")
Back to Journals » Diabetes, Metabolic Syndrome and Obesity » Volume 17
Type 2 Diabetes Mellitus Aggravates Complement Dysregulation and Affects Cortisol Response in Patients with Post-COVID-19
Authors Ji W, Xie X , Bai G, Fan Y, He Y, Zhang L, Zhou H, Li L, Qiang D, Li H
Received 29 May 2024
Accepted for publication 1 October 2024
Published 19 October 2024 Volume 2024:17 Pages 3849—3861
DOI https://doi.org/10.2147/DMSO.S480457
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Prof. Dr. Muthuswamy Balasubramanyam
Wenrui Ji,1,* Xiaomin Xie,1,* Guirong Bai,1 Yalei Fan,2 Yanting He,1 Li Zhang,1 Haiyan Zhou,1 Ling Li,1 Dan Qiang,1 Huan Li1
1Department of Endocrinology, the First People’s Hospital of Yinchuan, Yinchuan, 750001, People’s Republic of China; 2The Second Clinical Medical School of Ningxia Medical University, Yinchuan, 750001, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Xiaomin Xie, Department of Endocrinology, the First People’s Hospital of Yinchuan, Liqun West Street No. 2, Xingqing District, Yinchuan, Ningxia Hui Autonomous Region, 750001, People’s Republic of China, Tel +8613895189599, Email [email protected]
Purpose: COVID-19 viral infection results in dysregulation of the complement system and a decrease in cortisol and adrenocorticotropin hormone (ACTH) levels. This study aimed to explore the complement system, as well as cortisol and ACTH responses in patients with post-COVID-19 conditions (PCC) and type 2 diabetes mellitus (T2DM).
Patients and Methods: This study recruited 31 patients with PCC and T2DM (PCC-T2DM), 19 patients with PCC (PCC), 10 patients with T2DM (T2DM), and 10 healthy participants (control). Cortisol and ACTH in the PCC and PCC-T2DM groups were assessed using the insulin tolerance test. In the fasting state, serum samples were collected for proteomic analyses. Spearman correlation analysis was performed between proteins and cortisol, as well as between proteins and ACTH.
Results: Cortisol and ACTH levels were consistently decreased in the PCC and PCC-T2DM groups. Proteomic analyses revealed that most of the differentially abundant proteins (DAPs) in the PCC vs control and PCC-T2DM vs T2DM were involved in the coagulation and complement cascade, and the essential complement C3 was significantly upregulated in the PCC and PCC-T2DM groups when compared to their controls. Additionally, complement-related DAPs in the PCC vs control and PCC-T2DM vs T2DM were significantly correlated with cortisol and ACTH levels. In comparing PCC-T2DM samples with PCC samples, we found that upregulated DAPs were linked to the complement system and other immune system, and most DAPs were negatively correlated with cortisol and ACTH.
Conclusion: Our study revealed that T2DM exacerbated dysregulation of the complement system in patients with PCC, and significant correlations were present between complement protein levels and those of cortisol and ACTH. These results provide novel insights into the dysregulation of complement and endocrine hormones in patients with PCC and T2DM.
Keywords: post-COVID-19 condition, type 2 diabetes mellitus, complement, cortisol, adrenocorticotropin hormone
Graphical Abstract:

Introduction
Long-term effects of COVID-19 have been recently observed a distressing sequelae and are recognized by the World Health Organization as post-COVID-19 condition (PCC).1 Patients with PCC have a confirmed SARS-CoV-2 infection, typically presenting 3 months of post-COVID-19 onset, with symptoms lasting for at least 2 months that cannot be explained by other factors.2 A number of cognitive and autonomic dysfunctions are commonly reported by individuals with PCC, including fatigue, post-exertional malaise, and pain after physical exertion.3
Diabetes mellitus (DM, mostly type 2) and COVID-19 show mutual interactions.4 DM is a well-known risk factor for worse clinical outcomes in patients with COVID-19.5 Compared with non-diabetics, diabetics infected with SARS-CoV-2 have a higher rate of hospitalization, severe pneumonia, and mortality.6 Conversely, COVID-19 alters energy metabolism in DM patients. When SARS-CoV-2 attaches to the pancreatic angiotensin-converting enzyme 2 (ACE2) system, insulin production is perturbed, resulting in hyperglycemia.7 Hyperglycemia further induces inflammation, promotes insulin resistance, and reduces insulin secretion in patients with DM.8 These studies indicate that the alterations in metabolic pathways may be a hub of interaction between PCC and DM. However, the differential proteins involved in metabolic alterations in patients with PCC and DM have not yet been investigated.
Studies have found immune dysregulation in individuals with PCC,3,9,10 such as complement immunity.11 Comparing the blood of patients with confirmed SARS-CoV-2 infection with that of uninfected controls, Cervia-Hasler et al found that patients experiencing long COVID exhibit dysregulated activation of complement system, increased markers of hemolysis and platelet activation, and altered coagulation cascade.12 Ma et al also suggested that enhanced activation of the alternative complement pathway is most prevalent in patients with severe COVID-19.13 An increasing evidence also points toward a role of the complement system in the pathogenesis of DM, and complement upregulation and activation is an important feature of insulin resistance and the development of type 2 diabetes mellitus (T2DM).14–16 Nonetheless, the complement system in patients with PCC and T2DM is unknown.
One of the primary immune invasive strategy employed by SARS-CoV is to knock down the host’s cortisol stress response.17 The gene encoding ACE2 is the host receptor that binds SARS-CoV-2.18 Several endocrine organs have been identified to be targeted by SARS-CoV-2,19 and the virus enters the target cells in the pituitary and adrenal glands via the ACE2 receptor.20 Recent reports indicated that cortisol level is lower among participants with long COVID.3,9,21 In addition, Ahmadi et al reported that low cortisol level is linked to a high risk of mortality in patients with COVID-19.22 Cortisol is widely understood to suppress inflammation and immunity.23 However, the underlying correlation between cortisol response and complement in patients with PCC and T2DM remains unclear.
