")
Back to Journals » Cancer Management and Research » Volume 17
Anticancer Potential of Bioactive Compounds in Premna serratifolia, Premna odorata, and Premna tomentosa: A Review of In Vitro Evidence
Authors Febriyanti RM , Rafif SN, Mikdar NN, Hikmatiana BN, Maisyarah IT , Khatib A, Muhaimin M
Received 22 January 2025
Accepted for publication 4 April 2025
Published 27 May 2025 Volume 2025:17 Pages 1029—1045
DOI https://doi.org/10.2147/CMAR.S516204
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Antonella D'Anneo
Raden Maya Febriyanti,1,2,* Syauqi Nawwar Rafif,3,* Nazwa Nuraizza Mikdar,3,* Billa Nidia Hikmatiana,3,* Intan Timur Maisyarah,1,2 Alfi Khatib,4 Muhaimin Muhaimin1,2
1Department of Biological Pharmacy, Faculty of Pharmacy, Universitas Padjadjaran, Bandung, West Java, Indonesia; 2Herbal Study Center, Universitas Padjadjaran, Bandung, West Java, Indonesia; 3Pharmacist Professional Study Program, Faculty of Pharmacy, Universitas Padjadjaran, Bandung, West Java, Indonesia; 4Department of Pharmaceutical Chemistry, Kulliyyah of Pharmacy, International Islamic University Malaysia, Kuantan, Malaysia
*These authors contributed equally to this work
Correspondence: Raden Maya Febriyanti, Department of Biological Pharmacy, Faculty of Pharmacy Universitas Padjadjaran, Jl. Ir. Soekarno KM. 21, Jatinangor, West Java, 45363, Indonesia, Email [email protected] Muhaimin Muhaimin, Department of Biological Pharmacy, Faculty of Pharmacy Universitas Padjadjaran, Jl. Ir. Soekarno KM. 21, Jatinangor, West Java, 45363, Indonesia, Email [email protected]
Abstract: The genus Premna (Lamiaceae), widely distributed across tropical and subtropical regions, is renowned for its ethnomedicinal applications, including cardiotonic, antimicrobial, hepatoprotective, and antitumor properties. Despite these promising uses, the current body of literature relies predominantly on in vitro evidence, with limited knowledge regarding in vivo validation, metabolism, and bioavailability. This review synthesizes findings from the past decade on the Premna serratifolia group (P. serratifolia, P. odorata, and P. tomentosa), focusing on their bioactive compounds and mechanisms of in vitro anticancer activity. The compounds identified—such as flavonoids, terpenoids, and steroids—exhibit diverse actions, including cell cycle arrest, apoptosis induction, inhibition of metastasis, oxidative stress modulation, and autophagy. Key compounds like quercetin, kaempferol, and stigmasterol demonstrate multi-targeted actions, effectively regulating pathways such as PI3K/AKT and NF-κB while selectively targeting cancer cells. These findings underscore chemopreventive potential from P. serratifolia group and their ability to complement conventional cancer therapies, potentially reducing side effects and overcoming drug resistance. Furthermore, the review validates the ethnomedicinal use of Premna species and bridges traditional knowledge with modern oncology. However, the absence of comprehensive in vivo and clinical data warrants further research to fully harness these compounds’ potential. This study highlights P. serratifolia, P. odorata, and P. tomentosa as promising sources for novel plant-derived anticancer agents, offering opportunities for future drug discovery.
Keywords: phytochemicals, natural product-based therapy, chemoprevention, cytotoxic activity, multi-target mechanisms
Introduction
Noncommunicable diseases, characterized by their chronic and complex nature, are responsible for more than 70% of global deaths annually, equating to approximately 41 million fatalities. Among these, cancer ranks as the second leading cause of death, following cardiovascular diseases.1 According to GLOBOCAN 2020, a global cancer data source, new cancer cases are projected to rise significantly by 2040, representing a 47% increase compared to 2020.2 Cancer includes a diverse group of over 277 diseases, generally defined by uncontrolled cell growth. Any somatic cell has the potential to undergo malignant transformation through a cascade of genetic mutations that disrupt normal cell functions.3 This process, known as carcinogenesis, occurs in three stages: initiation, where genetic mutations occur; promotion, where mutated cells become malignant; and progression, marked by rapid, uncontrolled tumor growth. Additionally, metastasis—the spread of cancer cells to distant organs—is a major contributor to cancer-related.4
The growing global cancer burden highlights the urgent need for innovative, cost-effective chemopreventive, and therapeutic strategies that are accessible to populations worldwide. Natural products, with their unparalleled chemical diversity, provide a rich source of bioactive compounds with therapeutic potential.5 Among these, phytochemicals have proven to be vital in the development of anticancer drugs, with more than 60% of current chemotherapeutic agents derived from these compounds.6
The genus Premna, first described by Linnaeus in 1771, comprises about 200 species found across Australia, Africa, Asia, and the Pacific Islands. The name Premna derives from the Greek word “premon”, meaning tree stump, reflect the short and twisted trunks of Premna serratifolia L., the first species described in this genus. Typically, Premna species are shrubs or trees, though some exist as pyroherbs or lianas.7 Morphologically, Premna species are divided into two groups based on their twig, calyx, and fruit characteristics: the P. serratifolia group and the P. trichostoma group. The P. serratifolia group includes commonly found species like P. serratifolia, P. odorata, and P. tomentosa. In contrast, the P. trichostoma group includes rarer, geographically restricted species.8 The P. serratifolia group are renowned for their phytochemical richness, including compounds such as iridoid glycosides, diterpenoids, phenylethanoids, lignans, sesquiterpenes, ceramides, megastigmanes, and glyceroglycolipids. These metabolites contribute to the diverse bioactivities of Premna species, including immunomodulatory, antimicrobial, anti-hyperglycemic, anti-inflammatory, and cytotoxic effects.9,10
Despite these promising biological activities, current research predominantly consists of in vitro assays, and there remains a significant gap regarding the comprehensive understanding of anticancer mechanisms, bioactive constituents, and their therapeutic potential in clinical contexts. To bridge this gap, this review aims to summarize and evaluate the available in vitro data regarding the anticancer activities and mechanisms of bioactive compounds from P. serratifolia, P. odorata, and P. tomentosa. Emphasis is placed on identifying bioactive compounds, summarizing reported mechanisms of action, and discussing the implications of these findings for future research. Given the limited in vivo studies available, our review focuses primarily on in vitro cytotoxic, apoptotic, anti-metastatic, and related mechanistic data, while underscoring the necessity for further preclinical and clinical validation.
Materials and Methods
Search Strategy
We conducted literature search in PubMed, EBSCO, and Scopus from 2013 to 2023 to identify relevant studies on Premna serratifolia, Premna odorata, and Premna tomentosa. The primary search terms included “Premna serratifolia”, “Premna odorata”, “Premna tomentosa”, combined with “bioactive compounds” and “anticancer activity”. Only peer-reviewed articles presenting in vitro and/or in vivo evidence for the anticancer effects of bioactive compounds explicitly isolated from these three Premna species were included. Reviews or commentaries lacking primary data and studies on non-serratifolia group species were excluded.
Data Extraction and Validation
Two investigators (RMF and SNR) independently screened the titles and abstracts and then reviewed the full text of potentially relevant articles. Discrepancies were resolved through discussion with a third investigator (ITM). Extracted data included information on the experimental design, cell lines models, treatment doses, identified bioactive compounds, and reported outcome. The data were cross-verified to ensure consistency and accuracy.
Although the focus is on in vitro efficacy, some studies that mention initial in vivo experiments were considered if they provided substantial mechanistic or cytotoxic data. Potential conflicts of interest in the cited papers were not always explicitly stated; however, we acknowledge such factors may influence reported outcomes and interpretations.
