")
Back to Journals » Drug Design, Development and Therapy » Volume 19
Decoding TMAO in the Gut-Organ Axis: From Biomarkers and Cell Death Mechanisms to Therapeutic Horizons
Authors Liu J, Ge P, Luo Y , Sun Z , Luo X, Li H , Pei B, Xun L, Zhang X, Jiang Y, Wen H, Liu J, Yang Q, Ma S, Chen H
Received 14 December 2024
Accepted for publication 8 April 2025
Published 29 April 2025 Volume 2025:19 Pages 3363—3393
DOI https://doi.org/10.2147/DDDT.S512207
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Prof. Dr. Tin Wui Wong
Jie Liu,1– 3,* Peng Ge,1– 3,* Yalan Luo,4,* Zhenxuan Sun,1– 3 Xinyu Luo,1– 3 Huijuan Li,5 Boliang Pei,1– 3 Lu Xun,1– 3 Xuetao Zhang,1– 3 Yunfei Jiang,1– 3 Haiyun Wen,1– 3 Jin Liu,1– 3 Qi Yang,6 Shurong Ma,1– 3 Hailong Chen1– 3
1Department of General Surgery, The First Affiliated Hospital of Dalian Medical University, Dalian, Liaoning, 116011, People’s Republic of China; 2Institute (College) of Integrative Medicine, Dalian Medical University, Dalian, Liaoning, 116044, People’s Republic of China; 3Laboratory of Integrative Medicine, The First Affiliated Hospital of Dalian Medical University, Dalian, Liaoning, 116011, People’s Republic of China; 4Department of Gastroenterology, The First Affiliated Hospital of Dalian Medical University, Dalian, Liaoning, 116011, People’s Republic of China; 5Department of Cardiology, The First Affiliated Hospital of Dalian Medical University, Dalian, Liaoning, 116011, People’s Republic of China; 6Department of Traditional Chinese Medicine, The Second Affiliated Hospital of Dalian Medical University, Dalian, 116023, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Hailong Chen, Email [email protected]
Abstract: The gut microbiota and its metabolites are bi-directionally associated with various human illnesses, which has received extensive attention. Trimethylamine N-oxide (TMAO) is a gut microbiota metabolite produced in the liver, which may serve the role of an “axis” connecting the gut and host organs. TMAO levels are significantly higher in the blood of individuals with cardiovascular, renal, neurological, and metabolic diseases. Endothelial cells are crucial for regulating microcirculation and maintaining tissue and organ barriers and are widely recognized as target cells for TMAO. TMAO not only induces endothelial dysfunction but also acts on various cell types, such as endothelial cells, epithelial cells, vascular smooth muscle cells, nerve cells, and pancreatic cells, triggering multiple cell death mechanisms, including necrosis and programmed cell death, thereby influencing host health. This paper thoroughly covers the origins, production, and metabolic pathways of TMAO, emphasizing its importance in the early detection and prognosis of human diseases in the “Gut-Organ” axis, as well as its mechanisms of influence on human diseases, particularly the cross-talk with cell death. Furthermore, we cover recent advances in treating human diseases by regulating gut microbiota structure and enzyme activity to influence TMAO metabolism and reduce TMAO levels, including the use of probiotics, prebiotics, antibiotics, anti-inflammatory drugs, antiplatelet drugs, hypoglycemic drugs, lipid-lowering drugs, and natural products.
Keywords: trimethylamine N-oxide, gut-organ axis, cell death, biomarker, therapeutic strategies
Introduction
The gut is a vital organ of the host. The gut microbiota plays a pivotal role in regulating host metabolism and immune responses and is closely associated with various human diseases. Metabolites produced by the gut microbiota serve as key mediators between the microbiota and the host, interacting with multiple systems in the body to regulate physiological and pathological functions.1 Trimethylamine N-oxide (TMAO) is a significant metabolite generated from dietary components by gut microbes through microbial enzymes (such as trimethylamine lyases). Dietary choline, L-carnitine, and betaine are metabolized by gut microbiota, generating trimethylamine (TMA); in the liver, TMA is metabolized by the enzyme flavin-containing monooxygenase 3 (FMO3) to produce TMAO, which then enters the bloodstream.2–6 In recent years, it has gradually been recognized that TMAO may act as an “axis” connecting the gut and host organs. Its research scope has expanded from cardiovascular diseases to other systemic diseases, including pulmonary, kidney, neurological, and pancreatic diseases. Elevated plasma TMAO levels are closely associated with the development and progression of various diseases. TMAO is expected to become a potential biomarker for the diagnosis and prognosis evaluation of specific diseases.
There is widespread recognition that TMAO causes endothelial cell (EC) dysfunction.7 TMAO can stimulate ECs to release nitric oxide (NO), adhesion molecules (ICAM-1, VCAM-1, E-selectin, etc), and interleukin-6 (IL-6) to control the adherence of immune cells.8 Additionally, while TMAO affects endothelial barrier function, it can also act on various types of cells, including vascular smooth muscle cells, epithelial cells, pancreatic cells, neurons, astrocytes, microglia, and oligodendrocytes, thereby exerting broad impacts on numerous human diseases. Notably, TMAO can induce various types of cell death, including necrosis and programmed cell death (PCD). Necrosis is a kind of cell death that occurs passively. PCD is a manner of cell death actively triggered by internal and external stimuli. Apoptosis is the most common form of PCD, and other kinds of PCD include autophagy, pyroptosis, and ferroptosis, among others.9
This paper presents a detailed overview of the biomarker function of TMAO in the “gut-organ” axis, as well as the regulation mechanism of TMAO on the “gut-organ” axis, particularly in relation to the crosstalk mechanisms involved in cell death. Concisely, we first present the generation, synthesis, and metabolic pathways of TMAO. After applying strict criteria for literature screening, we selected clinical trials focused on TMAO to determine its potential as a biomarker for specific disorders through the “gut-organ” axis. We also provide a concise overview of the mechanism by which TMAO triggers PCD. Ultimately, we offer a summary of some medications that specifically aim to inhibit or regulate the generation, synthesis, or metabolism of TMAO. Our review offers new insights into the potential mechanisms of TMAO in disease onset and progression while emphasizing its crucial role in health regulation.
Methods
This narrative review included a literature search using the PubMed and Web of Science databases with the keywords: “(TMAO OR Trimethylamine N-oxide)” AND (“clinical study” OR “animal model” OR “in vitro study”). We included high-quality, peer-reviewed literature over the last 20 years (2004–2024), including cohort studies, randomized controlled trials, systematic reviews, meta-analyses, and original research publications. Studies were rejected if they were not in English, were duplicates, lacked full-text access, or consisted just of abstracts or comments without empirical data. It is essential to emphasize that, in contrast to systematic reviews or meta-analyses, we concentrated on presenting pertinent clinical and preclinical data on TMAO to provide complete insights. In Regulation Mechanism of TMAO in the “Gut-Organ” Axis: Crosstalk in Cell Death, we retrieved pertinent information about clinical features and the assessment of TMAO as a biomarker from chosen clinical trials. Subsequently, we qualitatively discussed the results within the literature in a narrative format, emphasizing significant findings to illustrate the potential use of TMAO in predicting, diagnosing, or prognosticating human illnesses. In Diverse Therapeutic Strategies Targeting TMAO, we curated the most pertinent preclinical and in vitro studies on TMAO, emphasizing findings associated with TMAO-induced cell death to deliver a thorough summary of its role in the development and progression of diseases along the “gut-organ (lung, pancreas, heart, kidney, and brain)” axis. In Conclusion and Future Directions, we reviewed clinical and preclinical data on therapeutic techniques aimed at TMAO, with the objective of elucidating the possible prospects of TMAO-targeted interventions in disease therapy.
Source, Synthesis and Metabolism of TMAO
Choline, L-carnitine, betaine, gamma-butyryl betaine in animal-derived foods (eg, red meat, eggs), and ergothione derived from mushrooms are critical precursors for the production of TMA.10–13 These compounds are primarily derived from the diet and all contain a trimethylamine group. They can be metabolized into TMA in the gut through the action of various enzymes in interaction with certain gut microbiota. TMAO, abundant in crustaceans, fish, and other seafood products, can also be directly assimilated into the human body through the gastrointestinal tract. Choline is a water-soluble nutrient that is essential for human health. Exogenous supplementation is required to attain the necessary choline levels for human health. Choline is primarily present in animal-derived foods as free, water-soluble compounds (choline phosphate, choline glycerophosphate) and fat-soluble compounds (phosphatidylcholine, sphingomyelin). The phospholipase D enzyme can convert phosphatidylcholine, also referred to as lecithin, into choline. Choline kinase can catalyze the conversion of choline to lecithin, and the process is bidirectional between PC and choline.5 Furthermore, the seeds of cruciferous plants contain choline in the form of phenolic cholinesterase. Choline erucate, abundant in cauliflower, is also a precursor of TMAO.3,4 Choline TMA-lyase (CutC) catalyzes the production of TMA from choline. It is important to note that the choline-TMA conversion process can be substantially influenced by the composition and diversity of the gut microbiome, which in turn impacts the production and accumulation of TMAO. L-carnitine is another precursor of TMA that is typically obtained from meat (red meat is exceptionally high in L-carnitine) and eggs. The primary function of L-carnitine is to transport long-chain fatty acids from the cytoplasm to the mitochondria for β-oxidation, which is essential for the body’s energy metabolism.2,6 Two critical enzymes involved in the conversion of L-carnitine to TMA are carnitine oxygenase (CntA) and carnitine reductase (CntB).14 Betaine is a choline derivative that is present in both animal and plant diets. Choline dehydrogenase and betaine aldehyde dehydrogenase can continuously convert choline to betaine. Carnitine dehydrogenase can also catalyze the conversion of L-carnitine to betaine. Betaine is converted to TMA through the action of betaine reductase. Ergothione is exclusively obtained through the diet (beans, livestock products, or mushrooms) and is converted to TMA by degrading the enzyme ergothionase.5 Briefly, the conversion of dietary precursors into TMA involves four enzymatic pathways, including the choline-utilizing TMA lyase system (CutC/D), the carnitine Rieske-type oxygenase/reductase system (CntA/B), betaine reductase, and TMAO reductase. Additionally, YeaW/X, a system homologous to CntA/B, can also metabolize carnitine, choline, γ-butyrobetaine, and betaine to generate TMA.15
In mammals, this process is catalyzed by enzymes encoded by genes such as CntA/B, CutC, and betaine reductase. Although less than 1% of gut microbiota possess the genes required for TMA synthesis, these relevant microbial communities are still sufficient to drive significant TMA production.16 As a core step in TMAO biosynthesis, the gut microbiota plays a pivotal role in regulating the generation and metabolism of TMA. Key microbial groups involved in this process include Firmicutes, such as Lachnoclostridium, Clostridium hathewayi, Clostridium asparagiformis, and Clostridium sporogenes, as well as Proteobacteria, such as Escherichia coli MS 200-1, Escherichia fergusonii, Proteus penneri, Edwardsiella tarda, and Desulfovibrio desulfuricans.17,18 Additionally, other gut microorganisms, including Deferribacteraceae, Anaeroplasmataceae, Prevotellaceae, and Enterobacteriaceae, also participate in this process.19–21 Considering the conserved nature of TMAO metabolism, we posited that fluctuations in TMAO levels throughout many illnesses, including cardiovascular, renal, and neurological disorders, are affected by alterations in gut microbiota. The gut microbiota significantly contributes to the progression of numerous illnesses and exhibits distinct alterations in response to different conditions. However, they may all result in the overexpression of TMAO, which may impact several organs.
TMA is absorbed into the liver through passive diffusion. In the liver, it is processed by FMOs, particularly FMO3, to create TMAO.22 After its formation, TMAO is excreted from the liver into the bloodstream via specialized transporters. ABCB1 (MDR1) and ABCG2 (BCRP), both members of the ATP-binding cassette (ABC) transporter family, play a pivotal role in facilitating the diffusion of TMAO.23 Notably, FMO3 is not only highly expressed in the liver but also significantly in the lungs, adrenal glands, and aortic tissues.24,25 This indicates the presence of several locations for the production of TMAO (Figure 1).
 |
Figure 1 Schematic representation of TMAO source, synthesis, and excretion. Initially, foods rich in L-carnitine, choline, betaine, ergothioneine, and γ-butyrobetaine, primarily fish, meat, eggs, and dairy products, are metabolized into TMA by the gut microbiota and their associated enzymes (Source). Most of the TMA ingested or generated in the gut is swiftly absorbed into the portal circulation via passive diffusion across intestinal epithelial cell membranes, subsequently undergoing oxidation to TMAO through the action of FMO3 in the liver. TMAO can be directly sourced from fish, shellfish, and marine invertebrates (Synthesis). Finally, TMAO is filtered by the kidneys and excreted via three main pathways: urinary excretion, fecal elimination, and respiratory clearance (Excretion). Created in BioRender. Ge, P. (2025) https://BioRender.com/k35g185. Abbreviations: TMA, Trimethylamine; TMAO, Trimethylamine-N-oxide; FMO3, Flavin-containing monooxygenase 3. |
The Biomarker Role of TMAO in “Gut-Organ” Axis: Clinical Reality
TMAO in Gut-Lung Axis
The gut-lung axis is a bidirectional pathway facilitating interaction between the gut and lungs via symbiotic microbes at a distance. Research on the gut-lung axis has revealed intricate interactions between the respiratory and gut microbiota, as well as between respiratory and gastrointestinal illnesses. The digestive system and the respiratory tract have a common origin in embryonic development. Moreover, in gut microbiota imbalance, the lung becomes the most susceptible organ. The process may include bacteria and their metabolites entering the bloodstream, disseminating throughout the body, initiating an inflammatory cascade, and culminating in sepsis.26 Wang et al found TMAO to be a possible biomarker for endoplasmic reticulum stress (ERS) in lung tissue after mice were given tunicamycin, which caused lung damage.27 Cao et al found that changing the gut microbiota, specifically by lowering the production of TMA and TMAO, may help reduce acute lung injury caused by sepsis by blocking the nuclear factor-κB signaling pathway.28 Chou et al originated the term “paradox of TMAO.” Researchers found a negative correlation between plasma TMAO levels and cardiovascular mortality in individuals suffering from severe sepsis. This indicates that TMAO levels represent the body’s inflammatory condition and nutritional status.29 The respiratory tract has its own microbiota, and pulmonary inflammation may lead to gut dysbiosis.30 Coronavirus disease 2019 (COVID-19) is an infectious disease caused by Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2). The virus primarily invades the respiratory system but also affects the gastrointestinal tract. SARS-CoV-2 has been detected in esophageal, gastric, duodenal, rectal, and fecal samples of COVID-19 patients. The gastrointestinal symptoms in COVID-19 patients are mainly manifested as diarrhea, which is associated with a decrease in gut microbiota richness and diversity, immune dysregulation, and delayed clearance of SARS-CoV-2.31 TMAO may serve as a potential biomarker for COVID-19, aiding in diagnosis, assessment of disease severity, and prognosis. Terruzzi et al noted that the gut microbiota-derived metabolites TMAO and lipopolysaccharides, which generate a pro-inflammatory microenvironment, may have a positive correlation with the severe risk of COVID-19.32 Marhuenda-Egea et al utilized 1H NMR to analyze urine samples from COVID-19 patients, revealing metabolic differences between COVID-19 patients and healthy controls. These differences primarily involve energy metabolism (such as glucose, ketone bodies, glycine, creatinine, and citrate), as well as processes related to the bacterial microbiota (such as TMAO and formic acid) and detoxification (such as hippuric acid).33 Isabel Tristán analyzed 80 serum and urine samples from COVID-19 patients (34 in intensive care and 46 hospitalized) and 32 healthy controls, identifying various metabolites, including TMAO, that could serve as biomarkers for the diagnosis, prognosis and severity assessment of COVID-19.34 Notably, the variation trend of TMAO among COVID-19 patients is not entirely consistent across all studies. In a nutshell, TMAO has shown promise within the gut-lung axis, warranting further exploration of its function (Figure 2).
 |
Figure 2 Application of TMAO in different body fluids as a potential biomarker for human diseases. TMAO levels have been related to various human diseases, such as pulmonary diseases (such as COVID-19), pancreatic diseases (such as diabetes mellitus), cardiovascular diseases, kidney diseases, and nervous system diseases, TMAO may serve as a potential biomarker for disease risk assessment and progression, underscoring its relevance in clinical settings. Created in BioRender. Ge, P. (2025) https://BioRender.com/f31k885. Abbreviations: TMAO, Trimethylamine N-Oxide; TMA, Trimethylamine; FMO, Flavin-containing monooxygenase. |
TMAO in Gut-Pancreas Axis
The pancreas has two main functions: endocrine and exocrine. The endocrine function is primarily responsible for producing and secreting hormones that regulate glucose homeostasis, while the exocrine function focuses on producing and secreting enzymes that facilitate digestion. Pancreatic tissue comprises various cellular and non-cellular components, forming a highly organized and complex microenvironment. When cells are stimulated, the dynamic balance of this microenvironment may be disrupted, leading to the development and progression of pancreatic diseases, such as diabetes and pancreatitis (Figure 2).