The purpose of this study was to analyze the complement function and cortisol response in PCC patients, T2DM patients, and patients with PCC and T2DM. Ultra-performance liquid chromatography (UPLC) and tandem mass spectrometry (MS/MS) were used for proteomic analysis to reveal changes in pivotal complement proteins in subjects. This study provides novel insights into the complement system and cortisol responses in patients with PCC and T2DM, and identifies potential therapeutic targets for this patient population.
Methods and Materials
Subjects
Fifty patients with PCC who visited the Endocrinology Department of the Second Affiliated Hospital of Ningxia Medical University from June, 2023 to December, 2023 were recruited, including 28 females and 22 males. The participants were 18–75 years old and divided into PCC patients (PCC group) and patients with PCC and T2DM (PCC-T2DM group). The PCC group contained 19 PCC patients with an average age of 44.21±15.5 years old. Group PCC-T2DM included 31 patients with PCC and T2DM and an average age of 55.1±15.5 years old. Patients with PCC and T2DM followed the diagnostic criteria of the American Diabetes Association and PCC,2,24 respectively. Information on all individual participants is shown in Table S1. Ten healthy participants with non-SARS-CoV-2 infection and non-DM (control group) and 10 patients with non-SARS-CoV-2 infection and T2DM (T2DM group) who visited the Physical Examination Center of the Second Affiliated Hospital of Ningxia Medical University from September, 2021 to November, 2021 were recruited, including 10 females and 10 males.
The exclusion criteria were as follows:25 (1) type 1 DM or other specific types of DM; (2) patients with acute and chronic complications of DM; (3) chronic kidney and liver disease or cancer; (4) secondary hypertension; (5) history of anemia, severe cardiac and cerebrovascular diseases, or tumors; and (6) pregnant or breastfeeding women. This study was conducted in accordance with the Declaration of Helsinki and approved by the Ethics Committee of Yinchuan First People’s Hospital (approval number KY-2024-034). Informed consent was obtained from all the patients.
Blood Tests
After an overnight fasting period of 8–12 hours, the patients were bedridden to collect peripheral venous blood. The Beckman Coulter Automated Chemistry Analyzer AU5821 was used to detect fasting blood glucose (FBG), alanine aminotransferase (ALT), aspartate aminotransferase (AST), total protein (TP), albumin (ALB), triglycerides (TG), total cholesterol (TC), high-density lipoprotein (HDL), low-density lipoprotein (LDL), creatinine (Cr), and uric acid (UA). Plasma cortisol and adrenocorticotropic hormone (ACTH) levels were measured using electrochemiluminescence (Cobas 6000 Automated Chemistry Immunological Analysis System). Partial blood samples were prepared for proteomic sequencing, and residual samples were stored at −80°C.
Insulin Tolerance Test (ITT)
According to a final dose of 0.1–0.15 U/kg of insulin for intravenous injection, insulin was accurately calculated and dissolved in the volume of the 2 mL glucose stock solution. Peripheral blood glucose, venous blood glucose, cortisol, and ACTH in patients with a fasting period of 8 hours were detected at 0 min, 30 min, 60 min, and 90 min after insulin injection. If there was no hypoglycemic reaction or blood glucose level did not drop below 2.2 mmol/L, the test was repeated (insulin increased to 0.3 IU/kg). Symptoms of hypoglycemia (generally appeared after 15–30 min of insulin injection) or blood glucose changes at 0, 15, 30, 45, and 60 min before and after insulin injection were observed. A blood glucose level falling to 50% of the baseline or < 2.6 mmol/L was considered a valid ITT test. All the blood samples were stored in an ice bath.
Sample Preparation
Blood samples were centrifuged to obtain serum. Serum samples were subsequently treated with lysate, protease inhibitor (ROCHE, 04693132001) was added at 1:50 (protease inhibitors: lysate), and 8 M of urea (Amresco, M123-1KG) was added. The samples were ultrasonically broken for 1 sec at 2 sec intervals, and the cumulative duration was 120 s. The samples were then separated by centrifugation at 14,000×g for 20 min at 4°C to obtain the protein samples. These proteins were then analyzed using UPLC-MS/MS.
UPLC-MS/MS Analysis
Mobile phase solutions A (100% MS-grade water with 0.1% formic acid) and B (80% acetonitrile with 0.1% formic acid) were prepared. Lyophilized proteins were dissolved in 10 μL of solution A and centrifuged at 14000×g at 4°C for 20 min. One microgram of supernatant was used for liquid quality detection using a RIGOL L-3000 high-performance liquid chromatography system. Q Exactive HF-X Mass Spectrometer and the Nanospray Flex ion source were used in this study.
Bioinformatics Analysis
Principal component analysis (PCA) was performed to visualize the separation of samples between groups using scales, ggplot2, ggrepel, and Scatterplot3d R packages. The Blast2GO software was used to identify enriched Gene Ontology (GO) terms in the biological process, molecular function, or cellular component categories. Kyoto Encyclopedia of Genes and Genomes (KEGG) was used to KEGG pathway analysis of proteins. Fisher’s exact test was used to calculate enrichment p values according to the method of Benjamini and Hochberg for multiple testing corrections. The Reactome database was used to explore the functional pathways of proteins. Statistical significance was considered when Benjamini- and Hochberg adjusted p values were < 0.05.
Differential Analysis of Proteins
Differentially abundant proteins (DAPs) between groups were determined by t-test and fold change (FC) using the ggplot2 software package. DAPs with FC ≥ 1.30 or 1.20 and p value ≤ 0.05 were considered to be upregulated DAPs. DAPs with FC ≤ 1/1.30 or 1/1.20 and p value ≤ 0.05 were considered downregulated DAPs.
Statistical Analysis
SPSS 27.0 (IBM, USA) was used to statistically analyze the data, and the results are presented as mean ± standard deviation. The t-test was used for comparisons between the two groups. Pearson’s correlation analysis was used to evaluate correlations. P < 0.05 was considered statistically significant difference.