Results and Discussion
Premna serratifolia, Premna odorata, and Premna tomentosa are widely distributed across tropical and subtropical regions and have been traditionally used to address a broad spectrum of ailments. Their phytochemical profiles, including flavonoids, sterols, and terpenoids, underline their potential as sources of bioactive compounds for therapeutic applications. Table 1 provides a comprehensive overview of three species within the Premna serratifolia group. Following, Table 2 summarizes recent in vitro anticancer studies examining the biological activities of various extracts and bioactive secondary metabolites derived from Premna serratifolia, Premna odorata, and Premna tomentosa. In the following paragraphs, we discuss anticancer mechanism of the bioactive compound such as induction of apoptosis, modulation of the cell cycle, inhibition of proliferation and migration, oxidative stress modulation, and autophagy induction. However, during compiling these findings, we recognized that certain references describing the anticancer mechanism of Premna species did not always provide in-depth of the identification of bioactive compound. To fill these gaps, we supplemented these mechanistic discussions with studies on the same compounds—albeit from other plant sources. While we acknowledge that subtle variations in phytochemical context can influence potency or synergy, the core biochemical pathways (eg, apoptosis, cell cycle arrest) generally remain consistent for well-characterized molecules. Thus, once a compound is structurally identified, its fundamental anticancer mechanism (eg, caspase activation, ROS modulation) is usually reproducible across different models.
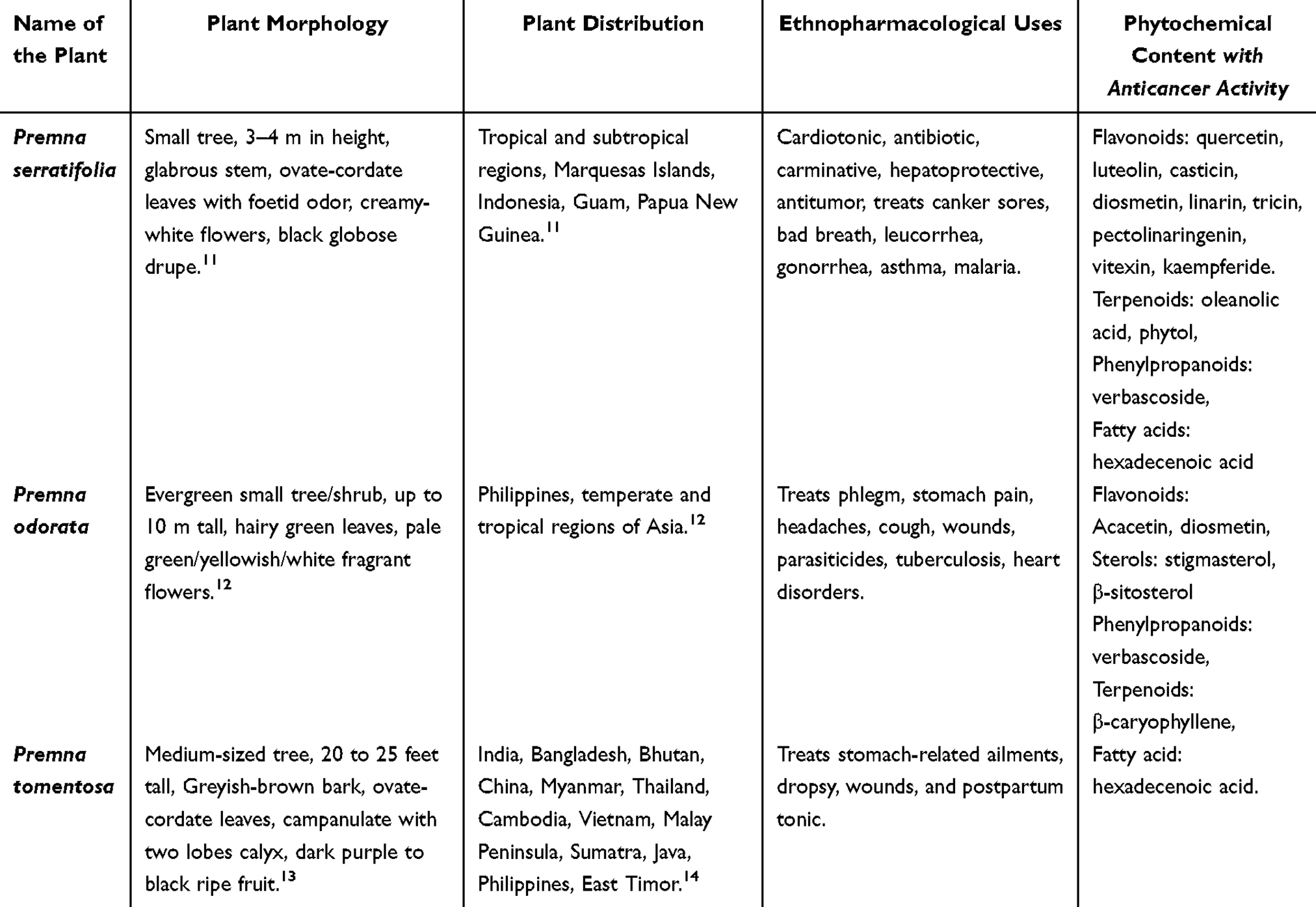 |
Table 1 Overview of Morphology, Distribution, Ethnopharmacological Uses, and Phytochemical Content of Premna Species |
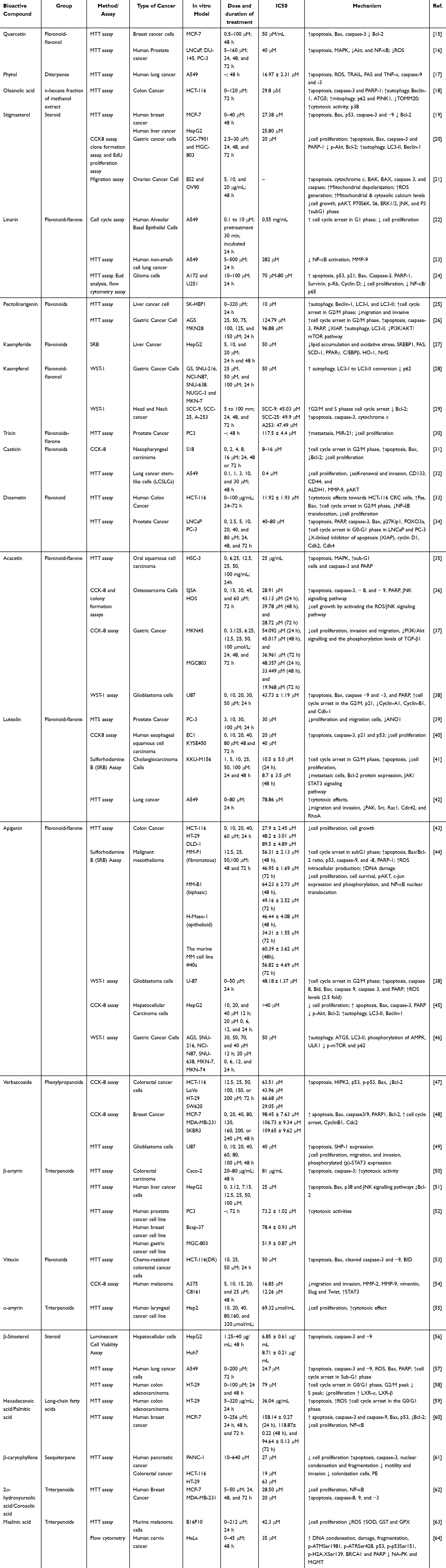 |
Table 2 Summary of Anticancer Mechanism of Premna Sp. Derived Bioactive Compound |
Mechanism of Anticancer
The anticancer potential of the three species within the Premna serratifolia group—P. serratifolia, P. odorata, and P. tomentosa—is attributed to their diverse array of bioactive compounds that target multiple cancer-related pathways. The Premna serratifolia group, exhibits a rich diversity of bioactive compounds categorized into several major classes of secondary metabolites. These compounds include flavonoids, terpenoids, steroids, phenylpropanoids, and long-chain fatty acids, all of which contribute to their anticancer potential.10 Among flavonoids, notable compounds such as quercetin, diosmetin, kaempferol-3-O-b-D-galactopyranoside, luteolin, and casticin have demonstrated potent anticancer activities through mechanisms like apoptosis induction, cell cycle arrest, and inhibition of proliferation and metastasis. Quercetin and kaempferol are particularly prominent for their ability to regulate oxidative stress and modulate signaling pathways such as NF-κB and PI3K/AKT.65
The terpenoid class is represented by compounds such as oleanolic acid and phytol which exhibit diverse anticancer mechanisms, including induction of apoptosis, autophagy, and modulation of mitochondrial function. Steroidal compounds such as stigmasterol and β-sitosterol play crucial roles in promoting apoptosis and disrupting cancer cell proliferation through mitochondrial depolarization and ROS generation. Phenylpropanoids, including verbascoside and pectolinarigenin, show remarkable potential in inhibiting cancer cell migration, invasion, and proliferation while promoting apoptosis and autophagy.10 Additionally, long-chain fatty acids, particularly hexadecanoic acid, have been identified for their capability to enhance ROS levels, induce apoptosis, and arrest cell cycle progression in cancer cells.66,67 A detailed overview of the anticancer mechanism of bioactive compounds extracted from Premna species focusing on their in vitro studies against various cancer types is presented in Table 2.
