")
Back to Journals » International Journal of Nanomedicine » Volume 20
Wolf in Sheep’s Clothing: Taming Cancer’s Resistance with Human Serum Albumin?
Authors Stukan I , Żuk A , Pukacka K, Mierzejewska J, Pawłowski J, Kowalski B, Dąbkowska M
Received 5 November 2024
Accepted for publication 26 February 2025
Published 19 March 2025 Volume 2025:20 Pages 3493—3525
DOI https://doi.org/10.2147/IJN.S500997
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Dong Wang
Iga Stukan,1 Anna Żuk,2 Kamila Pukacka,3 Julia Mierzejewska,4 Jakub Pawłowski,4 Bogusław Kowalski,4 Maria Dąbkowska4
1Department of General Pathology, Pomeranian Medical University in Szczecin, Szczecin, Poland; 2Independent Laboratory of Community Pharmacy, Pomeranian Medical University in Szczecin, Szczecin, Poland; 3Department of Pharmaceutical Technology, Pomeranian Medical University in Szczecin, Szczecin, Poland; 4Independent Laboratory of Pharmacokinetic and Clinical Pharmacy, Pomeranian Medical University in Szczecin, Szczecin, Poland
Correspondence: Maria Dąbkowska, Independent Laboratory of Pharmacokinetic and Clinical Pharmacy, Pomeranian Medical University in Szczecin, Szczecin, Poland, Email [email protected] Iga Stukan, Department of General Pathology, Pomeranian Medical University in Szczecin, Szczecin, Poland, Email [email protected]
Abstract: Human serum albumin (HSA) has emerged as a promising carrier for nanodrug delivery, offering unique structural properties that can be engineered to overcome key challenges in cancer treatment, especially resistance to chemotherapy. This review focuses on the cellular uptake of albumin-based nanoparticles and the modifications that enhance their ability to bypass resistance mechanisms, particularly multidrug resistance type 1 (MDR1), by improving targeting to cancer cells. In our unique approach, we integrate the chemical properties of albumin, its interactions with cancer cells, and surface modifications of albumin-based delivery systems that enable to bypass resistance mechanisms, particularly those related to MDR1, and precisely target receptors on cancer cells to improve treatment efficacy. We discuss that while well-established albumin receptors such as gp60 and gp18/30 are crucial for cellular uptake and transcytosis, their biology remains underexplored, limiting their translational potential. Additionally, we explore the potential of emerging targets, such as cluster of differentiation 44 (CD44), cluster of differentiation (CD36) and transferrin receptor TfR1, as well as the advantages of using dimeric forms of albumin (dHSA) to further enhance delivery to resistant cancer cells. Drawing from clinical examples, including the success of albumin-bound paclitaxel (Abraxane) and new formulations like Pazenir and Fyarro (for Sirolimus), we identify gaps in current knowledge and propose strategies to optimize albumin-based systems. In conclusion, albumin-based nanoparticles, when tailored with appropriate modifications, have the potential to bypass multidrug resistance and improve the targeting of cancer cells. By enhancing albumin’s ability to efficiently deliver therapeutic agents, these carriers represent a promising approach to addressing one of oncology’s most persistent challenges, with substantial potential to improve cancer treatment outcomes.
Keywords: HSA, drug delivery system, drug resistance, nanomedicine, gp60, HSA dimer, pazenir, fyarro, abraxane
Introduction
Human serum albumin (HSA) is among the most extensively studied carriers in cancer therapy because of its unique characteristics, such as biocompatibility and biodegradability.1 Albumin is also actively scavenged by cancer cells due to its high capacity for fatty acid binding and its glutamine content, which makes up approximately 10% of its amino acid composition. This makes albumin a valuable source for fueling the Krebs cycle and lipid biosynthesis, as well as providing nitrogen necessary for amino acid and nucleotide synthesis.2–7 Interestingly, hypoalbuminemia in cancer is frequently associated with cancer chemoresistance due to reduced drug transportation (reviewed in).8
Albumin structure is amenable to modifications, also allows for the efficient design of active targeting strategies, specifically directed at particular receptors. Thus, it enables both passive and active (via receptor-mediated pathways) targeting of cancer cells.
Passive targeting results from the prolonged circulation time of human serum albumin (HSA). The growth of cancerous tissue causes local swelling (due to a reduction in lymphatic flow), increasing the accumulation of albumin in the interstitial space. This accumulation is caused by a significant increase in vascular permeability called the enhanced permeability and retention (EPR) effect, reported by Maeda and Matsumura in 1986. Hoogenboezem and Duvall provided a comprehensive review of the relationship between the EPR effect and albumin accumulation in tumor tissues, highlighting its role as a drug carrier in oncology.9 Despite passive drug targeting, clinical evidence suggests that tumor accumulation of organic compounds, including albumin-based nanoparticles, typically only reaches less than 1% of the administered dose, and satisfactory outcomes are seldom achieved solely through the enhanced permeability and retention effect strategy.10,11
This is because mere accumulation at the target site is insufficient for effective drug delivery. The issue of drug penetration into tumor tissue remains complex and is dependent on factors such as the tumor’s vascularization. Furthermore, a critical question is – is it possible for the nanoparticle or drug to enter the cancer cell to excerpt therapeutic effect? To precisely address this question and tackle it effectively, it is essential to understand how a cancer cell internalizes a given particle and whether the process could be enhanced or bypassed. Targeting and uptake can be improved by incorporating ligands into the nanoparticle that are known to effectively bind to receptors on the cancer cell surface. On the other hand, resistance mechanisms can actively expel the particle from the cell, preventing it from interfering with the cancer cell’s life pathways.
It is estimated that therapy resistance is responsible for almost 90% of deaths caused by cancer.12 The resistance can be pre-existing, also described as primary, innate or intrinsic. This means that the patient is characterized with a certain quality of cancer cells or tissues that causes therapy to be ineffective. Also, in terms of chemotherapy, such resistance is present before the treatment. On the other hand, due to the extraordinary plasticity, cancer cells can get resistant during the treatment. Such resistance is called acquired, secondary or extrinsic.13 One of the factors that can emerge as a result of both primary and secondary resistance is perturbation of cell influx/efflux mechanisms. In 1976, Juliano and Ling reported a specific membrane protein of 170 kDa, which was the reason for colchicine resistance in cell culture.14 They called it P-glycoprotein (P-gp, multiple drug resistance type 1 – MDR1). MDR1 belongs to the ATP-binding cassette (ABC) transporters, responsible for the distribution, absorption and secretion of unspecific substrate repertoire.15,16 Once MDR1 is expressed on a cancer cell, its nonspecificity enables cross-resistance to various drugs, a phenomenon known as multidrug resistance. MDR1 is not only a concern confined to cytostatic therapy; evidence has demonstrated that certain targeted therapeutics, such as tyrosine kinase inhibitors, are able to interact with MDR pumps.16 Using nanoparticles that prevent expulsion through MDR, enable drug release within the cancer cell and are readily internalized by the cancer cell, like albumin, a seemingly essential nutrient, may serve as a “wolf in sheep’s clothing”—a strategy that deceives cancer cells and significantly hinders the development of drug resistance.
This review focuses on the unique molecular features of human serum albumin that make it a versatile carrier for drug delivery, particularly in overcoming cancer cell resistance. While much has been written about the general effectiveness and methods for synthesizing albumin-based delivery systems,9,11,17–20 our review takes an interdisciplinary approach, integrating insights from chemistry, cell biology, and translational medicine. We highlight how specific chemical modifications to albumin can improve its ability to bypass resistance mechanisms, especially those related to MDR1 (Figure 1). By examining the interplay between albumin’s chemical properties, its cellular interactions, and the potential for manipulating these to overcome specific oncological barriers, we emphasize the translational relevance of this work. Rather than focusing solely on how the carrier reaches cancer cells, we investigate how the choice of receptor can influence the success of overcoming resistance mechanisms. Drawing on key studies, including the clinically used formulation Abraxane and new formulations – Pazenir and Fyarro, we identify areas where current knowledge is lacking and propose strategies to enhance the therapeutic impact of albumin-based drug delivery systems. Our goal is to shed light on how albumin can be optimized to break through cancer resistance and improve treatment outcomes. Although various sources of albumin are commonly used (as reviewed extensively in21), our attention will be given to human origin, as there is some evidence that the source of albumin carrier might influence the risk of allergic reactions,20 which is highly important for potential use in immunotherapy.9
 |
Figure 1 Human serum albumin as a modifiable carrier to target cancer cells. Folate receptor, gp60 (glycoprotein 60), CD44 (cluster of differentiation 44), and FcRn (neonatal Fc receptor) are involved in the uptake and transport of albumin-based carriers. |
Basic Properties of Human Serum Albumin
Human Serum Albumin Function
Human albumin is the most abundant, with an average concentration of 42 grams per liter (equivalent to 632 µM)22 and a half-life of 19 days, which allows it to maintain a prolonged presence in circulation.23 During its life of 27 days, an average albumin molecule makes about 15000 trips around the circulatory system plus about 15 trips through extravascular spaces.24,25 Approximately 120 grams of albumin are present within blood vessels, and due to its relatively low molecular weight compared to other plasma proteins, it plays a crucial role in maintaining about 80% of the colloid osmotic pressure. This function facilitates the movement of fluids between tissues and the bloodstream, helping to retain fluid within the blood vessels.19,26,27
The highly organized structure and multiple binding sites of HSA enable it to transport various compounds, including ions, fatty acids, hormones, and other molecules, serving as a high-capacity reservoir that stabilizes the concentration of free ligands. Albumin is indeed a crucial protein in the human body functions primarily as a carrier protein, which is vital for transporting various substances in the bloodstream and controlling their active concentration, making them an important part of drug delivery.28–30 The transport of metabolites involves uptake by albumin at one organ and release to the cells of another. However, the failure of protein therapy in clinical trials has been attributed to poor knowledge of protein pharmacokinetics, as well as the uncontrolled spread of proteins when they are administered systemically.
Furthermore, HSA extends beyond its traditional functions in transport, osmotic regulation, and detoxification, playing a crucial role in cellular defense through its significant antioxidant properties. HSA is central to combating oxidative stress, a process closely linked to various diseases and aging. It contributes to cellular antioxidant defense by scavenging free radicals, binding metals, participating in redox reactions, and preventing lipid peroxidation.31–33 Additionally, HSA serves as a biomarker for several diseases, including cancer, rheumatoid arthritis, ischemia, and obesity, and is clinically used to treat conditions such as blood loss, hemorrhage, chronic and acute liver failure, and hypoalbuminemia.34
Human Serum Albumin Structure
The liver synthesizes HSA, a non-glycosylated protein chain of 585 amino acids with a molar mass of 66,439 Daltons.35 Human serum albumin and its derivatives, including recombinant HSA (rHSA) and the albumin dimer (dHSA), have well-known primary and secondary chemical structures. These molecules have a globular shape and are relatively stable under normal physiological conditions.
The crystalline structure of HSA is predominantly composed of α-helix formations, which make up 69% of its overall structure. The sequence of albumins is characterized by a unique arrangement of subunits linked by disulfide bonds. This configuration includes 35 cysteine residues that are integral in forming 17 disulfide bridges, which are crucial for maintaining the molecule’s spherical stability.31,33
The molecule is organized into three repeating domains: I (residues 1 to 195), II (residues 196 to 383), and III (residues 384 to 585), each displaying a similar structural motif (Figure 2).
 |
Figure 2 Modular domain structure of the HSA molecule. The panel illustrates the structure of the HSA molecule, with its subdomains distinctly color-coded. The molecule is organized into three repeating domains, each exhibiting a similar structural motif: Domain I (IA: residues 1–112, IB: residues 112–195), Domain II (IIA: residues 196–303, IIB: residues 303–383), and Domain III (IIIA: residues 384–500, IIIB: residues 500–585). |
Across these three domains, a total of ten helices are arranged in antiparallel, presenting a surface of hydrophobic residues oriented toward the interior of each domain. These helices are further grouped into two subdomains: Subdomain A consists of six helices (h1 to h6), whereas Subdomain B contains four helices (h7 to h10).5,30,36,37
The amino acid composition of albumins is distinguished by a low abundance of tryptophan, glycine, and methionine, along with a high concentration of cystine and charged amino acids, such as aspartic acid, glutamic acid, lysine, and arginine.25 The albumin molecule is highly polar, possessing approximately 100 negative and 82 positive charges, with the net negative charge being most pronounced in the N-terminal domain and lowest in the C-terminal domain, resulting in an asymmetric charge distribution. Human albumin’s high solubility and stability are attributed to its overall net charge of −15 at neutral pH and the numerous disulfide bonds within its structure. Therefore, developing an HSA-based delivery system requires determining the electrokinetic charge of HSA, as this information is crucial for understanding drug adsorption mechanisms and ensuring optimal functionality. It is important to note that both electrokinetic and surface charges should be understood as net (averaged) values representative of the entire molecule, forming regions of positive and negative charges across a broad pH range.Various techniques are employed to estimate or predict a protein’s isoelectric point (IEP), with one approach involving computational methods that consider factors such as crystallographic structure, electrostatic interactions, and amino acid properties to determine the IEP of human serum albumin, which ranges from 5.67 to 5.9.38,39 When dispersed in aqueous solutions, HSA has a positive electrokinetic charge at pH values below 5 because of specific amino acid residues, such as cysteine (35), tyrosine (19), lysine (59), and arginine (23). However, when the pH of these solutions reaches 5, the electrokinetic charge of the molecule becomes negative, primarily because of the presence of glutamic acid (59) and aspartic acid (41) residues.40 This shift establishes the molecule’s isoelectric point at 5.1.41 Depending on the methodology used, the isoelectric point values for human albumin can range from 4.7 to 5.8.38,40,42,43 The differences observed when comparing experimental and computational methods for determining IEP can be attributed to variations in amino acid composition, the presence of fatty acids, or potential errors in measurement techniques.44–46
Ligands Binding and Functional Sites
A defining characteristic of albumin is its ability to reversibly bind a wide range of endogenous and exogenous ligands. Most of these interactions rely on hydrophobic bonds that affect albumin’s structural flexibility, enabling it to adapt its shape to accommodate various ligands, which likely explains its extensive binding capacity. The majority of ligands bind reversibly, with typical association constants (Ka) ranging from 104 to 106 M−1.