Results
Cortisol and ACTH Responses are Impaired in PCC Patients, as Well as Patients with PCC and T2DM
The metabolic parameters of the PCC and PCC-T2DM groups are shown in Table S2. Patients with PCC and T2DM had significantly higher fasting blood glucose (FBG) and glycosylated hemoglobin (HbA1c) levels than PCC patients (Table S2). No significant differences were found in other metabolic parameters between diabetic and non-diabetic patients with PCC (Table S2). ITT is a gold standard for evaluating the function of the hypothalamus-pituitary-adrenal (HPA) axis.26 It stimulates the release of cortisol and ACTH by inducing hypoglycemia. Thus, ITT was used to assessed the cortisol response in patients with PCC. Fasting plasma cortisol and ACTH levels were considered as baseline cortisol and ACTH. The cortisol and ACTH values are shown in Table 1. We found that ΔACTH0 in the PCC-T2DM group was significantly lower than that in the PCC group. Importantly, both cortisol and ACTH levels showed negative growth after 30 min of ITT, implying impaired cortisol and ACTH responses in patients with PCC.
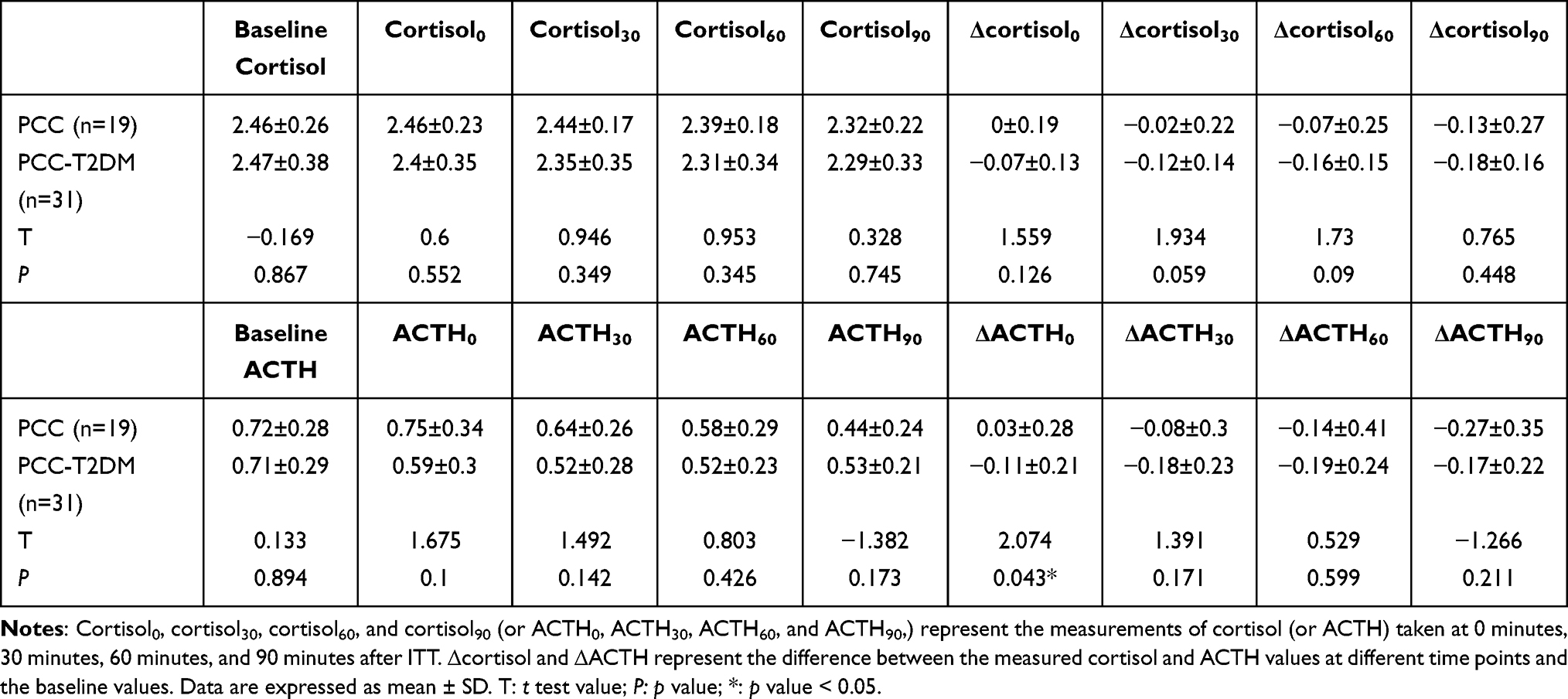 |
Table 1 Values of Cortisol and ACTH in the PCC and PCC-T2DM Groups |
PCC Induces Dysregulated Coagulation and Complement Cascade in Healthy and T2DM subjects
Serum samples from PCC, PCC-T2DM, control (subjects with no SARS-CoV-2 infection and T2DM), and T2DM groups were analyzed using a UPLC-MS/MS-based assay to quantitatively determine a panel of proteins with potential roles in patients with PCC and T2DM. Unsupervised analysis using PCA showed a distinction among the groups: the control and T2DM samples were clearly separated from the PCC and PCC-T2DM samples (Figure S1a). The precision of our procedures was measured as RSD (Figure S1b). Subsequently, we compared DAPs between PCC patients and controls as well as between PCC-T2DM and T2DM patients. A total of 24 DAPs were found to be upregulated, while 154 DAPs were downregulated in PCC patients compared with controls (Figure 1a). Similarly, 23 upregulated and 154 downregulated DAPs were observed in the PCC-T2DM vs T2DM (Figure 1b). Notably, GO terms of DAPs in both PCC vs control and PCC-T2DM vs T2DM were enriched in complement and coagulation cascades, such as complement activation, lectin pathway, and positive regulation of blood coagulation (Figure 1c and d). These results indicate that PCC induces dysregulation of the coagulation and complement cascade in both healthy subjects and T2DM patients.