Induction of Cell Cycle Arrest
Dysregulation of the cell cycle is a hallmark of cancer, leading to unchecked proliferation. Compounds such as stigmasterol, linarin, kaempferol, and diosmetin derived from Premna species have been shown to arrest cancer cells at G0/G1, S, or G2/M phases, often accompanied by alterations in cyclin-dependent kinases and cell-cycle regulatory proteins. CDK inhibitors have emerged as promising therapeutic agents, forcing malignant cells into senescence or apoptosis by halting the cell cycle.68 Cancer cells, like L1-LW6, are notably more susceptible to cell death caused by CDK inhibition compared to their normal counterparts.
Findings from various in vitro studies highlight the role of bioactive compounds in targeting specific phases of the cell cycle. For instance, stigmasterol-induced subG1 arrest in ES2 cells, signaling apoptosis.21 Similarly, linarin disrupted the G1-to-S phase transition, resulting in G1 arrest in A549 cells.22 Kaempferol −3-O-b-D-galactopyranoside exhibited dual effects by halting SCC-9 cells in the S-phase and arresting HT-29 and LNCaP cells in the G0/G1 phase, linked to the downregulation of cyclin D1, cyclin E, and CDKs.29,59 Diosmetin also arrested the G1 and S phases in HCT-116 and PC-3 cells, respectively, by downregulating cyclin-dependent genes and increasing CDK inhibitors like p27Kip1, confirming its potential as a CDK inhibitor.33,34
Hexadecanoic acid and β-sitosterol significantly reduced the S and G2/M phases in HT-29 cells, suggesting these compounds arrest colon cancer cells in the G0/G1 phase.58,59 Pectolinarigenin caused G2/M arrest in SK-HEP-1, AGS, and MKN28 cells through the downregulation of cyclin B1 and CDK1, along with the upregulation of p21 and p53.25,26 Similarly, verbascoside-induced G2/M arrest in MCF-7 and MDA-MB-231 cells, which involved downregulating cyclin B1 and CDC2 and modulating the PI3K/AKT signaling pathway.48
Furthermore, casticin treatment-elevated p21 expression in S18 cells, further supporting its role in G2/M phase arrest.31 Additionally, diosmetin-suppressed genes associated with mitotic division, such as ttk, pttg2, and mad2l2, while enhancing inhibitors of cyclin A and B, such as p21 and gadd45. This led to the inhibition of mitosis and accumulation of cells in the G2/M phase. These findings strongly suggest that compounds from the P. serratifolia group could effectively regulate the cell cycle and hold significant potential for developing anticancer therapies.
Inhibition of Proliferation
Cancer cells exhibit an abnormal ability to survive beyond their normal lifespan and proliferate uncontrollably, making the inhibition of proliferation a key strategy in cancer treatment. Targeting the high proliferative rate of cancer cells through specific interventions has proven effective in slowing or stopping cancer progression.69 Several factors contribute to cancer cell proliferation, including epithelial-to-mesenchymal transition (EMT), dysregulated cell cycle proteins, and constitutive activation of signal transduction pathways such as Wnt, Notch, IGF, PI3K/Akt, NF-κB, and Hedgehog (Hh).69 These factors promote uncontrolled cell proliferation, metastasis, and stem cell growth. Bioactive flavonoids like quercetin, linarin, casticin, and luteolin reduce cancer cell viability in in vitro assays, often by downregulating pro-survival signaling (eg, Akt, NF-κB) and upregulating cell cycle inhibitors (eg, p21, p27).
Genus Premna was reported rich in flavonoids such as quercetin, linarin, tricin, casticin, diosmetin, luteolin, acacetin, and apigenin. Flavonoids are known as key mediators of antiproliferative activity. Quercetin has been shown to inhibit prostate cancer (PCa) cell proliferation by modulating ROS production and interfering with MAPK, Akt, and NF-κB signaling pathways.16 Similarly, linarin suppresses proliferation in human lung carcinoma (A549) cells by downregulating Akt activation and upregulating cyclin-dependent kinase inhibitor p27Kip1.22 In glioma cells, linarin exerts antiproliferative effects by upregulating p53 and downregulating NF-κB/p65, thereby inhibiting cell growth.24
Casticin effectively suppresses proliferation in lung cancer stem-like cells (LCSLCs) and nasopharyngeal carcinoma (NPC) cells by inducing G2/M arrest and selectively inhibiting PI3K pathways.31,32 Diosmetin, a plant flavonoid, exerts antiproliferative effects on prostate and colon cancer cells by regulating proteins such as c-Myc, Bax, p27Kip1, and FOXO3a while inhibiting BMP and NF-κB pathways.33,34
Tricin reduces proliferation in prostate cancer cells by downregulating key proliferative markers, while luteolin exhibits antiproliferative activity through the suppression of migration and G2/M phase arrest, mediated by p21 and p53 regulation.39,40 Additionally, luteolin inhibits metastasis in cholangiocarcinoma cells by regulating JAK/STAT3 signaling pathways.41
Beyond flavonoids, other classes of secondary metabolites, including triterpenoids, sesquiterpenes, phenylpropanoids, and fatty acids, also demonstrate antiproliferative effects. β-caryophyllene suppresses colorectal cancer cell growth by disrupting mitochondrial membrane potential and inducing apoptosis.61 Hexadecanoid acid inhibits breast cancer cell proliferation by enhancing apoptosis-related proteins such as caspase-3, Bax, and p53 while downregulating anti-apoptotic protein Bcl-2.60
Stigmasterol inhibits gastric and ovarian cancer cell proliferation by targeting the PI3K and Akt/mTOR signaling cascades.20,21 Verbascoside further demonstrates antiproliferative effects by inducing cell cycle arrest, apoptosis, and inhibiting glioblastoma cell migration and invasion through SHP-1 activation and STAT3 pathway inhibition.48,49
Inhibition of Migration, Invasion, and Metastasis
Metastasis accounts for a large percentage of cancer-related deaths. This process involves cancer cell migration, invasion, and eventual colonization of secondary tissues through intravasation into the blood or lymphatic systems.70 The ability of cancer cells to migrate and invade tissues is a hallmark of metastatic progression and is largely driven by pathways such as STAT3 activation, which upregulates MMP-2 and promotes epithelial–mesenchymal transition (EMT).71
Flavonoids, a group of secondary metabolites abundant in Premna species, have demonstrated significant anti-metastatic properties in various in vitro cancer models. These effects are mediated by the regulation of signaling pathways and molecules associated with migration and invasion, including MMPs, TGF-β, and NF-κB.72 Among the bioactive flavonoids in P. serratifolia leaves with demonstrated antimetastatic activity are linariin, pectolinaringenin, tricin, and casticin.