Studying the pH-dependent conformational isomerization of HSA offers valuable insights into its properties and interactions with various molecules, including drugs. These pH-induced changes in HSA are particularly significant in pharmacology and biochemistry, as shifts in pH lead to notable alterations in protein shape and structure. The abundance of acidic and basic amino acid residues in HSA allows it to undergo reversible conformational changes across different pH values (Figure 3).
 |
Figure 3 Proposed configurations and interrelation of the N, F, and E isomeric forms of HSA by Carter and Ho (1994) and Peters (1985). |
At pH values below 2.7, HSA takes on an elongated structure known as the “E-form.” Interestingly, between pH 2.7 and 4.3, HSA adopts a different form, termed the “F-form”, which is characterized by increased viscosity, reduced solubility, and a decrease in the alpha-helix content. From pH 4.3 to 8.0, HSA primarily exists in the “N-form”, recognized by its heart-shaped structure. When the pH exceeds 8.0, HSA transitions to the “B-form”, which is characterized by a reduced alpha-helix content and a greater affinity for certain ligands than the “N-form”.30,34 The spherical arrangement of domains in HSA gives each HSA monomer allosteric properties, enabling it to interact effectively with a diverse array of ligands. This allosteric nature allows HSA to undergo conformational changes, which alter its binding sites and enhance its ability to bind and transport various molecules efficiently. This structural flexibility and adaptability in ligand interactions make HSA essential for transporting substances within the bloodstream.
 |
Figure 4 Modification and ligand binding sites on human serum albumin molecules: Tryptophan Residue 214 (Purple), Lysine Residues and N-terminal Primary Amine (Yellow) as Potential mPEG/PEG-NHS Binding Sites, and FA Binding Site (Light Green) with Structural Illustration (Orange) and Labeled Surrounding Amino Acids.37 |
Tryptophan in the HSA Structure
The tryptophan residue in HSA (Figure 4) is particularly noteworthy because of its strategic position near the hydrophobic cavity of subdomain IIA, the principal active binding region of HSA (eg aspirin, warfarin, diazepam, ibuprofen).25 This area also contains two binding sites, Sudlow I and II, known for their significant binding affinities classified based on studies on molecular interactions with fluorescent probes. The binding locus of large heterocyclic compounds possessing a negative charge was identified as Site I, with Site II showing a preference for small aromatic carboxylic acids. Consequently, the tryptophan residue is often utilized in spectroscopic studies and is regarded as a valuable tool in this area of research.36,47,48
Lysines in the HSA Structure
HSA contains 59 lysine residues with free N-termini (primary amino groups) that have become a key focus for structural modifications of the protein (Figure 4). Lys-199 interacts with drugs of Sudlow’s Site I, corresponds to the hydrophobic pocket in subdomain IIA, and his Site II to the pocket within IIIA. The reactivity of Lys-199 may be attributed to its unusually low pKa of 7.9,24,25 which can be rationalized based on its close interaction with His-242. These residues readily react with N-hydroxysuccinimide (NHS) esters and propionaldehydes, allowing significant alterations to the protein structure without causing denaturation. One common modification involves conjugating polyethylene glycol (PEG) or polyethylene glycol monomethyl ether (mPEG) through reactions between the primary amines of lysine residues and NHS esters or propionaldehydes.49,50
Folic Acid Binding Site in the HSA Structure
Human albumin has a binding site for folic acid (FA) located within domain I (Figure 4). The ligand fits into the protein’s structure through noncovalent interactions, primarily hydrogen bonds. Structural analysis revealed that folic acid binds to HSA through a combination of hydrophilic and hydrophobic contacts, with a total binding constant in the range of 104 to 105 M−1.51 In the folic acid-HSA complex, folic acid is surrounded by Ile-142, Leu-185, Tyr-138, and Trp-161.51 These findings indicate that HSA can effectively bind and transport folic acid, highlighting its potential for further exploration in the context of folic acid utilization in complex with HSA.
Albumin in Cancer
Despite more than 30 years since identification of albumin receptors, and numerous studies and scientific reviews on their expression and function,1,9,19,34,52,53 the precise mechanisms by which they operate remain poorly understood.54,55 This knowledge is however crucial to identify cancer types that respond efficiently to albumin-based drug delivery systems and also to improve efficacy of existing therapies, also in the context of drug resistance.56
Mechanisms of Albumin Uptake in Healthy and Cancer Cells
Albumin is not merely a passive protein circulating in bodily fluids, it plays a dynamic role in cellular physiology, tailored to the needs of the environment. Albumin uptake operates through two general mechanisms: endocytosis and micropinocytosis. Endocytosis is specific to substrate-receptor binding, which involves frequently auxillary role of clathrin or caveolin-1 (CAV1). Clathrin-mediated endocytosis operates as a stochastic process, suggesting a more random and ubiquitous mechanism for internalizing molecules, whereas CAV1-dependent endocytosis is spatially restricted to lipid raft regions, indicating a more organized and compartmentalized pathway. This distinction highlights the potential for differential targeting strategies in drug delivery, depending on the desired endocytic route and cellular localization.57,58 Albumin uptake in healthy cells is primarily mediated by caveolin-1 (CAV-1), however clathrin-mediated endocytosis and macropinocytosis-like processes have been described in pulmonary epithelium or skeletal muscle primary cells.59,60 Macropinocytosis is not specific to the substrate, but it involves cytoskeleton movement it is actin-dependent. This process enables sampling of large volumes of extracellular fluid and is considered a preferred mechanism for albumin scavenging by cancer cells.61 It is a form of nutrient sensing and may facilitate survival in nutrient-poor environments.2,4,7
Cancer patients often experience hypoalbuminemia, a condition associated with cachexia.62 This reduction in albumin levels is thought to stem, at least in part, from the disruption of albumin recycling mechanisms that normally maintain its balance in bodily fluids. Under physiological conditions, this recycling is orchestrated by glycoprotein 60 (gp60), the neonatal fragment crystallizable (Fc) receptor (FcRn) and the cubulin/megalin complex in kidneys. However, in the tumor microenvironment, these mechanisms face significant challenges. Driven by the demands of unchecked growth, tumor cells scavenge albumin from their environment. They employ micropinocytosis and upregulate receptor-mediated pathways to access essential nutrients, such as fatty acids and iron. Notably, receptors like glycoprotein 18/30 (gp18/30), Cluster of Differentiation 36 (CD36) or Transferrin receptor protein 1 (TfR1) play key roles in this process, making them attractive targets for therapeutic strategies (Table 1). A deeper understanding of these pathways is essential to justify modifications that circumvent resistance mechanisms, paving the way for more effective interventions.
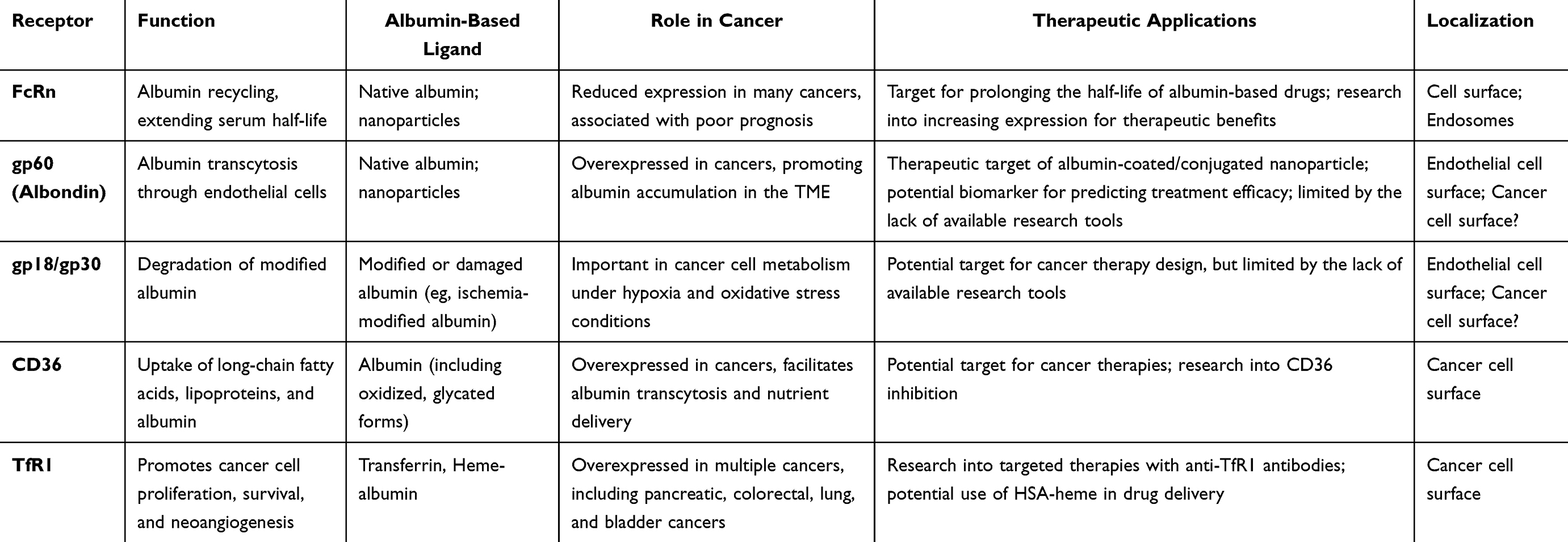 |
Table 1 The Ability of Selected Albumin-Recognizing Receptors to Improve Cancer Cell’s Targetability and Nanoparticle Design |
gp60
The first and most described receptor responsible for native albumin binding is gp60, also known as albondin. It serves as a key mediator of native albumin transcytosis across endothelial cells, which is essential for maintaining vascular oncotic pressure and delivering nutrients bound to albumin to tissues where they are needed, including tumor microenvironment63 (Table 1). Binding of albumin to gp60 initiates a CAV-1-dependent formation of caveolae – a structure that facilitates transcellular transport (Figures 5 and 6).64 CAV-1 correlates with a favorable response to Abraxane, suggesting that gp60 might be involved in Abraxane uptake, it is unclear whether gp60 would be able to recognize albumin-based nano carriers.64 Despite its importance, research on gp60 remains limited by the lack of specific tools to study its mechanisms in detail.56,65 Several sources reference a 1984 paper by Schnitzer et al, describing a sequence of gp60 derived from bovine leukemia virus envelope proteins,66 later identified as the gp60 albumin receptor.53 However, up to date this sequence is not deposited in any database. Some groups have reported the use of antibodies for immunofluorescent labeling directed against gp60/110 isolated from human herpesvirus 6,56,67 but it is not suggested by the antibody producer that it can be used to visualize human gp60. Another discrepancy in the literature concerns the localization of the gp60 receptor. While it is widely accepted that gp60 receptors are located on endothelial cells, some sources, perhaps erroneously, report their presence on the surface of cancer cells.68 This possibility cannot be entirely excluded, given the lack of appropriate investigative tools and limited data on gp60 expression. Understanding the nuances of gp60-mediated albumin transcytosis could reveal new therapeutic targets, particularly in diseases where albumin transport is co-opted for pathological purposes.
 |
Figure 5 Gp60 recycles undamaged albumin. Albumin is recognized by gp60 glycoprotein receptors on the cell surface (I). After binding, albumin is internalized via caveolin-1-mediated endocytosis (II) and becomes entrapped in an endosome (III). Because pH changes due to H+ ATPase activity, the FcRn receptor located in the endosome subsequently recognizes albumin (IV), which prevents it from further lysosomal degradation. Endosomes merge with the cellular membrane, and the pH increases, resulting in albumin release (V). |
 |
Figure 6 Pathways of albumin acquisition by cancer cells. Albumin or albumin-based nanoparticles present in the bloodstream (A) can enter the tumor interstitial space through the leaky vasculature or via transcytosis mediated by the gp60/caveolin-1/FcRn pathway. (B) Alternatively, albumin may be internalized by macropinocytosis, where the invagination of the cell membrane encloses both albumin and the FcRn receptor within an endosome. This process protects albumin from degradation, allowing it to be released into the extracellular milieu. A detailed description of this process is provided in Figure 5. In the interstitial space, albumin or albumin-based nanoparticles can interact with the SPARC (secreted protein acidic and rich in cysteine) protein and be endocytosed in a clathrin-dependent manner, potentially entering the lysosomal degradation pathway. (C) An additional pathway involves the gp18/30 system, where albumin or albumin-based nanoparticles are internalized and degraded within the lysosomal compartment. This process is further detailed in Figure 7. |
Neonatal Fc Receptors
Albumin recycling is possible because of FcRn receptors, that are widely expressed in various tissues.69–75 In mice, FcRn deficiency caused rapid albumin turnover and 50% reduction in the half-life in blood serum.76,77 This effect was irrespective of increased albumin production in those animals.26,76
FcRn-mediated albumin recycling is an elegant, pH-dependent mechanism (Figure 5) Figure 3. FcRn does not interact with albumin at neutral pH (pH 7.0), but binds it efficiently at acidic pH (6.5–5.5). When albumin is internalized either by gp60-receptor or by micropinocytosis, it enters a cell with an endosome at neutral pH. When pH within endosome decreases, it activates FcRn to bind albumin. This event prevents albumin degradation in the lysosome. The endosome then releases its contents into the serum, where the pH is approximately 7.0–7.4. The binding affinity between albumin and FcRn decreases again, allowing the release of albumin back into the extracellular space.27,77,78 Notably, the tumor extracellular microenvironment is characterized by intense acidification, with pH values ranging between 6.4 and 7.0.79 While this acidic milieu theoretically may influence the release of albumin into the extracellular space, this aspect remains unexplored in the literature. The literature is also inconsistent in the meaning of the FcRn expression level in cancer. There is a significant reduction in FcRn expression, associated with poor prognosis in many cancer types.80,81 Swiercz et al has also demonstrated that increased FcRn expression prevents albumin scavenging and instead promotes albumin recycling, which reduces glutamine levels in cancer cells and decreases the number of metastases.80 In contrast, Larsen et al reported that FcRn overexpression in human cancer cells promotes albumin recruitment and cellular growth in breast and colorectal cancer cell lines. As proof of principle, human FcRn knockout in a mouse xenograft significantly decreased the tumor volume. Additionally, the authors reported that FcRn was significantly overexpressed at the protein level in eight out of ten investigated cancer types, including breast, cervical, pancreatic, lung, colorectal, head and neck and renal cancer. In the discussion, the authors underline the need for further research in terms of albumin acquisition by cancer cells in the context of future targeted albumin-based therapies.82 The evidence points to a deeper story, where the interaction between FcRn and cancer emerges as a pivotal factor in shaping the success of albumin-based therapies. This discrepancy hints that the therapeutic outcome may depend significantly on FcRn expression levels, varying across cancer types and individual patients.
SPARC
Research conducted from the late 1990s revealed that secreted protein acidic and rich in cysteine (SPARC, osteonectin, BM-40) is highly expressed in stromal tissue and various advanced cancers.83–85 Indeed, SPARC plays a significant role in neovascularization,53,86,87 metastasis, and cancer cell survival, underscoring its critical contribution to tumor progression.53,88–90 SPARC regulates cancer interactions with the extracellular matrix (ECM) and has a strong affinity for albumin, which can lead to albumin accumulation in inflamed and tumorous tissues.91 SPARC-bound albumin facilitates the gp60/CAV-1-mediated transcytosis and simultaneously protects and directs albumin to cancer cells (Figure 6). Main SPARC-albumin receptor on the cancer cell surface is α5β1 integrin complex.60,92 The internalization of SPARC depends on clathrin, and the subsequent fate of albumin remains uncertain.60,93 SPARC regulates transport from early endosomes to late endosomes, suggesting that albumin enters the lysosomal pathway.3,52,60,94 However, some studies report that albumin may become trapped in endolysosomal or autophagosomal compartments.95 Interestingly, Abraxane appears to be rescued from lysosomal degradation by SPARC, which inhibits the cytotoxic effect of Abraxane but not paclitaxel.65
gp18/gp30
In contrast to gp60, gp18/30 are membrane receptors that bind modified or damaged albumin,96–98 for instance ischemia-modified albumin (IMA).99 These receptors were identified via agarose gel electrophoresis98 and first described by Schnitzer JE & Bravo J in 1986.98 After receptor recognition, albumin is internalized through macropinocytosis and directed to the lysosomal degradation pathway, making it a key route used by cancer cells for albumin scavenging (Figure 7). Damaged vasculature and the pressure exerted by the growing tumor on surrounding tissues, leads to inadequate nutrient supply through blood vessels. As a result, tissues resort to alternative means for essential amino acids to suffice cellular anabolism. It is unknown to what extent are those receptors involved in HSA nanoparticles intake, however Abraxane, initially hypothesized to be internalized through the gp60/CAV-1 system, is actually transported via the denatured albumin transport system involving gp18/30.65 The pathway of nanoparticle internalization is critical, as it determines the intracellular fate of the cargo. While degradation before reaching the molecular target can reduce therapeutic efficacy, this is not the case for Abraxane. Transport via gp18/30 still allows for efficient delivery of paclitaxel, outperforming administration without a carrier. This mechanism also holds significance for overcoming treatment resistance, a topic explored further in the subsequent sections.
 |
Figure 7 Gp18/30 recognizes damaged albumin and targets it for degradation. Damaged albumin is recognized by glycoprotein receptors on the cell surface, specifically by gp18/30 (I). Upon binding, albumin is internalized via clathrin-mediated endocytosis (II) and becomes entrapped in an endosome (III). The activity of H+ ATPase leads to a reduction in pH (IV). The endosome subsequently merges with the lysosome, and degradation enzymes are released (IV). Albumin is digested into amino acids, and its fatty acid cargo is salvaged which can be further used by cancer cells (V). |
CD36
The interaction between albumin and the cluster of differentiation 36 (CD36) transmembrane receptor is well-documented in the literature, yet therapeutic approaches targeting this specific interaction remain largely unexplored. Cancer cells, notably, often overexpress CD36, a receptor essential for the uptake of long-chain fatty acids and lipoproteins, and which plays a pivotal role in immunological recognition, inflammation, cell adhesion, and apoptosis.100,101 Albumin, both in its native and modified forms, serves as one of its primary ligands.102 A series of experiments conducted by Raheel et al shed light on the functional relevance of this interaction, demonstrating that CD36 mediates albumin transcytosis across the dermis, and, to a lesser extent, the pulmonary microvascular endothelium. Interestingly, they found that this process of transcytosis could be saturable and exhibited variability depending on the tissue type, suggesting the potential for targeting this pathway in therapeutic applications.54
Transferrin Receptor Protein 1
TfR1 plays a key role in cancer cell survival through NF-κB activation,103 stimulation of neoangiogenesis104 and altered energy metabolism, leading to its overexpression in various cancers.103,105,106 Despite efforts to use anti-TfR1 antibodies for targeted drug delivery, safety concerns have hindered their success.107 TfR1 can also recognizes heme-albumin, facilitating the transport of nutrients or drugs into cells.108 HSA binds heme at two stites: one unaffected by fatty acids.109–111 The role of HSA in heme scavenging and delivery has been extensively reviewed.112 Brell et al demonstrated that TfR1 recognizes heme-albumin in Abraxane, promoting endocytosis and proliferation as efficiently as iron-loaded transferrin. Inhibition of TfR1 with the monoclonal antibody mAb 5–528, educed apoptotic cells by 25%, likely due to impaired endocytosis and reduced cytotoxicity.108 The potential of heme-albumin in targeting tumors compared to unmodified albumin remains to be explored. Additionally, HSA-based nanoparticles may spontaneously incorporate hemin, enabling TfR1-mediated uptake and improving drug delivery in cancer therapy.