 |
Figure 1 Dysregulation of complement and coagulation cascade in PCC patients and patients with PCC and T2DM. (a and b) Volcano plots indicate the DAPs in PCC vs control (a) and PCC-T2DM vs T2DM (b). The orange, green, and black dots represent upregulated, downregulated, and normal DAPs, respectively. (c and d) GO enrichment analysis of DAPs in the PCC vs control (c) and PCC-T2DM vs T2DM (d). |
Next, we depicted a network of shared 42 DAPs identified in both PCC vs control and PCC-T2DM vs PCC, which were involved in the coagulation and complement cascade pathway in the KEGG database (Figure 2). Prothrombin (F2) is known to contribute significantly to the prothrombinase-catalyzed activation of prothrombin,27 and can regulate C3 and C5 to activate complement pathway.28 C3 and C5 are pivotal components of the complement cascade. In the PCC vs control and PCC-T2DM vs T2DM, the upregulation of F2 and C3 was observed in PCC and PCC-T2DM patients (Figure 2). Complement factor H (CFH or FH) can accelerate the decay of the complement alternative pathway C3 convertase C3bBb.29,30 The upregulation of FH in this study may further promote the decay of C3 convertase C3bBb (Figure 2). Additionally, C1qr2 in classical pathway and mannose binding lectin (MBL) and MBL associated serine protease 1/2 (MASP1/2) in lectin pathway exhibited significant downregulation (Figure 2), which further inhibited the levels of C2 and C4, indicating the block of classical pathway and lectin pathway. Interestingly, complement pathways were found to be connected to the downstream B cell receptor signaling pathway (Figure 2). DAPs associated with the B cell response, such as immunoglobulin heavy variable 3–7 (IGHV3-7) and immunoglobulin heavy constant alpha 1 (IGHA1), exhibited significant downregulation in the PCC-T2DM group when compared to the T2DM group (Figure 1a and b). These findings further validate the dysregulation of complement system in PCC and PCC-T2DM subjects.
 |
Figure 2 KEGG map of coagulation and complement cascades. KEGG map of coagulation and complement cascades (ko04610) depicts the relationships among 42 DAPs between PCC and control groups, as well as T2DM and PCC-T2DM groups. The red and green boxes represent upregulated and downregulated DAPs in the PCC and PCC-T2DM groups compared to the control and T2DM groups, respectively. |
Complement Proteins are Significantly Correlated with Cortisol and ACTH
Subsequently, several complement-related DAPs were selected for further excavation, as listed in Table 2. Interestingly, classical and lectin pathway activation markers, such as complement C1r subcomponent-like protein (C1S) and complement C1q subcomponent subunit C (C1QC), were downregulated in PCC and PCC-T2DM patients compared to control and T2DM patients, respectively (Table 2). In contrast, complement C3 (C3) was found an upregulation (Table 2). To better understand the association of complement-related DAPs with cortisol and ACTH values, we performed Spearman correlation analysis. The findings demonstrated positive and negative correlations between MASP2 and C6 and Δcortisol30, Δcortisol60, and Δcortisol90, respectively (Figure 3a). C1S, C2, and C3 were negatively correlated with cortisol60 and cortisol90 (Figure 3a). Intriguingly, complement-related DAPs such as CFP, MASP2, and FCN3 were positively correlated with ACTH0, ACTH30, ACTH60, and ACTH90, and C3 and C6 were negatively correlated with most ACTH and its growth (Figure 3b). These results demonstrate a potential correlation between the complement system and the production of cortisol and ACTH.
 |
Table 2 Complement-Related DAPs |
 |
Figure 3 Correlation analysis between complement proteins and levels of cortisol ACTH. (a and b) Correlations between complement-related DAPs and cortisol (a) and ACTH (b) levels. The “×” in each box indicates that the p value is > 0.05, with color intensity indicating the Spearman correlation intensity. P < 0.05 is considered statistically significant correlations between two factors. |
T2DM May Exacerbate the Complement System Dysregulation in Patients with PCC
We learned that PCC results in dysregulation of the coagulation and complement cascade in healthy and T2DM subjects. To further explore the vital DAPs between PCC and PCC-T2DM patients, we extracted their UPLC-MS/MS data for differential analysis. A total of 10 DAPs were acquired, including targets of Nesh-SH3 (ABI3BP), apolipoprotein M (APOM), periostin (POSTN), neural cell adhesion molecule L1-like protein (CHL1), neuropilin-1 (NRP1), ACE, fructose-bisphosphate aldolase B (ALDOB), collectin-11 (COLEC11), IgGFc-binding protein (FCGBP), and immunoglobulin kappa variable 1D-16 (IGKV1D-16), which were upregulated in PCC-T2DM patients compared with those in patients with PCC (Figure 4a). These DAPs comprise the domains of fasciclin, lectin C-type, immunoglobulin, immunoglobulin V-set, and angiotensin-converting enzyme (Figure 4b). In addition, DAPs were enriched in the renin-angiotensin system and renin secretion pathways (Figure S2a), and the KEGG map shows that upregulated ACE is involved in the regulation of complement and coagulation cascades via the renin-angiotensin system (Figure S2b). In addition, DAPs were not only associated with complement-related Reactome enrichment pathways but also with other immune-related pathways like antigen activates B cell Receptor leading to generation of second messengers, adaptive immune system, and innate immune system (Figure 4c). Upregulated DAPs were also found to be associated with nervous system development (Figure 4c), suggesting an effect of T2DM on the nervous system in patients with PCC. Collectively, T2DM may exacerbate the complement system dysregulation in patients with PCC.
 |
Figure 4 Enrichment analysis of DAPs between the PCC and PCC-T2DM groups. (a) Volcano plot showing fold change (log2 values) and probability (log10 values) for individual DAPs between the PCC-T2DM and PCC groups. Upregulated and normal DAPs are highlighted by red and black dots, respectively. (b and c) Bubble diagrams showing the enriched protein domains (b) and Reactome pathways (c) of DAPs between the PCC-T2DM and PCC subjects. |
Most of DAPs Between PCC and PCC-T2DM Groups Show Negative Correlations with Levels of Cortisol and ACTH
To gain a deeper understanding of the relationship between the 10 DAPs in Figure 4a and cortisol and ACTH, Spearman correlation analysis was performed. We observed that APOM expression positively correlated with values of cortisol0, cortisol30, and Δcortisol0 (Figure 5a). DAPs, including ABI3BP, NRP1, and ALDOB, were significantly negatively correlated with Δcortisol30, Δcortisol60, and Δcortisol90 (Figure 5b). In addition, NRP1 levels were negatively correlated with both cortisol and ACTH levels (Figure 5a and b). Interestingly, COLEC11 was the only DAP with a significant positive correlation with ACTH levels at different time points (Figure 5b). Together, most of the DAPs between the PCC and PCC-T2DM groups showe negative correlations with cortisol and ACTH levels.