Diabetes mellitus (DM) is a class of metabolic diseases marked by elevated blood glucose levels. It is the third largest chronic disease in the world, second only to tumors and cardiovascular and cerebrovascular diseases. DM is categorized into T1DM, T2DM, gestational DM, and other special types of DM. Among them, T2DM is the most common type, accounting for more than 90% of DM patients.35 T2DM is mainly characterized by pancreatic β cell dysfunction, insulin resistance leading to glucose metabolism deficiency, and chronic low-grade inflammation.36 Lifestyle, environment, and genetics are closely related to the occurrence and development of T2DM.37 Additionally, diet and gut microbiota are also significantly associated with T2DM susceptibility.38 Accumulated clinical evidence supports a significant association between TMAO and DM (Table 1). A cross-sectional study reported that plasma TMAO levels in T2DM patients were higher than in controls.39–41 This concurs with the findings of a previous report.42 A long-term prospective cohort study involving 2088 participants found that higher serum TMAO levels were associated with an increased risk of T2DM and elevated fasting blood glucose.43 Another meta-analysis also reported that TMAO may be a novel biomarker for DM.44 However, a cohort study of 892 participants found that plasma TMAO levels were negatively associated with T2DM risk.45 Preclinical studies have shown a significant association between TMAO and the onset and progression of T2DM.46–48 Mechanistically, TMAO may induce T2DM by inducing pancreatic β-cell dysfunction, chronic inflammation, oxidative stress, and disrupting glucose and lipid homeostasis.36,49 The latest research indicates that TMAO can impair β-cell function and glucose tolerance, and the mechanism may be related to the promotion of β-cell ERS, dedifferentiation, and apoptosis.36
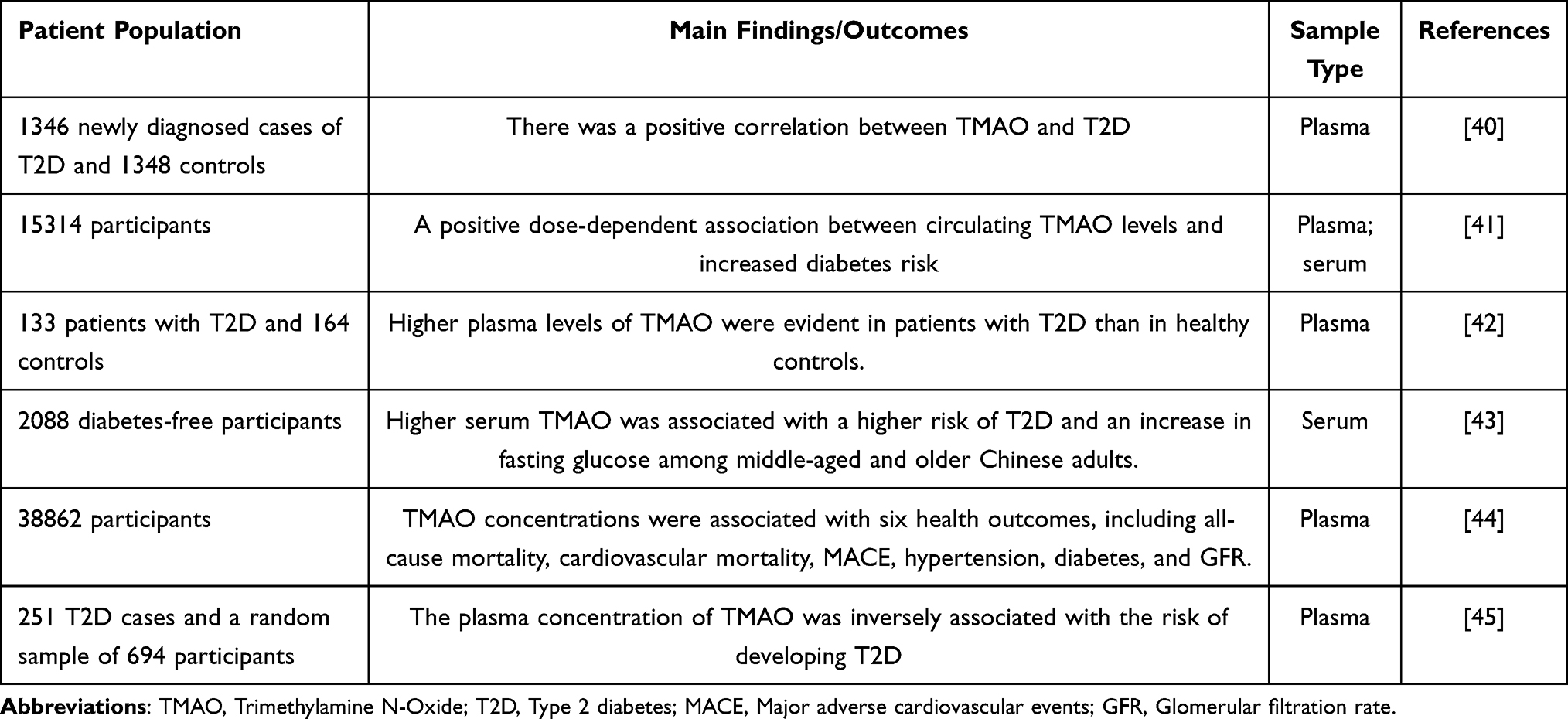 |
Table 1 The Effects of TMAO on Clinical Behavior and Prognosis in Diabetes |
In acute pancreatitis, the autodigestion of pancreatic acinar cells and the abnormal activation of pancreatic enzymes are the primary pathological features. Yang et al described the damaging effects of TMAO on pancreatic acinar cells.50 More and more evidence shows that the abnormalities in intercellular communication within the pancreatic microenvironment play a critical role in the pathophysiological response of the pancreas to various stimuli. TMAO may be a key mediator in the cross-talk between “endocrine cells and exocrine cells” in the pancreas.
TMAO in Gut-Heart Axis
Cardiovascular disease (CVD), including coronary atherosclerotic heart disease (CHD), heart failure (HF), arrhythmia, and cardiomyopathy, is a significant public health problem threatening human life and health. Despite the widespread use of secondary prevention drugs for CVD, the residual cardiovascular risk of patients remains high. Recent epidemiological evidence shows that the residual cardiovascular risk of CVD patients is closely linked to gut microbiota disturbances. Imbalances in the gut microbiota may regulate lipid metabolism by changing the level of bacterial metabolites and then induce the occurrence and development of CVD. In 2011, researchers using non-targeted metabolomics methods found that TMAO was associated with an increased risk of cardiovascular events in subjects.12 Different groups have demonstrated a strong association between TMAO and CVD and CVD-related risk factors in recent years. Hypertension is a major risk factor for CVD. Recently, the potential link between TMAO and Hypertension has garnered increasing attention. A Mendelian randomization study found a causal relationship between TMAO and its precursors and susceptibility to hypertension.51 A meta-analysis study reported that elevated TMAO levels were significantly associated with the risk of hypertension in a dose-dependent manner.52 Atrial fibrillation (AF) is not only a complication of various cardiovascular diseases but also significantly increases the risk of cardiovascular events. A growing number of preclinical and clinical studies have shown that TMAO is associated with AF53–56 (Table 2). Compared with AF patients without thrombosis, AF patients with thrombosis had significantly increased circulating TMAO levels. Mechanistically, TMAO may accelerate thrombosis by promoting platelet activation, inducing platelet hyperreactivity, and platelet aggregation.57,58 Several research groups have previously summarized the role of TMAO in the early diagnosis and prognosis assessment of atherosclerosis (AS),12,59–65 CHD,66–76 and HF.77–88 Therefore, we have generated Tables 2–4 to complement the latest research findings.
 |
Table 2 The Effects of TMAO on Clinical Behavior and Prognosis in Atrial Fibrillation and Atherosclerosis |
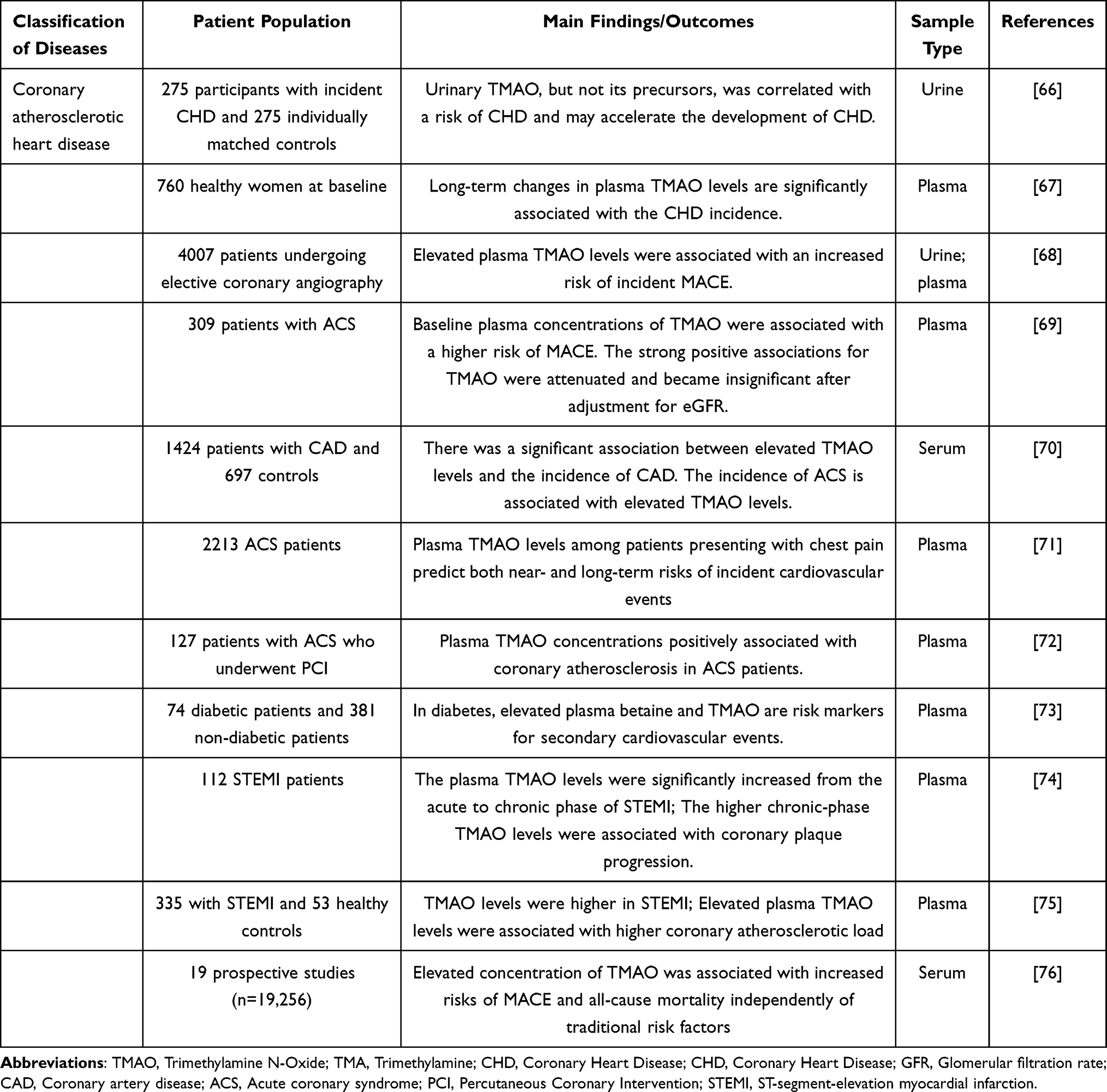 |
Table 3 The Effects of TMAO on Clinical Behavior and Prognosis in Coronary Atherosclerotic Heart Disease |
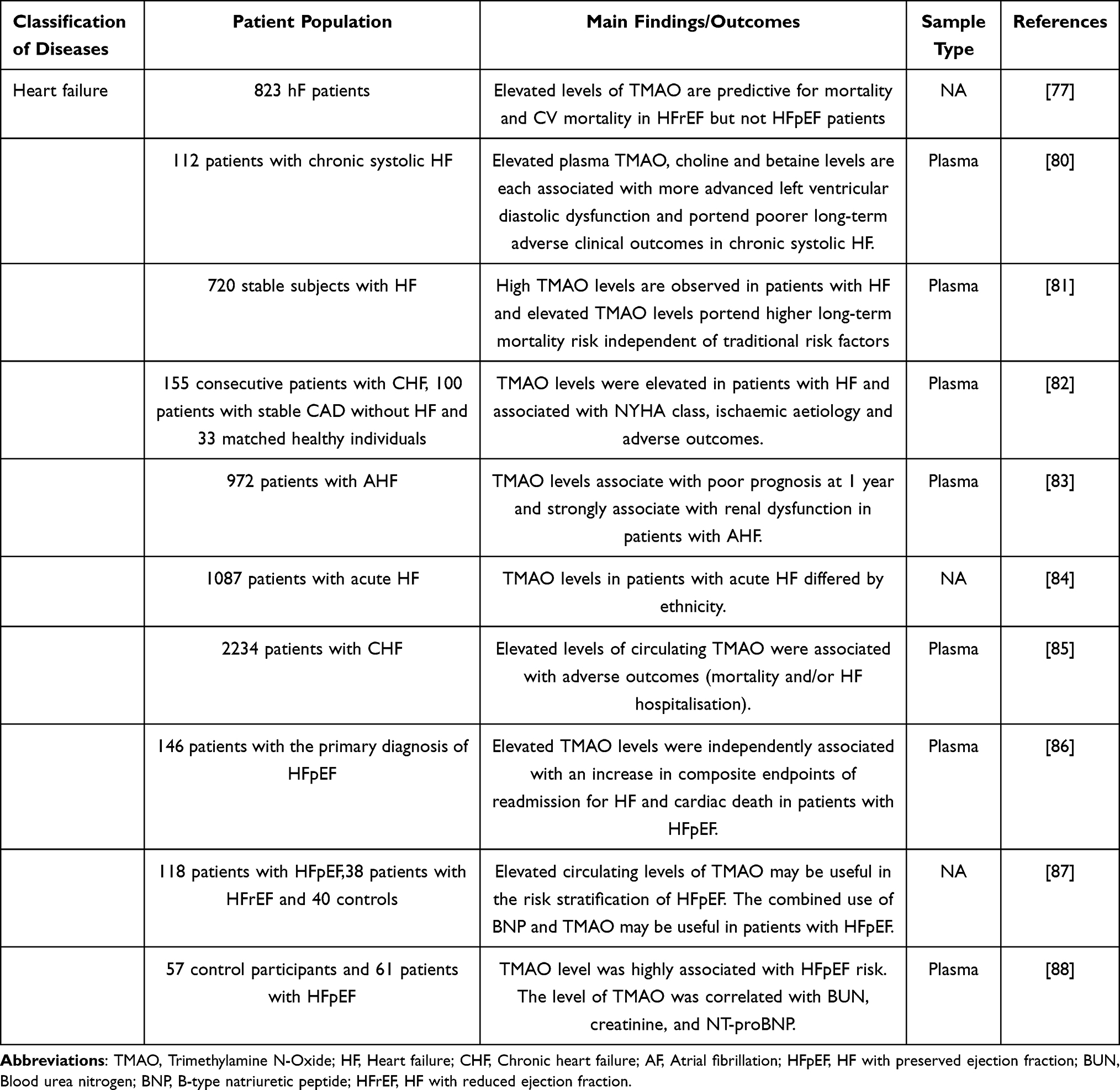 |
Table 4 The Effects of TMAO on Clinical Behavior and Prognosis in Heart Failure |
The value of TMAO in CVD is multifaceted. On the one hand, TMAO is a potential diagnostic marker for CVD, including atherosclerosis, coronary artery disease, and acute coronary syndrome. However, circulating TMAO levels are dynamically variable and are tightly regulated by diet, gut microbiome composition, liver enzyme activity, and renal excretion. Therefore, more sophisticated clinical studies are still needed to confirm the value of TMAO in diagnosing CVD. In addition, Additionally, given TMAO’s critical role in the development and progression of CVD, it is significantly associated with adverse events. Follow-up detection of circulating TMAO levels is helpful in screening out high-risk populations. However, existing clinical studies are mostly single-center and subject to racial limitations. Future studies should include larger and more ethnic groups of patients. Based on previous data, lowering plasma TMAO levels may reduce adverse outcomes of CVD. Targeting TMAO is a potential strategy for CVD therapy.