Bae et al proposed another potential application of increased TfR expression on cancer cell surfaces. In their study, they developed doxorubicin-loaded nanoparticles functionalized with transferrin using sulfosuccinimidyl-4-(N-maleimidomethyl)cyclohexane-1-carboxylate (Sulfo-SMCC). The transferrin modification aimed to enhance receptor-mediated endocytosis and successfully overcame multidrug resistance in MCF-7/ADR cells, which is significant given that this resistance is typically mediated by MDR1. This highlights the critical importance of targeting TfR1 as a viable strategy for bypassing MDR1-associated resistance and improving therapeutic efficacy in drug-resistant cancers.113
Albumin and Multi Drug Resistance in Cancer
Cancers possess the ability to expel harmful substances from their cells by inducing multidrug resistance (MDR) systems, such as MDR1. When albumin delivers toxic drugs to cancer cells, it raises the question of whether tumors counteract this by upregulating MDR1. Insights into this question come from years of research on Abraxane, the first albumin-based drug approved by the US Food and Drug Administration (FDA) and the European Medicines Agency (EMA). A related and equally critical question is whether albumin can circumvent an already active MDR1 system, effectively delivering the drug despite the presence of functional efflux pumps. These issues will be addressed in this section.
Resistance to Abraxane
Abraxane (ABI-007, nab-paclitaxel) is a 130 nm albumin-bound paclitaxel formulation used for various solid tumors.114 It is twice as effective as unbound paclitaxel, eliminates toxic solvents, and allows for higher dosing.114–116 Paclitaxel is a well-known substrate for MDR1.117 In a Phase 3 clinical trial (NCT00046514), Abraxane was administered to patients with metastatic breast cancer who had likely developed taxane resistance likely due to MDR1 induction. The study demonstrated that Abraxane effectively bypassed cross-resistance, even in patients heavily pretreated with solvent-based paclitaxel or docetaxel.118
 |
Table 2 HSA-Based Nanoparticles in MDR1-Expressing Cancer Cell Lines and Their Outcomes |
The potential of Abraxane to circumvent multidrug resistance (MDR) was further investigated in vitro (Table 2). Zhao et al established a non-small cell lung cancer cell line resistant to Abraxane (A549/Abr) and used RNA-Seq to compare its transcriptome with that of the parental line. They reported significant MDR1 overexpression in A549/Abr cells, a common mechanism of resistance to unbound paclitaxel, and confirmed that MDR1 contributes to resistance by reversing the phenotype with verapamil, an MDR1 inhibitor.119 In another study, Zhao et al analyzed global protein expression changes between treatment-naïve and treatment-resistant A549/Abr cells.120 These authors reported MDR1 overexpression and suggested that resistance arises from the release of paclitaxel from albumin, as MDR1 cannot export the large albumin‒paclitaxel complex. This study identified additional proteins related to lipid metabolism, the cell cycle, the cytoskeleton and apoptosis processes, including novel resistance markers such as the E3-ubiquitin ligase ring finger protein 139 (RNF139) and an enzyme involved in cholesterol synthesis, 3-hydroxy-3-methylglutaryl-CoA synthase 1 (HMGCS1).120 Similarly, Vallo et al reported that MDR1 overexpression in urothelial cell lines caused resistance to nab-paclitaxel.121 While Abraxane remains pioneering, analogous systems have been developed for the controlled delivery of various chemotherapeutics. However, the MDR1 substrate repertoire accommodates a variety of hydrophobic substances. Kong et al tested gemcitabine-loaded HSA nanoparticles in pancreatic cancer cells resistant to gemcitabine due to MDR1 and MRP1 overexpression. The nanoparticles inhibited proliferation and induced apoptosis in these resistant cells.122 The authors proposed that this effect was likely due to ATP consumption during albumin degradation, which limits ATP availability for ATP-dependent ABC transporters such as MDR1 or MRP1.
Onafuye et al developed HSA-based nanoparticles encapsulating doxorubicin and tested them in neuroblastoma cells resistant to vincristine and doxorubicin respectively. The doxorubicin-loaded nanoparticles were effective in MDR1-expressing cell lines, particularly in vincristine-resistant cells. In doxorubicin-resistant cells, the effect was present but less pronounced, possibly because the released drug faces a resistance mechanism similar to that of doxorubicin in solution.123 This highlights the possibility that multiple resistance mechanisms may be involved depending on the specific chemotherapeutic agent used.
Yuan et al investigated triple-negative breast cancer, which is characterized by a high percentage of cancer stem cells (CSCs), which are key drivers of tumor progression and metastasis.124 CSCs are chemoresistant due to mechanisms such as MDR1 overexpression and the activity of detoxifying enzymes, such as aldehyde hydroxygenase (ALDH). The authors reported that while Taxol and Abraxane had similar efficacy in reducing tumor size, Abraxane significantly reduced the frequency of CSCs by 3--9-fold and was more effectively taken up by ALDH+ cells than Taxol.125 Although multidrug resistance has not been specifically studied, the use of albumin as a carrier may enhance chemotherapy outcomes, even in cases of drug resistance.
Overall, despite conflicting evidence, albumin-based delivery systems show promise in facilitating drug transport, potentially overcoming multidrug resistance independent of active MDR1. There are possible specific modifications that would allow for active MDR1 targeting.
HSA-Based Drug Delivery in Presence of Active MDR1
Cancer cells often overexpress MDR1, reducing the efficacy of therapy by decreasing the bioavailability of therapeutics.126 Specific allosteric MDR1 inhibitors (reviewed in127), have been developed, but clinical trials have shown that they can cause significant toxic side effects due to MDR1’s role in regulating the distribution of various substances in the body.128 Therefore, enhancing drug delivery into cells may be more beneficial than inhibiting MDR1 directly (Table 2).
Yang et al developed HSA-based nanoparticles decorated with the EGFR-targeting antibody cetuximab to deliver short interfering RNA (siRNA) targeting MDR1 (siMDR1) and doxorubicin to breast cancer tissues. This nanoparticle successfully reversed multidrug resistance in vitro and in vivo with minimal off-target toxicity.129 Similarly, Liu et al coencapsulated paclitaxel and chemosensitizer-2 in folate-conjugated HSA nanoparticles. This approach aimed to reduce multidrug resistance and improve antitumor efficiency. The nanoparticles showed enhanced target specificity and reduced damage to noncancerous tissues, suggesting that folic acid receptor-mediated uptake limited the action of MDR1 (folic acid as a target for HSA modification is further described in Section 4.3).130
PEG, which is commonly used as an excipient in pharmaceuticals, can be efficiently conjugated to HSA and HSA-core nanoparticles through specific binding sites on albumin. This involves the use of functional groups, such as the -NHS ester of PEG, which bind to primary amines on the free N-termini of lysine residues within the albumin structure. This modification enables HSA nanoparticles to enhance drug delivery and inhibit MDR1 activity, improving drug bioavailability.131,132 Hodaei et al demonstrated that the effectiveness of PEG depends on its molecular weight.133 While the exact mechanism is not fully understood, it may be related to changes in membrane fluidity134,135 and direct inhibition of MDR1 through alkyl and unsaturated C‒C bonds in PEG.136
HSA Utilization in Receptor-Based Targeting Strategies
The intrinsic properties of albumin as a drug carrier offer a unique advantage in bypassing MDR1-mediated drug resistance by facilitating efficient cargo delivery to cancer cells. Yet, the challenge of multidrug resistance extends beyond simple transport—it demands strategies that simultaneously enhance membrane translocation and therapeutic specificity. By addressing these dual requirements, it becomes possible to disrupt the mechanisms that enable cancer cells to expel therapeutic agents. HSA-based nanotherapeutics employ passive targeting, mainly EPR effect, and active targeting strategies. Due to its large molecular size, passive diffusion of albumin is not feasible; instead, tissue accumulation occurs primarily through endocytosis.137 Active targeting involves functionalizing the surface of albumin nanoparticles with ligands that bind to receptors overexpressed on cancer cells (Figure 1). This approach enhances specific targeting of affected tissues, increasing the uptake of drug-loaded nanoparticles by cancer cells while reducing systemic exposure and toxicity. Thanks to albumin’s inherent properties and ease of modification, ligands can be conjugated to the protein, facilitating the internalization of nanoparticles and their cargo via endocytosis (Figure 8).
 |
Figure 8 Utilization of human serum albumin as a versatile drug carrier. |
In pharmaceutical applications, albumin serves as a crucial carrier for drugs through two primary mechanisms: albumin–drug conjugation and drug encapsulation within albumin nanoparticles. Albumin–drug conjugates include albumin-fusion proteins, drugs coupled to exogenous albumin, and prodrugs designed to bind efficiently to endogenous serum albumin. While modified albumin molecules can enhance the pharmacokinetic profiles of drugs, nanoparticle formulations improve their stability and solubility. Fan et al proposed a preclinical optimization strategy that correlates a drug’s binding affinity to albumin with its serum half-life, influencing dosing regimens and therapeutic effectiveness.138
Protein modification techniques can enhance drug delivery precision and reduce chemotherapy side effects. By chemically modifying the hydroxyl, amine, and carboxyl groups on HSA, selective binding to cancer cell receptors can be achieved. Strategies targeting tumor-specific markers, such as transferrin receptors, aptamers, targeting peptides, CD44, folate receptors, and others, have been explored in nanocarrier systems.139 Despite promising reports, only a few of these modifications have been widely implemented (Table 3). Advances in albumin modification, as described in recent studies, represent a critical step toward achieving this objective (Figure 8). In this section, we delve into the innovative modifications of albumin that not only improve drug delivery efficiency but also hold potential to overcome resistance mechanisms, paving the way for more effective cancer therapies.
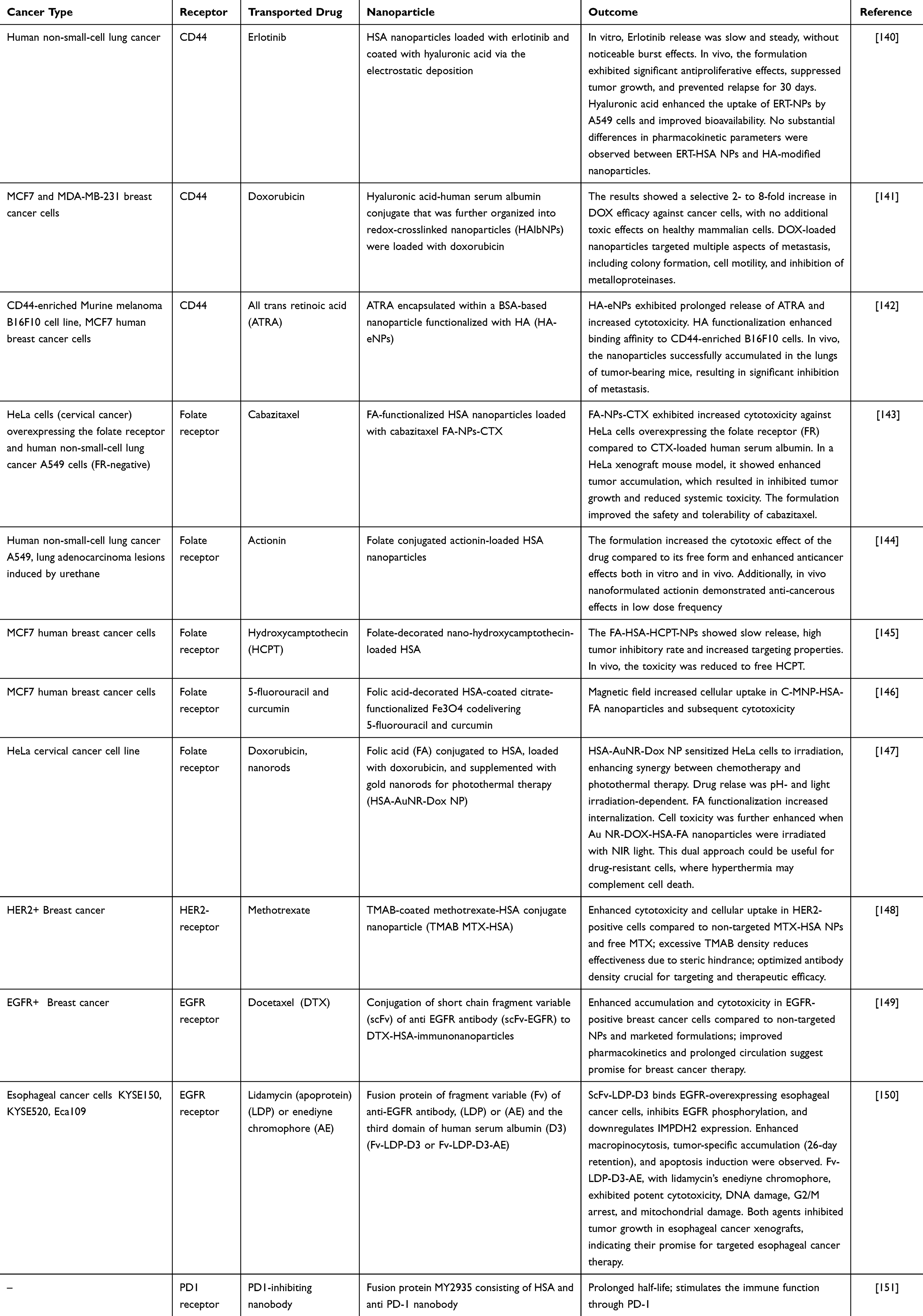 |
Table 3 Examples of Cancer-Specific Modifications of Albumin |
Aptamers
Aptamers are short single-stranded oligonucleotides capable of selectively binding to tumor tissue, offering advantages over monoclonal antibodies, such as lower cost, reduced immunogenicity, and easier high-throughput screening and chemical synthesis.152 However, their small size (approximately 20 kDa) leads to rapid renal clearance, limiting their clinical use. Conjugating aptamers with larger molecules, such as albumin, extends their circulation time in the bloodstream.153 Although aptamers have shown potential to improve cancer therapy (as reviewed in154–156), human serum albumin‒aptamer conjugates have yet to be explored in this context. There have been attempts to enhance immunotherapy with bovine serum albumin conjugated with PD-L1157,158 or CTLA-4 aptamers.159 In non-cancer applications, Zhang et al reported the double conjugation of a newly designed bone anabolic aptamer targeting sclerostin, which exhibits high binding affinity for human serum albumin, thereby extending the system’s half-life.153 These findings, along with results from aptamer-conjugated BSA nanoparticles, underscore the potential of albumin as an aptamer carrier. This research holds significant promise and is crucial for advancing translational oncology.