 |
Figure 5 Correlation analysis between DAPs in the PCC-T2DM vs PCC and levels of cortisol ACTH. (a and b) Heatmap visualizing Spearman correlations between the expression of DAPs in the PCC-T2DM vs PCC and levels of cortisol (a) and ACTH (b). |
Discussion
Complement dysregulation has been observed in patients with PCC and patients with T2DM and PCC. PCC is characterized by dysregulation of the terminal complement system and ongoing activation of the alternative and classical complement pathways.12 Previous study suggested that patients undergoing COVID-19 infection have elevated levels of alternative pathway activation markers, C3bBbP, and C5a.31 Additionally, antibodies against SARS-CoV-2 antigens can activate the classical complement pathway.32 In our results, DAPs involved in the classical and lectin complement pathways were downregulated in patients with PCC and those with both PCC and T2DM. The classical complement pathway is known as “antibody dependent” because IgM/IgG antibodies are involved in complement activation. C1 is shown to comprise three subcomponent proteins: C1q, C1r, and C1s, in which C1q binds to the Fc domain that leads to the activation of C1r and C1s.33,34 Then, C1 cleaves the next components of C4 and C2 to produce C4b2a complex, also known as the C3 convertase.33 In the lectin pathway, hexamers of MBLs bind to bacterial carbohydrate motifs. MASPs are complexed with large pattern recognition molecules such as MBL, collectins, and ficolins.35 MASPs also function similarly to C1r and C1s to cleave C4 and then C2, generating the C4bC2b C3 convertase.36 In this study, classical pathway markers such as C1S, C1QC, and C1RL and lectin pathway markers such as MASP2 and FCN3 exhibited downregulation in patients with PCC and patients with PCC and T2DM, as compared to both their controls, suggesting suppressed classical complement and lectin pathways. In contrast, complement C3 was upregulated in PCC and PCC-T2DM subjects. The central effector molecule of the complement system, C3, mediates multiple complement functions through different binding sites and receptors.37 C3 dysregulation implies the blocked complement pathways. Skendros et al reported that complement C3 activation contributes to COVID-19 pathology, and C3 targeting has emerged as a promising therapeutic strategy.38 Nonetheless, other studies have shown that low complement C3 levels are associated with poor prognosis in COVID-19 patients and a higher risk for clinical worsening among patients with COVID-19.39,40 The role of complement in COVID-19 is still limited by controversial results.
Complement regulation of lymphoid cell function has evolved greatly since lymphocytes synthesize the most soluble complement components. Effectors of the complement system are involved in the regulation of B lymphocyte function. Specifically, the products of complement system activation, particularly C3-derived fragments such as C3b, C3a, iC3b, and C3d, play a pivotal role in regulating B cell responses by binding to their corresponding receptors, including C3aR, CR1, CR2, CR3, and CR4.41–43 In this study, IGHV3-7 and IGHA1, which are immunoglobulins produced by B lymphocytes, were significantly downregulated in patients with PCC and T2DM compared to those in T2DM patients. In addition, compared to PCC patients, FCGBP and immunoglobulin IGKV1D-16 exhibited a significant upregulation in patients with PCC and T2DM. Regulatory trends of these immunoglobulins were similar to those observed in complement proteins between samples, suggesting an association between complement dysregulation and impaired B lymphocyte function in patients with PCC and T2DM.
We used ITT to evaluate the stress function of cortisol and ACTH in PCC patients and patients with PCC and T2DM, and found that cortisol and ACTH levels did not rise normally following ITT. HPA axis dysregulation is characterized by impaired ACTH secretion due to a disease or injury to the hypothalamus or the pituitary gland, leading to reduced cortisol production.44 HPA is a center of homeostasis, stress responses, energy metabolism, and neuropsychiatric function.45 Mounting evidence of disruption of the HPA axis in COVID-19 is known. For example, SARS-CoV can induce the injury of the adrenal glands.46,47 Patients who had COVID-19 underwent impaired pituitary functions.48 A relative hypo-responsiveness of the HPA axis with persistently low cortisol levels appears in PCC participants.49 Our findings were consistent with current studies that cortisol and ACTH levels in PCC participants and participants with PCC and T2DM consistently decreased following ITT and were lower than the baseline. We speculate that the impaired HPA axis may be one of the reasons for the abnormal cortisol and ACTH production.
Cortisol and ACTH are closely related to hemostasis.50–53 Hypercoagulability has been identified as a common complication in patients with Cushing’s syndrome.54 In this study, enrichment of coagulation pathways has been observed in PCC patients and patients with PCC and T2DM. Several studies have reported abnormal coagulation parameters, imbalance of thrombotic and thrombolytic factors, and endothelial cell injury in patients with COVID-19, and severe COVID-19 infection is associated with significant coagulopathy that correlates with disease severity.55–58 We noted that DAPs involved in the coagulation cascade were mostly downregulated in patients with PCC and patients with PCC and T2DM compared to their controls, implying that COVID-19 infection or abnormal regulation of cortisol responses may both lead to coagulation abnormalities.
Several complement proteins exhibited significant positive and negative correlations with cortisol and ACTH as well as the increase in cortisol and ACTH. HPA axis-mediated communication between the immune and endocrine system is important for the control of inflammation.59,60 Interestingly, Francis et al have reported the existence of C3a receptors in the anterior pituitary gland and the presence of C5a receptors in the gland.60,61 Our results found a significant upregulation of C3 in patients with PCC and patients with PCC and T2DM compared to their controls, and C3 exhibited a negative correlation with both cortisol and ACTH levels. The relationship between the complement system and the cortisol response warrants further investigation.