TMAO in Gut-Kidney Axis
Chronic kidney disease (CKD) is a progressive form of kidney structural damage and loss of function. The early clinical symptoms of CKD are not obvious, and the renal function gradually deteriorates in the later stage and eventually develops into uremia, that is, end-stage renal disease (ESRD). Advanced age, hypertension, dyslipidemia, and diabetes are recognized risk factors for CKD (Table 5). The “gut-kidney” axis plays a crucial role in the pathogenesis of kidney diseases.89 TMAO, as a metabolite derived from gut microbiota, may be a critical mediator in the “gut-kidney” axis.90 In recent years, increased TMAO levels have been found to be a novel risk factor and early diagnosis biomarker for CKD.91 Compared to healthy individuals, CKD patients’ TMAO levels significantly increased in blood and urine.92–94 TMAO levels are also significantly associated with several measures of renal function.95–98 Mechanistically, in the absence of impaired kidney function, more than 95% of TMAO in the human body is excreted through renal tubules or glomerular filtration,99,100 suggesting a potential mechanism for elevated circulating TMAO levels when renal function is impaired. GFR is an important indicator of renal function. Studies have found a strong negative correlation between plasma TMAO levels and measured GFR (mGFR) in patients with CKD.93,101 Stubbs et al also described a negative correlation between serum TMAO concentration and estimated GFR (eGFR).102 In patients with CKD, elevated TMAO levels accelerate the decline of renal function and affect long-term survival.94 Consistent with the above findings, Wang et al’s study also suggested that increased TMAO levels are a crucial risk factor for decreased renal function in CKD patients.103
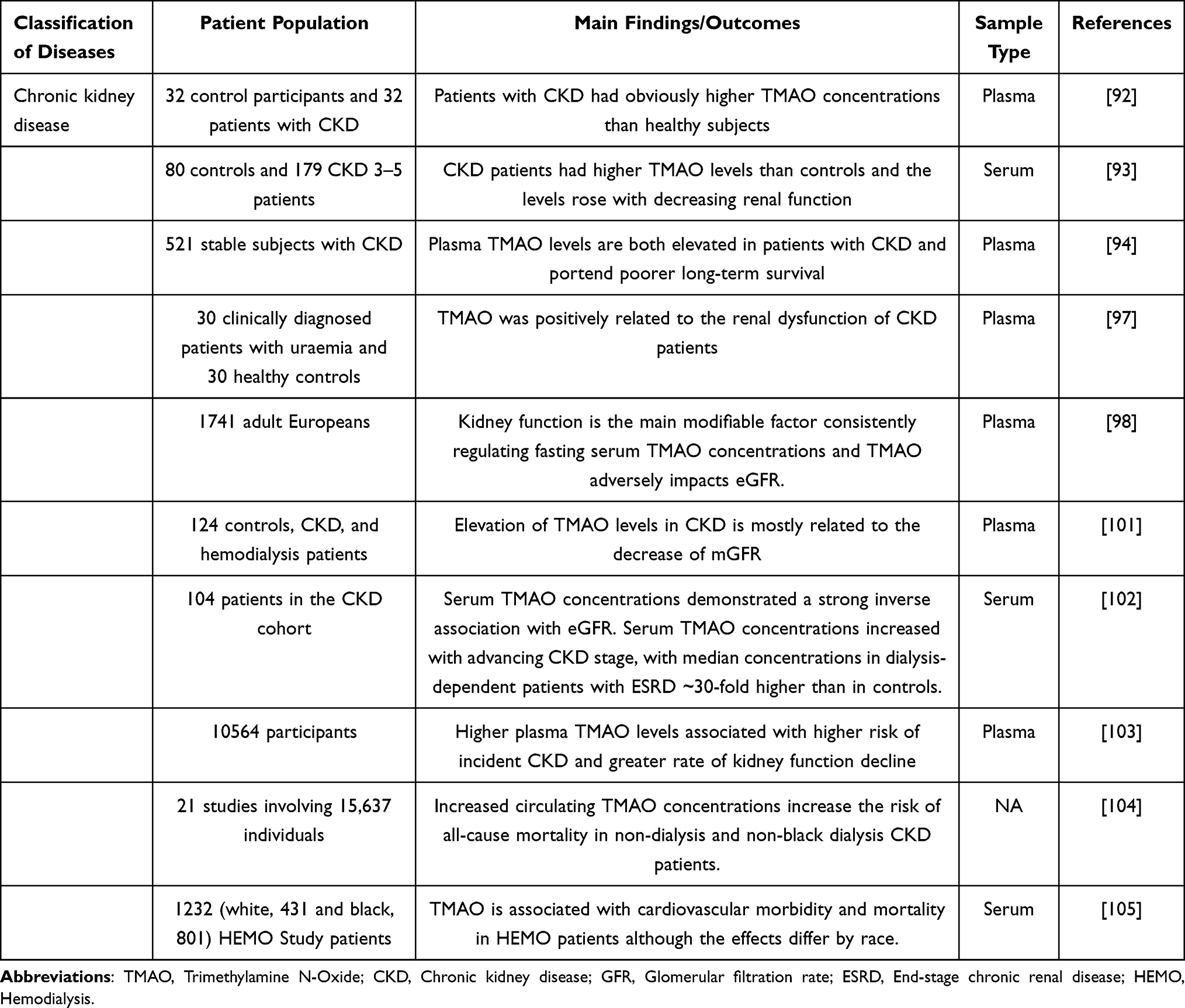 |
Table 5 The Effects of TMAO on Clinical Behavior and Prognosis in Kidney Disease |
It is well known that the incidence of adverse cardiovascular events increases with the progression of CKD.106 The deterioration of renal function is the main reason for accelerating the occurrence and development of adverse cardiovascular events in patients with CKD.107 TMAO is a potential risk factor for cardiovascular complications in patients with CKD.25,102 A meta-analysis reported that increased circulating TMAO levels were closely associated with an increased risk of all-cause mortality in patients with CKD.104 In CKD patients undergoing coronary angiography, TMAO is an independent predictor of coronary atherosclerotic burden and can also predict long-term mortality. A study of 1232 hemodialysis patients revealed no significant variation in TMAO levels between white and black individuals. Whites had 2-fold higher TMAO levels, which were strongly related to cardiac death, first cardiovascular event, and all-cause mortality. However, no significant association was seen among blacks.105 Mechanistically, on the one hand, TMAO may promote cardiac remodeling in hemodialysis patients by accelerating myocardial fibrosis and myocardial hypertrophy. On the other hand, TMAO may increase the incidence of adverse cardiovascular events in hemodialysis patients by increasing visceral fat.108 Notably, in patients undergoing peritoneal dialysis, researchers also identified a significant correlation between elevated serum TMAO levels and an increased risk of cardiovascular disease-related mortality. However, no significant association was found between TMAO level and mortality in peritoneal dialysis patients.109
Overall, there is a close association between TMAO and renal function, CKD, and cardiovascular outcomes in patients with CKD. Specifically, the levels of TMAO exhibit dynamic changes at different stages of CKD patients.97 In the early stages of CKD, circulating TMAO levels are relatively low because kidney function has not yet been significantly impaired. As CKD progresses, the TMAO levels in patients gradually increase. This may be related to decreased renal function, leading to reduced excretion of TMAO, along with alterations in the composition and metabolic function of the gut microbiota. In addition, elevated TMAO levels are closely associated with an increased risk of cardiovascular disease in CKD patients, possibly due to TMAO enhancing platelet reactivity and amplifying endothelial inflammatory responses, thereby increasing the risk of thrombosis. In conclusion, TMAO levels hold promise as a potential biomarker for predicting CKD progression and the risk of associated complications.
TMAO in Gut-Brain Axis
The “gut-brain” axis has progressively become a research hotspot. This bidirectional, stable axis connects the gut with the central nervous system. TMAO exists in human blood circulation and cerebrospinal fluid (CSF)110–112 and serves as a critical mediator in the “gut-brain” axis. Studies have found that high levels of TMAO have damaging effects on the neurovascular unit.113–115 “Neurovascular unit (NVU)” consists of three fundamental units: vascular cells, glial cells, and neurons, as well as the extracellular matrix in the cerebrovascular system. NVU emphasizes the interdependent symbiotic relationship between brain cells and cerebrovascular in terms of development, structure, and function, as well as the importance of cell-to-cell interactions in maintaining normal brain function and facilitating brain repair after injury. Dysfunction within NVU may lead to the onset and progression of various pathological reactions in the brain.116–118 Deng et al found that TMAO may induce neuroinflammation and neuronal apoptosis, leading to NVU dysfunction, which in turn exacerbates cognitive impairment and neuropathological changes in vascular dementia rats.119 Blood-brain barrier (BBB) disruption and white matter lesions are precursor events of neurological disorders. White matter primarily consists of myelinated nerve fibers and glial cells responsible for producing myelin sheaths. Oligodendrocytes are the cells in the central nervous system that form myelin sheaths. Myelin sheath surrounds myelin nerve fibers, acts as electrical insulation, and provides structural protection. It is crucial for maintaining physiological functions such as neuronal nutritional metabolism and information processing.120 Cell pyroptosis is also involved in oligodendrocyte damage and inflammatory demyelination.121,122 Ji et al found that oral administration of TMAO significantly exacerbated demyelination in spontaneously hypertensive rats, with the underlying mechanism possibly related to TMAO-induced pyroptosis of oligodendrocytes and inflammation.123 TMAO may influence the crosstalk of different cells in the NVU and participate in the occurrence and development of neurological disorders. This offers a potential direction for the study of the gut-brain axis in neurological diseases.
The potential of TMAO in the early diagnosis, disease monitoring, and prognosis assessment of neurological disorders is equally noteworthy. Plasma TMAO levels accumulate with age. As is well known, aging is a significant risk factor for cognitive deterioration. Therefore, Alzheimer’s disease (AD), characterized by cognitive impairment and memory loss, is inevitably influenced by the accumulation of TMAO. Moreover, TMAO can cross the BBB. Compared with healthy volunteers, CSF TMAO levels in AD patients were also significantly increased. Multiple clinical studies have repeatedly demonstrated a significant causal association between high plasma and CSF levels of TMAO and cognitive decline110,111,115,124–126 (Table 6).
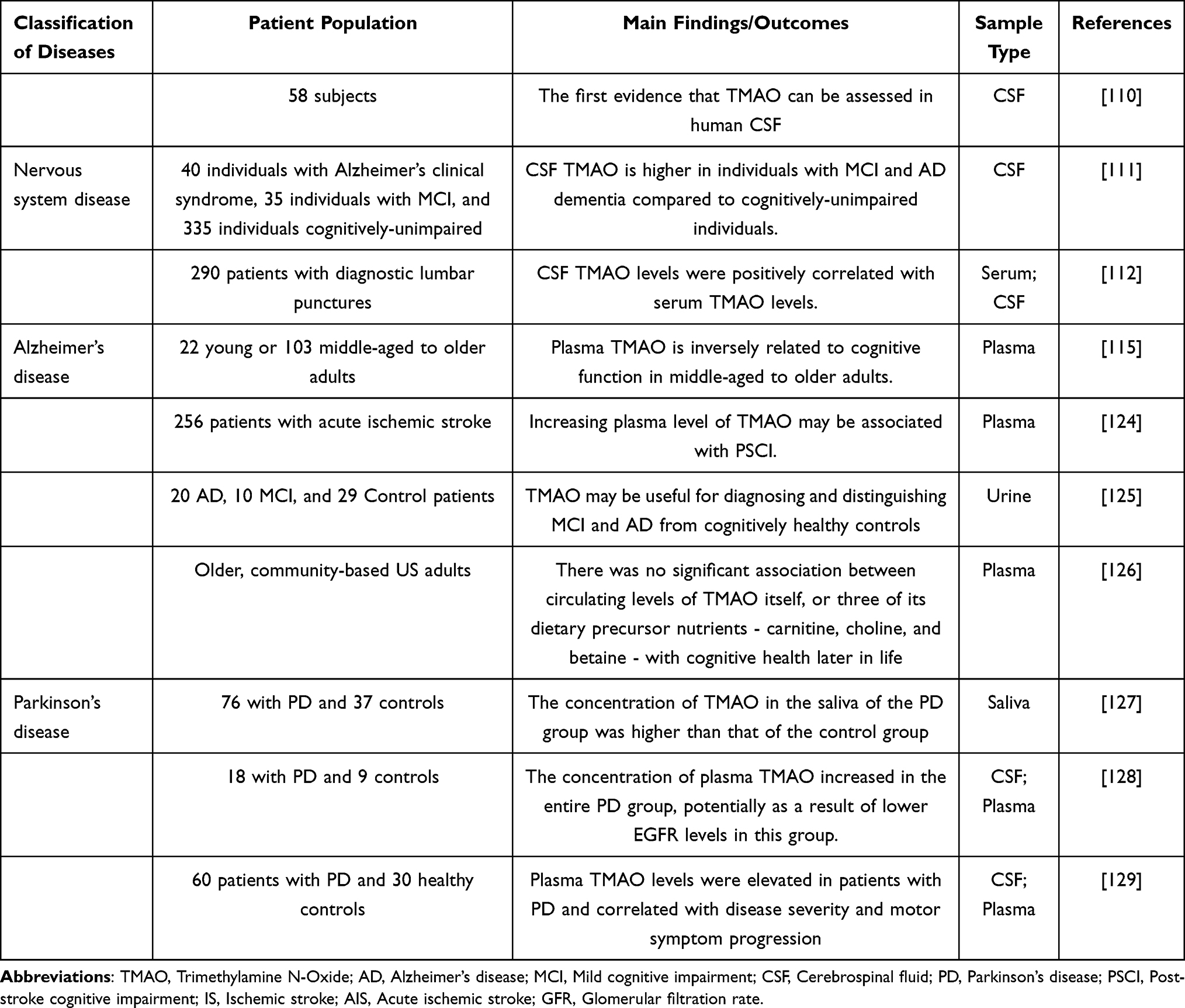 |
Table 6 The Effects of TMAO on Clinical Behavior and Prognosis in Nervous System Disease |
As a common neurodegenerative disease, the onset and progression of Parkinson’s disease (PD) are closely related to dysbiosis of the gut microbiota. Gut microbiota-related metabolites (especially TMAO) may be the key mediators that mediate the changes in brain structure and function of PD mediated by gut microbiota.130,131 On the one hand, TMAO is intimately linked to PD risk factors such as metabolic syndrome and cardiovascular disease.132,133 On the other hand, TMAO can directly promote alpha-synuclein folding and aggregation, neuroinflammation, mitochondrial dysfunction, and neuronal injury.134–136 Clinical studies have reported that compared with healthy controls, plasma, CSF, and saliva TMAO levels in PD patients are significantly elevated.127–129
Stroke seriously affects human health and quality of life.137 Among them, ischemic stroke (IS) is the most common type, mainly manifested as brain parenchyma injury, neuroinflammation, and immune response caused by middle cerebral artery occlusion in the affected area.138 IS is primarily associated with neuronal excitotoxicity, oxidative stress, neuroinflammation, apoptosis, mitochondrial damage, amyloid production, and tau protein dysfunction.139–143 Accumulated evidence emphasizes the critical role of gut microbiota and metabolic disorders in the occurrence, development, prognosis, and rehabilitation of stroke.144,145 Mechanistically, TMAO may mediate the occurrence and development of stroke by promoting endothelial dysfunction,7 inflammatory response,146 oxidative stress, and lipid metabolism disorders.147 Additionally, multiple clinical studies have reported the associations between plasma TMAO levels and their dynamic changes and stroke risk, severity, and long-term adverse outcomes.148–152 Additional studies have revealed a nonlinear dose-dependent relationship between circulating TMAO concentrations and stroke risk.153–155
Regulation Mechanism of TMAO in the “Gut-Organ” Axis: Crosstalk in Cell Death
Cell death is usually divided into two types: accidental cell death and regulated cell death (RCD). RCD includes apoptosis, regulatory necrosis, autophagy-dependent cell death, pyroptosis, ferroptosis, etc.