Hyaluronic Acid (HA)
HA is an unbranched glycosaminoglycan that is an important component of the extracellular matrix. Its synthesis increases in proliferating cells, which indicates its role in the metabolism of cancer cells.160 The HA receptor CD44 is present to varying degrees on multiple cells in the body. It plays a role in cell adhesion and migration, inhibits apoptotic pathways, promotes cell proliferation, and may also enhance early angiogenesis in tumors (detailed in161,162). While present in normal tissue, CD44 is overexpressed in multiple tumors and has been demonstrated to be correlated with increased aggressiveness and a greater risk of metastasis.163 Spadea et al reported that cancer cell lines that express standard CD44 (the shortest CD44 isoform164), which is present in high quantities on the membrane of cancer cells, can efficiently take up HA. Furthermore, owing to heightened receptor activity, the uptake of HA by malignant cells surpasses that of noncancerous cells, as evidenced by various studies.164–166 This phenomenon may enhance drug accumulation within cancerous tissues relative to healthy tissues. Importantly, CD44 is also a molecular marker specific for cancer stem cells (CSCs). Moreover, hyaluronic acid is a nontoxic, biocompatible, and biodegradable polymer, which makes it an excellent candidate for use as a targeting ligand in delivering drugs to tumors, including HSA-based delivery systems (Table 3). Shen Y et al synthesized spherical albumin nanoparticles loaded with erlotinib and coated them with hyaluronic acid via the electrostatic deposition technique. This approach enhanced drug uptake and anticancer effects in the lung cancer cell line A549, which was further confirmed in experiments using a mouse model.140 Brindisi M et al designed a similar system in which they synthesized a hyaluronic acid-human serum albumin conjugate that was further organized into redox-crosslinked nanoparticles (HAlbNPs). The HA-HSA NPs were loaded with doxorubicin (DOX). The authors tested several breast cancer cell lines with various CD44 receptor expressions and evaluated cancer cell viability and the main molecular drug resistance and metastasis pathways. The results demonstrated a selective, 2- to 8-fold increase in the efficacy of DOX against cancer cells, without an increased toxic effect on healthy mammal cells.141 HA demonstrates excellent targeting capabilities for CSCs, as demonstrated by Li et al, through the utilization of all-trans retinoic acid encapsulated within a BSA-based nanoparticle functionalized with HA (HA-eNPs). These HA-eNPs exhibited heightened binding affinity to the CD44-enriched B16F10 cell population and effectively accumulated in the lungs of tumor-bearing mice, leading to significant inhibition of metastasis.142
Folic Acids
Folates play crucial roles in cell growth, survival and proliferation.167 Because of their hydrophilic properties, folates rely on folate receptors (FRs) for the cellular internalization through receptor-mediated transcytosis.168,169 Tumor cells require folates for essential processes, a vulnerability exploited by widely used anticancer drugs such as folate antagonists.170 FOLR1 and FOLR2 folate receptor isoforms are overexpressed in many types of carcinomas, including ovarian, endometrial, kidney, lung, and brain carcinomas; mesothelioma; and myeloid leukemia.171,172 This heightened expression could also be utilized in nanoparticle design (Table 3). Ulbrich et al demonstrated that, in general, cancer cells exhibit greater binding and uptake of HSA nanoparticles conjugated with folic acid than noncancer cells do.173 Sun et al demonstrated that FA-functionalized HSA nanoparticles loaded with cabazitaxel (FA-NPs-CTX), compared with CTX-loaded human serum albumin alone, induced greater cytotoxic effects against HeLa cells overexpressing the folate receptor (FR). Moreover, in a HeLa xenograft mouse model, researchers reported a significant increase in the accumulation of FA-NPs-CTX at the tumor site, leading to inhibited tumor growth and reduced toxicity. Importantly, FA conjugation enhanced the safety and tolerability of cabazitaxel.143 In another study, Ahlawat et al reported that the functionalization of HSA nanoparticles with folic acid enhanced the targeting of hydroxycaptothecin (HCPT) in lung adenocarcinoma and increased the cytotoxic effect of the drug (actinonin) encapsulated within the nanoparticles compared with that of the free drug.144 Wang et al reported a similar enhanced anticancer effect. The authors developed a folate-decorated HSA nanoparticle loaded with nano-HCPT. Although the drug loading efficacy was 7.8%, compared with the free drug, the nanoparticles presented enhanced anticancer effects both in vitro and in vivo. More HCPT was delivered to the tumor tissue, resulting in an increased tumor inhibitory rate.145 FA-decorated nanoparticles can also enhance the delivery of magnetic nanoparticles that contain iron oxide. Hiremath et al designed human serum albumin-coated citrate-functionalized iron oxide decorated with folic acid that also delivers two anticancerous compounds, 5-fluorouracil and curcumin. When a magnetic field was present, the uptake was further enhanced, especially in FA-decorated nanoparticles, increasing the sensitivity of cancer cell lines.146 An intriguing application was presented by Encinas-Basurto et al. The authors introduced a nanoparticle comprising folic acid (FA) conjugated to human serum albumin (HSA), loaded with doxorubicin, and supplemented with gold nanorods for photothermal therapy. This formulation sensitized HeLa cells to irradiation, facilitating synergy between chemotherapeutic and photothermal therapies.147
Antibodies
Monoclonal antibodies (mAbs) have transformed cancer therapy by selectively targeting receptors overexpressed on tumor cells. Examples such as trastuzumab (anti-HER2) and bevacizumab (anti-VEGF) illustrate their role in improving drug delivery.174 However, their potential expands further when integrated with nanocarriers (Table 3). Taheri et al synthesized trastuzumab-decorated methotrexate-albumin nanoparticles (TMAB-MTX-HSA NPs), demonstrating that antibody density on the nanoparticle surface significantly influences cytotoxicity.148 Notably, a moderate density of trastuzumab yielded superior cytotoxic effects compared with high-density configurations, suggesting that overcrowded antibodies may hinder receptor-ligand interactions. This underscores the need for precise control in nanoparticle engineering.
Overcoming Size Limitations with Recombinant Antibodies
Nanoparticle size plays a critical role in cellular uptake and therapeutic efficacy. Ideally, particles ranging from 30 to 50 nm can bypass lysosomal degradation via caveolin-mediated endocytosis.175,176 Larger particles, however, risk lysosomal disposal when taken up through macropinocytosis or phagocytosis.177,178 Recombinant antibody derivatives such as single-chain variable fragments (scFvs), diabodies, and nanobodies address this challenge by offering targeted binding capabilities with reduced particle size.179,180
In an innovative study, Yewale et al developed docetaxel-loaded HSA nanoparticles conjugated with scFvs targeting EGFR-overexpressing breast cancer cells.149 The resulting immunonanoparticles (approximately 160 nm) exhibited enhanced uptake, prolonged circulation, and greater cytotoxicity compared with unconjugated formulations. These results highlight the potential of scFvs to balance specificity and size in antibody-nanoparticle systems.
Further advancements integrate scFvs with albumin to create multifunctional systems. Wang et al combined an EGFR-targeting scFv with domain 3 of HSA and the LDP apoprotein of lidamycin, leveraging the macropinocytosis pathway upregulated in Ras-driven cancers like pancreatic carcinoma.181,182 This bifunctional system not only enhanced drug internalization but also synergistically inhibited EGFR signaling while promoting apoptosis and cell cycle arrest.
Similarly, Xian et al developed a fusion protein of anti-PD-1 nanobody (Nb97) and HSA to extend the half-life of immune checkpoint inhibitors while maintaining PD-1/PD-L1 pathway inhibition.151 This approach demonstrates how HSA can act as both a drug carrier and a stabilizing agent for therapeutic molecules prone to rapid renal clearance.
HSA Dimers
Some studies have demonstrated not only the utility of HSA monomers but also the use of HSA dimers as effective carriers for enhancing the pharmacokinetic profiles of low-molecular-weight drugs and increasing their accumulation within tumor tissues through the EPR phenomenon. Human serum albumin dimers are two HSA molecules linked together and have gained attention in recent years as potential drug carriers for various therapeutic applications.
The increased uptake of albumin dimer-based drug delivery systems in solid tumors is facilitated by the pathophysiology of the tumor tissue. The accumulation of HSA dimers is attributed not only to transcytosis initiated by the binding of albumin to 60-kDa glycoprotein (gp60) and SPARC but also to the presence of these proteins in various types of tumor stroma. Therefore, the endogenous albumin transport system presents significant potential as an effective drug delivery approach for addressing resistant forms of cancer.
Ishima et al183 demonstrated the potential of poly-nitric oxide (NO)-conjugated human serum albumin (Poly-SNO-HSA) as a drug delivery system (DDS) capable of accumulating NO in tumors. However, they noted the need for improved stability of poly-SNO-HSA in circulation and emphasized the importance of evaluating its optimal molecular size for maximizing the EPR effect. This study employed two murine tumor models: one with Colon 26 (C26) cells, which originate from mouse colon cancer and exhibit a pronounced endogenous EPR effect, and the other with B16 cells, which are derived from mouse melanoma and demonstrate a comparatively smaller EPR effect. By dimerizing with albumin, they achieved a tenfold increase in antitumor activity, enhancing NO delivery in Colon 26 tumor-bearing mice. Their research focused on substantial accumulation of albumin in tumors, leveraging the S-nitrosated human serum albumin dimer (SNO-HSA dimer) to increase the EPR effect. They aimed to develop a DDS strategy utilizing the endogenous albumin transport (EAT) system to target tumor cells, underscoring its potential for future advancements. Furthermore, the same group investigated the effectiveness of PEGylated liposomal doxorubicin (Doxil), a typical example of a stealth liposome approved for medical usage.184 In this study, the antitumor efficacy of Doxil® was enhanced when Doxil® was combined with the S-nitrosated form of HSA dimer in the treatment of solid tumors in B16-bearing mice with poorly permeable vasculature, which corresponded with increased survival rates and diminished tumor sizes. To validate the tumor response, the doxorubicin levels in the tumor were quantified. The concentrations of doxorubicin in the tumor increased by more than threefold in the C26 (murine colon 26 adenocarcinoma) and sixfold in the B16 (murine melanoma) tumor-bearing models. These findings suggest that albumin dimers have the potential in enhancing the EPR effect in highly resistant cancer, likely due to their ability to utilize the natural albumin transport mechanism. Consequently, the endogenous albumin transport system appears to be a promising drug delivery strategy for combating refractory cancers.184 In 2017, Ryo Kinoshita et al studied an albumin dimer incorporating the chemotherapeutic agent doxorubicin (HSA-d-DOX) as a next-generation drug delivery system for tumor treatment, employing state-of-the-art nab (nanoparticle albumin-bound) technology.185 The results showed that HSA-dimers-DOX demonstrated greater cytotoxicity than HSA-monomers-DOX did in vitro in SUIzo Tumor-2 (SUIT2) cells. Furthermore, comparative analysis revealed that the HSA dimer significantly improved tumor migration and cellular uptake abilities compared with its counterpart, the HSA monomer, which was synthesized via conventional nab techniques. Notably, HSA-d-DOX efficiently delivered DOX to a human pancreatic tumor model (SUIT2) and effectively inhibited tumor growth without causing severe side effects. Consequently, the dimerization of HSA has emerged as a multifaceted strategy, facilitating the solubilization of previously insoluble drugs and serving as an active targeting carrier, which is particularly valuable in low vascular permeability or in the context of intractable pancreatic tumors.67 HSA-d, a dimer of HSA, has a greater tumor distribution than the HSA monomer does, making it a more effective drug carrier for antitumor therapy.186 Recent research suggests that dimeric albumin formulations can more efficiently target anticancer drugs to tumor tissues, such as chemically synthesized dimers created using a 1.6-bis(maleimido)hexane (BMH) spacer at the Cys-34 residue.187–189
Managing Pharmacokinetics
Throughout its 27-day lifespan, an average albumin molecule completes approximately 15,000 circulatory trips and around 15 journeys through extravascular spaces.24 The use of biomacromolecule-based albumin carriers has been shown to improve the pharmacokinetics of payloads and reduce systemic toxicity and immunogenicity. Albumin, with its long half-life, serves as an effective biomimetic drug delivery carrier, enhancing solubility, stability, and targeted delivery to tissues. Its natural affinity for tumor cells and EPR effect enable preferential accumulation of anticancer drugs in tumor tissues. The strategy for controlling the EPR may apply not only to liposome preparations, such as Doxil®, and albumin-binding drugs, such as Abraxane® but also to micelle preparations and antibodies, such as OPDIVO®, which has recently received prominent attention. Albumin-based systems also reduce the likelihood of adverse side effects by controlling the release and distribution of the drug. Moreover, the reduced immunogenicity associated with albumin-based drugs is particularly advantageous for avoiding unwanted immune responses, which can further complicate cancer treatment. Overall, managing the pharmacokinetics of albumin-based anticancer drugs is crucial for addressing the challenges posed by the complex microenvironment of refractory tumors. By leveraging the unique properties of albumin, these drug delivery systems enhance the efficacy and safety of anticancer therapies.
There are three approved drugs formulated as albumin-bound particles: Abraxane, Pazenir, and Fyarro (Table 4). Owing to the unique pharmacokinetics of these formulations, dosing adjustments may be required on the basis of patient-specific factors such as body weight, renal and hepatic function, and overall health. Regular monitoring for efficacy and side effects is essential, as these drugs have different toxicity profiles than nonalbumin-bound versions do. Additionally, potential drug interactions should be carefully considered, particularly with respect to medications that could influence the binding or metabolism of albumin-bound drugs.