DM has been recognized as a significant risk factor for the rapid progression and poor prognosis of COVID-19.58 After SARS-CoV-2 infection, individuals with DM exhibit significantly elevated serum levels of IL-6, C-reactive protein, ferritin, and D-dimer compared to those without DM, which obliges the cytokine storm and contributes to the rapid deterioration of patients.62,63 Hypercoagulability and hypo-fibrinolysis are common features of both DM and COVID-19 because of the variations in hemostatic factors and overactivation of the endothelium.64,65 Similarly, complement dysregulation occurred in both PCC patients and patients with PCC and T2DM. When DM and COVID-19 overlapped, dysregulation of procoagulant factors and deficiency of fibrinolytic mechanisms were exacerbated in patients with PCC and T2DM. Additionally, these upregulated DAPs also participate in the renin-angiotensin system and renin secretion, and ACE is linked to the complement pathways via the renin-angiotensin system. The renin-angiotensin system responds to blood pressure. Under pathophysiological conditions, the effects of the renin-angiotensin system can intensify to trigger inflammation and structural remodeling, thus promoting cardiac and vascular damage.66 Renin-angiotensin system exerts a pivotal role in the pathogenesis of COVID-19.67 DM may respond to COVID-19 by regulating the cardiovascular system through the renin-angiotensin system combined with the complement system, exacerbating host pathological progression. This may be an important cause of cardiovascular complications in PCC patients.
Conclusion
Our study not only uncovered dysregulation in the complement and coagulation cascades and abnormal cortisol and ACTH responses in patients with PCC, but also revealed significant correlations between complement proteins and the levels of cortisol and ACTH. Importantly, T2DM further promotes complement disorders in patients with PCC. These results provide novel insights into the dysregulation of complement and endocrine hormones in PCC patients.
Data Sharing Statement
The datasets generated and/or analyzed during the current study are available from the corresponding author upon reasonable request.
Ethics Approval and Informed Consent
This study was approved by the Ethics Committee of The First People’s Hospital of Yinchuan (approval number: KY-2024-034). Informed consent was obtained from all the participants.
Consent for Publication
The authors declare that the details of this study can be published and that the person (s) providing consent have been shown in the article content to be published.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This project was supported by the 2024 Key Research and Development Project of the Autonomous Region (grant number: 2024BEG02015).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Sukocheva OA, Maksoud R, Beeraka NM. et al. Analysis of post COVID-19 condition and its overlap with myalgic encephalomyelitis/chronic fatigue syndrome. J Adv Res. 2022;40:179–196. doi:10.1016/j.jare.2021.11.013
2. Soriano JB, Murthy S, Marshall JC, et al. A clinical case definition of post-COVID-19 condition by a Delphi consensus. Lancet Infect Dis. 2022;22(4):e102–e07. doi:10.1016/S1473-3099(21)00703-9
3. Klein J, Wood J, Jaycox JR, et al. Distinguishing features of long COVID identified through immune profiling. Nature. 2023;623(7985):139–148. doi:10.1038/s41586-023-06651-y
4. Conte C, Cipponeri E, Roden M. Diabetes mellitus, energy metabolism, and COVID-19. Endocr Rev. 2024;45(2):281–308 doi:10.1210/endrev/bnad032.
5. Nassar M, Daoud A, Nso N, et al. Diabetes mellitus and COVID-19: review Article. Diabetes Metab Syndr. 2021;15(6):102268. doi:10.1016/j.dsx.2021.102268
6. Lima-Martinez MM, Carrera Boada C, Madera-Silva MD, Marin W, Contreras M. COVID-19 and diabetes: a bidirectional relationship. Clin Investig Arterioscler. 2021;33(3):151–157. doi:10.1016/j.arteri.2020.10.001
7. Somasundaram NP, Ranathunga I, Ratnasamy V, et al. The impact of SARS-Cov-2 virus infection on the endocrine system. J Endocr Soc. 2020;4(8):bvaa082. doi:10.1210/jendso/bvaa082
8. Ceriello A. Hyperglycemia and COVID-19: what was known and what is really new? Diabet Res Clin Pract. 2020;167:108383. doi:10.1016/j.diabres.2020.108383
9. Davis HE, McCorkell L, Vogel JM, Topol EJ. Long COVID: major findings, mechanisms and recommendations. Nat Rev Microbiol. 2023;21(3):133–146. doi:10.1038/s41579-022-00846-2
10. Yong SJ. Long COVID or post-COVID-19 syndrome: putative pathophysiology, risk factors, and treatments. Infect Dis. 2021;53(10):737–754. doi:10.1080/23744235.2021.1924397
11. Hagiya H, Tokumasu K, Otsuka Y, et al. Relevance of complement immunity with brain fog in patients with long COVID. J Infect Chemother. 2024;30(3):236–241. doi:10.1016/j.jiac.2023.10.016
12. Cervia-Hasler C, Bruningk SC, Hoch T, et al. Persistent complement dysregulation with signs of thromboinflammation in active long covid. Science. 2024;383(6680):eadg7942. doi:10.1126/science.adg7942
13. Ma L, Sahu SK, Cano M, et al. Increased complement activation is a distinctive feature of severe SARS-CoV-2 infection. Sci Immunol. 2021;6(59). doi:10.1126/sciimmunol.abh2259.