Linariin has been shown to significantly reduce IR-induced migration and invasion of A549 cells by downregulating MMP-9 expression and suppressing NF-κB activation through inhibition of NF-κB and IκB-α phosphorylation.23 Pectolinaringenin inhibited the invasion and migration of human SK-HEP1 carcinoma cells, as evidenced by wound healing and transwell assays, further validating its anti-metastatic potential.25 Tricin, through its ability to reduce MiR-21 overexpression, effectively decreased metastasis and chemoresistance in prostate cancer cells, indicating its potential in combating metastatic progression.30 Similarly, casticin inhibited the invasion of sphere-forming cells (SFCs) derived from A549 lung cancer cells by reducing MMP-9 activity in a concentration-dependent manner.32
In addition to flavonoids, other secondary metabolites from Premna species have shown anti-metastatic effects. For instance, β-caryophyllene from Aquilaria crassna essential oil significantly reduced cell invasion in colorectal cancer, underscoring its potential as an anti-metastatic agent.61 Similarly, verbascoside decreased glioblastoma cell migration and invasion by increasing SHP-1 expression, downregulating phosphorylated STAT3, and reducing MMP-2 and MMP-9 levels.49
Induction of Apoptosis
Apoptosis, or programmed cell death, is a tightly regulated process essential for maintaining cellular homeostasis, development, and the removal of damaged or unwanted cells. Unlike necrosis, apoptosis is a controlled and orderly mechanism that prevents inflammation and minimizes damage to surrounding tissues. Apoptosis occurs through two interconnected pathways: the intrinsic (mitochondrial) pathway and the extrinsic (death receptor) pathway, both of which involve the activation of caspase family proteases. Caspases serve as both initiators and executioners in the apoptotic process, breaking down cellular components to facilitate cell death.73
The intrinsic pathway is regulated by the BCL-2 protein family, which includes anti-apoptotic (eg, BCL-2, BCL-xL), pro-apoptotic (eg, BAX, BAK), and BH3-only proteins (eg, BIM, BID). Triggers for apoptosis increase BH3-only protein expression, leading to mitochondrial membrane disruption, cytochrome c release, and apoptosome formation, which activates caspase-9 and downstream executioner caspases-3 and -7. The extrinsic pathway involves death receptor ligands such as Fas-L and TNF binding to receptors, forming the death-inducing signaling complex (DISC) and activating caspases-8 and -10. These pathways converge to initiate apoptosis via cleavage of structural and regulatory proteins, aided by inhibitors such as XIAP and regulators like SMAC.73,74
Several bioactive compounds from the Premna genus (P. serratifolia group) have shown strong apoptotic effects in various cancer cell models. Quercetin, at a dose of 50 μM/mL, effectively activates the apoptotic signaling pathway in MCF-7 cells by increasing Bax and caspase-3 expression while downregulating Bcl-2 expression.15 Similarly, stigmasterol enhances Bax expression, reduces Bcl-2 levels, and upregulates caspase-9 and -3 mRNA expression in a dose-dependent manner in ovarian and gastric cancer cells. It also increases mitochondrial depolarization and ROS generation in ES2 and OV90 cell line.19,21
Phytol induces apoptosis in A549 lung cancer cells via ROS-mediated activation of TRAIL, FAS, and TNF-α receptors and caspases-9 and -3, operating through both intrinsic and extrinsic pathways (Thakor, 2017). In colon cancer HCT116 cells, oleanolic acid activates cleaved caspase-3 and PARP-1, inducing apoptosis.18 Linarin induces apoptosis in glioma cells by activating p53, Bax, and caspase-3 and cleaving PARP.24 Pectolinaringenin decreases XIAP levels, activates caspase-3, and cleaves PARP in AGS and MKN28 gastric carcinoma cells.26 Kaempferol −3-O-b-D-galactopyranoside induces apoptosis by triggering cytochrome c release and activating caspase-3 in head and neck cancer cells.29
Diosmetin activates apoptosis in colon and prostate cancer cells by upregulating Fas and Bax expression, releasing cytochrome c, and cleaving caspase cascades while inhibiting XIAP.33,34 Acacetin induces apoptosis in oral squamous cell carcinoma, osteosarcoma, and glioblastoma through caspase activation, Bcl-2/Bax regulation, and mitochondrial depolarization.35,36 Luteolin triggers apoptosis in cholangiocarcinoma cells by increasing p21 and p53 levels and activating caspase-3 and related pro-apoptotic proteins such as CYT-c and cPARP.40
Other flavonoids such as apigenin [ref] regulate apoptosis via the Bax/Bcl-2 ratio, p53 expression, and caspase-3 and PARP cleavage in colon and hepatocellular carcinoma cells.45 Casticin increases Bax/Bcl-2 expression and decreases the Bcl-2/Bax ratio in nasopharyngeal carcinoma cells, promoting apoptosis.31 Vitexin [ref] induces caspase-9 and −3 activation while upregulating pro-apoptotic proteins such as BID and Bax in colorectal cancer cells.53 Verbascoside induces apoptosis by modulating apoptosis-related proteins, increasing pro-apoptotic factors such as HIPK2, p53, and cleaved PARP1 while reducing anti-apoptotic proteins like Bcl-2 and survivin in colorectal and glioblastoma cells.48
Additionally, β-sitosterol and hexadecenoic acid induce apoptosis by increasing ROS levels, modulating apoptotic signaling pathways, and activating caspase cascades in colon and breast cancer cells.56,59 Furthermore, β-amyrin exerts its anticancer properties via apoptosis induction and activation of JNK and P38 signalling pathway.50,51 β-caryophyllene disrupts mitochondrial membrane potential, promoting apoptosis via nuclear condensation and fragmentation.61
Effect on Oxidative Stress
Reactive oxygen species (ROS), known as partially reduced oxygen metabolites, exhibit potent oxidizing properties that, in high concentrations, can damage cellular components. However, at moderate levels, ROS play essential roles in cellular signaling and homeostasis. Elevated ROS levels in cancer cells can induce cell cycle arrest, senescence, and apoptosis through intrinsic (mitochondrial) and extrinsic (death receptor) apoptotic pathways. These effects are mediated by the activation of signaling cascades such as ASK1/JNK and ASK1/p38, which suppress anti-apoptotic factors and promote cell death. ROS also regulate apoptotic effectors, such as the Bcl-2 protein family and cytochrome c, leading to caspase activation, DNA fragmentation, and PARP cleavage, hallmark events in apoptosis.75
In cancer therapy, ROS can serve a dual role: while excessive ROS levels promote apoptosis in malignant cells, their suppression can protect normal cells from oxidative damage. Flavonoids, as polyphenolic compounds, play a critical role in modulating ROS levels and protecting cells against oxidative stress. Interestingly, flavonoids can also induce oxidative stress in cancer cells, triggering apoptosis. Several flavonoid compounds in the Premna serratifolia group, such as quercetin, kaempferide, acacetin, and apigenin, have demonstrated ROS-modulating properties in various cancer models.27,38,44
Quercetin demonstrated differential effects on ROS in prostate cancer cells, quenching ROS in LNCaP and PC-3 cells while elevating ROS levels in DU-145 cells, leading to the activation of Akt and NF-κB pathways and subsequent cancer cell death.16 Acacetin induced ROS-mediated activation of the JNK/c-Jun signaling pathway in osteosarcoma cells, amplifying apoptotic signaling.36 Similarly, apigenin induced a dose-dependent ROS increase in multiple myeloma (MM) and glioblastoma U87 cells, triggering apoptosis through caspase activation and DNA fragmentation.38,44
Steroidal compounds like stigmasterol and β-sitosterol also exhibit ROS-modulating activity. Stigmasterol induces ROS production in ES2 and OV90 ovarian cancer cells, contributing to mitochondrial depolarization and apoptosis.21 β-Sitosterol triggers apoptosis in HT-29 colon cancer cells through ROS-mediated mitochondrial dysregulation, evidenced by caspase-3 and −9 activation, Bcl-2/Bax ratio alteration, and cytochrome c release.57
Other secondary metabolites from the P. serratifolia group also demonstrate ROS-related activities. Phytol induces ROS-mediated apoptosis in A549 lung cancer cells by activating TRAIL, FAS, and TNF-α receptors and caspase cascades.17 Hexadecanoic acid significantly enhances ROS generation in HT-29 colon cancer cells, leading to apoptosis through mitochondrial pathways.59
Modulation of Autophagy
Autophagy, a cellular degradation process crucial for maintaining homeostasis, involves the sequestration and lysosomal degradation of damaged organelles and protein aggregates. This process, derived from the Greek words “auto” (self) and “phagia” (eating), is vital for cellular health and survival under stress conditions.76 Autophagy is regulated by several upstream mechanisms, including the mTORC1 and AMPK pathways, which respond to cellular energy and nutrient levels. Under nutrient depletion, mTORC1 inhibition and AMPK activation promote autophagy initiation via phosphorylation of ULK1/2 and Atg13, enabling autophagosome formation. Dysfunctional regulation of autophagy has been implicated in cancer progression, where cancer cells bypass regulatory circuits to sustain high autophagic activity, promoting tumor survival.77
The role of natural products in modulating autophagy has been increasingly recognized in cancer therapy. Bioactive compounds from the Premna serratifolia group have demonstrated potent autophagy-regulating effects in various cancer models, either promoting autophagic cell death or inducing selective degradation of cellular components to inhibit tumor growth. Key autophagic markers such as LC3-I, LC3-II, and Beclin-1 are commonly evaluated to assess autophagy activation.