TMAO and Apoptosis
Apoptosis is a natural, non-inflammatory, programmed form of cell death. TMAO-induced apoptosis is closely related to ERS and mitochondrial dysfunction.156 The unfolded protein response (UPR) helps to re-establish endoplasmic reticulum (ER) homeostasis and promote cell survival. Moderate ERS can play a protective role through UPR. If ERS persists and does not alleviate, UPR will induce apoptosis.157 In the mitochondrial pathway, disruption of the outer mitochondrial membrane integrity can lead to mitochondrial outer membrane permeabilization (MOMP). MOMP plays a critical role in the mechanism of disease development by contributing to mitochondrial dysfunction. On the one hand, the occurrence of MOMP will lead to the release of apoptosis-related proteins into the cytoplasm, thus causing the opening of the mitochondrial permeability transition pore, which in turn results in the disruption of mitochondrial respiration, matrix swelling, and rupture of the mitochondrial outer membrane, thereby inducing apoptosis. On the other hand, cytochrome C released during MOMP can activate the caspase cascade and rapidly lead to apoptosis. Preclinical studies have demonstrated that TMAO can induce apoptosis in various target cells, such as human aortic vascular smooth muscle cells,158 pancreatic acinous cells, renal tubular epithelial cells,159 human aortic endothelial cells,160,161 and neurons,119 contributing to the development and progression of cardiovascular, renal, neurological, and pancreatic diseases.
Protein kinase RNA-like endoplasmic reticulum kinase (PERK) is a type I transmembrane protein located in the ER, consisting of an ER lumen domain and a cytoplasmic protein kinase domain. PERK-eukaryotic initiation factor 2 alpha (eIF2α), inositol-requiring enzyme 1 alpha (IRE1α)-X-box binding protein 1 (XBP1), and activating transcription factor 6 alpha (ATF6α) serve as key proteinS involved in ERS-related signaling pathways. Several studies have demonstrated that TMAO can stabilize protein metabolism, thereby reducing PERK activity and alleviating ER stress.162 TMAO may accelerate the progression or rupture of abdominal aortic aneurysm by inducing VSMC apoptosis through the activation of PERK-mediated UPR. Yang et al found that TMAO may promote pancreatic acinous cell apoptosis by inducing sustained oxidative stress and regulating the IRE1α/XBP-1 pathway. Additionally, TMAO may exacerbate the severity of CKD in rats by promoting renal tubular epithelial cell apoptosis through the activation of the ASK1-JNK pathway. Mitochondria are important components of cell metabolism, and the changes in mitochondrial outer membrane permeability are closely related to apoptosis. The accumulation of TMAO is a potential irritant to mitochondrial dysfunction. Studies have found that TMAO can significantly reduce the number of mitochondria, membrane potential, and ATP content of oocytes in polycystic ovary syndrome rat models, thus accelerating the apoptosis of oocytes.163 TMAO in circulation seems to never stop at all in its assault on vascular endothelial cells.164–167 In addition to direct damaging effects, TMAO-stimulated exosomes released by hepatocytes also play a crucial role in vascular inflammation and endothelial dysfunction.168 Liu et al demonstrated that exosomes released by TMAO-stimulated hepatocytes can promote HAEC apoptosis, inhibit cell migration, and endothelium-dependent vasodilation.160 It is noteworthy that TMAO’s role is not a one-way street. It also has an anti-apoptotic effect. In 2009, it was reported that TMAO degraded misfolded G98R mutant CRYAA by activating the ubiquitin-proteasome pathway, alleviating ERS and apoptosis of human lens epithelial B3 cells induced by aggregated G98R CrYAA.169 High concentrations of TMAO can promote the proliferation of human colon cancer cells, but have no significant effect on apoptosis.170 We speculate that the above differences are related to TMAO dosage and cell type heterogeneity, but there is currently no direct evidence to prove this. Figure 3 illustrates the regulatory pathways of TMAO-induced cell apoptosis in different cell types.
 |
Figure 3 Mechanisms of TMAO inducing cell Apoptosis. The mechanisms through which TMAO induces cell apoptosis via the endoplasmic reticulum (ER) and mitochondrial pathways. TMAO induces ER stress, activating the PERK/ATF-4 and IRE-1α/XBP-1s signaling pathways, leading to apoptosis, inflammation, and vascular injury. Additionally, TMAO-induced apoptosis may result from reduced SERCA activity and disrupted calcium homeostasis. It also promotes mitochondrial reactive oxygen species (mtROS) production and activates NLRP3 inflammasomes, which upregulate IL-1β and further drive apoptosis. TMAO triggers apoptosis by activating the ASK-JNK and SIRT1 pathways. Conversely, TMAO may inhibit apoptosis by mitigating aggregation and ER stress caused by G98R αA-crystallin. Created in BioRender. Ge, P. (2025) https://BioRender.com/t03q624. Abbreviations: TMAO, Trimethylamine-N-oxide; ERS, Endoplasmic reticulum stress; PERK, RNA-dependent protein kinase-like endoplasmic reticulum kinase; eIF2α, Eukaryotic translation initiation factor 2 subunit-α; ATF4, Activating transcription factor 4; CHOP, C/EBP homologous protein; IRE1α, Inositol-requiring enzyme 1α; XBP1, X-box binding protein 1; GRP78, Glucose-regulated protein 78; UPR, Unfolded protein response; Serca2, Sarco/endoplasmic reticulum ATPase type 2; ROS, Reactive oxygen species; NLRP3, Nod-like receptor family pyrin domain containing 3; ASC, Apoptosis-associated speck-like protein; IL-1β, Interleukin 1β; SIRT1, Sirtuin 1; JNK, C-Jun N-terminal kinase; ASK, Apoptosis signal-regulating kinase; MMP, Mitochondrial membrane permeabilization; LncRNA, Long non-coding RNA; NEAT1, Nuclear enriched abundant transcript 1; STAT3, Signal transducer and activator of transcription 3. |
TMAO and Autophagy
Autophagy is a cellular self-degradation process, which is an important process for cells to cope with nutritional stress. Under the regulation of autophagy-associated genes (ATG), misfolded or aggregated proteins and damaged organelles are removed and recycled by lysosomes or mitochondria to preserve cellular homeostasis.171,172 Moderate autophagy is a degradation system that maintains cellular homeostasis,173 and cells can achieve renewal and avoid death through basal levels of autophagy. However, during cell growth and development, autophagy acts as a double-edged sword. Insufficient autophagy may lead to the accumulation of toxic proteins and damaged organelles, affecting normal cell function.174 Conversely, excessive autophagy can result in the erroneous engulfment of essential proteins or normal organelles such as mitochondria.175 Insufficient and excessive autophagy are detrimental to cell viability and may cause cell apoptosis.176 In addition to non-selective autophagy induced under starvation conditions, researchers have found that cells can also target the degradation of damaged or redundant organelles, protein aggregates, and invading pathogens for selective autophagy. Selective autophagy includes mitophagy, endoplasmic reticulum autophagy, proteasome autophagy, ribosome autophagy, etc.177
Recent studies have found that the interactions between gut microbiota and their metabolites, especially TMAO, and autophagy not only regulate intestinal epithelial function and gut homeostasis but also play a role in regulating physiological functions in distant organs. Epithelial cells, ECs, and vascular smooth muscle cells (VSMCs) in multiple organ systems may be the primary targets of TMAO-induced autophagy-dependent cell death. In this process, mTOR and ATG16L1 may serve as vital regulatory factors. Kidney stone is a common urological disorder, which can cause renal insufficiency, atrophy kidney, and empyema. Calcium oxalate is the major component in the formation of kidney stones, causing tubular epithelial injury. A recent research discovered that serum TMAO levels were significantly higher in hyperoxaluria mice. Exogenous TMAO administration exacerbated kidney stones and inflammation in mice. Mechanistically, TMAO may promote autophagy-dependent cell death in human renal proximal tubular epithelial cells by activating the PERK/ROS pathway, thereby aggravating calcium oxalate crystal deposition and kidney injury.178 Additionally, Yue et al found that TMAO can inhibit autophagy in colonic epithelial cells, thereby exacerbating inflammatory bowel disease. The mechanism may be related to inhibiting ATG16L1-mediated autophagy of normal colon epithelial cells, promoting ROS accumulation and NLRP3 inflammasome assembly.179
TMAO mediates the onset and progression of CVD through its interaction with autophagy. Intervention in vascular remodeling is an important strategy to prevent and alleviate atherosclerosis.180 Vascular remodeling refers to a series of structural and functional abnormalities in the vascular wall, including the proliferation, migration, and apoptosis of cells within the vessel wall, and changes in the matrix components. The phenotypic transformation of VSMCs and vascular ECs plays a significant role in vascular remodeling. Early vascular remodeling is an adaptive protective mechanism, and vascular remodeling during decompensation can promote the progression of atherosclerosis and the occurrence of complications. Moderate activation of VSMC autophagy can significantly alleviate the atherosclerosis process induced by statins or cholesterol.181,182 PIK/AKT/mTOR signaling pathway is involved in the formation of autophagosomes.183 Studies have shown that TMAO can inhibit oxidative low-density lipoprotein (ox-LDL)-induced VSMC autophagy by activating the PIK/AKT/mTOR signaling pathway, thereby aggravating atherosclerosis.184 Besides VSMCs, the autophagy of ECs is also closely associated with the occurrence of atherosclerosis.185 Once ECs are damaged, various pathophysiological changes, such as thrombosis, inflammation, and excessive proliferation of smooth muscle cells, will be induced.186 Studies have shown that ox-LDL may exacerbate atherosclerosis by inhibiting HUVEC autophagy through the activation of the PI3K/AKT/mTOR signaling pathway.183 Hong et al found that TMAO may promote neointimal hyperplasia after balloon injury by activating Beclin1-mediated autophagy flow abnormality (the conversion of autophagosomes to autolysosomes).187 It should be emphasized that moderate autophagy is a positive process in combating atherosclerosis. However, excessive stress or overactive autophagy can still lead to autophagy-related cell death in VSMCs, negatively affecting the cells.188 In conclusion, the complex mechanisms involving TMAO, autophagy, and the crosstalk between ECs and VSMCs in the progression of atherosclerosis still require further research. The regulatory mechanisms of TMAO-induced autophagy in different cells are shown in Figure 4.
 |
Figure 4 Mechanisms of TMAO inducing autophagic cell death. TMAO upregulates Beclin1, a key regulator of autophagy, but this overexpression impairs autophagic flux, disrupting the normal degradation and recycling processes within the cell. Additionally, TMAO promotes autophagy inhibition by activating the PI3K/AKT/mTOR signaling pathway, a well-known negative regulator of autophagy. TMAO also triggers the PERK/ROS pathway, which paradoxically enhances autophagy, suggesting a complex role for TMAO in modulating autophagic processes. Furthermore, TMAO significantly suppresses the expression of ATG16L1, a critical component of autophagosome formation. This suppression prevents the proper multimerization of the ATG16L1-ATG5-ATG12 complex, further inhibiting autophagy and contributing to cellular dysfunction. These mechanisms highlight TMAO’s dual role in enhancing and impairing autophagy, ultimately leading to autophagic cell death. Created in BioRender. Ge, P. (2025) https://BioRender.com/i74e270. Abbreviations: TMAO, Trimethylamine-N-oxide; PI3K, Phosphoinositide 3-kinases; AKT, Protein kinase B; mTOR, Mammalian target of rapamycin; ERS, Endoplasmic reticulum stress; PERK, RNA-dependent protein kinase-like endoplasmic reticulum kinase; eIF2α, Eukaryotic translation initiation factor 2 subunit-α; ATF4, Activating transcription factor 4; LC3, Light chain 3; ROS, Reactive oxygen species; ATG, Autophagy-related gene. |
TMAO and Pyroptosis
ECs form the lumen of capillaries, arteries, and veins via tight junctions between cellular and matrix components. Originally considered an inactive resting layer, the endothelium disseminates throughout the circulatory system. ECs are crucial in the physiological regulation of vascular homeostasis. Pathophysiological processes such as vascular permeability, vasomotor tone, vascular growth, inflammation and immunology, coagulation homeostasis, and low-density lipoprotein oxidation are all under the control of the endothelium. ECs keep blood vessels in balance by releasing substances that narrow and widen blood vessels through autocrine, endocrine, and paracrine pathways. This includes the secretion of nitric oxide for vasodilation and endothelin-1 for vasoconstriction, which modulates vascular tension and diameter, directly influencing the balance of oxygen supply to meet tissue metabolic demands.189 The barrier function constitutes the fundamental role of ECs. The endothelium acts as a selective permeability barrier, controlling the steady movement of blood cell parts and different macromolecules across the vascular wall. This is made easier by the surface glycocalyx, intercellular junctions, and the basement membrane working together.190 EC damage or death is the main cause of illness. It can cause vasomotor imbalance, inflammatory responses, oxidative stress, and fat buildup, and it is strongly linked to the prevalence of diseases in many body systems.191 Researchers have recognized blood flow shear stress, cytokines, reactive oxygen species, pathogen-associated molecular patterns, and injury-associated molecular patterns as elements that trigger EC inflammation and dysfunction.192 Ongoing research on gut microbiota indicates that TMAO may play a crucial role in the stimulation of EC inflammation (Especially pyroptosis).
