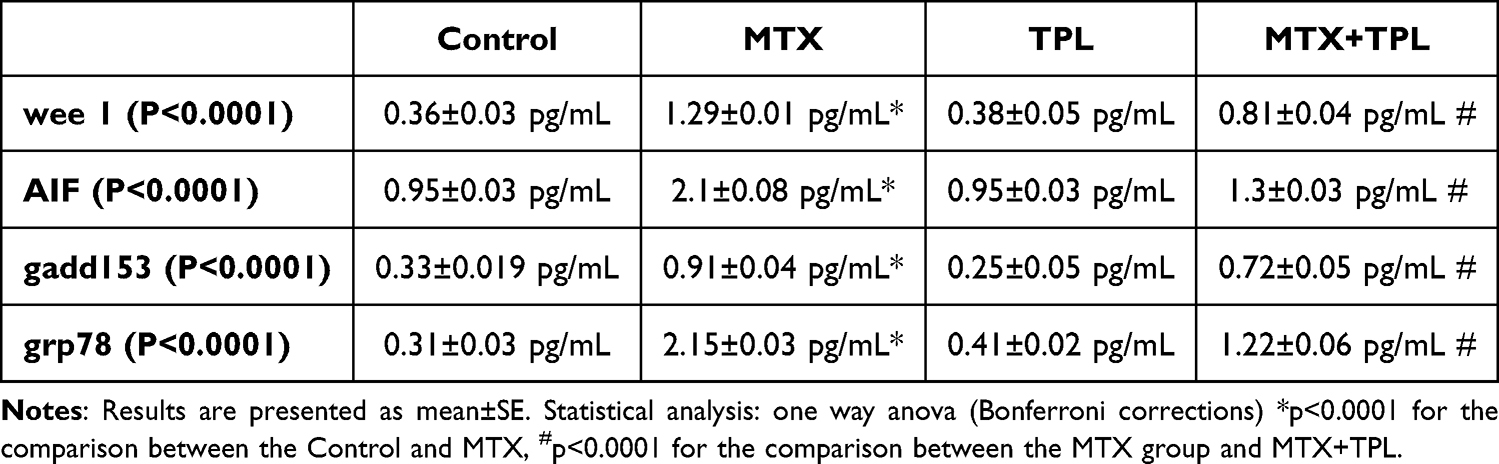 |
Table 1 Effect of TPL and MTX treatment on wee 1, gadd153, grp78 and AIF expression. |
 |
Figure 1 Effect of MTX and TPL on caspase-3 activity (n=8). Statistical analysis: ANOVA. Post hoc: Bonferroni. (*For control P < 0.05. # For I/R P < 0.05). |
 |
Figure 2 Effect of MTX and TPL on bax levels (n=8). Statistical analysis: ANOVA. Post hoc: Bonferroni. (*For control P < 0.05. # For I/R P < 0.05). |
 |
Figure 3 Effect of MTX and TPL on bcl-2 levels (n=8). Statistical analysis: ANOVA. Post hoc: Bonferroni. (*For control P < 0.05. # For I/R P < 0.05). |
The Mitogen-Activated Protein Kinase (MAPK) pathway components, specifically JNK and ERK, were also examined due to their pivotal role in mediating cellular signaling to the nucleus.13,14 Our findings showed that both JNK (p-JNK) and ERK (p-ERK) activities were significantly increased in the MTX group compared to the control group. Importantly, the application of Tempol effectively suppressed the MTX-induced activation of JNK (Figure 4, P<0.0001) and ERK (Figure 5, P<0.0001).
 |
Figure 4 Effect of MTX and TPL on JNK activity (n=8). Statistical analysis: ANOVA. Post hoc: Bonferroni. (*For control P < 0.05. # For I/R P < 0.05). |
 |
Figure 5 Effect of MTX and TPL on ERK activity (n=8). Statistical analysis: ANOVA. Post hoc: Bonferroni. (*For control P < 0.05. # For I/R P < 0.05). |
We assessed various oxidative stress parameters and found that the levels of Total Antioxidant Status (TAS) were significantly reduced in the MTX-treated group when contrasted with control subjects. In contrast, the TAS levels were markedly higher in the group treated with Tempol compared to the MTX group (Figure 6, p < 0.0001).
 |
Figure 6 Effect of MTX and TPL on Tas (n=8). Statistical analysis: ANOVA. Post hoc: Bonferroni. (*For control P < 0.05. # For I/R P < 0.05). |
Additionally, we observed that Total Oxidant Status (TOS) levels were notably elevated in the MTX group relative to controls. Conversely, TOS levels were significantly decreased in the Tempol-treated group compared to those receiving MTX (Figure 7, P < 0.0001).