Oleanolic acid, a triterpenoid from leaf extract of P. serratifollia [ref oleanolic acid], induces autophagy by upregulating Beclin-1, ATG5, and LC3B expression, thereby promoting autophagosome formation. It also facilitates mitophagy by increasing p62 and PTEN-induced kinase 1 expression while reducing TOMM20 levels, which contributes to oxidative stress mitigation and cancer inhibition.18 Apigenin enhances autophagy in HepG2 cells by increasing LC3-II levels and GFP-LC3 puncta, alongside upregulating ATG5 and AMPK phosphorylation and downregulating p-mTOR and p62 in gastric cancer cells, leading to autophagic cell death.45,46
Kaempferol-3-O-b-D-galactopyranoside exhibits similar effects by increasing LC3-II expression and decreasing p62 levels in gastric cancer cells. It activates the IRE1-JNK-CHOP signaling pathway, inducing an ER stress response and autophagic cell death.28 Pectolinaringenin and stigmasterol modulate autophagy by inhibiting the PI3K/AKT/mTOR signaling pathway, reducing p-4EBP1, p-p70S6K, and p-eIF4E levels. These effects are associated with increased Beclin-1, LC3-I, and LC3-II expression, alongside the formation of autophagosomes and acidic vesicular organelles, as observed via electron microscopy.20,25,26
Table 3 summarizes the diverse anticancer mechanisms of bioactive compounds identified in the Premna serratifolia group, categorized into six major pathways. Cell cycle arrest is mediated by compounds like β-sitosterol, casticin, diosmetin, kaempferol-3-O-b-D-galactopyranoside, and linarin, which target cyclin-dependent kinases, leading to disruption in the G1, S, or G2/M phases. Induced apoptosis, a critical pathway for eliminating cancer cells, involves compounds such as apigenin, diosmetin, and stigmasterol. These compounds trigger both intrinsic (mitochondrial) and extrinsic (death receptor) apoptotic pathways through the activation of caspases and modulation of pro- and anti-apoptotic proteins. The modulation of oxidative stress plays a dual role in cancer treatment, where compounds like quercetin and acacetin either mitigate oxidative damage in normal cells or induce excessive ROS production in cancer cells to promote apoptosis. Autophagy regulation, a process that facilitates cellular homeostasis, is influenced by compounds such as kaempferol-3-O-b-D-galactopyranoside, oleanolic acid, and pectolinagerin, which target mTOR and AMPK signaling to promote autophagic cell death in cancer cells. Compounds such as β-caryophyllene, tricin, and stigmasterol inhibit migration, invasion, and metastasis by suppressing key signaling pathways like NF-κB and matrix metalloproteinases (MMPs), thereby reducing the metastatic potential of cancer cells. Finally, the inhibition of cell proliferation is achieved by compounds like diosmetin, verbascoside, and luteolin, which regulate oncogenic pathways such as PI3K/AKT and Wnt. Collectively, the Premna sp. represents a promising avenue toward the development of innovative, targeted, and less toxic cancer therapies that meet the goals of modern oncology.
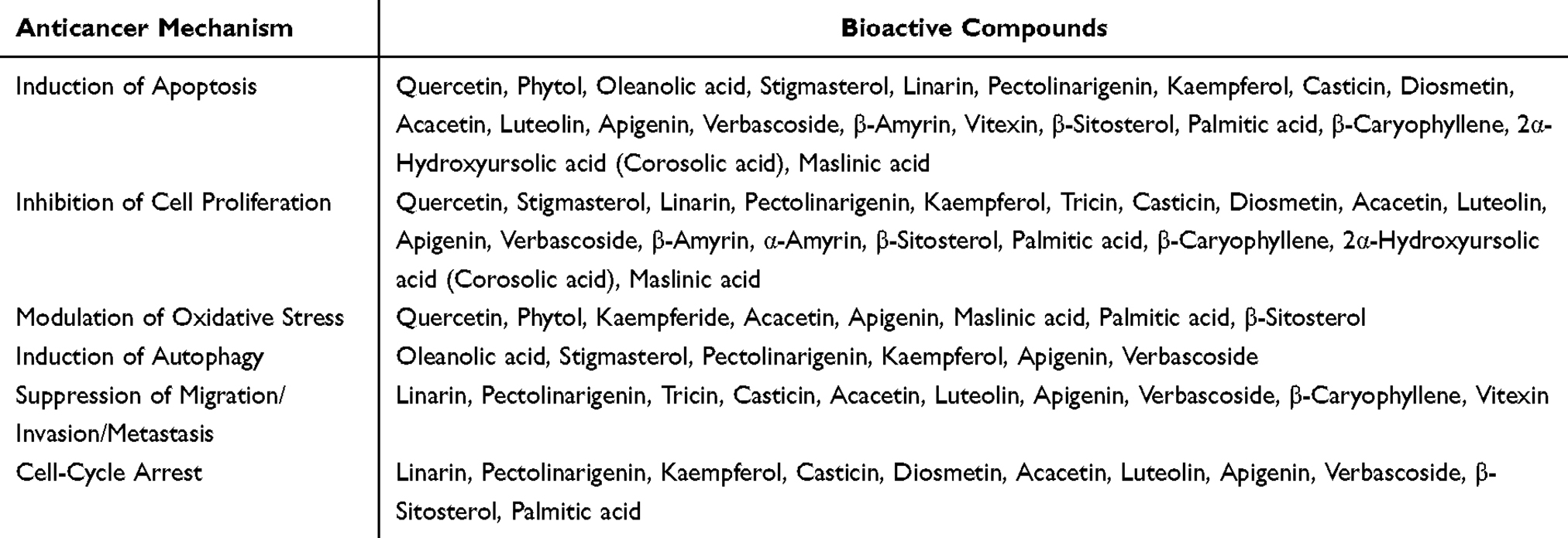 |
Table 3 Mechanism-Based Classification Table for the Bioactive Compounds Derived from Premna Serratifolia Group |
However, although the bioactive compounds from the Premna serratifolia group exhibit promising anticancer effects in vitro, their clinical translation requires careful assessment of pharmacokinetics and toxicology in in vivo models. Key parameters such as absorption, distribution, metabolism, excretion (ADME), and bioavailability strongly influence therapeutic efficacy. Many flavonoids, terpenoids, and steroids, for instance, undergo extensive metabolic modifications (eg, glucuronidation, sulfation) that can attenuate or alter their cytotoxic potential. Moreover, limited water solubility and rapid clearance often hamper systemic availability in animal models. To date, only a few studies have addressed these aspects in detail for Premna species. Future research should include well-designed in vivo experiments to validate anticancer efficacy and examine optimal delivery methods, pharmacokinetic profiles, and potential toxicity. Ultimately, incorporating both metabolic and bioavailability investigations will more definitively position these compounds for clinical application.