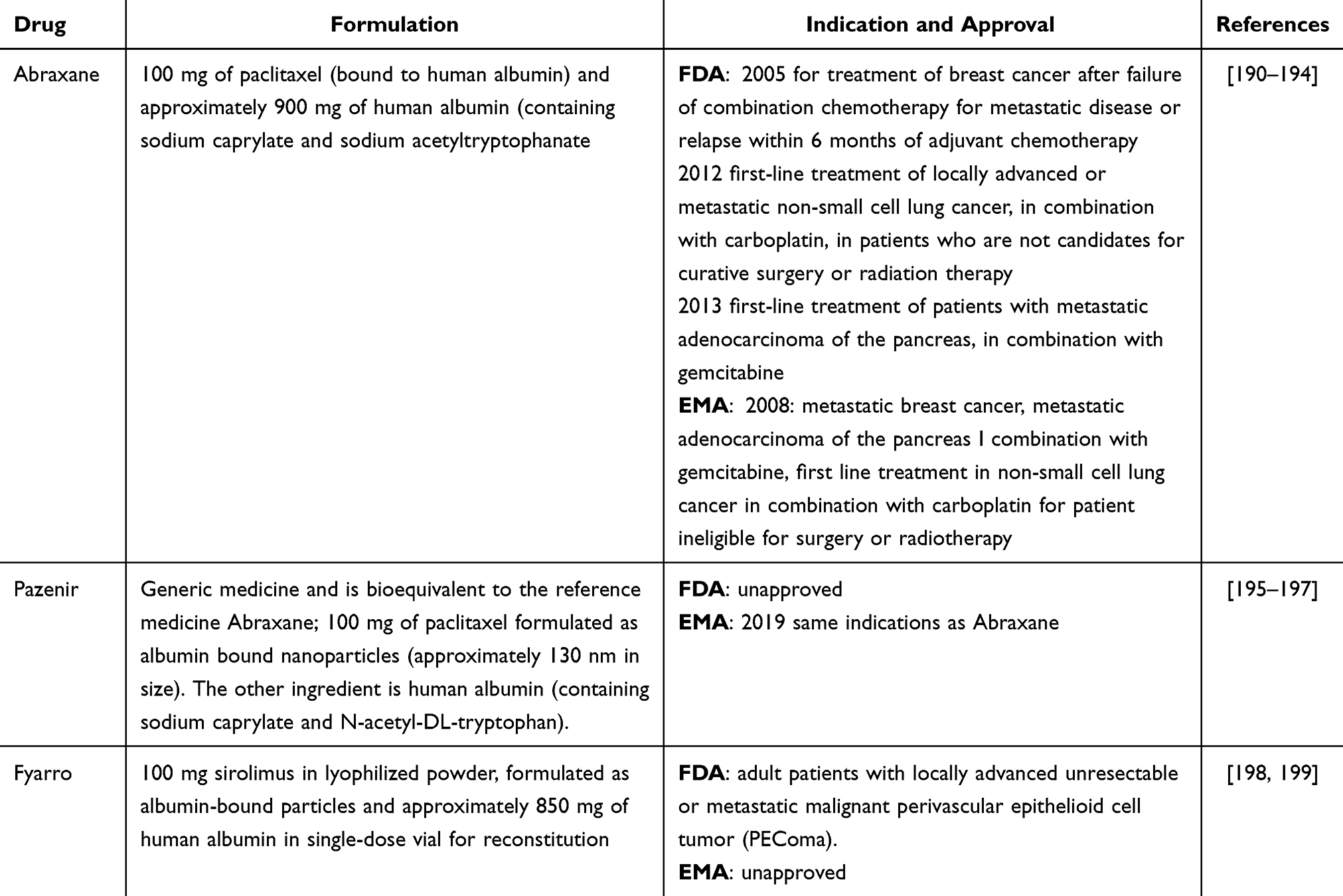 |
Table 4 Regulatory Approvals and Therapeutic Applications of Albumin-Based Anticancer Therapies for Improved Efficacy and Safety |
Abraxane and Pazenir
Both Abraxane and Pazenir contain human serum albumin‒paclitaxel nanoparticles of approximately 130 nm in size, where paclitaxel is present in a noncrystalline, amorphous state (Table 4). Upon intravenous administration, the nanoparticles dissociate rapidly into soluble, albumin-bound paclitaxel complexes approximately 10 nm in size. As described in Section 2, albumin is known to facilitate endothelial caveolar transcytosis of plasma components, and in vitro studies have shown that albumin in Abraxane improves the transport of paclitaxel across endothelial cells. It is hypothesized that this enhanced transendothelial caveolar transport is driven by gp-60, leading to increased paclitaxel accumulation in the tumor area due to the albumin-binding SPARC proteins.190
Pharmacokinetics of Abraxane
The pharmacokinetics of total paclitaxel have been determined in clinical studies. Following intravenous administration of Abraxane, paclitaxel concentrations in plasma declined in a biphasic manner, with the initial rapid decline representing distribution to the peripheral compartment and the slower second phase representing drug elimination (Table 5). The terminal half-life was approximately 27 hours. Drug exposure (area under the curve, AUC) was proportional to the dose over 80–375 mg/m2, and the pharmacokinetics of paclitaxel were independent of the duration of Abraxane administration. The mean total clearance was 15 L/hr/m2. Abraxane is characterized by extensive extravascular distribution and/or tissue binding of paclitaxel.
 |
Table 5 Comparison of Pharmacokinetics Between Chemotherapeutic-Loaded HSA Nanoparticles and Chemotherapeutic Agent Alone: Emphasizing the Role of HSA Nanoparticles |
Between 89% and 98% of the drug is bound to human serum proteins, and the presence of cimetidine, ranitidine, dexamethasone, or diphenhydramine does not affect the protein binding of paclitaxel.
Abraxane is metabolized primarily to 6α-hydroxypaclitaxel by CYP2C8 and to two minor metabolites, 3′-p-hydroxypaclitaxel and 6 α, 3′-p-dihydroxypaclitaxel, by CYP3A4. The pharmacokinetics of paclitaxel may be altered in vivo as a result of interactions with compounds that are substrates, inducers, or inhibitors of CYP2C8 and/or CYP3A4. The metabolism of paclitaxel is catalyzed by CYP2C8 and CYP3A4. In the absence of formal clinical studies on drug interactions, caution is advised when administering Abraxane alongside medications known to inhibit (eg, ketoconazole and other imidazole antifungals, erythromycin, fluoxetine, gemfibrozil, cimetidine, ritonavir, saquinavir, indinavir, and nelfinavir) or induce (eg, rifampicin, carbamazepine, phenytoin, efavirenz, and nevirapine) the enzymes CYP2C8 or CYP3A4. The mean values for cumulative urinary recovery of unchanged drugs (4%) indicated extensive nonrenal clearance. Less than 1% of the total administered dose was excreted in the urine as the metabolites 6 α-hydroxypaclitaxel and 3′-p-hydroxypaclitaxel.190,191
Pharmacokinetics of Pazenir
The plasma clearance of paclitaxel administered as Pazenir was 43% greater than that of solvent-based paclitaxel injections, with a 53% increase in volume of distribution. The mean terminal half-life ranges between 13 and 27 hours, and the average plasma clearance of paclitaxel is between 13 and 30 L/h/m² (Table 5). No evidence of paclitaxel accumulation was observed over multiple treatment cycles.195
After Pazenir is administered to patients with solid tumors, paclitaxel is evenly distributed between blood cells and plasma, with a high plasma protein binding rate of 94%. Population pharmacokinetic analysis estimated the total volume of distribution to be approximately 1741 L, suggesting significant extravascular distribution and/or extensive tissue binding of paclitaxel.195
Pazenir is metabolized primarily to 6α-hydroxypaclitaxel and to two minor metabolites, 3′-p-hydroxypaclitaxel and 6 α-3′-p-dihydroxypaclitaxel. The formation of these hydroxylated metabolites is catalyzed by CYP2C8, CYP3A4, and both the CYP2C8 and the CYP3A4 isoenzymes, respectively.195 The average cumulative urinary excretion of unchanged active substances accounted for 4% of the total administered dose, with less than 1% excreted as the metabolites 6α-hydroxypaclitaxel and 3′-p-hydroxypaclitaxel, indicating extensive nonrenal clearance. Pazenir is primarily eliminated through hepatic metabolism and biliary excretion.195
Caution is advised when administering paclitaxel alongside medications known to inhibit CYP2C8 or CYP3A4 (eg, ketoconazole and other imidazole antifungals, erythromycin, fluoxetine, gemfibrozil, clopidogrel, cimetidine, ritonavir, saquinavir, indinavir, and nelfinavir), as increased exposure to paclitaxel may increase its toxicity. Coadministration of paclitaxel with medicines known to induce CYP2C8 or CYP3A4 (eg, rifampicin, carbamazepine, phenytoin, efavirenz, nevirapine) is not recommended, as it may reduce paclitaxel exposure and compromise its efficacy, since paclitaxel clearance is driven primarily by CYP2C8- and CYP3A4-mediated metabolism, followed by biliary excretion.195
Fyarro
Fyarro is a nanoparticle albumin-bound formulation of sirolimus, also known as nab-sirolimus. The active ingredient, sirolimus, inhibits the mammalian target of rapamycin (mTOR) pathway, which is vital for cell growth and proliferation. By blocking this pathway, sirolimus helps slow or halt the growth of cancer cells. The protein binding ability of sirolimus is greater than 99%, and sirolimus primarily attaches to serum albumin in vitro. Additionally, sirolimus has a mean elimination half-life of approximately 59 hours, is metabolized by CYP3A4, and is primarily eliminated through feces (91%) and urine (2%) (Table 5). Fyarro’s active ingredient, sirolimus, is also a substrate for P-gp.198 CYP3A4 and/or P-gp inhibitors may increase sirolimus concentrations, increasing the risk of Fyarro-related adverse reactions, whereas CYP3A4 and/or P-gp inducers may lower sirolimus levels, potentially reducing Fyarro’s effectiveness. To avoid these effects, the concomitant use of Fyarro with strong CYP3A4 and/or P-gp inhibitors or inducers, as well as grapefruit or grapefruit juice (a strong CYP3A4 inhibitor), should be avoided. If Fyarro is used with a moderate or weak CYP3A4 inhibitor, a dosage reduction is recommended. Additionally, using moderate or weak CYP3A4 inducers with Fyarro may decrease sirolimus effectiveness.198,205,206
Conclusion
Albumin is a well-studied carrier in cancer therapy, serving as an alternative energy source for cancer cells that adapt to its uptake. Its physical structure can be modified without denaturation, for instance, through pH manipulation or lysine modification with NHS ester, enabling the design of novel drug delivery systems. Such modifications support a “wolf in sheep’s clothing” strategy for effective therapy.
The cellular uptake of HSA-based nanotherapeutics, though promising, requires further investigation. While gp60-mediated uptake and its interplay with SPARC enhance HSA accumulation in cancer cells, the underlying mechanisms remain unclear, as discussed in Section 2.2. A key challenge is the lack of detailed genetic and molecular information on gp60 and gp18/30 receptors, limiting therapeutic advancements targeting these pathways. Emerging albumin receptors, such as CD36 and transferrin, present additional targets for HSA modifications.
Overcoming multidrug resistance (MDR1) remains critical, particularly regarding resistance to Abraxane, linked to paclitaxel release from albumin. HSA-based nanoparticles, however, have demonstrated efficacy in MDR1-resistant cell lines. Strategies such as active targeting of overexpressed cancer cell surface proteins—including transferrin receptors, CD44, folic acid receptors, and specific antibodies—offer potential solutions. Dimeric HSA has shown enhanced cellular uptake and migratory properties, further advancing therapeutic opportunities.
Drugs like Abraxane, Pazenir, and Fyarro require careful pharmacokinetic optimization to balance efficacy and minimize adverse effects, although little is known about Pazenir’s effectiveness in drug-resistant cancers. HSA-based nanoparticles hold significant promise for overcoming resistance, but understanding uptake mechanisms in specific cancer types remains vital to developing multi-receptor-targeting delivery systems for improved outcomes Overall, we believe that HSA-based nanoparticles show potential for bypassing resistance, but a deeper understanding of uptake mechanisms in specific cancer types is crucial for designing delivery systems targeting multiple receptors for enhanced efficacy. Overall, we believe that HSA-based nanoparticles show potential for bypassing resistance, but a deeper understanding of uptake mechanisms in specific cancer types is crucial for designing delivery systems targeting multiple receptors for enhanced efficacy.
Abbreviations
(ABC) proteins, ATP-binding cassette; ABCB1, ABC-binding protein 1 (MDR1); Akt, Protein kinase B; BCRP (ABCG2), Breast Cancer Resistance Protein; CAV1, caveolin-1; CD36, cluster of differentiation 36,; CD44, cluster of differentiation 44; CSCs, cancer stem cells; CTX, Cabazitaxel; CYP2C8, Cytochrome P450 2C8; CYP3A4, Cytochrome P450 3A4; Db, diabodies; DDS, drug delivery system; dHSA, human serum albumin dimer; DTX, docetaxel; EGFR, epidermal growth factor receptor; EMA, European Medicines Agency; EPR, enhanced permeability and retention; FA, folic acid; FcRn, neonatal fragment crystallizable (Fc) receptor; FDA, US Food and Drug Administration; FR, Folate Receptor; gp18/30, glycoprotein 18/30; gp60, glycoprotein 60; HA, Hyaluronic acid; HCPT, hydroxycaptothecin; HER2, Receptor tyrosine-protein kinase erbB-2; HSA, human serum albumin; MAPK, mitogen-activated protein kinase; MDR1, multidrug resistance type 1; MRP (ABCC), multiple drug resistance protein; mTOR, mammalian target of rapamycin; MTX, Methotrexate; Nb, nanobodies; NF-κB, Nuclear factor kappa-light chain enhancer of activated B cells; NPs, nanoparticles; OVA, ovalbumin; PEG, polyethylene glycol; PgP, glycoprotein P (MDR1); PI3Ks, Phosphoinositide 3-kinase; rHSA, recombinant human serum albumin; scFv, single-chain variable fragment; SPARC, secreted protein acidic and rich in cysteine; TfR1, transferrin receptor 1, CD71; TMAB, trastuzumab; TME, tumor microenvironment; VEGF, Vascular endothelial growth factor.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This work was funded by Pomeranian Medical University in Szczecin, WFB-409/S/2023.
Disclosure
The authors declare that they have no competing interests.
References
1. Spada A, Emami J, Tuszynski JA, Lavasanifar A. The Uniqueness of Albumin as a Carrier in Nanodrug Delivery. Mol Pharm. 2021;18(5):1862–1894. doi:10.1021/acs.molpharmaceut.1c00046
2. Kamphorst JJ, Nofal M, Commisso C, et al. Human Pancreatic Cancer Tumors Are Nutrient Poor and Tumor Cells Actively Scavenge Extracellular Protein. Cancer Res. 2015;75(3):544–553. doi:10.1158/0008-5472.CAN-14-2211
3. Stehle G, Sinn H, Wunder A, et al. Plasma protein (albumin) catabolism by the tumor itself—implications for tumor metabolism and the genesis of cachexia. Crit rev oncol/hematol. 1997;26(2):77–100. doi:10.1016/S1040-8428(97)00015-2
4. Recouvreux MV, Commisso C. Macropinocytosis: a Metabolic Adaptation to Nutrient Stress in Cancer. Front Endocrinol. 2017;8:261. doi:10.3389/fendo.2017.00261
5. Curry S, Mandelkow H, Brick P, Franks N. Crystal structure of human serum albumin complexed with fatty acid reveals an asymmetric distribution of binding sites. Nat Struct mol Biol. 1998;5(9):827–835. doi:10.1038/1869
6. Gurachevsky A, Muravskaya E, Gurachevskaya T, Smirnova L, Muravsky V. Cancer-Associated Alteration in Fatty Acid Binding to Albumin Studied by Spin-Label Electron Spin Resonance. Cancer Invest. 2007;25(6):378–383. doi:10.1080/07357900701407947
7. Commisso C, Davidson SM, Soydaner-Azeloglu RG, et al. Macropinocytosis of protein is an amino acid supply route in Ras-transformed cells. Nature. 2013;497(7451):633–637. doi:10.1038/nature12138
8. Bagheri S, Saboury AA. Hypothesis of using albumin to improve drug efficacy in cancers accompanied by hypoalbuminemia. Xenobiotica. 2021;51(7):778–785. doi:10.1080/00498254.2021.1929557
9. Hoogenboezem EN, Duvall CL. Harnessing albumin as a carrier for cancer therapies. Adv. Drug Delivery Rev. 2018;130:73–89. doi:10.1016/j.addr.2018.07.011
10. Wilhelm S, Tavares AJ, Dai Q, et al. Analysis of nanoparticle delivery to tumours. Nat Rev Mater. 2016;1(5):16014. doi:10.1038/natrevmats.2016.14
11. Cho H, Jeon SI, Ahn C-H, Shim MK, Kim K. Emerging Albumin-Binding Anticancer Drugs for Tumor-Targeted Drug Delivery. Current Understandings and Clinical Translation Pharmaceutics. 2022;14:728.
12. Bukowski K, Kciuk M, Kontek R. Mechanisms of Multidrug Resistance in Cancer Chemotherapy. IJMS. 2020;21(9):3233. doi:10.3390/ijms21093233
13. Emran TB, Shahriar A, Mahmud AR, et al. Multidrug Resistance in Cancer: understanding Molecular Mechanisms, Immunoprevention and Therapeutic Approaches. Front Oncol. 2022;12:891652.