 |
Figure 7 Effect of MTX and TPL on TOS (n=8). Statistical analysis: ANOVA. Post hoc: Bonferroni. (*For control P < 0.05. # For I/R P < 0.05). |
Furthermore, the activities of superoxide dismutase (SOD) (Figures 8, P < 0.0001) and glutathione peroxidase (GPx) (Figures 9, P < 0.0001) two vital endogenous scavengers of reactive oxygen species (ROS) were considerably lower in the MTX group compared to the control group. However, these enzyme levels were significantly elevated in the Tempol-treated group in relation to the MTX group.
 |
Figure 8 Effect of MTX and TPL on SOD (n=8). Statistical analysis: ANOVA. Post hoc: Bonferroni. (*For control P < 0.05. # For I/R P < 0.05). |
 |
Figure 9 Effect of MTX and TPL on Gpx (n=8). Statistical analysis: ANOVA. Post hoc: Bonferroni. (*For control P < 0.05. # For I/R P < 0.05). |
Discussion
Apoptotic Mediators
The study evaluated the effects of Methotrexate (MTX) treatment on key proteins and enzymatic activities associated with programmed cell death. The results indicate a significant increase in caspase-3 activity, suggesting that MTX effectively promotes apoptotic pathways in target cells. Additionally, the elevated expression of the pro-apoptotic protein bax further strengthens the pro-apoptotic signaling cascade initiated by MTX.1,9,15
The decrease in bcl-2 expression is particularly noteworthy, as bcl-2 functions to inhibit apoptosis.12,16 This reduction disrupts the delicate balance between pro-apoptotic (bax) and anti-apoptotic (bcl-2) factors, resulting in a bax/bcl-2 ratio that favors apoptosis an indication that MTX may lead to increased cell death in treated populations.
This study has demonstrated that Tempol (TPL) exhibits a protective effect at low doses, effectively mitigating oxidative stress and enhancing cellular viability. This protective effect is particularly evident in lung cancer cells and juxtaglomerular cells, where Tempol helps prevent cell death in response to cytotoxic insults. The underlying mechanisms of this protection are hypothesized to stem from Tempol’s potential antioxidant properties, particularly its function as a superoxide dismutase (SOD) mimic. By scavenging superoxide radicals (O2 •−) and reducing the intracellular concentration of ferrous iron (Fe II), Tempol facilitates the reduction of hydroxyl radical (•OH) formation, both of which are involved in generating harmful oxidative species. However, emerging evidence indicates that at higher concentrations, Tempol can induce apoptotic effects, acting as a nitroxide compound. This dual behavior underscores the complexity of Tempol’s action and reflects its dose-dependent influence on redox (oxidation-reduction) reactions within the cell. The toxicity of Tempol appears to be dose-dependent, with studies suggesting that it preferentially targets cancer cells while sparing normal cells under certain conditions. At millimolar concentrations, Tempol has been shown to inhibit cell growth and induce apoptosis, with the effects varying significantly based on the specific cell type and its origin. For instance, high concentrations of Tempol or prolonged exposure have been linked to increased levels of reactive oxygen species (ROS) in juxtaglomerular cells, breast cancer cells, and ovarian cancer cells. This paradoxical effect raises important questions about Tempol’s dual role as both an antioxidant and a potential pro-oxidant in different biological contexts. Collectively, this variation in response highlights the importance of dosage when considering Tempol for therapeutic applications. While the beneficial effects at lower concentrations are notable, they may shift toward cytotoxic effects at higher doses. Therefore, further research is needed to clarify the conditions under which Tempol operates as either a protective antioxidant or a harmful pro-oxidant, paving the way for its rational application in clinical settings.15,17,18
Furthermore, the diminished expression of other apoptotic mediators, such as wee 1, grp78, gadd153, and AIF, provides further insight into the multifaceted mechanisms through which MTX exerts its cytotoxic effects. Wee 1 is involved in cell cycle regulation and its reduction could lead to unchecked cell cycle progression, while gadd153 (also known as CHOP) is a stress-responsive protein linked to apoptosis.19 AIF is crucial for caspase-independent apoptosis, and its decline indicates that MTX may exploit multiple pathways to initiate cell death.20
Importantly, the ability of Tempol to counteract these effects suggests a promising protective mechanism. By restoring the balance of these apoptotic mediators, Tempol may help to preserve cell viability during MTX therapy, underscoring its potential as an adjunctive treatment.
MAPK Mediators
Numerous extracellular and intracellular stimuli have been demonstrated to activate MAPK (Mitogen-Activated Protein Kinase) pathways. One significant contributor to this activation is reactive oxygen species (ROS); however, the precise mechanisms through which ROS exert this effect remain unclear. ROS are believed to play crucial roles as modulators and signaling molecules within cells.