Limitations of the Study
Although the present review provides comprehensive insights into the anticancer potential of bioactive compounds from Premna serratifolia, P. odorata, and P. tomentosa, several limitations need to be acknowledged. First, most of the evidence presented relies on in vitro studies, and there is a notable scarcity of robust in vivo data and clinical studies to confirm efficacy, safety, pharmacokinetics, and therapeutic relevance in more complex biological systems. Secondly, variability in the experimental design—such as differences in cell lines, experimental conditions, concentrations, and methodologies—may introduce inconsistencies or biases into the interpretation of results. Furthermore, the pharmacokinetic properties and bioavailability of identified compounds remain largely unexplored, posing challenges for translating laboratory findings into clinical applications. Thus, future research should incorporate standardized methods, systematic evaluation of pharmacokinetics and toxicity in animal models, and eventually clinical validation to substantiate the therapeutic utility of these promising natural compounds.
Conclusion
This review highlights the in vitro anticancer potential of bioactive compounds from the Premna serratifolia group, encompassing P. serratifolia, P. odorata, and P. tomentosa. Flavonoids, terpenoids, and steroids present in these species exhibit varied mechanisms of action—inducing cell cycle arrest, promoting apoptosis, inhibiting metastasis, modulating oxidative stress, and regulating autophagy—making them valuable prospects for the development of novel plant-based anticancer drugs. Compounds such as quercetin, kaempferol, and stigmasterol stand out for their ability to target multiple pathways in tumorigenesis. Nevertheless, the current evidence is predominantly from in vitro research, and thorough in vivo and clinical evaluations remain necessary to confirm therapeutic utility. By investigating pharmacokinetics, toxicity, and long-term safety, future studies can help realize the chemopreventive and therapeutic potential of these Premna species, potentially overcoming drug resistance and minimizing side effects in conventional cancer treatments.
Acknowledgments
This study is funded by Fundamental Research Grant 2024 from Indonesian Ministry of Education, Culture, Research, and Technology awarded to MM.
Disclosure
The authors report no conflicts of interest in this work.
References
1. World Health Organization. Noncommunicable Diseases. 2023.
2. Sung H, Ferlay J, Siegel RL, et al. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 Cancers in 185 Countries. CA Cancer J Clin. 2021;71(3):209–249. doi:10.3322/caac.21660
3. Hassanpour SH, Dehghani M. Review of cancer from perspective of molecular. Journal of Cancer Research and Practice. 2017;4(4):127–129. doi:10.1016/j.jcrpr.2017.07.001
4. Cadoná FC, Dantas RF, de Mello GH, Silva-Jr FP. Natural products targeting into cancer hallmarks: an update on caffeine, theobromine, and (+)-catechin. Crit Rev Food Sci Nutr. 2022;62(26):7222–7241. doi:10.1080/10408398.2021.1913091
5. Huang M, Lu JJ, Ding J. Natural products in cancer therapy: past, present and future. Nat Prod Bioprospect. 2021;11(1):5–13. doi:10.1007/s13659-020-00293-7
6. Buyel JF. Plants as sources of natural and recombinant anti-cancer agents. Biotechnol Adv. 2018;36(2):506–520. doi:10.1016/j.biotechadv.2018.02.002
7. de Kok R. The genus Premna L. (Lamiaceae) in the Flora Malesiana area. Kew Bull. 2013;68(1):55–84. doi:10.1007/s12225-013-9433-5
8. Amran AN, Noor NNM, Mohamed F. Micromorphological analyses of stomata and epidermal cell among different species in the genus premna l. (Lamiaceae). Malaysian Journal of Microscopy. 2023;19(1).
9. Altyar AE, Ashour ML, Youssef FS. Premna odorata: seasonal metabolic variation in the essential oil composition of its leaf and verification of its anti-ageing potential via in vitro assays and molecular modelling. Biomolecules. 2020;10(6):1–17. doi:10.3390/biom10060879
10. Dianita R, Jantan I. Ethnomedicinal uses, phytochemistry and pharmacological aspects of the genus Premna: a review. Pharm Biol. 2017;55(1):1715–1739. doi:10.1080/13880209.2017.1323225
11. Francis S, Anand Gideon V, Britto SJ, Vj D. A review on two endemic species of genus premna and their conservational importance. J Drug Delivery Ther. 2019;9:666–669. doi:10.22270/jddt.v9i4
12. Youssef FS, Ovidi E, Al MNM, Morphology AML. Anatomy and secondary metabolites investigations of premna odorata blanco and evaluation of its anti-tuberculosis activity using in vitro and in silico studies. Plants. 2021;10(9):1953. doi:10.3390/plants10091953
13. N VGM, Atmakur H, Katragadda SB, et al. Antioxidant, hepatoprotective and cytotoxic effects of icetexanes isolated from stem-bark of Premna tomentosa. Phytomedicine. 2014;21(4):497–505. doi:10.1016/j.phymed.2013.09.025
14. Kew Science. Premna tomentosa. Plants of the World Online.
15. Khorsandi L, Orazizadeh M, Niazvand F, Abbaspour MR, Mansouri E, Khodadadi A. Quercetin induces apoptosis and necroptosis in MCF-7 breast cancer cells. Bratislava Medical Journal. 2017;118(2):123–128. doi:10.4149/BLL_2017_025
16. Ward AB, Mir H, Kapur N, Gales DN, Carriere PP, Singh S. Quercetin inhibits prostate cancer by attenuating cell survival and inhibiting anti-apoptotic pathways. World J Surg Oncol. 2018;16(1). doi:10.1186/s12957-018-1400-z
17. Thakor P, Subramanian RB, Thakkar SS, Ray A, Thakkar VR. Phytol induces ROS mediated apoptosis by induction of caspase 9 and 3 through activation of TRAIL, FAS and TNF receptors and inhibits tumor progression factor Glucose 6 phosphate dehydrogenase in lung carcinoma cell line (A549). Biomedicine and Pharmacotherapy. 2017;87:92. doi:10.1016/j.biopha.2017.05.066
18. Poto’cnjak I, Simi´c L, Vukeli´c I, Bati’ci´c L, Domitrovi´c R. Oleanolic acid induces HCT116 colon cancer cell death through the p38/ FOXO3a/Sirt6 pathway. Chem Biol Interact. 2022, 1–10:110010
19. Abbas A, Al-Fatlawi Y. Stigmasterol inhibits proliferation of cancer cells via apoptotic regulatory genes. Bioscience Research. 2019;16(1).