Pyroptosis is a pro-inflammatory programmed cell death that can be induced by exogenous microbial infection and endogenous injury-related signals. The key event in pyroptosis is the cleavage of gasdermin proteins by caspases. The gasdermin proteins share similar N-terminal and C-terminal domain structures, where the C-terminal domain acts as an inhibitory domain, restraining the activity of the N-terminal domain. However, not all gasdermin proteins can be cleaved by caspases. GSDMD and GSDME are the most extensively studied proteins in the gasdermin family. The linker region of GSDMD is cleaved by inflammatory caspases, releasing its active N-terminal domain. This N-terminal domain of GSDMD can be transported to the cell membrane, assembling into pore structures that release IL-1β and IL-18, leading to excessive inflammation and cell death.193 Additionally, caspase-3 cleaves GSDME at its linker region, releasing an N-terminal fragment that activates pyroptosis through a mechanism similar to that of the GSDMD N-terminal domain. Pyroptosis was initially discovered in immune cells, but subsequent studies have shown that other cell types, including epithelial and endothelial cells, can also undergo pyroptosis.194 Pyroptosis of renal parenchymal cells is considered a critical event exacerbating kidney injury. Yi et al found that TMAO aggravated high glucose-induced pyroptosis in human renal proximal tubular epithelial cells.195 Endothelial cell pyroptosis is a critical triggering event for vascular dysfunction.196,197 TMAO contributes to the onset and progression of atherosclerosis by inducing ECs pyroptosis. Mitochondria serve as the powerhouse and energy metabolism center of the cell, playing a pivotal role in sustaining cellular homeostasis and proper function. When mitochondrial homeostasis and integrity are compromised, it can lead to excessive production of ROS.198 ROS is a critical molecule that triggers pyroptosis.199 Succinate dehydrogenase complex subunit B (SDHB) is a subunit of the succinate dehydrogenase (SDH) family. A recent study showed that TMAO may induce atherosclerosis by upregulating the expression of SDHB in ECs, promoting the destruction of mitochondrial structure and function, and excessive production of ROS, accelerating the process of cell pyroptosis.164 Acetaldehyde dehydrogenase 2 (ALDH2) is a nuclear-encoded aldehyde oxidase present in the mitochondrial matrix.200 It has been found to play a protective role in various organ injuries.201–204 ALDH2 can inhibit the activation of NLRP3 inflammasome by reducing the excessive accumulation of mitochondrial ROS (mtROS) and total intracellular ROS, thereby mitigating high glucose-induced pyroptosis in cardiomyocytes. A study has found that TMAO may induce pyroptosis in HUVECs by inhibiting ALDH2 activity, which subsequently activates the ROS/NLRP3/GSDMD pathway.205 Recently, researchers have found a non-classical pathway of pyroptosis mediated by caspase-3 and gasdermin E (GSDME) in human primary cells and tumor cells.206–208 Although Caspase-3 is a pro-apoptotic caspase, existing evidence shows that after treatment with chemotherapy drugs, GSDME can be cleaved and activated by caspase-3, leading to pyroptosis.209,210 Another study reported that TMAO may induce endothelial progenitor cell pyroptosis through the Bax/caspase-3/GSDME pathway in vitro, thus participating in endothelial dysfunction. Figure 5 summarizes the regulatory mechanisms of TMAO-induced pyroptosis in various cells.
 |
Figure 5 Mechanisms of TMAO inducing cell pyroptosis. ROS plays a crucial role in activating the NLRP3 inflammasome, which subsequently triggers Caspase-1-dependent pyroptosis. TMAO can directly induces excessive ROS production and indirectly enhances ROS generation by activating the SDHB pathway and inhibiting the ADLH2 pathway, ultimately triggering cell pyroptosis. Furthermore, TMAO may also induce cell pyroptosis through the Bax/caspase-3/GSDME pathway, highlighting the multifaceted roles of TMAO in mediating inflammatory cell death. Created in BioRender. Ge, P. (2025) https://BioRender.com/i00w506. Abbreviations: TMAO, Trimethylamine-N-oxide; ROS, Reactive oxygen species; NLRP3, Nod-like receptor family pyrin domain containing 3; ASC, Apoptosis-associated speck-like protein; GSDMD, Gasdermin D; GSDME, Gasdermin E; NEK7, (NIMA)-related kinase 7. |
TMAO and Ferroptosis
Ferroptosis is a form of PCD characterized by iron dependency and lipid peroxidation, which primarily involves three pathways: iron metabolism, lipid metabolism, and antioxidant system. Iron and oxygen are fundamental drivers of metabolism. Iron metabolism disorders trigger the Fenton reaction and induce ROS production, an inevitable byproduct. The sustained accumulation of ROS and the ROS-induced buildup of lipid peroxides further compromises the integrity of the plasma membrane, and ultimately, the cell will die due to ferroptosis. Therefore, Ferroptosis can be considered as a byproduct of cellular metabolism. Currently, evidence directly linking TMAO to ferroptosis is limited. Wang et al found that in diabetic myocardial ischemia-reperfusion injury (DIR) rats, TMAO promoted the ferroptosis of DIR cardiocytes by up-regulating the expression of ALB, PPARG and HMOX1 (ferroptosis related genes).211 TMAO is closely related to excessive oxidative stress within cells. Luo et al found that TMAO may inhibit the expression of nuclear factor E2-related factor 2 (Nrf2), HO-1, and glutathione peroxidase 4 (GPX4), resulting in the overproduction of ROS and the reduction of SOD activity, thus promoting oxidative stress.212 Similarly, Cristina Gonzalez-Correa et al found that in a mouse model of systemic lupus erythematosus, TMAO may induce elevated NADPH oxidase activity and vascular endothelial oxidative stress by inhibiting Nrf2-mediated antioxidant defense mechanism.213 GPX4 is a negative ferroptosis regulator, playing a critical role in converting lipid hydroperoxides to non-toxic lipids.214 In CKD, TMAO may induce ferroptosis-related changes in renal tubular epithelial cells, including increased MDA and iron levels, excessive ROS accumulation, reduced GSH, and decreased GPX4 protein levels.215 Nrf2 is a key transcription factor that regulates cellular redox homeostasis and inflammatory response. In many chronic diseases, activation of Nrf2 signaling can also protect cells from ferroptosis.216 Additionally, levels of the anabolic reductant NADPH are biomarkers of ferroptosis sensitivity.217 It can be seen that TMAO may participate in iron death by regulating the expression of ferroptosis-related genes and inducing oxidative stress (Figure 6).
 |
Figure 6 Mechanisms of TMAO inducing cell ferroptosis. TMAO promotes ferroptosis through two major pathways: Modulation of the GSH/GPX4 axis–TMAO inhibits system Xc−, leading to reduced GSH synthesis and impaired GPX4 activity. Dysregulation of iron metabolism–TMAO alters the expression of key iron-related proteins (eg, TRF-1, FPN-1, and FTH-1) and suppresses the NRF2 signaling pathway, exacerbating iron accumulation and lipid peroxidation. Created in BioRender. Ge, P. (2025) https://BioRender.com/upipqo4. Abbreviations: TMAO, Trimethylamine-N-oxide; SLC7A11, solute carrier family 7 member 11; GSH, glutathione; GPX4, Glutathione peroxidase 4; GSSG, Glutathione disulfide; TFR1, Transferrin receptor protein 1; TF, Transferrin; HO-1, Haem oxygenase-1; NRF2, Nuclear factor-erythroid 2-related factor 2; FPN, Ferroportin; FTH1, Ferritin heavy chain 1; ROS, Reactive oxygen species; STEAP3, Six-transmembrane epithelial antigen of the prostate 3; DMT1, Divalent metal transporter 1; NCOA4, nuclear receptor coactivator 4. |
In conclusion, within the “gut-organ” axis, TMAO is closely associated with the pathological changes of various chronic diseases by regulating processes such as apoptosis, pyroptosis, abnormal autophagy, and ferroptosis. Although current research has unveiled multiple mechanisms by which TMAO affects cellular function, its specific roles in different tissues and pathological conditions require further investigation.
Diverse Therapeutic Strategies Targeting TMAO
Probiotics, Prebiotics, and Antibiotics
The role of probiotics in reducing TMAO levels has been in the spotlight, shining brightly over the past decade.218 Currently, the most common probiotics include Lactobacillus and Bifidobacterium. Lactobacillus and Bifidobacterium exhibit anti-inflammatory, antioxidant, anti-thrombotic, and endothelial protective effects.219 They can prevent and cure illnesses, such as atherosclerosis,220 cardiovascular diseases,221 kidney diseases,222 and metabolic syndrome.223 Existing studies have indicated that probiotics, including Bifidobacterium longum, Lactobacillus plantarum, Lactobacillus amylovorus, Enterobacter aerogenes, Lactobacillus rhamnosus, Lactobacillus acidophilus, Bifidobacterium lactis, Bifidobacterium breve, Bifidobacterium animalis, and Lactobacillus casei, have a certain downregulatory effect on circulating TMAO levels. Table 7 shows the direct or indirect regulatory effects of probiotics on circulating TMAO level.222,224–233 Notably, probiotics can be utilized to modify the gut microbiota targeting TMAO, but the impact of different strains on TMAO levels varies. The effect of probiotics on TMAO is strain-dependent, and the specific outcome is determined by the influence of the particular strain on TMAO metabolism, and not all probiotics exhibit this effect. Therefore, further large-scale clinical studies are needed to provide strong evidence that specific probiotic strains can target TMAO metabolism and improve disease outcomes.
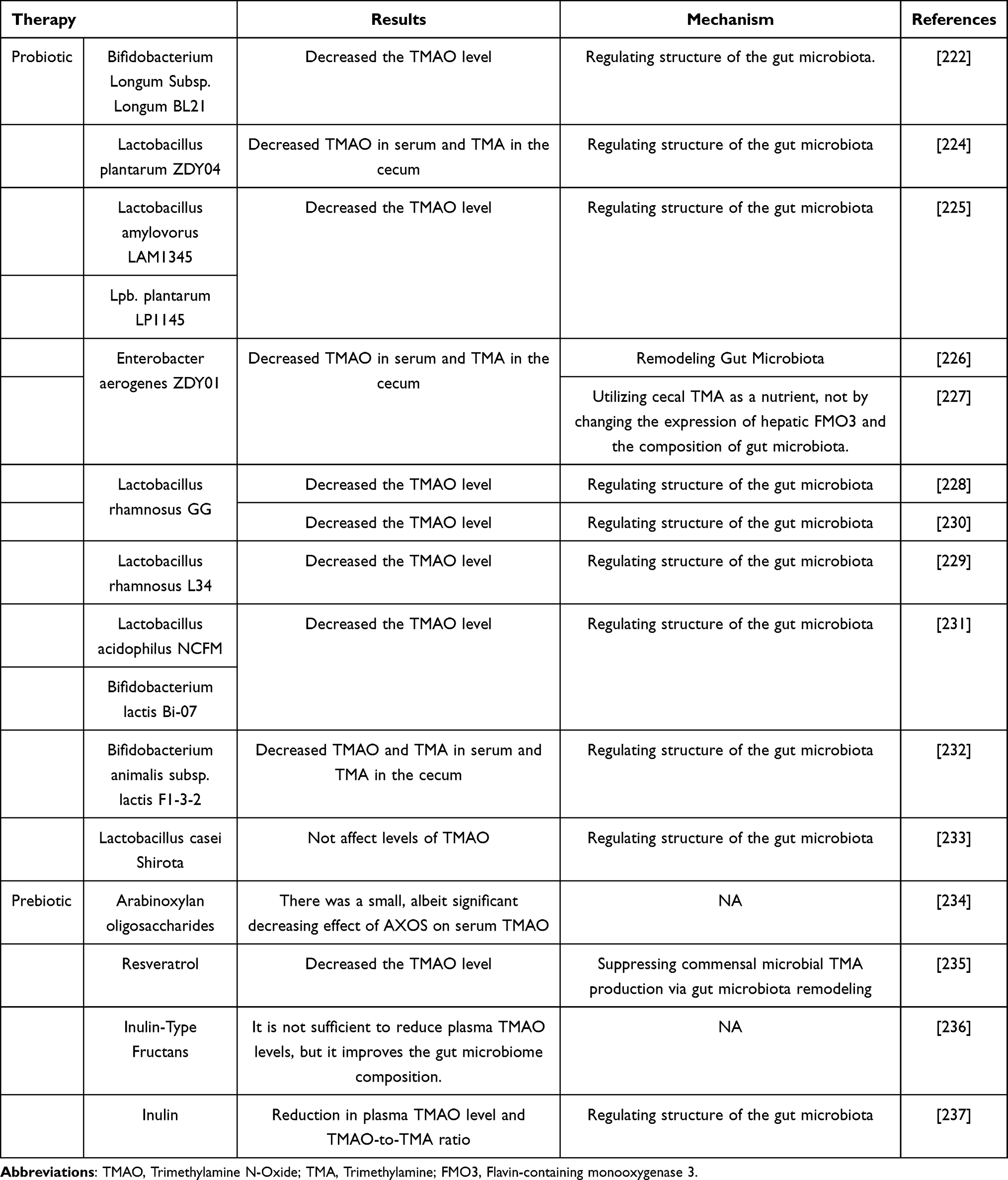 |
Table 7 The Regulatory Effects of Probiotics on Circulating TMAO Level |
Prebiotics are a class of non-digestible food ingredients that exerting positive effects on host health.238 Prebiotic compounds include oligosaccharides, fructooligosaccharides, galactooligosaccharides, beta-glucans, and inulin. Recent research has also revealed that prebiotics may have the potential to regulate TMAO levels, thus improving related diseases234–237 (Table 7).
Probiotics and prebiotics have shown great potential in targeting TMAO production. Nevertheless, this field continues to encounter numerous challenges, such as the need to identify safe and effective probiotic strains to ensure that they can significantly reduce TMAO levels. Concurrently, it is also necessary to determine the optimal dosage, duration, and frequency of probiotic and prebiotic administration.
Antibiotics are the most potent agents for inhibiting the production of TMAO. In pediatric patients with trimethylaminuria (FMO3 deficiency), the use of metronidazole, amoxicillin, and neomycin can partially inhibit the conversion of choline to TMA, with neomycin being the most effective, followed by amoxicillin, and metronidazole being the weakest effective among them.239 It is worth noting that using antibiotics to target TMAO for disease treatment belongs to a strategy of “Pyrrhic victory.” Briefly, the use of broad-spectrum antibiotics such as ciprofloxacin and metronidazole almost completely suppress TMAO levels. However, once antibiotics are discontinued, TMAO levels rise again within a month. The combination of vancomycin, neomycin sulfate, metronidazole, ampicillin, and other broad-spectrum antibiotics can alleviate choline-induced atherosclerosis in mice, and the mechanism may be related to the inhibition of choline conversion to TMA, the reduction of TMAO levels, and the inhibition of macrophage foaming. However, long-term use of antibiotics may lead to the emergence of drug-resistant strains and the depletion of beneficial bacteria.12 Hence, the rational use of antibiotics, combined with probiotics and prebiotics, or the exploitation of new intervention strategies, should be important directions for future research.
Clinical Compounds
Antidiabetic drugs (such as metformin and Empagliflozin), antiplatelet drugs (such as aspirin and clopidogrel), as well as statins (such as atorvastatin and rosuvastatin) have been shown to target TMAO to exert therapeutic effects. Metformin has been shown to reduce TMA and TMAO levels by remodeling gut microbiota.240 Notably, empagliflozin, unexpectedly gives rise to a rapid and significant elevate in TMAO levels in patients with AMI.241 Low-dose aspirin mitigates platelet hyperreactivity linked to high TMAO and may decrease TMAO’s dependence on choline.57 Clopidogrel resistance is associated with elevated TMAO levels, as shown by Ge et al, who found that dietary choline and TMAO inhibit the metabolic activation of clopidogrel via the NOX/ROS/Nrf2/CES1 pathway.242 Statins not only significantly improve blood lipid levels but also influence gut microbiota and the production of its metabolite, TMAO. A meta-analysis involving 244 subjects showed a significant reduction in plasma TMAO levels after statin treatment.242 Two independent cohort studies found that patients with dyslipidemia experienced a decrease in plasma TMAO levels after initiating statin treatment, but TMAO levels returned to baseline after a four-week discontinuation of the medication.243 In patients with atherosclerotic cardiovascular disease, rosuvastatin not only significantly reduced blood lipids but also decreased TMAO levels. Additionally, research also observed a significant elevation in the levels of TMAO precursors during rosuvastatin treatment.244 Therefore, a complex regulatory mechanism may underlie the modulation of TMAO metabolism by statins, necessitating further research to clarify the precise pathways involved and their clinical implications (Table 8).
 |
Table 8 Other Drugs Regulating TMAO Levels and Their Mechanisms |
Natural Products
Natural products, including alkaloids, polyphenols, and flavonoids, exhibit various biological activities, such as anti-inflammatory, antitumor, and anti-oxidative stress effects, providing highly valuable lead compounds for new drug research and development. Studies indicate that the bioactivity of natural products may be closely related to their biotransformation by the gut microbiota. Additionally, compared to antibiotics and other drugs, natural products not only inhibit harmful bacteria but also enhance the abundance of certain beneficial bacteria, thereby improving disease conditions. In this section, we summarize the research progress on various natural products that regulate gut microbiota composition and structure, reduce TMAO levels, and ultimately improve disease outcomes (Table 8).