A substantial body of evidence supports the concept that ROS act as “second messengers” within intracellular signaling pathways that regulate essential cellular processes such as growth, proliferation, migration, and apoptosis. Their involvement in these pathways underscores the importance of ROS in mediating cellular responses to various stimuli and highlights their dual role as both functional signals and potentially damaging agents within the cell. Understanding the exact mechanisms by which ROS influence MAPK signaling could provide valuable insights into their physiological roles and implications for various biological contexts.
The results concerning MAPK signaling pathways illustrate a critical aspect of how MTX influences cellular responses.4,21 The significant increases in p-JNK and p-ERK activities in the MTX-treated group highlight the activation of stress-related signaling pathways. JNK is often associated with pro-apoptotic signals, while ERK can promote cell survival under certain conditions.22 The observed elevation in these markers suggests that MTX triggers pathways that can lead to cell death while also activating survival signals in a context-dependent manner.
The subsequent inhibition of JNK and ERK activation by TPL reinforces its role as a protective agent. By modulating these signaling cascades, TPL may reduce both oxidative stress and apoptosis, promoting a healthier cellular environment under MTX treatment. This dual-mode action could serve as a foundational strategy in managing side effects associated with cancer therapies.
Oxidative Stress Parameters
The assessment of oxidative stress parameters reveals critical insights into the cellular environment in response to MTX. The significant reduction in Total Antioxidant Status (TAS) and the increase in Total Oxidant Status (TOS) indicate that MTX exposure induces oxidative stress, which can lead to cellular damage and apoptosis. The imbalance between oxidants and antioxidants is a well-documented pathway for drug-induced toxicity.
Furthermore, the decrease in the activities of superoxide dismutase (SOD) and glutathione peroxidase (GPx) underscores the overwhelming oxidative stress faced by cells undergoing MTX treatment. These enzymes play crucial roles in detoxifying reactive oxygen species (ROS), and their decline signifies a diminished cellular defense against oxidative injury.12
Tempol’s effectiveness in restoring TAS and lowering TOS levels indicates its role as a potent antioxidant.9 The elevation of SOD and GPx activities in the Tempol-treated group further strengthens the argument for its protective role against oxidative stress induced by MTX. This suggests that Tempol may actively enhance the cellular antioxidant defense system, thereby preventing oxidative damage and apoptosis.
Conclusion
In conclusion, this study provides valuable insights into the mechanisms underlying MTX-induced toxicity and highlights the potential of Tempol as a protective agent. The observed alterations in apoptotic mediators, MAPK signaling, and oxidative stress parameters underscore the complex interplay between drug action and cellular response. These findings support the hypothesis that Tempol may serve as a beneficial adjunct to MTX therapy, protecting cellular integrity and potentially improving treatment outcomes. However, the broader clinical implications of these findings necessitate the translation of our in vitro results to in vivo and clinical studies to verify Tempol’s efficacy and safety in a real-world context. Emphasizing the importance of dose optimization for Tempol is crucial, as achieving the right balance is essential to leveraging its protective effects while minimizing potential cytotoxicity. Future studies should focus not only on the long-term impacts of Tempol on cell survival and health in the context of MTX therapy but also on determining the optimal dosing strategies to maximize therapeutic benefits. Addressing these aspects will significantly advance therapeutic strategies and enhance the clinical relevancy of our findings.