20. Zhao H, Zhang X, Wang M, Lin Y, Zhou S. Stigmasterol simultaneously induces apoptosis and protective autophagy by inhibiting Akt/mTOR pathway in gastric cancer cells. Front Oncol. 2021;11. doi:10.3389/fonc.2021.629008
21. Bae H, Song G, Lim W. Stigmasterol causes ovarian cancer cell apoptosis by inducing endoplasmic reticulum and mitochondrial dysfunction. Pharmaceutics. 2020;12(6):488. doi:10.3390/pharmaceutics12060488
22. Seo DW, Cho YR, Kim W, Eom SH. Phytochemical linarin enriched in the flower of chrysanthemum indicum inhibits proliferation of A549 human alveolar basal epithelial cells through suppression of the akt-dependent signaling pathway. J Med Food. 2013;16(12):1086–1094. doi:10.1089/jmf.2012.2674
23. Jung CH, Han AR, Chung HJ, Ha IH, Um HD. Linarin inhibits radiation-induced cancer invasion by downregulating MMP-9 expression via the suppression of NF-κB activation in human non-small-cell lung cancer A549. Nat Prod Res. 2019;33(24):3582–3586. doi:10.1080/14786419.2018.1484460
24. Zhen ZG, Ren SH, Ji HM, et al. Linarin suppresses glioma through inhibition of NF-κB/p65 and up-regulating p53 expression in vitro and in vivo. Biomedicine and Pharmacotherapy. 2017:95. doi:10.1016/j.biopha.2017.08.023.
25. Liu S, Zhang J, Yang H, Zhang Q, Chen M. Pectolinarigenin flavonoid exhibits selective anti-proliferative activity in cisplatin-resistant hepatocellular carcinoma, autophagy activation, inhibiting cell migration and invasion, G2/M phase cell cycle arrest and targeting ERK1/2 MAP kinases. Journal of BUON. 2020;25(1):415–420.
26. Lee HJ, Saralamma VVG, Kim SM, et al. Pectolinarigenin induced cell cycle arrest, autophagy, and apoptosis in gastric cancer cell via PI3K/AKT/mTOR signaling pathway. Nutrients. 2018;10(8):1043. doi:10.3390/nu10081043
27. Tie F, Ding J, Hu N, Dong Q, Chen Z, Wang H. Kaempferol and kaempferide attenuate oleic acid‐induced lipid accumulation and oxidative stress in HepG2 cells. Int J Mol Sci. 2021;22(16):8847. doi:10.3390/ijms22168847
28. Kim TW, Lee SY, Kim M, Cheon C, Ko SG. Kaempferol induces autophagic cell death via IRE1-JNK-CHOP pathway and inhibition of G9a in gastric cancer cells. Cell Death Dis. 2018;9(9):1–14. doi:10.1038/s41419-018-0930-1
29. Kubina R, Krzykawski K, Dziedzic A, Kabała-Dzik A. Kaempferol and fisetin-related signaling pathways induce apoptosis in head and neck cancer cells. Cells. 2023;12(12):1568. doi:10.3390/cells12121568
30. Ghasemi S, Lorigooini Z, Wibowo JP, Amini-khoei H. Tricin isolated from Allium atroviolaceum potentiated the effect of docetaxel on PC3 cell proliferation: role of miR-21. Nat Prod Res. 2019;33(12):1828–1831. doi:10.1080/14786419.2018.1437439
31. Liu J, Yang J, Hou Y, et al. Casticin inhibits nasopharyngeal carcinoma growth by targeting phosphoinositide 3-kinase. Cancer Cell Int. 2019;19(1):1–16. doi:10.1186/s12935-019-1069-6
32. Liu F, Cao X, Liu Z, et al. Casticin suppresses self-renewal and invasion of lung cancer stem-like cells from A549 cells through down-regulation of pAkt. Acta Biochim Biophys Sin. 2014;46(1):15–21. doi:10.1093/abbs/gmt123
33. Koosha S, Mohamed Z, Sinniah A, Alshawsh MA. Investigation into the molecular mechanisms underlying the anti-proliferative and anti-tumorigenesis activities of diosmetin against HCT-116 human colorectal cancer. Sci Rep. 2019;9(1):1–17. doi:10.1038/s41598-019-41685-1
34. Oak C, Khalifa AO, Isali I, Bhaskaran N, Walker E, Shukla S. Diosmetin suppresses human prostate cancer cell proliferation through the induction of apoptosis and cell cycle arrest. Int J Oncol. 2018;53(2):835–843. doi:10.3892/ijo.2018.4407
35. Kim CD, Cha JD, Li S, Cha IH. The mechanism of acacetin-induced apoptosis on oral squamous cell carcinoma. Arch Oral Biol. 2015;60(9):1283–1298. doi:10.1016/j.archoralbio.2015.05.009
36. Wang S, Lin B, Liu W, et al. Acacetin induces apoptosis in human osteosarcoma cells by modulation of ros/jnk activation. Drug Des Devel Ther. 2020;14:5077–5085. doi:10.2147/DDDT.S275148
37. Zhang G, Li Z, Dong J, et al. Acacetin inhibits invasion, migration and TGF-β1-induced EMT of gastric cancer cells through the PI3K/Akt/Snail pathway. BMC Complement Med Ther. 2022;22(1). doi:10.1186/s12906-021-03494-w
38. Shendge AK, Chaudhuri D, Mandal N. The natural flavones, acacetin and apigenin, induce Cdk-Cyclin mediated G2/M phase arrest and trigger ROS-mediated apoptosis in glioblastoma cells. Mol Biol Rep. 2021;48(1):539–549. doi:10.1007/s11033-020-06087-x
39. Seo Y, Ryu K, Park J, et al. Inhibition of ANO1 by luteolin and its cytotoxicity in human prostate cancer PC-3 cells. PLoS One. 2017;12(3):e0174935. doi:10.1371/journal.pone.0174935
40. Chen P, Zhang JY, Sha BB, et al. Luteolin inhibits cell proliferation and induces cell apoptosis via down-regulation of mitochondrial membrane potential in esophageal carcinoma cells EC1 and KYSE450. Oncotarget. 2017;8(16). doi:10.18632/oncotarget.15832
41. Aneknan P, Kukongviriyapan V, Prawan A, Kongpetch S, Sripa B, Senggunprai L. Luteolin arrests cell cycling, induces apoptosis and inhibits the JAK/STAT3 pathway in human cholangiocarcinoma cells. Asian Pacific Journal of Cancer Prevention. 2014;15(12):5071–5076. doi:10.7314/APJCP.2014.15.12.5071
42. Masraksa W, Tanasawet S, Hutamekalin P, Wongtawatchai T, Sukketsiri W. Luteolin attenuates migration and invasion of lung cancer cells via suppressing focal adhesion kinase and non-receptor tyrosine kinase signaling pathway. Nutr Res Pract. 2020;14(2):127. doi:10.4162/nrp.2020.14.2.127
43. Shan S, Shi J, Yang P, et al. Apigenin restrains colon cancer cell proliferation via targeted blocking of pyruvate kinase M2-dependent glycolysis. J Agric Food Chem. 2017;65(37):8136–8144. doi:10.1021/acs.jafc.7b02757
44. Masuelli L, Benvenuto M, Mattera R, et al. In Vitro and in vivo anti-tumoral effects of the flavonoid apigenin in malignant mesothelioma. Front Pharmacol. 2017;8(JUN). doi:10.3389/fphar.2017.00373
45. Yang J, Pi C, Wang G. Inhibition of PI3K/Akt/mTOR pathway by apigenin induces apoptosis and autophagy in hepatocellular carcinoma cells. Biomedicine and Pharmacotherapy. 2018;103. doi:10.1016/j.biopha.2018.04.072
46. Kim TW, Lee HG. Apigenin induces autophagy and cell death by targeting EZH2 under hypoxia conditions in gastric cancer cells. Int J Mol Sci. 2021;22(24):13455. doi:10.3390/ijms222413455
47. Zhou L, Feng Y, Jin Y, et al. Verbascoside promotes apoptosis by regulating HIPK2-p53 signaling in human colorectal cancer. BMC Cancer. 2014;14(1). doi:10.1186/1471-2407-14-747
48. Ou Y, Zhang Y, Zhu X, Zhu D. Verbascoside induces the cell cycle arrest and apoptosis of breast cancer via suppression of the PI3K/AKT signaling pathway. doi:10.21203/rs.3.rs-1544829/v1
49. Jia WQ, Wang ZT, Zou MM, et al. Verbascoside inhibits glioblastoma cell proliferation, migration and invasion while promoting apoptosis through upregulation of protein tyrosine phosphatase SHP-1 and inhibition of STAT3 phosphorylation. Cellular Physiology and Biochemistry. 2018;47(5):1871–1882. doi:10.1159/000491067
50. Maiyo F, Moodley R, Singh M. Phytochemistry, cytotoxicity and apoptosis studies of Β-sitosterol-3-oglucoside and Β -amyrin from Prunus Africana. African Journal of Traditional, Complementary and Alternative Medicines. 2016;13(4):105–112. doi:10.21010/ajtcam.v13i4.15
51. Wen S, Gu D, Zeng H. Antitumor effects of beta-amyrin in Hep-G2 liver carcinoma cells are mediated via apoptosis induction, cell cycle disruption and activation of JNK and P38 signalling pathways. Journal of BUON. 2018;23(4):965–970.