14. Juliano RL, Ling V. A surface glycoprotein modulating drug permeability in Chinese hamster ovary cell mutants. Biochimica et Biophysica Acta. 1976;455(1):152–162. doi:10.1016/0005-2736(76)90160-7
15. Lee TD, Lee OW, Brimacombe KR, et al. A High-Throughput Screen of a Library of Therapeutics Identifies Cytotoxic Substrates of P-glycoprotein. Mol Pharmacol. 2019;96(5):629–640. doi:10.1124/mol.119.115964
16. Hegedűs T, Őrfi L, Seprődi A, Váradi A, Sarkadi B, Kéri G. Interaction of tyrosine kinase inhibitors with the human multidrug transporter proteins, MDR1 and MRP1. Biochim Biophys Acta. 2002;1587(2–3):318–325. doi:10.1016/S0925-4439(02)00095-9
17. Karami E, Behdani M, Kazemi-Lomedasht F. Albumin nanoparticles as nanocarriers for drug delivery: focusing on antibody and nanobody delivery and albumin-based drugs. J Drug Delivery Sci Technol. 2020;55:101471. doi:10.1016/j.jddst.2019.101471
18. Hassanin I, Elzoghby A Albumin-based nanoparticles: a promising strategy to overcome cancer drug resistance. CDR. 2020. Available from: https://www.oaepublish.com/articles/cdr.2020.68.
19. Sleep D, Cameron J, Evans LR. Albumin as a versatile platform for drug half-life extension. Biochim Biophys Acta. 2013;1830(12):5526–5534. doi:10.1016/j.bbagen.2013.04.023
20. Asrorov AM, Mukhamedov N, Kayumov M, et al. Albumin is a reliable drug-delivering molecule: highlighting points in cancer therapy. Medicine in Drug Discovery. 2024;22:100186. doi:10.1016/j.medidd.2024.100186
21. Hutapea TPH, Madurani KA, Syahputra MY, et al. Albumin: source, preparation, determination, applications, and prospects. J Sci. 2023;8:100549.
22. H M, Azzazy E, Christenson RH. All About Albumin: biochemistry, Genetics, and Medical Applications. Clin. Chem. 1996;432:2014–2015.
23. Sleep D. Albumin and its application in drug delivery. Expert Opin Drug Delivery. 2015;12(5):793–812. doi:10.1517/17425247.2015.993313
24. Peters T. Serum Albumin. Advances in Protein Chemistry. 1985. 161–245. Available from: https://linkinghub.elsevier.com/retrieve/pii/S0065323308600650.
25. Carter DC, Ho JX. Structure of Serum Albumin. Adv Potein Chem. 1994:153–203. Available from.
26. Kim J, Hayton WL, Robinson JM, Anderson CL. Kinetics of FcRn-mediated recycling of IgG and albumin in human: pathophysiology and therapeutic implications using a simplified mechanism-based model. Clin Immunol. 2007;122(2):146–155. doi:10.1016/j.clim.2006.09.001
27. Anderson CL, Chaudhury C, Kim J, Bronson CL, Wani MA, Mohanty S. Perspective – fcRn transports albumin: relevance to immunology and medicine. Trends in Immunology. 2006;27(7):343–348. doi:10.1016/j.it.2006.05.004
28. Nicholson JP, Wolmarans MR, Park GR. The role of albumin in critical illness. Br J Anaesth. 2000;85(4):599–610. doi:10.1093/bja/85.4.599
29. Reynolds EC, Wong A. Effect of adsorbed protein on hydroxyapatite zeta potential and Streptococcus mutans adherence. Infect Immun. 1983;39(3):1285–1290. doi:10.1128/iai.39.3.1285-1290.1983
30. Adamczyk Z, Nattich-Rak M, Dąbkowska M, Kujda-Kruk M. Albumin adsorption at solid substrates: a quest for a unified approach. J Colloid Interface Sci. 2018;514:769–790. doi:10.1016/j.jcis.2017.11.083
31. Roche M, Rondeau P, Singh NR, Tarnus E, Bourdon E. The antioxidant properties of serum albumin. FEBS Lett. 2008;582(13):1783–1787. doi:10.1016/j.febslet.2008.04.057
32. Anraku M, Chuang VTG, Maruyama T, Otagiri M. Redox properties of serum albumin. Biochim Biophys Acta. 2013;1830(12):5465–5472. doi:10.1016/j.bbagen.2013.04.036
33. Plantier J-L, Duretz V, Devos V, Urbain R, Jorieux S. Comparison of antioxidant properties of different therapeutic albumin preparations. Biologicals. 2016;44(4):226–233. doi:10.1016/j.biologicals.2016.04.002
34. Fanali G, Di Masi A, Trezza V, Marino M, Fasano M, Ascenzi P. Human serum albumin: from bench to bedside. mol Aspect Med. 2012;33(3):209–290. doi:10.1016/j.mam.2011.12.002
35. Dąbkowska M, Adamczyk Z. Human Serum Albumin Monolayers on Mica: electrokinetic Characteristics. Langmuir. 2012;28(44):15663–15673. doi:10.1021/la3036677
36. Rabbani G, Ahn SN. Structure, enzymatic activities, glycation and therapeutic potential of human serum albumin: a natural cargo. Int J Biol Macromol. 2019;123:979–990. doi:10.1016/j.ijbiomac.2018.11.053
37. Sugio S, Kashima A, Mochizuki S, Noda M, Kobayashi K. Crystal structure of human serum albumin at 2.5 Å resolution. Protein Eng Des Sel. 1999;12(6):439–446. doi:10.1093/protein/12.6.439
38. Turell L, Botti H, Bonilla L, et al. HPLC separation of human serum albumin isoforms based on their isoelectric points. J Chromatogr B. 2014;944:144–151. doi:10.1016/j.jchromb.2013.11.019
39. Olsson MHM, Søndergaard CR, Rostkowski M, Jensen JH. PROPKA3: consistent Treatment of Internal and Surface Residues in Empirical pKa Predictions. J Chem Theory Comput. 2011;7(2):525–537. doi:10.1021/ct100578z
40. Peters T. All About Albumin: Biochemistry, Genetics, and Medical Applications. Academic Press; 1996.
41. Jachimska B, Wasilewska M, Adamczyk Z. Characterization of Globular Protein Solutions by Dynamic Light Scattering, Electrophoretic Mobility, and Viscosity Measurements. Langmuir. 2008;24(13):6866–6872. doi:10.1021/la800548p
42. Hawe A, Friess W. Stabilization of a hydrophobic recombinant cytokine by human serum albumin. J Pharmaceut Sci. 2007;96(11):2987–2999. doi:10.1002/jps.20909
43. Kujda M, Adamczyk Z, Ciesla M. Monolayers of the HSA dimer on polymeric microparticles-electrokinetic characteristics. Colloids Surf. B. 2016;148:229–237. doi:10.1016/j.colsurfb.2016.08.017
44. Maciążek-Jurczyk M, Pawliszyn J. The automatic use of capillary isoelectric focusing with whole column imaging detection for carbamazepine binding to human serum albumin. Journal of Pharmaceutical & Biomedical Analysis. 2016;127:9–17. doi:10.1016/j.jpba.2016.01.023
45. Genoud V, Castañon M, Lauricella AM, Quintana I. Characterization of N-homocysteinylated Albumin Adducts. Protein J. 2014;33(1):85–91. doi:10.1007/s10930-013-9540-z
46. Kohlheyer D, Eijkel JCT, Schlautmann S, Van Den Berg A, Schasfoort RBM. Microfluidic High-Resolution Free-Flow Isoelectric Focusing. Anal Chem. 2007;79(21):8190–8198. doi:10.1021/ac071419b
47. Joshi R, Jadhao M, Kumar H, Ghosh SK. Is the Sudlow site I of human serum albumin more generous to adopt prospective anti-cancer bioorganic compound than that of bovine: a combined spectroscopic and docking simulation approach. Bioorg Chem. 2017;75:332–346. doi:10.1016/j.bioorg.2017.10.013
48. Park J, Kim M-S, Park T, Kim YH, Shin DH. Crystal structure of pharmaceutical-grade human serum albumin. Int J Biol Macromol. 2021;166:221–228. doi:10.1016/j.ijbiomac.2020.10.152
49. Zhao. N-terminal PEGylation of human serum albumin and investigation of its pharmacokinetics and pulmonary microvascular retention. 2012. Available from: http://www.biosciencetrends.com/getabstract.php?id=533.
50. Fahrländer E, Schelhaas S, Jacobs AH, Langer K. PEGylated human serum albumin (HSA) nanoparticles: preparation, characterization and quantification of the PEGylation extent. Nanotechnology. 2015;26(14):145103. doi:10.1088/0957-4484/26/14/145103
51. Bourassa P, Hasni I, Tajmir-Riahi HA. Folic acid complexes with human and bovine serum albumins. Food Chem. 2011;129(3):1148–1155. doi:10.1016/j.foodchem.2011.05.094
52. Merlot AM, Kalinowski DS, Richardson DR. Unraveling the mysteries of serum albumin—more than just a serum protein. Front Physiol. 2014. 5. doi:10.3389/fphys.2014.00299/abstract
53. Ji Q, Zhu H, Qin Y, et al. GP60 and SPARC as albumin receptors: key targeted sites for the delivery of antitumor drugs. Front Pharmacol. 2024;15:1329636. doi:10.3389/fphar.2024.1329636
54. Raheel H, Ghaffari S, Khosraviani N, et al. CD36 mediates albumin transcytosis by dermal but not lung microvascular endothelial cells: role in fatty acid delivery. Am J Physiol Lung Cell mol Physiol. 2019;316:L740–50.
55. Bern M, Sand KMK, Nilsen J, Sandlie I, Andersen JT. The role of albumin receptors in regulation of albumin homeostasis: implications for drug delivery. J Control Release. 2015;211:144–162. doi:10.1016/j.jconrel.2015.06.006
56. Meng R, Hao S, Sun C, et al. Reverse-QTY code design of active human serum albumin self-assembled amphiphilic nanoparticles for effective anti-tumor drug doxorubicin release in mice. Proc Natl Acad Sci USA. 2023;120:e2220173120.
57. Dalton CM, Schlegel C, Hunter CJ. Caveolin-1: a Review of Intracellular Functions, Tissue-Specific Roles, and Epithelial Tight Junction Regulation. Biology. 2023;12(11):1402. doi:10.3390/biology12111402
58. Kaksonen M, Roux A. Mechanisms of clathrin-mediated endocytosis. Nat Rev mol Cell Biol. 2018;19(5):313–326. doi:10.1038/nrm.2017.132
59. Li -H-H, Li J, Wasserloos KJ, et al. Caveolae-Dependent and -Independent Uptake of Albumin in Cultured Rodent Pulmonary Endothelial Cells. PLoS One. 2013;8(11):e81903. doi:10.1371/journal.pone.0081903
60. Nakamura K, Yamanouchi K, Nishihara M. Secreted protein acidic and rich in cysteine internalization and its age-related alterations in skeletal muscle progenitor cells. Aging Cell. 2014;13(1):175–184. doi:10.1111/acel.12168
61. Jayashankar V, Edinger AL. Macropinocytosis confers resistance to therapies targeting cancer anabolism. Nat Commun. 2020;11(1):1121. doi:10.1038/s41467-020-14928-3
62. Soeters PB, Wolfe RR, Shenkin A. Hypoalbuminemia: pathogenesis and Clinical Significance. J Parenter Enteral Nutr. 2019;43(2):181–193. doi:10.1002/jpen.1451
63. Schnitzer JE, Oh P. Albondin-mediated capillary permeability to albumin. Differential role of receptors in endothelial transcytosis and endocytosis of native and modified albumins. J Biol Chem. 1994;269(8):6072–6082. doi:10.1016/S0021-9258(17)37571-3
64. Chatterjee M, Ben-Josef E, Robb R, et al. Caveolae-Mediated Endocytosis Is Critical for Albumin Cellular Uptake and Response to Albumin-Bound Chemotherapy. Cancer Res. 2017;77(21):5925–5937. doi:10.1158/0008-5472.CAN-17-0604
65. Hama M, Ishima Y, Chuang VTG, Ando H, Shimizu T, Ishida T. Evidence for Delivery of Abraxane via a Denatured-Albumin Transport System. ACS Appl Mater Interfaces. 2021;13(17):19736–19744. doi:10.1021/acsami.1c03065
66. Schultz AM, Copeland TD, Oroszlanz S. The envelope proteins of bovine leukemia virus: purification and sequence analysis. Virology. 1984;135(2):417–427. doi:10.1016/0042-6822(84)90197-1
67. Kinoshita R, Ishima Y, Chuang VTG, et al. Improved anticancer effects of albumin-bound paclitaxel nanoparticle via augmentation of EPR effect and albumin-protein interactions using S-nitrosated human serum albumin dimer. Biomaterials. 2017;140:162–169. doi:10.1016/j.biomaterials.2017.06.021
68. An JM, Moon H, Verwilst P, et al. Human Glioblastoma Visualization: triple Receptor-Targeting Fluorescent Complex of Dye, SIWV Tetra-Peptide, and Serum Albumin Protein. ACS Sens. 2021;6(6):2270–2280. doi:10.1021/acssensors.1c00320
69. Antohe F, Rădulescu L, Gafencu A, Gheţie V, Simionescu M. Expression of functionally active FcRn and the differentiated bidirectional transport of IgG in human placental endothelial cells. Hum Immunol. 2001;62(2):93–105. doi:10.1016/S0198-8859(00)00244-5
70. Li Z, Palaniyandi S, Zeng R, Tuo W, Roopenian DC, Zhu X. Transfer of IgG in the female genital tract by MHC class I-related neonatal Fc receptor (FcRn) confers protective immunity to vaginal infection. Proc Natl Acad Sci USA. 2011;108(11):4388–4393. doi:10.1073/pnas.1012861108
71. Zhu X, Meng G, Dickinson BL, et al. MHC Class I-Related Neonatal Fc Receptor for IgG Is Functionally Expressed in Monocytes, Intestinal Macrophages, and Dendritic Cells. J Immunol. 2001;166(5):3266–3276. doi:10.4049/jimmunol.166.5.3266
72. Powner MB, McKenzie JAG, Christianson GJ, Roopenian DC, Fruttiger M. Expression of Neonatal Fc Receptor in the Eye. Invest Ophthalmol Vis Sci. 2014;55(3):1607. doi:10.1167/iovs.13-12574
73. Schlachetzki F, Zhu C, Pardridge WM. Expression of the neonatal Fc receptor (FcRn) at the blood–brain barrier. Journal of Neurochemistry. 2002;81(1):203–206. doi:10.1046/j.1471-4159.2002.00840.x
74. Cianga P, Cianga C, Cozma L, Ward ES, Carasevici E. The MHC class I related Fc receptor, FcRn, is expressed in the epithelial cells of the human mammary gland. Hum Immunol. 2003;64(12):1152–1159. doi:10.1016/j.humimm.2003.08.025
75. Cauza K, Hinterhuber G, Dingelmaier-Hovorka R, et al. Expression of FcRn, the MHC Class I-Related Receptor for IgG, in Human Keratinocytes. J Invest Dermatol. 2005;124(1):132–139. doi:10.1111/j.0022-202X.2004.23542.x
76. Kim J, Bronson CL, Hayton WL, et al. Albumin turnover: fcRn-mediated recycling saves as much albumin from degradation as the liver produces. Am J Physiol Gastrointest Liver Physiol. 2006;290(2):G352–360. doi:10.1152/ajpgi.00286.2005
77. Chaudhury C, Mehnaz S, Robinson JM, et al. The Major Histocompatibility Complex–related Fc Receptor for IgG (FcRn) Binds Albumin and Prolongs Its Lifespan. J Exp Med. 2003;197(3):315–322. doi:10.1084/jem.20021829
78. Ward ES, Ober RJ. Commentary: “There’s been a Flaw in Our Thinking. Front Immunol”. 2015;6. Available from: http://journal.frontiersin.org/Article/10.3389/fimmu.2015.00351/abstract.