Alkaloids are nitrogenous secondary metabolites found in plants, exhibiting diverse biological activity. Berberine may reduce the risk of thrombosis and improve atherosclerosis by modulating the gut microbiota, inhibiting CutC enzyme activity and decreasing TMAO production.245–247 Additionally, it protects hypertensive mice by remodeling gut microbiota and preventing TMAO-induced endothelial dysfunction.161 Trigonelline may exert anti-atherosclerotic effects by inhibiting Citrobacter freundii and the activity of FMO3 and blocking the choline-TMA-TMAO metabolic pathway.248 Curcumin (Cur) is a natural polyphenolic compound derived from the rhizome of plants such as Curcuma longa L. (turmeric), which has anti-inflammatory, antioxidant, anti-apoptosis, and anti-cancer effects.257 The concentration of Cur in plasma is relatively low, it is significantly higher in the gut.258 Studies have shown that curcumin can reduce TMAO production by regulate the abundance, diversity, and composition of gut microbiota, thereby regulating macrophage polarization and mitigating cadmium-induced atherosclerosis in mice.250 Docosahexaenoic acid is a n-3 long-chain polyunsaturated fatty acid widely found in Marine animals. Shi and his team recombined DHA and Cur, and demonstrated that DHA-acylated Cur diesters significantly improved cisplatin-induced acute kidney injury in mice compared to a single DHA or Cur treatment group. The mechanism is linked to the regulation of gut microbiota and inhibition of LPS and TMAO-induced PI3K/AKT/NF-κB signaling pathways.251 Additionally, Resveratrol (RSV), Polyphenol extracts of hickory nut and Mangiferin can regulate the diversity of gut microbiota, promote the metabolism of TMA, decrease TMAO levels, thus alleviates TMAO-induced atherosclerosis.249,259 Taurine is a natural sulfur-containing amino acid. Numerous preclinical studies have reported the pharmacological activities of taurine, such as anti-inflammatory, anti-oxidative stress, anti-depression, hepatoprotective, and anti-tumor. Supplementing with taurine can help prevent aging, mitochondrial diseases, metabolic syndrome, cancer, cardiovascular diseases, and neurological disorders. Yang et al showed that taurine may alleviate atherosclerosis by inhibiting TMAO. Specifically, taurine significantly reduced TMAO-induced atherosclerosis lesions and chronic inflammation in mice, and the mechanism may be related to increasing the ratio of conjugated bile acids to unconjugated bile acids in the serum.253
Satheesh et al reported that the effects of a moderate blueberry (rather than strawberry) diet on TMAO were not mediated by metabolic changes (TMAO levels in the circulation can be reduced without changing choline or TMA). The sequencing results showed that blueberry significantly increased the abundance of bacterial taxa negatively correlated with circulating TMA/TMAO, suggesting that it may affect TMAO synthesis by affecting the composition of gut microbiota.252 Further analysis of freeze-dried blueberries and strawberries revealed a significant increase in phenolic compounds, particularly chlorogenic acid, which may be a critical factor in blueberries’ inhibition of TMAO production. Asparagus contains various phytochemicals, including polyphenols, saponins, asparagusic acid, and alkaloids, which have anti-inflammatory, antibacterial, antioxidant, neuroprotective, anti-anxiety and anti-asthmatic properties. In vitro experiments have demonstrated that asparagus extract can alleviate TMAO-induced endothelial dysfunction, and the mechanism may be related to improving the TMAO-induced decrease in EC proliferation and regulating the molecular circadian clock controlled by NEAT1 and MAPK signaling pathways.254 Other natural plants or plant-derived chemical compounds have likewise been demonstrated to reduce plasma TMAO levels and alleviate TMAO-induced tissue and organ damage. Commelina communis L., a plant widely distributed in tropical regions, is used in its dried aerial part known as Yazhicao (Commelina communis L., YZC). YZC has been shown to improve metabolic function, reshape and regulate gut microbiota, reduce TMAO synthesis, and inhibit NLRP3 inflammasome activation, thereby ameliorating LPS-induced lung inflammation and injury in septic ALI mice.28 In addition, Decaisnea insignis seed oil mitigated liver dysfunction in L-carnitine-fed mice, potentially by modulating the composition of the gut microbiota, reducing the abundance of Firmicutes, Proteobacteria, and Erysipelotrichaceae, as well as the proportions of Lactobacillus and Akkermansia, thereby preventing TMAO generation.255 Similarly, Sandalwood seed oil has been shown to improve insulin resistance in rats induced by a high fat/high sugar diet, with mechanisms related to changes in gut microbiota composition and the production of microbial metabolites (including TMAO).256
Conclusion and Future Directions
TMAO may drive the cross-talk between inflammatory response and oxidative stress by mediating cell “death” (including apoptosis, pyroptosis, autophagy, and ferroptosis, etc) and then induce pathological processes such as foam cell activation, massive secretion of cytokines and adhesion molecules, overaccumulation of ROS, enhanced platelet reactivity and abnormal vascular tone regulation. It is deeply involved in the pathogenesis and progression of diseases across the “gut-organ” axis pulmonary diseases, including cardiovascular diseases (such as atherosclerosis, heart failure, and hypertension), renal diseases, neurodegenerative disorders, and metabolic diseases, including diabetes. As a risk factor for various chronic diseases, the detection of TMAO and related metabolites can provide a potential basis for the prevention, diagnosis, and treatment of cardiovascular diseases, kidney diseases, neurological diseases, and metabolic diseases. We recommend incorporating TMAO and its related metabolites into routine health examinations. Additionally, this paper also summarized a variety of measures to intervene in related diseases by targeting the reduction of TMAO levels, including probiotics, prebiotics, antibiotics, anti-inflammatory drugs, antiplatelet agents, hypoglycemic agents, lipid-lowering drugs, Chinese herbal monomers, herbal extracts, and formulas. The above strategies may lower the circulating TMAO levels by targeting gut microbiota, inhibiting the activity of TMA/TMAO production and conversion-related enzymes, and intervening in downstream effects, thereby providing a theoretical basis for the clinical treatment of TMAO-related diseases.
Notably, there are still many limitations in current research on TMAO. On the one hand, TMAO levels are dynamically changing in human circulation (this physiological concentration range is also controversial) and are strictly regulated by factors such as age, diet, gut microbiota composition, hepatic enzyme activity, as well as liver and kidney function. In the early diagnosis and prognosis evaluation of various human diseases, the specificity and sensitivity of TMAO have constantly been challenged. Moreover, different tissues and organs have varying tolerance thresholds for TMAO, and further clarification is still required on whether the dynamic fluctuations of TMAO exert beneficial roles or detrimental effects in human organs. Second, although numerous observational studies have shown causal associations between TMAO and cardiovascular, kidney, neurological, and metabolic diseases, due to the inherent limitations of statistical methods (bias due to confounding factors and reverse causality), and the causal relationship between TMAO and human diseases still requires validation through multicenter, multi-ethnic randomized controlled trials. Finally, the pathological process of endothelial cell dysfunction induced by TMAO may be regulated by various forms of cell death, yet the specific mechanism of this process is still unclear. To provide a more thorough theoretical foundation for treating disorders associated to TMAO, larger-scale clinical trials are needed to validate TMAO’s potential as a biomarker for various diseases along the “gut-organ” axis. In-depth mechanistic studies are also required to fully understand the interactions between TMAO, cell death, and the “gut-organ” axis.
Funding
This study was supported by the National Natural Science Foundation of China (NO. 82274311, 82074158, and 82304943) and 2023 Dalian Life and Health field guidance Plan (2023C027).
Disclosure
The authors declare no conflicts of interest.
References
1. Connell E, Le Gall G, Pontifex MG, et al. Microbial-derived metabolites as a risk factor of age-related cognitive decline and dementia. Mol Neurodegener. 2022;17(1):43. doi:10.1186/s13024-022-00548-6
2. Steiber A, Kerner J, Hoppel CL. Carnitine: a nutritional, biosynthetic, and functional perspective. Mol Aspects Med. 2004;25(5–6):455–473. doi:10.1016/j.mam.2004.06.006
3. Böttcher C, von Roepenack-Lahaye E, Schmidt J, et al. Analysis of phenolic choline esters from seeds of Arabidopsis thaliana and Brassica napus by capillary liquid chromatography/electrospray- tandem mass spectrometry. J Mass Spectrom. 2009;44(4):466–476. doi:10.1002/jms.1522
4. Chen H, Peng L, Pérez de Nanclares M, et al. Identification of sinapine-derived choline from a rapeseed diet as a source of serum trimethylamine N-oxide in pigs. J Agric Food Chem. 2019;67(27):7748–7754. doi:10.1021/acs.jafc.9b02950
5. Fennema D, Phillips IR, Shephard EA. Trimethylamine and trimethylamine N-Oxide, a Flavin-Containing Monooxygenase 3 (FMO3)-mediated host-microbiome metabolic axis implicated in health and disease. Drug Metab Dispos. 2016;44(11):1839–1850. doi:10.1124/dmd.116.070615
6. Zeisel SH, Warrier M. Trimethylamine N-Oxide, the microbiome, and heart and kidney disease. Annu Rev Nutr. 2017;37:157–181. doi:10.1146/annurev-nutr-071816-064732
7. Shanmugham M, Bellanger S, Leo CH. Gut-derived metabolite, Trimethylamine-N-Oxide (TMAO) in cardio-metabolic diseases: detection, mechanism, and potential therapeutics. Pharmaceuticals. 2023;16(4):504. doi:10.3390/ph16040504
8. Querio G, Antoniotti S, Geddo F, et al. Modulation of endothelial function by TMAO, a gut microbiota-derived metabolite. Int J Mol Sci. 2023;24(6):5806. doi:10.3390/ijms24065806
9. Galluzzi L, Vitale I, Aaronson SA, et al. Molecular mechanisms of cell death: recommendations of the nomenclature committee on cell death 2018. Cell Death Differ. 2018;25(3):486–541. doi:10.1038/s41418-017-0012-4
10. Thomas MS, Fernandez ML. Trimethylamine N-Oxide (TMAO), diet and cardiovascular disease. Curr Atheroscler Rep. 2021;23(4):12. doi:10.1007/s11883-021-00910-x
11. Koeth RA, Wang Z, Levison BS, et al. Intestinal microbiota metabolism of L-carnitine, a nutrient in red meat, promotes atherosclerosis. Nat Med. 2013;19(5):576–585. doi:10.1038/nm.3145
12. Wang Z, Klipfell E, Bennett BJ, et al. Gut flora metabolism of phosphatidylcholine promotes cardiovascular disease. Nature. 2011;472(7341):57–63. doi:10.1038/nature09922
13. Simó C, García-Cañas V. Dietary bioactive ingredients to modulate the gut microbiota-derived metabolite TMAO. New opportunities for functional food development. Food Funct. 2020;11(8):6745–6776. doi:10.1039/d0fo01237h
14. Zhu Y, Jameson E, Crosatti M, et al. Carnitine metabolism to trimethylamine by an unusual Rieske-type oxygenase from human microbiota. Proc Natl Acad Sci U S A. 2014;111(11):4268–4273. doi:10.1073/pnas.1316569111
15. Koeth RA, Levison BS, Culley MK, et al. γ-Butyrobetaine is a proatherogenic intermediate in gut microbial metabolism of L-carnitine to TMAO. Cell Metab. 2014;20(5):799–812. doi:10.1016/j.cmet.2014.10.006
16. Rath S, Heidrich B, Pieper DH, et al. Uncovering the trimethylamine-producing bacteria of the human gut microbiota. Microbiome. 2017;5(1):54. doi:10.1186/s40168-017-0271-9
17. Romano KA, Vivas EI, Amador-Noguez D, et al. Intestinal microbiota composition modulates choline bioavailability from diet and accumulation of the proatherogenic metabolite Trimethylamine-N-Oxide. mBio. 2015;6(2):e02481. doi:10.1128/mBio.02481-14
18. Arias N, Arboleya S, Allison J, et al. The relationship between choline bioavailability from diet, intestinal microbiota composition, and its modulation of human diseases. Nutrients. 2020;12(8):2340. doi:10.3390/nu12082340
19. Ley RE. Gut microbiota in 2015: prevotella in the gut: choose carefully. Nat Rev Gastroenterol Hepatol. 2016;13(2):69–70. doi:10.1038/nrgastro.2016.4
20. Craciun S, Balskus EP. Microbial conversion of choline to trimethylamine requires a glycyl radical enzyme. Proc Natl Acad Sci U S A. 2012;109(52):21307–21312. doi:10.1073/pnas.1215689109
21. Velasquez MT, Ramezani A, Manal A, et al. Trimethylamine N-Oxide: the good, the bad and the unknown. Toxins. 2016;8(11):326. doi:10.3390/toxins8110326
22. Catucci G, Querio G, Sadeghi SJ, et al. Enzymatically produced trimethylamine N-Oxide: conserving it or eliminating it. Catalysts. 2019;9(12):1028. doi:10.3390/catal9121028
23. Teft WA, Morse BL, Leake BF, et al. Identification and characterization of trimethylamine-N-oxide uptake and efflux transporters. Mol Pharm. 2017;14(1):310–318. doi:10.1021/acs.molpharmaceut.6b00937
24. Saaoud F, Liu L, Xu K, et al. Aorta- and liver-generated TMAO enhances trained immunity for increased inflammation via ER stress/mitochondrial ROS/glycolysis pathways. JCI Insight. 2023;8(1):e158183. doi:10.1172/jci.insight.158183
25. Bennett BJ, de Aguiar Vallim TQ, Wang Z, et al. Trimethylamine-N-oxide, a metabolite associated with atherosclerosis, exhibits complex genetic and dietary regulation. Cell Metab. 2013;17(1):49–60. doi:10.1016/j.cmet.2012.12.011
26. Ziaka M, Exadaktylos A. Gut-derived immune cells and the gut-lung axis in ARDS. Crit Care. 2024;28(1):220. doi:10.1186/s13054-024-05006-x