52. Luo H, Cai Y, Peng Z, Liu T, Yang S. Chemical composition and in vitro evaluation of the cytotoxic and antioxidant activities of supercritical carbon dioxide extracts of pitaya (dragon fruit) peel. Chem Cent J. 2014;8(1). doi:10.1186/1752-153X-8-1
53. Bhardwaj M, Cho HJ, Paul S, et al. Vitexin induces apoptosis by suppressing autophagy in multidrug resistant colorectal cancer cells. Oncotarget. 2018;9(3):3278–3291. doi:10.18632/oncotarget.22890
54. Zhang W, Zhou L, Liu G. Vitexin inhibited the invasion, metastasis, and progression of human melanoma cells by targeting STAT3 signaling pathway. bioRxiv.2020:2020–09
55. Mirunalini S, Isabella S, Vaithiyanathan V. Antiproliferative effect of alpha amyrin on hep2 cells by inducing cytotoxicity and oxidant antioxidant status modifications. FoodSci: Indian Journal of Research in Food Science and Nutrition. 2016;3(2). doi:10.15613/fijrfn/2016/v3i2/139485
56. Vo TK, Ta QTH, Chu QT, Nguyen TT, Vo VG. Anti-hepatocellular-cancer activity exerted by β-sitosterol and β-sitosterol-glucoside from Indigofera zollingeriana miq. Molecules. 2020;25(13):3021. doi:10.3390/molecules25133021
57. Rajavel T, Packiyaraj P, Suryanarayanan V, Singh SK, Ruckmani K, Pandima Devi K. β-Sitosterol targets Trx/Trx1 reductase to induce apoptosis in A549 cells via ROS mediated mitochondrial dysregulation and p53 activation. Sci Rep. 2018;8(1). doi:10.1038/s41598-018-20311-6
58. La Paz S M-D, Fernández-Arche MA, Bermúdez B, García-Giménez MD. The sterols isolated from evening primrose oil inhibit human colon adenocarcinoma cell proliferation and induce cell cycle arrest through upregulation of LXR. J Funct Foods. 2015;12:64–69. doi:10.1016/j.jff.2014.11.004
59. Bharath B, Perinbam K, Devanesan S, AlSalhi MS, Saravanan M. Evaluation of the anticancer potential of Hexadecanoic acid from brown algae Turbinaria ornata on HT–29 colon cancer cells. J Mol Struct. 2021;1235. doi:10.1016/j.molstruc.2021.130229
60. Zafaryab M, Fakhri KU, MdA K, Hajela K, Rizvi MMA. In vitro assessment of cytotoxic and apoptotic potential of palmitic acid for breast cancer treatment. Int J Life Sci Res. 2019;7(1).
61. Dahham SS, Tabana YM, Ahmed Hassan LE, Khadeer Ahamed MB, Abdul Majid AS, Abdul Majid AMS. In vitro antimetastatic activity of Agarwood (Aquilaria crassna) essential oils against pancreatic cancer cells. Alexandria Journal of Medicine. 2016;52(2):141–150. doi:10.1016/j.ajme.2015.07.001
62. Jasim SA, Khalaf OZ, Alshahrani SH, et al. An in vitro investigation of the apoptosis-inducing activity of corosolic acid in breast cancer cells. Iran J Basic Med Sci. 2023;26(4):453–460. doi:10.22038/IJBMS.2023.67783.14834
63. Mokhtari K, Pérez-Jiménez A, García-Salguero L, Lupiáñez JA, Rufino-Palomares EE. Unveiling the differential antioxidant activity of maslinic acid in murine melanoma cells and in rat embryonic healthy cells following treatment with hydrogen peroxide. Molecules. 2020;25(17):4020. doi:10.3390/molecules25174020
64. Lu KW, Yang MD, Peng SF, et al. Maslinic acid induces DNA damage and impairs DNA repair in human cervical cancer HeLa cells. Anticancer Res. 2020;40(12):6869–6877. doi:10.21873/anticanres.14709
65. Mir SA, Dar A, Hamid L, et al. Flavonoids as promising molecules in the cancer therapy: an insight. Current Research in Pharmacology and Drug Discovery. 2023;6:100167. doi:10.1016/j.crphar.2023.100167
66. Singh C, Anand SK, Tiwari KN, Mishra SK, Kakkar P. Phytochemical profiling and cytotoxic evaluation of Premna serratifolia L. against human liver cancer cell line. 3 Biotech. 2021;11(3):115. doi:10.1007/s13205-021-02654-6
67. Hadiarti D, Haryadi W, Matsjeh S, Swasono RT, Awaliyah N. Profiling of α-glucosidase inhibitors from ethyl acetate fraction of Buas-buas (Premna serratifolia) leaves using UHPLC-Q-Orbitrap HRMS and protein–ligand interaction with molecular docking. J Appl Pharm Sci. 2023;13(2):89–98. doi:10.7324/JAPS.2023.130210
68. Wang Z. Cell Cycle Progression and Synchronization: an Overview. Methods in Molecular Biology. 2022;2579:3–23. doi:10.1007/978-1-0716-2736-5_1
69. Feitelson MA, Arzumanyan A, Kulathinal RJ, et al. Sustained proliferation in cancer: mechanisms and novel therapeutic targets. Semin Cancer Biol. 2015:35. doi:10.1016/j.semcancer.2015.02.006.
70. Fares J, Fares MY, Khachfe HH, Salhab HA, Fares Y. Molecular principles of metastasis: a hallmark of cancer revisited. Signal Transduct Target Ther. 2020;5(1). doi:10.1038/s41392-020-0134-x
71. Belmehdi O, Taha D, Abrini J, et al. Anticancer properties and mechanism insights of α-hederin. Biomedicine and Pharmacotherapy. 2023:165. doi:10.1016/j.biopha.2023.115205.
72. Liskova A, Koklesova L, Samec M, et al. Flavonoids in cancer metastasis. Cancers. 2020;12(6):1498. doi:10.3390/cancers12061498
73. Baig S, Seevasant I, Mohamad J, Mukheem A, Huri HZ, Kamarul T. Potential of apoptotic pathway-targeted cancer therapeutic research: where do we stand ? Nature publishing group. Cell death disease. 1–11. doi:10.1038/cddis.2015.275
74. Pfeffer CM, Singh ATK. Apoptosis: a target for anticancer therapy. Int J Mol Sci. 2018;19(2):448. doi:10.3390/ijms19020448
75. Arfin S, Jha NK, Jha SK, et al. Oxidative stress in cancer cell metabolism. Antioxidants. 2021;10(5):642. doi:10.3390/antiox10050642
76. Mahapatra KK, Panigrahi DP, Praharaj PP, et al. Molecular interplay of autophagy and endocytosis in human health and diseases. Biological Reviews. 2019;94(4):1576–1590. doi:10.1111/brv.12515
77. Hernandez GA, Perera RM. Autophagy in cancer cell remodeling and quality control. Mol Cell. 2022;82(8):1514–1527. doi:10.1016/j.molcel.2022.03.023
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms.php
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms.php
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.