14. Ajjan RA, Schroeder V. Role of complement in diabetes. Mol Immunol. 2019;114:270–277. doi:10.1016/j.molimm.2019.07.031
15. Flyvbjerg A. The role of the complement system in diabetic nephropathy. Nat Rev Nephrol. 2017;13(5):311–318. doi:10.1038/nrneph.2017.31
16. King BC, Blom AM. Non-traditional roles of complement in type 2 diabetes: metabolism, insulin secretion and homeostasis. Mol Immunol. 2017;84:34–42. doi:10.1016/j.molimm.2016.12.009
17. Pal R. COVID-19, hypothalamo-pituitary-adrenal axis and clinical implications. Endocrine. 2020;68(2):251–252. doi:10.1007/s12020-020-02325-1
18. Lan J, Ge J, Yu J, et al. Structure of the SARS-CoV-2 spike receptor-binding domain bound to the ACE2 receptor. Nature. 2020;581(7807):215–220. doi:10.1038/s41586-020-2180-5
19. Oguz SH, Yildiz BO. Endocrine Disorders and COVID-19. Annu Rev Med. 2023;74(1):75–88. doi:10.1146/annurev-med-043021-033509
20. MS C, Ramesh J. Evaluation of the hypothalamo-pituitary-adrenal axis during the post-COVID-19 period in patients treated with steroids during the illness. Arch Endocrinol Metab. 2023;68:e220207. doi:10.20945/2359-4292-2022-0207
21. Alzahrani AS, Mukhtar N, Aljomaiah A, et al. The Impact of COVID-19 viral infection on the hypothalamic-pituitary-adrenal axis. Endocr Pract. 2021;27(2):83–89. doi:10.1016/j.eprac.2020.10.014
22. Ahmadi I, Estabraghnia Babaki H, Maleki M, et al. Changes in physiological levels of cortisol and adrenocorticotropic hormone upon hospitalization can predict SARS-CoV-2 mortality: a cohort study. Int J Endocrinol. 2022;2022:4280691. doi:10.1155/2022/4280691
23. Yeager MP, Guyre CA, Sites BD, et al. The stress hormone cortisol enhances interferon-upsilon-mediated proinflammatory responses of human immune cells. Anesth Analg. 2018;127(2):556–563. doi:10.1213/ANE.0000000000003481
24. Chung WK, Erion K, Florez JC, et al. Precision medicine in diabetes: a consensus report from the American Diabetes Association (ADA) and the European Association for the Study of Diabetes (EASD). Diabetes Care. 2020;43(7):1617–1635. doi:10.2337/dci20-0022
25. Ji W, Xie X, Bai G, et al. Metabolomic approaches to dissect dysregulated metabolism in the progression of pre-diabetes to T2DM. Mol Omics. 2024;20(5):333–347. doi:10.1039/D3MO00130J
26. Drummond JB, Soares BS, Vieira ELM, et al. Interleukin-6 response to insulin-induced hypoglycemia is associated with hypothalamic-pituitary-adrenal axis activation. J Neuroimmunol. 2020;350:577446. doi:10.1016/j.jneuroim.2020.577446
27. Friedmann AP, Koutychenko A, Wu C, et al. Identification and characterization of a factor Va-binding site on human prothrombin fragment 2. Sci Rep. 2019;9(1):2436. doi:10.1038/s41598-019-38857-4
28. Huber-Lang M, Sarma JV, Zetoune FS, et al. Generation of C5a in the absence of C3: a new complement activation pathway. Nat Med. 2006;12(6):682–687. doi:10.1038/nm1419
29. Wu J, Wu YQ, Ricklin D, et al. Structure of complement fragment C3b-factor H and implications for host protection by complement regulators. Nat Immunol. 2009;10(7):728–733. doi:10.1038/ni.1755
30. Kennedy AT, Schmidt CQ, Thompson JK, et al. Recruitment of factor h as a novel complement evasion strategy for blood-stage plasmodium falciparum infection. J Immunol. 2016;196(3):1239–1248. doi:10.4049/jimmunol.1501581
31. Kowalska D, Kuzniewska A, Senent Y, et al. C5a elevation in convalescents from severe COVID-19 is not associated with early complement activation markers C3bBbP or C4d. Front Immunol. 2022;13:946522. doi:10.3389/fimmu.2022.946522
32. Jarlhelt I, Nielsen SK, Jahn CXH, et al. SARS-CoV-2 antibodies mediate complement and cellular driven inflammation. Front Immunol. 2021;12:767981. doi:10.3389/fimmu.2021.767981
33. Wallis R, Mitchell DA, Schmid R, Schwaeble WJ, Keeble AH. Paths reunited: initiation of the classical and lectin pathways of complement activation. Immunobiology. 2010;215(1):1–11. doi:10.1016/j.imbio.2009.08.006
34. Gaboriaud C, Thielens NM, Gregory LA, et al. Structure and activation of the C1 complex of complement: unraveling the puzzle. Trends Immunol. 2004;25(7):368–373. doi:10.1016/j.it.2004.04.008
35. Dobo J, Pal G, Cervenak L, Gal P. The emerging roles of mannose-binding lectin-associated serine proteases (MASPs) in the lectin pathway of complement and beyond. Immunol Rev. 2016;274(1):98–111. doi:10.1111/imr.12460
36. Mathern DR, Heeger PS. Molecules Great and Small: the Complement System. Clin J Am Soc Nephrol. 2015;10(9):1636–1650. doi:10.2215/CJN.06230614
37. Zarantonello A, Revel M, Grunenwald A, Roumenina LT. C3-dependent effector functions of complement. Immunol Rev. 2023;313(1):120–138. doi:10.1111/imr.13147
38. Skendros P, Germanidis G, Mastellos DC, et al. Complement C3 inhibition in severe COVID-19 using compstatin AMY-101. Sci Adv. 2022;8(33):eabo2341. doi:10.1126/sciadv.abo2341
39. Fang S, Wang H, Lu L, Jia Y, Xia Z. Decreased complement C3 levels are associated with poor prognosis in patients with COVID-19: a retrospective cohort study. Int Immunopharmacol. 2020;89(Pt A):107070. doi:10.1016/j.intimp.2020.107070
40. Jiang H, Chen Q, Zheng S, et al. Association of Complement C3 with clinical deterioration among hospitalized patients with COVID-19. Int J Gen Med. 2022;15:849–857. doi:10.2147/IJGM.S348519
41. Kremlitzka M, Macsik-Valent B, Erdei A. Regulation of B cell functions by Toll-like receptors and complement. Immunol Lett. 2016;178:37–44. doi:10.1016/j.imlet.2016.07.015
42. Marin AV, Cardenas PP, Jimenez-Reinoso A, Munoz-Ruiz M, Regueiro JR. Lymphocyte integration of complement cues. Semin Cell Dev Biol. 2019;85:132–142. doi:10.1016/j.semcdb.2018.02.005
43. West EE, Kolev M, Kemper C. Complement and the regulation of T cell responses. Annu Rev Immunol. 2018;36(1):309–338. doi:10.1146/annurev-immunol-042617-053245
44. Hamazaki K, Nishigaki T, Kuramoto N, Oh K, Konishi H. Secondary adrenal insufficiency after COVID-19 diagnosed by insulin tolerance test and corticotropin-releasing hormone test. Cureus. 2022;14(3):e23021 doi:10.7759/cureus.23021.