27. Wang Z, Ma P, Wang Y, et al. Untargeted metabolomics and transcriptomics identified glutathione metabolism disturbance and PCS and TMAO as potential biomarkers for ER stress in lung. Sci Rep. 2021;11(1):14680. doi:10.1038/s41598-021-92779-8
28. Cao X, Zhao M, Wang X, et al. Multi-metabolomics and intestine microbiome analysis: YZC extract ameliorates septic-ALI by modulating intestine microbiota to reduce TMAO/NLRP3 signaling. Phytomedicine. 2024;130:155345. doi:10.1016/j.phymed.2024.155345
29. Chou RH, Wu PS, Wang SC, et al. Paradox of trimethylamine-N-oxide, the impact of malnutrition on microbiota-derived metabolites and septic patients. J Intensive Care. 2021;9(1):65. doi:10.1186/s40560-021-00581-5
30. Groves HT, Cuthbertson L, James P, et al. Respiratory disease following viral lung infection alters the murine gut microbiota. Front Immunol. 2018;9:182. doi:10.3389/fimmu.2018.00182
31. de Oliveira G, Oliveira C, Pinzan CF, et al. Microbiota modulation of the gut-lung axis in COVID-19. Front Immunol. 2021;12:635471. doi:10.3389/fimmu.2021.635471
32. Terruzzi I, Senesi P. Does intestinal dysbiosis contribute to an aberrant inflammatory response to severe acute respiratory syndrome coronavirus 2 in frail patients. Nutrition. 2020;79–80:110996. doi:10.1016/j.nut.2020.110996
33. Marhuenda-Egea FC, Narro-Serrano J, Shalabi-Benavent MJ, et al. A metabolic readout of the urine metabolome of COVID-19 patients. Metabolomics. 2023;19(2):7. doi:10.1007/s11306-023-01971-6
34. Tristán AI, Jiménez-Luna C, Abreu AC, et al. Metabolomic profiling of COVID-19 using serum and urine samples in intensive care and medical ward cohorts. Sci Rep. 2024;14(1):23713. doi:10.1038/s41598-024-74641-9
35. Zheng Y, Ley SH, Hu FB. Global aetiology and epidemiology of type 2 diabetes mellitus and its complications. Nat Rev Endocrinol. 2018;14(2):88–98. doi:10.1038/nrendo.2017.151
36. Kong L, Zhao Q, Jiang X, et al. Trimethylamine N-oxide impairs β-cell function and glucose tolerance. Nat Commun. 2024;15(1):2526. doi:10.1038/s41467-024-46829-0
37. Chatterjee S, Khunti K, Davies MJ. Type 2 diabetes. Lancet. 2017;389(10085):2239–2251. doi:10.1016/S0140-6736(17)30058-2
38. Sonnenburg JL, Bäckhed F. Diet-microbiota interactions as moderators of human metabolism. Nature. 2016;535(7610):56–64. doi:10.1038/nature18846
39. Dambrova M, Latkovskis G, Kuka J, et al. Diabetes is associated with higher trimethylamine N-oxide plasma levels. Exp Clin Endocrinol Diabetes. 2016;124(4):251–256. doi:10.1055/s-0035-1569330
40. Shan Z, Sun T, Huang H, et al. Association between microbiota-dependent metabolite trimethylamine-N-oxide and type 2 diabetes. Am J Clin Nutr. 2017;106(3):888–894. doi:10.3945/ajcn.117.157107
41. Zhuang R, Ge X, Han L, et al. Gut microbe-generated metabolite trimethylamine N-oxide and the risk of diabetes: a systematic review and dose-response meta-analysis. Obes Rev. 2019;20(6):883–894. doi:10.1111/obr.12843
42. Kalagi NA, Thota RN, Stojanovski E, et al. Association between plasma trimethylamine N-oxide levels and type 2 diabetes: a case control study. Nutrients. 2022;14(10):2093. doi:10.3390/nu14102093
43. Li SY, Chen S, Lu XT, et al. Serum trimethylamine-N-oxide is associated with incident type 2 diabetes in middle-aged and older adults: a prospective cohort study. J Transl Med. 2022;20(1):374. doi:10.1186/s12967-022-03581-7
44. Li D, Lu Y, Yuan S, et al. Gut microbiota-derived metabolite trimethylamine-N-oxide and multiple health outcomes: an umbrella review and updated meta-analysis. Am J Clin Nutr. 2022;116(1):230–243. doi:10.1093/ajcn/nqac074
45. Papandreou C, Bulló M, Zheng Y, et al. Plasma trimethylamine-N-oxide and related metabolites are associated with type 2 diabetes risk in the Prevención con Dieta Mediterránea (PREDIMED) trial. Am J Clin Nutr. 2018;108(1):163–173. doi:10.1093/ajcn/nqy058
46. Gurung M, Li Z, You H, et al. Role of gut microbiota in type 2 diabetes pathophysiology. EBioMedicine. 2020;51:102590. doi:10.1016/j.ebiom.2019.11.051
47. Gao X, Liu X, Xu J, et al. Dietary trimethylamine N-oxide exacerbates impaired glucose tolerance in mice fed a high fat diet. J Biosci Bioeng. 2014;118(4):476–481. doi:10.1016/j.jbiosc.2014.03.001
48. Miao J, Ling AV, Manthena PV, et al. Flavin-containing monooxygenase 3 as a potential player in diabetes-associated atherosclerosis. Nat Commun. 2015;6(1):6498. doi:10.1038/ncomms7498
49. Scheithauer T, Rampanelli E, Nieuwdorp M, et al. Gut microbiota as a trigger for metabolic inflammation in obesity and type 2 diabetes. Front Immunol. 2020;11:571731. doi:10.3389/fimmu.2020.571731
50. Yang G, Zhang X. TMAO promotes apoptosis and oxidative stress of pancreatic acinar cells by mediating IRE1α-XBP-1 pathway. Saudi J Gastroenterol. 2021;27(6):361–369. doi:10.4103/sjg.sjg_12_21
51. Wang H, Luo Q, Ding X, et al. Trimethylamine N-oxide and its precursors in relation to blood pressure: a Mendelian randomization study. Front Cardiovasc Med. 2022;9:922441. doi:10.3389/fcvm.2022.922441
52. Ge X, Zheng L, Zhuang R, et al. The gut microbial metabolite trimethylamine N-oxide and hypertension risk: a systematic review and dose-response meta-analysis. Adv Nutr. 2020;11(1):66–76. doi:10.1093/advances/nmz064
53. Svingen G, Zuo H, Ueland PM, et al. Increased plasma trimethylamine-N-oxide is associated with incident atrial fibrillation. Int J Cardiol. 2018;267:100–106. doi:10.1016/j.ijcard.2018.04.128
54. Nguyen BO, Meems L, van Faassen M, et al. Gut-microbe derived TMAO and its association with more progressed forms of AF: results from the AF-RISK study. Int J Cardiol Heart Vasc. 2021;34:100798. doi:10.1016/j.ijcha.2021.100798
55. Yang W-T, Yang R, Zhao Q, et al. A systematic review and meta-analysis of the gut microbiota-dependent metabolite trimethylamine N-oxide with the incidence of atrial fibrillation. Ann Palliat Med. 2021;10(11):11512–11523. doi:10.21037/apm-21-2763
56. Gong D, Zhang L, Zhang Y, et al. Gut microbial metabolite trimethylamine N-oxide is related to thrombus formation in atrial fibrillation patients. Am J Med Sci. 2019;358(6):422–428. doi:10.1016/j.amjms.2019.09.002
57. Zhu W, Wang Z, Tang W, et al. Gut Microbe-Generated Trimethylamine N-Oxide From Dietary Choline Is Prothrombotic in Subjects. Circulation. 2017;135(17):1671–1673. doi:10.1161/CIRCULATIONAHA.116.025338
58. Hasan RA, Koh AY, Zia A. The gut microbiome and thromboembolism. Thromb Res. 2020;189:77–87. doi:10.1016/j.thromres.2020.03.003
59. Shan Z, Clish CB, Hua S, et al. Gut Microbial-Related Choline Metabolite Trimethylamine-N-Oxide Is Associated With Progression of Carotid Artery Atherosclerosis in HIV Infection. J Infect Dis. 2018;218(9):1474–1479. doi:10.1093/infdis/jiy356
60. Bogiatzi C, Gloor G, Allen-Vercoe E, et al. Metabolic products of the intestinal microbiome and extremes of atherosclerosis. Atherosclerosis. 2018;273:91–97. doi:10.1016/j.atherosclerosis.2018.04.015
61. Randrianarisoa E, Lehn-Stefan A, Wang X, et al. Relationship of serum trimethylamine N-oxide (TMAO) levels with early atherosclerosis in humans. Sci Rep. 2016;6(1):26745. doi:10.1038/srep26745
62. Yin J, Liao SX, He Y, et al. Dysbiosis of gut microbiota with reduced trimethylamine-N-oxide level in patients with large-artery atherosclerotic stroke or transient ischemic attack. J Am Heart Assoc. 2015;4(11):e002699. doi:10.1161/JAHA.115.002699
63. Meyer KA, Benton TZ, Bennett BJ, et al. Microbiota-dependent metabolite trimethylamine N-oxide and coronary artery calcium in the coronary artery risk development in young adults study (CARDIA). J Am Heart Assoc. 2016;5(10):e003970. doi:10.1161/JAHA.116.003970
64. Skagen K, Trøseid M, Ueland T, et al. The Carnitine-butyrobetaine-trimethylamine-N-oxide pathway and its association with cardiovascular mortality in patients with carotid atherosclerosis. Atherosclerosis. 2016;247:64–69. doi:10.1016/j.atherosclerosis.2016.01.033
65. Sardar SW, Nam J, Kim TE, et al. Identification of novel biomarkers for early diagnosis of atherosclerosis using high-resolution metabolomics. Metabolites. 2023;13(11):1160. doi:10.3390/metabo13111160
66. Yu D, Shu XO, Rivera ES, et al. Urinary levels of trimethylamine-N-oxide and incident coronary heart disease: a prospective investigation among urban Chinese adults. J Am Heart Assoc. 2019;8(1):e010606. doi:10.1161/JAHA.118.010606
67. Heianza Y, Ma W, DiDonato JA, et al. Long-term changes in gut microbial metabolite trimethylamine N-oxide and coronary heart disease risk. J Am Coll Cardiol. 2020;75(7):763–772. doi:10.1016/j.jacc.2019.11.060
68. Tang WH, Wang Z, Levison BS, et al. Intestinal microbial metabolism of phosphatidylcholine and cardiovascular risk. N Engl J Med. 2013;368(17):1575–1584. doi:10.1056/NEJMoa1109400
69. Sanchez-Gimenez R, Peiró ÓM, Bonet G, et al. Plasma trimethylamine-N-oxide, its precursors and risk of cardiovascular events in patients with acute coronary syndrome: mediating effects of renal function. Front Cardiovasc Med. 2022;9:1000815. doi:10.3389/fcvm.2022.1000815
70. Dean YE, Rouzan SS, Loayza Pintado JJ, et al. Serum trimethylamine N-oxide levels among coronary artery disease and acute coronary syndrome patients: a systematic review and meta-analysis. Ann Med Surg Lond. 2023;85(12):6123–6133. doi:10.1097/MS9.0000000000001426
71. Li XS, Obeid S, Klingenberg R, et al. Gut microbiota-dependent trimethylamine N-oxide in acute coronary syndromes: a prognostic marker for incident cardiovascular events beyond traditional risk factors. Eur Heart J. 2017;38(11):814–824. doi:10.1093/eurheartj/ehw582
72. Kong W, Ma J, Lin Y, et al. Positive association of plasma trimethylamine-N-oxide and atherosclerosis in patient with acute coronary syndrome. Cardiovasc Ther. 2022;2022:2484018. doi:10.1155/2022/2484018
73. Lever M, George PM, Slow S, et al. Betaine and trimethylamine-N-oxide as predictors of cardiovascular outcomes show different patterns in diabetes mellitus: an observational study. PLoS One. 2014;9(12):e114969. doi:10.1371/journal.pone.0114969
74. Matsuzawa Y, Nakahashi H, Konishi M, et al. Microbiota-derived trimethylamine N-oxide predicts cardiovascular risk after STEMI. Sci Rep. 2019;9(1):11647. doi:10.1038/s41598-019-48246-6
75. Sheng Z, Tan Y, Liu C, et al. Relation of circulating trimethylamine N-oxide with coronary atherosclerotic burden in patients with ST-segment elevation myocardial infarction. Am J Cardiol. 2019;123(6):894–898. doi:10.1016/j.amjcard.2018.12.018
76. Heianza Y, Ma W, Manson JE, et al. Gut microbiota metabolites and risk of major adverse cardiovascular disease events and death: a systematic review and meta-analysis of prospective studies. J Am Heart Assoc. 2017;6(7):e004947. doi:10.1161/JAHA.116.004947
77. Schuett K, Kleber ME, Scharnagl H, et al. Trimethylamine-N-oxide and heart failure with reduced versus preserved ejection fraction. J Am Coll Cardiol. 2017;70(25):3202–3204. doi:10.1016/j.jacc.2017.10.064
78. Tang WH, Kitai T, Hazen SL. Gut microbiota in cardiovascular health and disease. Circ Res. 2017;120(7):1183–1196. doi:10.1161/CIRCRESAHA.117.309715
79. Zabell A, Tang WH. Targeting the microbiome in heart failure. Curr Treat Options Cardiovasc Med. 2017;19(4):27. doi:10.1007/s11936-017-0528-4
80. Tang WH, Wang Z, Shrestha K, et al. Intestinal microbiota-dependent phosphatidylcholine metabolites, diastolic dysfunction, and adverse clinical outcomes in chronic systolic heart failure. J Card Fail. 2015;21(2):91–96. doi:10.1016/j.cardfail.2014.11.006
81. Tang WH, Wang Z, Fan Y, et al. Prognostic value of elevated levels of intestinal microbe-generated metabolite trimethylamine-N-oxide in patients with heart failure: refining the gut hypothesis. J Am Coll Cardiol. 2014;64(18):1908–1914. doi:10.1016/j.jacc.2014.02.617
82. Trøseid M, Ueland T, Hov JR, et al. Microbiota-dependent metabolite trimethylamine-N-oxide is associated with disease severity and survival of patients with chronic heart failure. J Intern Med. 2015;277(6):717–726. doi:10.1111/joim.12328
83. Suzuki T, Heaney LM, Bhandari SS, et al. Trimethylamine N-oxide and prognosis in acute heart failure. Heart. 2016;102(11):841–848. doi:10.1136/heartjnl-2015-308826
84. Yazaki Y, Aizawa K, Israr MZ, et al. Ethnic differences in association of outcomes with trimethylamine N-oxide in acute heart failure patients. ESC Heart Fail. 2020;7(5):2373–2378. doi:10.1002/ehf2.12777
85. Suzuki T, Yazaki Y, Voors AA, et al. Association with outcomes and response to treatment of trimethylamine N-oxide in heart failure: results from BIOSTAT-CHF. Eur J Heart Fail. 2019;21(7):877–886. doi:10.1002/ejhf.1338
86. Kinugasa Y, Nakamura K, Kamitani H, et al. Trimethylamine N-oxide and outcomes in patients hospitalized with acute heart failure and preserved ejection fraction. ESC Heart Fail. 2021;8(3):2103–2110. doi:10.1002/ehf2.13290
87. Salzano A, Israr MZ, Yazaki Y, et al. Combined use of trimethylamine N-oxide with BNP for risk stratification in heart failure with preserved ejection fraction: findings from the DIAMONDHFpEF study. Eur J Prev Cardiol. 2020;27(19):2159–2162. doi:10.1177/2047487319870355
88. Dong Z, Zheng S, Shen Z, et al. Trimethylamine N-oxide is associated with heart failure risk in patients with preserved ejection fraction. Lab Med. 2021;52(4):346–351. doi:10.1093/labmed/lmaa075
89. Lin J, Cheng A, Cheng K, et al. New insights into the mechanisms of pyroptosis and implications for diabetic kidney disease. Int J Mol Sci. 2020;21(19):7057. doi:10.3390/ijms21197057
90. Yang T, Richards EM, Pepine CJ, et al. The gut microbiota and the brain-gut-kidney axis in hypertension and chronic kidney disease. Nat Rev Nephrol. 2018;14(7):442–456. doi:10.1038/s41581-018-0018-2
91. Cañadas-Garre M, Anderson K, McGoldrick J, et al. Proteomic and metabolomic approaches in the search for biomarkers in chronic kidney disease. J Proteomics. 2019;193:93–122. doi:10.1016/j.jprot.2018.09.020