79. Boedtkjer E, Pedersen SF. The Acidic Tumor Microenvironment as a Driver of Cancer. Annu Rev Physiol. 2020;82(1):103–126. doi:10.1146/annurev-physiol-021119-034627
80. Swiercz R, Mo M, Khare P, Schneider Z, Ober RJ, Ward ES. Loss of expression of the recycling receptor, FcRn, promotes tumor cell growth by increasing albumin consumption. Oncotarget. 2017;8(2):3528–3541. doi:10.18632/oncotarget.13869
81. Dalloneau E, Baroukh N, Mavridis K, et al. Downregulation of the neonatal Fc receptor expression in non-small cell lung cancer tissue is associated with a poor prognosis. Oncotarget. 2016;7(34):54415–54429. doi:10.18632/oncotarget.10074
82. Larsen MT, Mandrup OA, Schelde KK, et al. FcRn overexpression in human cancer drives albumin recycling and cell growth; a mechanistic basis for exploitation in targeted albumin-drug designs. J Control Release. 2020;322:53–63. doi:10.1016/j.jconrel.2020.03.004
83. Gradishar WJ. Albumin-bound paclitaxel: a next-generation taxane. Expert Opinion on Pharmacotherapy. 2006;7(8):1041–1053. doi:10.1517/14656566.7.8.1041
84. Botti G, Scognamiglio G, Marra L, et al. SPARC/osteonectin is involved in metastatic process to the lung during melanoma progression. Virchows Arch. 2014;465(3):331–338. doi:10.1007/s00428-014-1616-4
85. Lindner JL, Loibl S, Denkert C, et al. Expression of secreted protein acidic and rich in cysteine (SPARC) in breast cancer and response to neoadjuvant chemotherapy. Ann Oncol. 2015;26(1):95–100. doi:10.1093/annonc/mdu487
86. Jendraschak E, Helene Sage E. Regulation of angiogenesis by SPARC and angiostatin: implications for tumor cell biology. Semi Cancer Biol. 1996;7(3):139–146. doi:10.1006/scbi.1996.0019
87. Volmer MW, Radacz Y, Hahn SA, et al. Tumor suppressor Smad4 mediates downregulation of the anti‐adhesive invasion‐promoting matricellular protein SPARC: landscaping activity of Smad4 as revealed by a “secretome” analysis. Proteomics. 2004;4(5):1324–1334. doi:10.1002/pmic.200300703
88. Tremble PM, Lane TF, Sage EH, Werb Z. SPARC, a secreted protein associated with morphogenesis and tissue remodeling, induces expression of metalloproteinases in fibroblasts through a novel extracellular matrix-dependent pathway. J Cell Biol. 1993;121(6):1433–1444. doi:10.1083/jcb.121.6.1433
89. Minn AJ, Gupta GP, Siegel PM, et al. Genes that mediate breast cancer metastasis to lung. Nature. 2005;436(7050):518–524. doi:10.1038/nature03799
90. Shi Q, Bao S, Maxwell JA, et al. Secreted Protein Acidic, Rich in Cysteine (SPARC), Mediates Cellular Survival of Gliomas through AKT Activation. J Biol Chem. 2004;279(50):52200–52209. doi:10.1074/jbc.M409630200
91. Kratz F. A clinical update of using albumin as a drug vehicle — a commentary. J Control Release. 2014;190:331–336. doi:10.1016/j.jconrel.2014.03.013
92. Nie J, Chang B, Traktuev DO, et al. IFATS Collection: combinatorial Peptides Identify α5β1 Integrin as a Receptor for the Matricellular Protein SPARC on Adipose Stromal Cells. Stem Cells. 2008;26(10):2735–2745. doi:10.1634/stemcells.2008-0212
93. Sanità G, Armanetti P, Silvestri B, et al. Albumin-Modified Melanin-Silica Hybrid Nanoparticles Target Breast Cancer Cells via a SPARC-Dependent Mechanism. Front Bioeng Biotechnol. 2020;8:765. doi:10.3389/fbioe.2020.00765
94. Kzhyshkowska J, Workman G, Cardó-Vila M, et al. Novel Function of Alternatively Activated Macrophages: stabilin-1-Mediated Clearance of SPARC. J Immunol. 2006;176(10):5825–5832. doi:10.4049/jimmunol.176.10.5825
95. Hoang B, Ernsting MJ, Roy A, Murakami M, Undzys E, Li S-D. Docetaxel-carboxymethylcellulose nanoparticles target cells via a SPARC and albumin dependent mechanism. Biomaterials. 2015;59:66–76. doi:10.1016/j.biomaterials.2015.04.032
96. Schnitzer JE, Sung A, Horvat R, Bravo J. Preferential interaction of albumin-binding proteins, gp30 and gp18, with conformationally modified albumins. Presence in many cells and tissues with a possible role in catabolism. J Biol Chem. 1992;267(34):24544–24553. doi:10.1016/S0021-9258(18)35799-5
97. Ghinea N, Fixman A, Alexandru D, et al. Identification of albumin-binding proteins in capillary endothelial cells. J Cell Biol. 1988;107(1):231–239. doi:10.1083/jcb.107.1.231
98. Schnitzer JE, Bravo J. High affinity binding, endocytosis, and degradation of conformationally modified albumins. Potential role of gp30 and gp18 as novel scavenger receptors. J Biol Chem. 1993;268(10):7562–7570. doi:10.1016/S0021-9258(18)53212-9
99. Shevtsova A, Gordiienko I, Tkachenko V, Ushakova G. Ischemia-Modified Albumin: origins and Clinical Implications. Dis. Markers. 2021;2021:1–18. doi:10.1155/2021/9945424
100. Wang J, Li Y. CD36 tango in cancer: signaling pathways and functions. Theranostics. 2019;9(17):4893–4908. doi:10.7150/thno.36037
101. Feng WW, Zuppe HT, Kurokawa M. The Role of CD36 in Cancer Progression and Its Value as a Therapeutic Target. Cells. 2023;12(12):1605. doi:10.3390/cells12121605
102. Ishima Y, Maruyama T, Otagiri M, Chuang VTG, Ishida T. The New Delivery Strategy of Albumin Carrier Utilizing the Interaction with Albumin Receptors. Chem Pharm Bull. 2022;70(5):330–333. doi:10.1248/cpb.c21-01024
103. Ryschich E, Huszty G, Knaebel HP, Hartel M, Büchler MW, Schmidt J. Transferrin receptor is a marker of malignant phenotype in human pancreatic cancer and in neuroendocrine carcinoma of the pancreas. Eur J Cancer. 2004;40(9):1418–1422. doi:10.1016/j.ejca.2004.01.036
104. Carlevaro MF, Albini A, Ribatti D, et al. Transferrin Promotes Endothelial Cell Migration and Invasion: implication in Cartilage Neovascularization. J Cell Biol. 1997;136(6):1375–1384. doi:10.1083/jcb.136.6.1375
105. Prutki M, Poljak-Blazi M, Jakopovic M, Tomas D, Stipancic I, Zarkovic N. Altered iron metabolism, transferrin receptor 1 and ferritin in patients with colon cancer. Cancer Lett. 2006;238(2):188–196. doi:10.1016/j.canlet.2005.07.001
106. Li C, Zhou L, Yin X. Pathophysiological aspects of transferrin-A potential nano-based drug delivery signaling molecule in therapeutic target for varied diseases. Front Pharmacol. 2024;15:1342181. doi:10.3389/fphar.2024.1342181
107. Daniels-Wells TR, Penichet ML. Transferrin receptor 1: a target for antibody-mediated cancer therapy. Immunotherapy. 2016;8(9):991–994. doi:10.2217/imt-2016-0050
108. Jennifer B, Berg V, Modak M, et al. Transferrin receptor 1 is a cellular receptor for human heme-albumin. Commun Biol. 2020;3(1):621. doi:10.1038/s42003-020-01294-5
109. Grinberg LN, O’Brien PJ, Hrkal Z. The effects of heme-binding proteins on the peroxidative and catalatic activities of hemin. Free Radic Biol Med. 1999;27(1–2):214–219. doi:10.1016/S0891-5849(99)00082-9
110. Morgan WT, Heng Liem H, Sutor RP, Muller-Eberhard U. Transfer of heme from heme-albumin to hemopexin. Biochim Biophys Acta. 1976;444(2):435–445. doi:10.1016/0304-4165(76)90387-1
111. Pasternack RF, Gibbs EJ, Hoeflin E, et al. Hemin binding to serum proteins and the catalysis of interprotein transfer. Biochemistry. 1983;22(8):1753–1758. doi:10.1021/bi00277a002
112. De Simone G, Varricchio R, Ruberto TF, Di Masi A, Ascenzi P. Heme Scavenging and Delivery: the Role of Human Serum Albumin. Biomolecules. 2023;13(3):575. doi:10.3390/biom13030575
113. Bae S, Ma K, Kim TH, et al. Doxorubicin-loaded human serum albumin nanoparticles surface-modified with TNF-related apoptosis-inducing ligand and transferrin for targeting multiple tumor types. Biomaterials. 2012;33(5):1536–1546. doi:10.1016/j.biomaterials.2011.10.050
114. Gardner ER, Dahut WL, Scripture CD, et al. Randomized Crossover Pharmacokinetic Study of Solvent-Based Paclitaxel and nab -Paclitaxel. Clin Cancer Res. 2008;14(13):4200–4205. doi:10.1158/1078-0432.CCR-07-4592
115. Emerich DF, Thanos CG. The pinpoint promise of nanoparticle-based drug delivery and molecular diagnosis. Biomol. Eng. 2006;23(4):171–184. doi:10.1016/j.bioeng.2006.05.026
116. Ibrahim NK, Samuels B, Page R, et al. Multicenter Phase II trial of ABI-007, an albumin-bound paclitaxel, in women with metastatic breast cancer. J Clin Oncol. 2005;23(25):6019–6026. doi:10.1200/JCO.2005.11.013
117. Dumontet C, Duran GE, Steger KA, Beketic-Oreskovic L, Sikic BI. Resistance mechanisms in human sarcoma mutants derived by single-step exposure to paclitaxel (Taxol). Cancer Res. 1996;56(5):1091–1097.
118. Blum JL, Savin MA, Edelman G, et al. Phase II Study of Weekly Albumin-Bound Paclitaxel for Patients with Metastatic Breast Cancer Heavily Pretreated with Taxanes. Clin Breast Cancer. 2007;7(11):850–856. doi:10.3816/CBC.2007.n.049
119. Zhao M, Lei C, Yang Y, et al. Abraxane, the Nanoparticle Formulation of Paclitaxel Can Induce Drug Resistance by Up-Regulation of P-gp. PLoS One. 2015;10(7):e0131429. doi:10.1371/journal.pone.0131429
120. Zhao M, Li H, Bu X, Lei C, Fang Q, Hu Z. Quantitative Proteomic Analysis of Cellular Resistance to the Nanoparticle Abraxane. ACS Nano. 2015;9(10):10099–10112. doi:10.1021/acsnano.5b03677
121. Vallo S, Köpp R, Michaelis M, et al. Resistance to nanoparticle albumin-bound paclitaxel is mediated by ABCB1 in urothelial cancer cells. Oncol Lett. 2017;13(6):4085–4092. doi:10.3892/ol.2017.5986
122. Kong L, Du J, Gu J, et al. Gemcitabine-Loaded Albumin Nanoparticle Exerts An Antitumor Effect on Gemcitabine-Resistant Pancreatic Cancer Cells Induced by MDR1 and MRP1 Overexpression in Vitro. Front Surg. 2022;9:890412. doi:10.3389/fsurg.2022.890412
123. Onafuye H, Pieper S, Mulac Jr D, et al. Doxorubicin-loaded human serum albumin nanoparticles overcome transporter-mediated drug resistance in drug-adapted cancer cells. Beilstein J Nanotechnol. 2019;10:1707–1715. doi:10.3762/bjnano.10.166
124. Fernandes Q, Therachiyil L, Khan AQ, et al. Shrinking the battlefield in cancer therapy: nanotechnology against cancer stem cells. Eur J Pharm Sci. 2023;191:106586. doi:10.1016/j.ejps.2023.106586
125. Yuan H, Guo H, Luan X, et al. Albumin Nanoparticle of Paclitaxel (Abraxane) Decreases while Taxol Increases Breast Cancer Stem Cells in Treatment of Triple Negative Breast Cancer. Mol Pharm. 2020;17(7):2275–2286. doi:10.1021/acs.molpharmaceut.9b01221
126. Bossennec M, Di Roio A, Caux C, Caux C. MDR1 in immunity: friend or foe? OncoImmunology. 2018;7:e1499388.
127. Lai J-I, Tseng Y-J, Chen M-H, Huang C-YF, Chang PM-H. Clinical Perspective of FDA Approved Drugs With P-Glycoprotein Inhibition Activities for Potential Cancer Therapeutics. Front Oncol. 2020;10:561936. doi:10.3389/fonc.2020.561936
128. Yergeri M, Kapse-Mistry S, Srivastava R, Govender T. Nanodrug delivery in reversing multidrug resistance in cancer cells. Front Pharmacol. 2014. 5. doi:10.3389/fphar.2014.00159.
129. Yang X, Wang Y, Chen S, Zhang S, Cui C. Cetuximab-Modified Human Serum Albumin Nanoparticles Co-Loaded with Doxorubicin and MDR1 siRNA for the Treatment of Drug-Resistant Breast Tumors. IJN. 2021;16:7051–7069. doi:10.2147/IJN.S332830
130. Liu X, Zhao T, Xu Y, et al. Co-administration of paclitaxel and 2-methoxyestradiol using folate-conjugated human serum albumin nanoparticles for improving drug resistance and antitumor efficacy. Pharmaceutical Development Technol. 2021;26(1):1–10. doi:10.1080/10837450.2020.1829640
131. Kodan A, Futamata R, Kimura Y, et al. ABCB1/MDR1/P‐gp employs an ATP‐dependent twist‐and‐squeeze mechanism to export hydrophobic drugs. FEBS Lett. 2021;595(6):707–716. doi:10.1002/1873-3468.14018
132. Kurimchak AM, Herrera-Montávez C, Montserrat-Sangrà S, et al. The drug efflux pump MDR1 promotes intrinsic and acquired resistance to PROTACs in cancer cells. Sci Signal. 2022;15:eabn2707.