45. Miller WL. The hypothalamic-pituitary-adrenal axis: a brief history. Horm Res Paediatr. 2018;89(4):212–223. doi:10.1159/000487755
46. Freire Santana M, Borba MGS, Baia-da-silva DC, et al. Case report: adrenal pathology findings in severe COVID-19: an autopsy study. Am J Trop Med Hyg. 2020;103(4):1604–1607. doi:10.4269/ajtmh.20-0787
47. Leyendecker P, Ritter S, Riou M, et al. Acute adrenal infarction as an incidental CT finding and a potential prognosis factor in severe SARS-CoV-2 infection: a retrospective cohort analysis on 219 patients. Eur Radiol. 2021;31(2):895–900. doi:10.1007/s00330-020-07226-5
48. Kaya MG, Alanli R, Kucukay MB, Ulukaya FB, Bakir F. Pituitary functions after recovery from covid-19. Acta Endocrinol. 2023;19(3):314–318. doi:10.4183/aeb.2023.314
49. Yavropoulou MP, Tsokos GC, Chrousos GP, Sfikakis PP. Protracted stress-induced hypocortisolemia may account for the clinical and immune manifestations of Long COVID. Clin Immunol. 2022;245:109133. doi:10.1016/j.clim.2022.109133
50. Isidori AM, Minnetti M, Sbardella E, Graziadio C, Grossman AB. Mechanisms in endocrinology: the spectrum of haemostatic abnormalities in glucocorticoid excess and defect. Eur J Endocrinol. 2015;173(3):R101–13. doi:10.1530/EJE-15-0308
51. Ferrante E, Serban AL, Clerici M, et al. Evaluation of procoagulant imbalance in Cushing’s syndrome after short- and long-term remission of disease. J Endocrinol Invest. 2022;45(1):9–16. doi:10.1007/s40618-021-01605-5
52. Swiatkowska-Stodulska R, Mital A, Wisniewski P, et al. Assessment of platelet function in endogenous hypercortisolism. Endokrynol Pol. 2015;66(3):207–213. doi:10.5603/EP.2015.0014
53. Fischli S, von Wyl V, Wuillemin W, et al. Impact of adrenal function on hemostasis/endothelial function in patients undergoing surgery. J Endocr Soc. 2021;5(5):bvab047. doi:10.1210/jendso/bvab047
54. Kastelan D, Dusek T, Kraljevic I, et al. Hypercoagulability in Cushing’s syndrome: the role of specific haemostatic and fibrinolytic markers. Endocrine. 2009;36(1):70–74. doi:10.1007/s12020-009-9186-y
55. Martin-Rojas RM, Perez-Rus G, Delgado-Pinos VE, et al. COVID-19 coagulopathy: an in-depth analysis of the coagulation system. Eur J Haematol. 2020;105(6):741–750. doi:10.1111/ejh.13501
56. Fogarty H, Townsend L, Ni Cheallaigh C, et al. COVID19 coagulopathy in caucasian patients. Br J Haematol. 2020;189(6):1044–1049. doi:10.1111/bjh.16749
57. Yang K, Holt M, Fan M, et al. Cardiovascular Dysfunction in COVID-19: association Between Endothelial Cell Injury and Lactate. Front Immunol. 2022;13:868679. doi:10.3389/fimmu.2022.868679
58. Valencia I, Lumpuy-Castillo J, Magalhaes G, et al. Mechanisms of endothelial activation, hypercoagulation and thrombosis in COVID-19: a link with diabetes mellitus. Cardiovasc Diabetol. 2024;23(1):75. doi:10.1186/s12933-023-02097-8
59. Zhang L, Lang J, Jin L, et al. Effect of hirsutella sinensis fungus on the hypothalamic-pituitary-adrenal axis in lewis rats with kidney-yang deficiency syndrome. Evid Based Complement Alternat Med. 2020;2020(1):5952612. doi:10.1155/2020/5952612
60. Francis K, Lewis BM, Monk PN, Ham J. Complement C5a receptors in the pituitary gland: expression and function. J Endocrinol. 2008;199(3):417–424. doi:10.1677/JOE-08-0110
61. Francis K, Lewis BM, Akatsu H, et al. Complement C3a receptors in the pituitary gland: a novel pathway by which an innate immune molecule releases hormones involved in the control of inflammation. FASEB J. 2003;17(15):2266–2268. doi:10.1096/fj.02-1103fje
62. Sardu C, D’Onofrio N, Balestrieri ML, et al. Outcomes in patients with hyperglycemia affected by COVID-19: can we do more on glycemic control? Diabetes Care. 2020;43(7):1408–1415. doi:10.2337/dc20-0723
63. Guo W, Li M, Dong Y, et al. Diabetes is a risk factor for the progression and prognosis of COVID-19. Diabetes Metab Res Rev. 2020;36(7):e3319. doi:10.1002/dmrr.3319
64. Nougier C, Benoit R, Simon M, et al. Hypofibrinolytic state and high thrombin generation may play a major role in SARS-COV2 associated thrombosis. J Thromb Haemost. 2020;18(9):2215–2219. doi:10.1111/jth.15016
65. Li X, Weber NC, Cohn DM, et al. Effects of hyperglycemia and diabetes mellitus on coagulation and hemostasis. J Clin Med. 2021;10(11):2419. doi:10.3390/jcm10112419
66. Paz Ocaranza M, Riquelme JA, Garcia L, et al. Counter-regulatory renin-angiotensin system in cardiovascular disease. Nat Rev Cardiol. 2020;17(2):116–129. doi:10.1038/s41569-019-0244-8
67. Lanza K, Perez LG, Costa LB, et al. Covid-19: the renin-angiotensin system imbalance hypothesis. Clin Sci (Lond). 2020;134(11):1259–1264. doi:10.1042/CS20200492
 © 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms.php
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms.php
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.