Acknowledgments
The authors thank all the staff at the Department Medicinal Pharmacology of Cukurova University, for their help in this study.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Uruc V, Salimov F, Kaplan HM. Protective effect of hypericum perforatum extract on methotrexate-induced osteotoxicity via reducing oxidative stress and MAPK activity. Int J Pharm. 2020;16(5):430–436. doi:10.3923/ijp.2020.430.436
2. Fan CM, Georgiou KR, King TJ, Xian CJ. Methotrexate toxicity in growing long bones of young rats: a model for studying cancer chemotherapy-induced bone growth defects in children. J Biomed Biotechnol. 2011. doi:10.1155/2011/903097
3. Iantomasi T, Romagnoli C, Palmini G, et al. Oxidative stress and inflammation in osteoporosis: molecular mechanisms involved and the relationship with microRNAs. Int J mol Sci. 2023;24(4):ARTN3772. doi:10.3390/ijms24043772
4. Mizuno N, Shiga S, Tanaka Y, et al. CDK8/19 inhibitor enhances arginase-1 expression in macrophages via STAT6 and p38 MAPK activation. Eur J Pharmacol. 2024;979:176852. doi:10.1016/j.ejphar.2024.176852
5. Prasadam I, Zhou Y, Du Z, et al. Osteocyte-induced angiogenesis via VEGF–MAPK-dependent pathways in endothelial cells. Mol Cell Biochem. 2014;386(1–2):15–25. doi:10.1007/s11010-013-1840-2
6. Noguchi S, Yamasaki R, Nagai-Yoshioka Y, et al. The mechanism of interleukin 33-induced stimulation of interleukin 6 in MLO-Y4 cells. Int J mol Sci. 2023;24(19):14842. doi:10.3390/ijms241914842
7. Park WH. tempol inhibits the growth of lung cancer and normal cells through apoptosis accompanied by increased O2•− levels and glutathione depletion. Molecules. 2022;27(21):7341. doi:10.3390/molecules27217341
8. Wang M, Li K, Zou Z, et al. Piperidine nitroxide Tempol enhances cisplatin-induced apoptosis in ovarian cancer cells. Oncol Lett. 2018;16(4):4847–4854. doi:10.3892/ol.2018.9289
9. Pinar N, Kaplan M, Özgür T, et al. Ameliorating effects of tempol on methotrexate-induced liver injury in rats. Biomed Pharmacother. 2018;102:758–764. doi:10.1016/j.biopha.2018.03.147
10. Erker L, Schubert R, Yakushiji H, et al. Cancer chemoprevention by the antioxidant tempol acts partially via the p53 tumor suppressor. Hum Mol Genet. 2005;14(12):1699–1708. doi:10.1093/hmg/ddi181
11. Pinar N, Soylu karapınar O, Özcan O, et al. Protective effects of tempol in an experimental ovarian ischemia-reperfusion injury model in female Wistar albino rats. Can J Physiol Pharmacol. 2017;95(7):861–865. doi:10.1139/cjpp-2016-0309
12. Izol V, Arıdoğan IA, Tansuğ Z, et al. Hypericum perforatum extract against oxidative stress, apoptosis and oedema in kidney induced by Gentamicin. Int J Pharm. 2019;15(1):66–73. doi:10.3923/ijp.2019.66.73
13. Okyay AG, Kaplan HM, Asil H, Singirik E. Saffron induces apoptosis in ovarian cancer cell via MAPK and AKT/mTOR pathways. Prog Nutr. 2020;22. doi:10.23751/pn.v22i2-S.10160
14. Celik E, Kaplan HM, Singirik E. The impact of propranolol on apoptosis in cutaneous squamous cell carcinomas. Bratisl Med J. 2020;121(11):801–804. doi:10.4149/Bll_2020_131
15. Kaplan HM, Pazarci P. Antiproliferative and apoptotic effects of tempol, methotrexate, and their combinations on the MCF7 breast cancer cell line. Acs Omega. 2024;9(6):6658–6662. doi:10.1021/acsomega.3c07624
16. Shandala T, Shen Ng Y, Hopwood B, et al. The role of osteocyte apoptosis in cancer chemotherapy-induced bone loss. J Cell Physiol. 2012;227(7):2889–2897. doi:10.1002/jcp.23034
17. Santos FR, Rossetto IMU, Montico F, et al. Differential tempol effects in prostatic cancer: angiogenesis and short- and long-term treatments. J Mol Histol. 2024;55(3):253–264. doi:10.1007/s10735-024-10187-4
18. Du JA. Tempol inhibits pancreatic cancer cell growth. Free Radic Biol Med. 2010;49:S58–S58. doi:10.1016/j.freeradbiomed.2010.10.133
19. Siyo V, Schäfer G, Hunter R, et al. The cytotoxicity of the ajoene analogue BisPMB in WHCO1 oesophageal cancer cells is mediated by CHOP/GADD153. Molecules. 2017;22(6):892. doi:10.3390/molecules22060892
20. Li X, Zhao Y, Jiang G. Aldosterone promotes calcification of vascular smooth muscle cells in mice through the AIF-1/Wnt/beta-catenin signaling pathway. Int Urol Nephrol. 2024;57(2):613–623. doi:10.1007/s11255-024-04213-3
21. Jin X, Zhu H, Chen X, et al. RON receptor tyrosine kinase regulates glycolysis through MAPK/CREB signaling to affect ferroptosis and chemotherapy sensitivity of thyroid cancer cells. Mol Med Rep. 2024;30(6). doi:10.3892/mmr.2024.13359
22. Ding W-J, Huang S-L, Huang S, et al. Di(2-ethylhexyl) phthalate mediates oxidative stress and activates p38MAPK/NF-kB to exacerbate diabetes-induced kidney injury in vitro and in vivo models. Toxicol Res. 2023;12(2):332–343. doi:10.1093/toxres/tfad022
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms.php
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms.php
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.