133. Hodaei D, Baradaran B, Valizadeh H, Zakeri-Milani P. Effects of polyethylene glycols on intestinal efflux pump expression and activity in Caco-2 cells. Braz J Pharm Sci. 2015;51(3):745–753. doi:10.1590/S1984-82502015000300026
134. Li M, Si L, Pan H, et al. Excipients enhance intestinal absorption of ganciclovir by P-gp inhibition: assessed in vitro by everted gut sac and in situ by improved intestinal perfusion. Int J Pharm. 2011;403(1–2):37–45. doi:10.1016/j.ijpharm.2010.10.017
135. Johnson BM, Charman WN, Porter CJH. An in vitro examination of the impact of polyethylene glycol 400, pluronic P85, and vitamin E d-a-tocopheryl polyethylene glycol 1000 succinate on P-glycoprotein efflux and enterocyte-based metabolism in excised rat intestine. AAPS J. 2002;4(4):193–205. doi:10.1208/ps040440
136. Shen Q, Lin Y, Handa T, et al. Modulation of intestinal P-glycoprotein function by polyethylene glycols and their derivatives by in vitro transport and in situ absorption studies. Int J Pharm. 2006;313(1–2):49–56. doi:10.1016/j.ijpharm.2006.01.020
137. Khot VM, Salunkhe AB, Pricl S, Bauer J, Thorat ND, Townley H. Nanomedicine-driven molecular targeting, drug delivery, and therapeutic approaches to cancer chemoresistance. Drug Discovery Today. 2021;26(3):724–739. doi:10.1016/j.drudis.2020.12.016
138. Fan J, Gilmartin K, Octaviano S, Villar F, Remache B, Regan J. Using Human Serum Albumin Binding Affinities as a Proactive Strategy to Affect the Pharmacodynamics and Pharmacokinetics of Preclinical Drug Candidates. ACS Pharmacol Transl Sci. 2022;5(9):803–810. doi:10.1021/acsptsci.2c00115
139. Bar-Zeev M, Livney YD, Assaraf YG. Targeted nanomedicine for cancer therapeutics: towards precision medicine overcoming drug resistance. Drug Resist Updates. 2017;31:15–30. doi:10.1016/j.drup.2017.05.002
140. Shen Y, Li W. HA/HSA co-modified erlotinib–albumin nanoparticles for lung cancer treatment. <![CDATA[Drug Design, Development and Therapy]]>. 2018;12:2285–2292. doi:10.2147/DDDT.S169734
141. Brindisi M, Curcio M, Frattaruolo L, et al. CD44-targeted nanoparticles with GSH-responsive activity as powerful therapeutic agents against breast cancer. Int J Biol Macromol. 2022;221:1491–1503. doi:10.1016/j.ijbiomac.2022.09.157
142. Li Y, Shi S, Ming Y, et al. Specific cancer stem cell-therapy by albumin nanoparticles functionalized with CD44-mediated targeting. J Nanobiotechnol. 2018;16(1):99. doi:10.1186/s12951-018-0424-4
143. Sun Y, Zhao Y, Teng S, et al. Folic acid receptor-targeted human serum albumin nanoparticle formulation of cabazitaxel for tumor therapy. IJN. 2018;14:135–148. doi:10.2147/IJN.S181296
144. Ahlawat P, Phutela K, Bal A, Singh N, Sharma S. Therapeutic potential of human serum albumin nanoparticles encapsulated actinonin in murine model of lung adenocarcinoma. Drug Delivery. 2022;29(1):2403–2413. doi:10.1080/10717544.2022.2067600
145. Wang W, Liang H, Sun B, et al. Pharmacokinetics and Tissue Distribution of Folate-Decorated Human Serum Albumin Loaded With Nano-Hydroxycamptothecin for Tumor Targeting. J Pharmaceut Sci. 2016;105(6):1874–1880. doi:10.1016/j.xphs.2016.03.016
146. Hiremath CG, Kariduraganavar MY, Hiremath MB. Synergistic delivery of 5-fluorouracil and curcumin using human serum albumin-coated iron oxide nanoparticles by folic acid targeting. Prog Biomater. 2018;7(4):297–306. doi:10.1007/s40204-018-0104-3
147. Encinas-Basurto D, Ibarra J, Juarez J, et al. Hybrid folic acid-conjugated gold nanorods-loaded human serum albumin nanoparticles for simultaneous photothermal and chemotherapeutic therapy. Mater Sci Eng C. 2018;91:669–678. doi:10.1016/j.msec.2018.06.002
148. Taheri A, Dinarvand R, Atyabi F, Ghahremani MH, Ostad SN. Trastuzumab decorated methotrexate–human serum albumin conjugated nanoparticles for targeted delivery to HER2 positive tumor cells. Eur J Pharm Sci. 2012;47(2):331–340. doi:10.1016/j.ejps.2012.06.016
149. Yewale C, Baradia D, Patil S, et al. Docetaxel loaded immunonanoparticles delivery in EGFR overexpressed breast carcinoma cells. J Drug Delivery Sci Technol. 2018;45:334–345. doi:10.1016/j.jddst.2018.03.027
150. He S, Zhao C, Tao H, et al. A recombinant scFv antibody-based fusion protein that targets EGFR associated with IMPDH2 downregulation and its drug conjugate show therapeutic efficacy against esophageal cancer. Drug Delivery. 2022;29(1):1243–1256. doi:10.1080/10717544.2022.2063454
151. Xian Z, Ma L, Zhu M, et al. Blocking the PD-1-PD-L1 axis by a novel PD-1 specific nanobody expressed in yeast as a potential therapeutic for immunotherapy. Biochem. Biophys. Res. Commun. 2019;519(2):267–273. doi:10.1016/j.bbrc.2019.08.160
152. Zhou J, Rossi J. Aptamers as targeted therapeutics: current potential and challenges. Nat Rev Drug Discov. 2017;16(3):181–202. doi:10.1038/nrd.2016.199
153. Zhang H, Yu S, Ni S, et al. A bimolecular modification strategy for developing long-lasting bone anabolic aptamer. mol Ther Nucleic Acids. 2023;34:102073. doi:10.1016/j.omtn.2023.102073
154. Venkatesan S, Chanda K, Balamurali MM. Recent Advancements of Aptamers in Cancer Therapy. ACS Omega. 2023;8(36):32231–32243. doi:10.1021/acsomega.3c04345
155. Sanati M, Afshari AR, Ahmadi SS, Kesharwani P, Sahebkar A. Aptamers against cancer drug resistance: small fighters switching tactics in the face of defeat. Biochim Biophys Acta. 2023;1869(6):166720. doi:10.1016/j.bbadis.2023.166720
156. Shigdar S, Schrand B, Giangrande PH, De Franciscis V. Aptamers: cutting edge of cancer therapies. Mol Ther. 2021;29(8):2396–2411. doi:10.1016/j.ymthe.2021.06.010
157. An Y, Li X, Yao F, Duan J, Yang X-D. Novel Complex of PD-L1 Aptamer and Albumin Enhances Antitumor Efficacy In Vivo. Molecules. 2022;27(5):1482. doi:10.3390/molecules27051482
158. Lai X, Yao F, An Y, Li X, Yang X-D. Novel Nanotherapeutics for Cancer Immunotherapy by PD-L1-Aptamer-Functionalized and Fexofenadine-Loaded Albumin Nanoparticles. Molecules. 2023;28(6):2556. doi:10.3390/molecules28062556
159. Yao F, An Y, Lai X, Li X, Yu Z, Yang X-D. Novel nanotherapeutics for cancer immunotherapy by CTLA-4 aptamer-functionalized albumin nanoparticle loaded with antihistamine. J Cancer Res Clin Oncol. 2023;149(10):7515–7527. doi:10.1007/s00432-023-04698-y
160. Laurent TC, Fraser JRE. Hyaluronan. FASEB j. 1992;6(7):2397–2404. doi:10.1096/fasebj.6.7.1563592
161. Michalczyk M, Humeniuk E, Adamczuk G, Korga-Plewko A. Hyaluronic Acid as a Modern Approach in Anticancer Therapy-Review. IJMS. 2022;24(1):103. doi:10.3390/ijms24010103
162. Ludwig N, Szczepanski MJ, Gluszko A, et al. CD44(+) tumor cells promote early angiogenesis in head and neck squamous cell carcinoma. Cancer Lett. 2019;467:85–95. doi:10.1016/j.canlet.2019.10.010
163. Senbanjo LT, Chellaiah MA. CD44: a Multifunctional Cell Surface Adhesion Receptor Is a Regulator of Progression and Metastasis of Cancer Cells. Front Cell Dev Biol. 2017. 5. doi:10.3389/fcell.2017.00018.
164. Spadea A, Rios De La Rosa JM, Tirella A, et al. Evaluating the Efficiency of Hyaluronic Acid for Tumor Targeting via CD44. Mol Pharm. 2019;16(6):2481–2493. doi:10.1021/acs.molpharmaceut.9b00083
165. Yang C, He Y, Zhang H, et al. Selective killing of breast cancer cells expressing activated CD44 using CD44 ligand-coated nanoparticles in vitro and in vivo. Oncotarget. 2015;6(17):15283–15296. doi:10.18632/oncotarget.3681
166. Dalla Pozza E, Lerda C, Costanzo C, et al. Targeting gemcitabine containing liposomes to CD44 expressing pancreatic adenocarcinoma cells causes an increase in the antitumoral activity. Biochimica et Biophysica Acta. 2013;1828(5):1396–1404. doi:10.1016/j.bbamem.2013.01.020
167. Ifergan I, Assaraf YG. Chapter 4 Molecular Mechanisms of Adaptation to Folate Deficiency. In: Vitamins & Hormones. Elsevier; 2008: 99–143. Available from: https://linkinghub.elsevier.com/retrieve/pii/S0083672908004044.
168. Zhao R, Matherly LH, Goldman ID. Membrane transporters and folate homeostasis: intestinal absorption and transport into systemic compartments and tissues. Expert Rev Mol Med. 2009;11:e4.
169. Holm J, Hansen SI. Characterization of soluble folate receptors (folate binding proteins) in humans. Biological roles and clinical potentials in infection and malignancy. Biochimica Et Biophysica Acta Proteins and Proteomics. 2020;1868(10):140466. doi:10.1016/j.bbapap.2020.140466
170. Assaraf YG. Molecular basis of antifolate resistance. Cancer Metastasis Rev. 2007;26(1):153–181. doi:10.1007/s10555-007-9049-z
171. Parker N, Turk MJ, Westrick E, Lewis JD, Low PS, Leamon CP. Folate receptor expression in carcinomas and normal tissues determined by a quantitative radioligand binding assay. Analytical Biochemistry. 2005;338(2):284–293. doi:10.1016/j.ab.2004.12.026
172. Ross JF, Chaudhuri PK, Ratnam M. Differential regulation of folate receptor isoforms in normal and malignant tissues in vivo and in established cell lines. Physiologic and Clinical Implications Cancer. 1994;73:2432–2443.
173. Ulbrich K, Michaelis M, Rothweiler F, et al. Interaction of folate-conjugated human serum albumin (HSA) nanoparticles with tumour cells. Int J Pharm. 2011;406(1–2):128–134. doi:10.1016/j.ijpharm.2010.12.023
174. Basu S, Chaudhuri P, Sengupta S. Targeting oncogenic signaling pathways by exploiting nanotechnology. Cell Cycle. 2009;8(21):3480–3487. doi:10.4161/cc.8.21.9851
175. Shang L, Nienhaus K, Nienhaus GU. Engineered nanoparticles interacting with cells: size matters. J Nanobiotechnol. 2014;12(1):5. doi:10.1186/1477-3155-12-5
176. Suen W-L-L, Chau Y. Size-dependent internalisation of folate-decorated nanoparticles via the pathways of clathrin and caveolae-mediated endocytosis in ARPE-19 cells. J Pharm Pharmacol. 2014;66(4):564–573. doi:10.1111/jphp.12134
177. Sousa De Almeida M, Susnik E, Drasler B, Taladriz-Blanco P, Petri-Fink A, Rothen-Rutishauser B. Understanding nanoparticle endocytosis to improve targeting strategies in nanomedicine. Chem Soc Rev. 2021;50(9):5397–5434. doi:10.1039/d0cs01127d
178. Wang B, Galliford CV, Low PS. Guiding principles in the design of ligand-targeted nanomedicines. Nanomedicine. 2014;9(2):313–330. doi:10.2217/nnm.13.175
179. Altintas I, Kok RJ, Schiffelers RM. Targeting epidermal growth factor receptor in tumors: from conventional monoclonal antibodies via heavy chain-only antibodies to nanobodies. Eur J Pharm Sci. 2012;45(4):399–407. doi:10.1016/j.ejps.2011.10.015
180. Jin B, Odongo S, Radwanska M, Magez S. Nanobodies: a Review of Generation, Diagnostics and Therapeutics. IJMS. 2023;24(6):5994. doi:10.3390/ijms24065994
181. Wang X, Sheng W, Wang Y, et al. A Macropinocytosis-Intensifying Albumin Domain-Based scFv Antibody and Its Conjugate Directed against K-Ras Mutant Pancreatic Cancer. Mol Pharm. 2018;15(6):2403–2412. doi:10.1021/acs.molpharmaceut.8b00234
182. Du Y, Shang B, Sheng W, et al. A recombinantly tailored β-defensin that displays intensive macropinocytosis-mediated uptake exerting potent efficacy against K-Ras mutant pancreatic cancer. Oncotarget. 2016;7(36):58418–58434. doi:10.18632/oncotarget.11170
183. Ishima Y, Fang J, Kragh-Hansen U, et al. Tuning of Poly-S-Nitrosated Human Serum Albumin as Superior Antitumor Nanomedicine. J Pharmaceut Sci. 2014;103(7):2184–2188. doi:10.1002/jps.24020
184. Kinoshita R, Ishima Y, Ikeda M, et al. S-Nitrosated human serum albumin dimer as novel nano-EPR enhancer applied to macromolecular anti-tumor drugs such as micelles and liposomes. J Control Release. 2015;217:1–9. doi:10.1016/j.jconrel.2015.08.036
185. Kinoshita R, Ishima Y, Chuang VTG, et al. The Therapeutic Effect of Human Serum Albumin Dimer-Doxorubicin Complex against Human Pancreatic Tumors. Pharmaceutics. 2021;13(8):1209. doi:10.3390/pharmaceutics13081209
186. Ishima Y, Maruyama T, Otagiri M, Ishida T. Drug Delivery System for Refractory Cancer Therapy via an Endogenous Albumin Transport System. Chem Pharm Bull. 2020;68(7):583–588. doi:10.1248/cpb.c20-00026
187. Taguchi K, Urata Y, Anraku M, et al. Superior Plasma Retention of a Cross-Linked Human Serum Albumin Dimer in Nephrotic Rats as a New Type of Plasma Expander. Drug Metab Dispos. 2010;38(12):2124–2129. doi:10.1124/dmd.109.031989
188. Komatsu T, Hamamatsu K, Tsuchida E. Cross-Linked Human Serum Albumin Dimer Incorporating Sixteen (Tetraphenylporphinato)iron(II) Derivatives: synthesis, Characterization, and O2-Binding Property. Macromolecules. 1999;32(25):8388–8391. doi:10.1021/ma990826p
189. Komatsu T, Oguro Y, Teramura Y, et al. Physicochemical characterization of cross-linked human serum albumin dimer and its synthetic heme hybrid as an oxygen carrier. Biochim Biophys Acta. 2004;1675(1–3):21–31. doi:10.1016/j.bbagen.2004.08.010
190. Abraxane Summary of Product Characteristics. 2005. Available from: https://www.ema.europa.eu/en/documents/product-information/abraxane-epar-product-information_en.pdf.
191. Abraxane Efficacy-New Indication 09/06/2013 SUPPL 37. 2013. Available from: https://www.accessdata.fda.gov/drugsatfda_docs/nda/2013/021660Orig1s037.pdf.
192. Abraxane Approval FDA. 2005. Available from: https://www.accessdata.fda.gov/drugsatfda_docs/nda/2005/21660_ABRAXANE_approv.PDF.
193. Abraxane EPAR summary for the public. 2015. Available from: https://www.ema.europa.eu/en/documents/overview/abraxane-epar-summary-public_en.pdf.
194. Abraxane Efficacy-New Indication. 2012. Available from: https://www.accessdata.fda.gov/drugsatfda_docs/nda/2012/021660Orig1s031.pdf.
195. Pazenir Summary of Product Characteristics EMA. 2019. Available from: https://www.ema.europa.eu/en/documents/product-information/pazenir-epar-product-information_en.pdf.
196. Pazenir An overview of Pazenir and why it is authorised in the EU. 2019. Available from: https://www.ema.europa.eu/en/documents/overview/pazenir-epar-medicine-overview_en.pdf.
197. December 2023 – approved Drug Product List. 2023. Available from: https://www.fda.gov/media/71494/download.
198. Fyarro Label. 2021. Available from: https://www.accessdata.fda.gov/drugsatfda_docs/label/2021/213312Orig1s000Corrected_lbl.pdf.
199. Fyarro Product Quality Review(s). 2021. Available from: https://www.accessdata.fda.gov/drugsatfda_docs/nda/2022/213312Orig1s000ChemR.pdf.
200. Paclitaxel - summary of product characteristics. 2024. Available from: https://www.medicines.org.uk/emc/product/3891/smpc/print.
201. Paclitaxel Kabi 6 mg/mL summary of product characteristics.
202. Paxene 6 mg/mL summary of product characteristics.
203. Rapamune - Clinical Pharmacology and Biopharmaceutics Review Application Number 21-110. 2000.
204. Rapamune - Summary of Product Characteristics. European Medicines Agency.
205. Fyarro. Available from: https://www.fyarrohcp.com/.
206. Fyarro. Available from: https://www.fyarro.com/.
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms.php
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms.php
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.