92. Xu KY, Xia GH, Lu JQ, et al. Impaired renal function and dysbiosis of gut microbiota contribute to increased trimethylamine-N-oxide in chronic kidney disease patients. Sci Rep. 2017;7(1):1445. doi:10.1038/s41598-017-01387-y
93. Missailidis C, Hällqvist J, Qureshi AR, et al. Serum trimethylamine-N-oxide is strongly related to renal function and predicts outcome in chronic kidney disease. PLoS One. 2016;11(1):e0141738. doi:10.1371/journal.pone.0141738
94. Tang WH, Wang Z, Kennedy DJ, et al. Gut microbiota-dependent trimethylamine N-oxide (TMAO) pathway contributes to both development of renal insufficiency and mortality risk in chronic kidney disease. Circ Res. 2015;116(3):448–455. doi:10.1161/CIRCRESAHA.116.305360
95. Mafune A, Iwamoto T, Tsutsumi Y, et al. Associations among serum trimethylamine-N-oxide (TMAO) levels, kidney function and infarcted coronary artery number in patients undergoing cardiovascular surgery: a cross-sectional study. Clin Exp Nephrol. 2016;20(5):731–739. doi:10.1007/s10157-015-1207-y
96. Manor O, Zubair N, Conomos MP, et al. A multi-omic association study of trimethylamine N-oxide. Cell Rep. 2018;24(4):935–946. doi:10.1016/j.celrep.2018.06.096
97. Hu DY, Wu MY, Chen GQ, et al. Metabolomics analysis of human plasma reveals decreased production of trimethylamine N-oxide retards the progression of chronic kidney disease. Br J Pharmacol. 2022;179(17):4344–4359. doi:10.1111/bph.15856
98. Andrikopoulos P, Aron-Wisnewsky J, Chakaroun R, et al. Evidence of a causal and modifiable relationship between kidney function and circulating trimethylamine N-oxide. Nat Commun. 2023;14(1):5843. doi:10.1038/s41467-023-39824-4
99. Al-Waiz M, Mitchell SC, Idle JR, et al. The metabolism of 14C-labelled trimethylamine and its N-oxide in man. Xenobiotica. 1987;17(5):551–558. doi:10.3109/00498258709043962
100. Obeid R, Awwad H, Heine GH, et al. Plasma concentrations of trimethylamine-N-oxide, choline, and betaine in patients with moderate to advanced chronic kidney disease and their relation to cardiovascular and renal outcomes. J Ren Nutr. 2024:S1051-2276(24)00060–00068 [pii]. doi:10.1053/j.jrn.2024.03.009
101. Pelletier CC, Croyal M, Ene L, et al. Elevation of trimethylamine-N-oxide in chronic kidney disease: contribution of decreased glomerular filtration rate. Toxins. 2019;11(11):635. doi:10.3390/toxins11110635
102. Stubbs JR, House JA, Ocque AJ, et al. Serum trimethylamine-N-oxide is elevated in CKD and correlates with coronary atherosclerosis burden. J Am Soc Nephrol. 2016;27(1):305–313. doi:10.1681/ASN.2014111063
103. Wang M, Tang W, Li XS, et al. The gut microbial metabolite trimethylamine N-oxide, incident CKD, and kidney function decline. J Am Soc Nephrol. 2024;35(6):749–760. doi:10.1681/ASN.0000000000000344
104. Li Y, Lu H, Guo J, et al. Gut microbiota-derived trimethylamine N-oxide is associated with the risk of all-cause and cardiovascular mortality in patients with chronic kidney disease: a systematic review and dose-response meta-analysis. Ann Med. 2023;55(1):2215542. doi:10.1080/07853890.2023.2215542
105. Shafi T, Powe NR, Meyer TW, et al. Trimethylamine N-oxide and cardiovascular events in hemodialysis patients. J Am Soc Nephrol. 2017;28(1):321–331. doi:10.1681/ASN.2016030374
106. Go AS, Chertow GM, Fan D, et al. Chronic kidney disease and the risks of death, cardiovascular events, and hospitalization. N Engl J Med. 2004;351(13):1296–1305. doi:10.1056/NEJMoa041031
107. Sarnak MJ, Levey AS, Schoolwerth AC, et al. Kidney disease as a risk factor for development of cardiovascular disease: a statement from the American Heart Association Councils on Kidney in Cardiovascular Disease, High Blood Pressure Research, Clinical Cardiology, and Epidemiology and Prevention. Hypertension. 2003;42(5):1050–1065. doi:10.1161/01.HYP.0000102971.85504.7c
108. Bi SH, Su C, Yang P, et al. Higher serum trimethylamine N-oxide (TMAO) levels are associated with increased visceral fat in hemodialysis patients. Clin Nephrol. 2023;100(6):275–283. doi:10.5414/CN111163
109. Chang D, Xu X, Yang Z, et al. Trimethylamine-N-oxide (TMAO) and clinical outcomes in patients with end-stage kidney disease receiving peritoneal dialysis. Perit Dial Int. 2022;42(6):622–630. doi:10.1177/08968608211051809
110. Del Rio D, Zimetti F, Caffarra P, et al. The gut microbial metabolite trimethylamine-N-oxide is present in human cerebrospinal fluid. Nutrients. 2017;9(10):1053. doi:10.3390/nu9101053
111. Vogt NM, Romano KA, Darst BF, et al. The gut microbiota-derived metabolite trimethylamine N-oxide is elevated in Alzheimer’s disease. Alzheimers Res Ther. 2018;10(1):124. doi:10.1186/s13195-018-0451-2
112. Enko D, Zelzer S, Niedrist T, et al. Assessment of trimethylamine-N-oxide at the blood-cerebrospinal fluid barrier: results from 290 lumbar punctures. EXCLI J. 2020;19:1275–1281. doi:10.17179/excli2020-2763
113. Su H, Fan S, Zhang L, et al. TMAO aggregates neurological damage following ischemic stroke by promoting reactive astrocytosis and glial scar formation via the Smurf2/ALK5 axis. Front Cell Neurosci. 2021;15:569424. doi:10.3389/fncel.2021.569424
114. Zhao L, Zhang C, Cao G, et al. Higher Circulating Trimethylamine N-oxide Sensitizes Sevoflurane-Induced Cognitive Dysfunction in Aged Rats Probably by Downregulating Hippocampal Methionine Sulfoxide Reductase A. Neurochem Res. 2019;44(11):2506–2516. doi:10.1007/s11064-019-02868-4
115. Brunt VE, LaRocca TJ, Bazzoni AE, et al. The gut microbiome-derived metabolite trimethylamine N-oxide modulates neuroinflammation and cognitive function with aging. Geroscience. 2021;43(1):377–394. doi:10.1007/s11357-020-00257-2
116. Yu X, Ji C, Shao A. Neurovascular unit dysfunction and neurodegenerative disorders. Front Neurosci. 2020;14:334. doi:10.3389/fnins.2020.00334
117. Wang L, Xiong X, Zhang L, et al. Neurovascular Unit: a critical role in ischemic stroke. CNS Neurosci Ther. 2021;27(1):7–16. doi:10.1111/cns.13561
118. Zlokovic BV. The blood-brain barrier in health and chronic neurodegenerative disorders. Neuron. 2008;57(2):178–201. doi:10.1016/j.neuron.2008.01.003
119. Deng Y, Zou J, Hong Y, et al. Higher circulating trimethylamine N-oxide aggravates cognitive impairment probably via downregulating hippocampal SIRT1 in vascular dementia rats. Cells. 2022;11(22):3650. doi:10.3390/cells11223650
120. Miyamoto N, Pham L-D-D, Seo JH, et al. Crosstalk between cerebral endothelium and oligodendrocyte. Cell Mol Life Sci. 2014;71(6):1055–1066. doi:10.1007/s00018-013-1488-9
121. Kiasalari Z, Afshin-Majd S, Baluchnejadmojarad T, et al. Ellagic acid ameliorates neuroinflammation and demyelination in experimental autoimmune encephalomyelitis: involvement of NLRP3 and pyroptosis. J Chem Neuroanat. 2021;111:101891. doi:10.1016/j.jchemneu.2020.101891
122. Yang F, Zhu W, Cai X, et al. Minocycline alleviates NLRP3 inflammasome-dependent pyroptosis in monosodium glutamate-induced depressive rats. Biochem Biophys Res Commun. 2020;526(3):553–559. doi:10.1016/j.bbrc.2020.02.149
123. Ji X, Tian L, Niu S, et al. Trimethylamine N-oxide promotes demyelination in spontaneous hypertension rats through enhancing pyroptosis of oligodendrocytes. Front Aging Neurosci. 2022;14:963876. doi:10.3389/fnagi.2022.963876
124. Zhu C, Li G, Lv Z, et al. Association of plasma trimethylamine-N-oxide levels with post-stroke cognitive impairment: a 1-year longitudinal study. Neurol Sci. 2020;41(1):57–63. doi:10.1007/s10072-019-04040-w
125. Yilmaz A, Ugur Z, Bisgin H, et al. Targeted metabolic profiling of urine highlights a potential biomarker panel for the diagnosis of Alzheimer’s disease and mild cognitive impairment: a pilot study. Metabolites. 2020;10(9):357. doi:10.3390/metabo10090357
126. de Oliveira Otto MC, Li XS, Wang Z, et al. Longitudinal associations of plasma TMAO and related metabolites with cognitive impairment and dementia in older adults: the cardiovascular health study. J Alzheimers Dis. 2022;89(4):1439–1452. doi:10.3233/JAD-220477
127. Kumari S, Goyal V, Kumaran SS, et al. Quantitative metabolomics of saliva using proton NMR spectroscopy in patients with Parkinson’s disease and healthy controls. Neurol Sci. 2020;41(5):1201–1210. doi:10.1007/s10072-019-04143-4
128. Sankowski B, Księżarczyk K, Raćkowska E, et al. Higher cerebrospinal fluid to plasma ratio of p-cresol sulfate and indoxyl sulfate in patients with Parkinson’s disease. Clin Chim Acta. 2020;501:165–173. doi:10.1016/j.cca.2019.10.038
129. Chen SJ, Kuo CH, Kuo HC, et al. The gut metabolite trimethylamine N-oxide is associated with Parkinson’s disease severity and progression. Mov Disord. 2020;35(11):2115–2116. doi:10.1002/mds.28246
130. Bai F, You L, Lei H, et al. Association between increased and decreased gut microbiota abundance and Parkinson’s disease: a systematic review and subgroup meta-analysis. Exp Gerontol. 2024;191:112444. doi:10.1016/j.exger.2024.112444
131. Bruggeman A, Vandendriessche C, Hamerlinck H, et al. Safety and efficacy of faecal microbiota transplantation in patients with mild to moderate Parkinson’s disease (GUT-PARFECT): a double-blind, placebo-controlled, randomised, Phase 2 trial. EClinicalMedicine. 2024;71:102563. doi:10.1016/j.eclinm.2024.102563
132. Potashkin J, Huang X, Becker C, et al. Understanding the links between cardiovascular disease and Parkinson’s disease. Mov Disord. 2020;35(1):55–74. doi:10.1002/mds.27836
133. Barrea L, Annunziata G, Muscogiuri G, et al. Trimethylamine-N-oxide (TMAO) as novel potential biomarker of early predictors of metabolic syndrome. Nutrients. 2018;10(12):1971. doi:10.3390/nu10121971
134. Uversky VN, Li J, Fink AL. Trimethylamine-N-oxide-induced folding of alpha-synuclein. FEBS Lett. 2001;509(1):31–35. doi:10.1016/s0014-5793(01)03121-0
135. Jahan I, Nayeem SM. Effect of osmolytes on conformational behavior of intrinsically disordered protein α-synuclein. Biophys J. 2019;117(10):1922–1934. doi:10.1016/j.bpj.2019.09.046
136. Chu CQ, Yu LL, Chen W, et al. Dietary patterns affect Parkinson’s disease via the microbiota-gut-brain axis. Trends Food Sci Technol. 2021;116:90–101. doi:10.1016/j.tifs.2021.07.004
137. Feigin VL, Stark BA, Johnson CO; GBD 2019 Stroke Collaborators. Global, regional, and national burden of stroke and its risk factors, 1990–2019: a systematic analysis for the Global Burden of Disease Study 2019. Lancet Neurol. 2021;20(10):795–820. doi:10.1016/S1474-4422(21)00252-0
138. Sekeljic V, Bataveljic D, Stamenkovic S, et al. Cellular markers of neuroinflammation and neurogenesis after ischemic brain injury in the long-term survival rat model. Brain Struct Funct. 2012;217(2):411–420. doi:10.1007/s00429-011-0336-7
139. Pluta R, Salínska E, Puka M, et al. Early changes in extracellular amino acids and calcium concentrations in rabbit hippocampus following complete 15-min cerebral ischemia. Resuscitation. 1988;16(3):193–210. doi:10.1016/0300-9572(88)90046-9
140. Pluta R, Ułamek-Kozioł M, Kocki J, et al. Expression of the tau protein and amyloid protein precursor processing genes in the CA3 area of the hippocampus in the ischemic model of Alzheimer’s disease in the rat. Mol Neurobiol. 2020;57(2):1281–1290. doi:10.1007/s12035-019-01799-z
141. Ułamek-Kozioł M, Czuczwar SJ, Januszewski S, et al. Proteomic and genomic changes in tau protein, which are associated with Alzheimer’s disease after ischemia-reperfusion brain injury. Int J Mol Sci. 2020;21(3):892. doi:10.3390/ijms21030892
142. Pluta R, Ułamek-Kozioł M, Januszewski S, et al. Shared genomic and proteomic contribution of amyloid and tau protein characteristic of Alzheimer’s disease to brain ischemia. Int J Mol Sci. 2020;21(9):3186. doi:10.3390/ijms21093186
143. Pluta R, Ułamek-Kozioł M, Januszewski S, et al. Participation of amyloid and tau protein in neuronal death and neurodegeneration after brain ischemia. Int J Mol Sci. 2020;21(13):4599. doi:10.3390/ijms21134599
144. Li N, Wang X, Sun C, et al. Change of intestinal microbiota in cerebral ischemic stroke patients. BMC Microbiol. 2019;19(1):191. doi:10.1186/s12866-019-1552-1
145. Chen YZ, Huang ZY, Zhou WW, et al. Uncovering the characteristics of the gut microbiota in patients with ischemic stroke and hemorrhagic stroke. Sci Rep. 2024;14(1):11776. doi:10.1038/s41598-024-62606-x
146. Ge P, Duan H, Tao C, et al. TMAO promotes NLRP3 inflammasome activation of microglia aggravating neurological injury in ischemic stroke through FTO/IGF2BP2. J Inflamm Res. 2023;16:3699–3714. doi:10.2147/JIR.S399480
147. Dolkar P, Deyang T, Anand N, et al. Trimethylamine-N-oxide and cerebral stroke risk: a review. Neurobiol Dis. 2024;192:106423. doi:10.1016/j.nbd.2024.106423
148. Rexidamu M, Li H, Jin H, et al. Serum levels of trimethylamine-N-oxide in patients with ischemic stroke. Biosci Rep. 2019;39(6):BSR20190515. doi:10.1042/BSR20190515
149. Zhai Q, Wang X, Chen C, et al. Prognostic value of plasma trimethylamine N-oxide levels in patients with acute ischemic stroke. Cell Mol Neurobiol. 2019;39(8):1201–1206. doi:10.1007/s10571-019-00714-3
150. Schneider C, Okun JG, Schwarz KV, et al. Trimethylamine-N-oxide is elevated in the acute phase after ischaemic stroke and decreases within the first days. Eur J Neurol. 2020;27(8):1596–1603. doi:10.1111/ene.14253
151. Haghikia A, Li XS, Liman TG, et al. Gut microbiota-dependent trimethylamine N-oxide predicts risk of cardiovascular events in patients with stroke and is related to proinflammatory monocytes. Arterioscler Thromb Vasc Biol. 2018;38(9):2225–2235. doi:10.1161/ATVBAHA.118.311023
152. Zhang H, Yao G. Significant correlation between the gut microbiota-derived metabolite trimethylamine-N-oxide and the risk of stroke: evidence based on 23 observational studies. Eur J Clin Nutr. 2023;77(7):731–740. doi:10.1038/s41430-022-01104-7
153. Farhangi MA, Vajdi M, Asghari-Jafarabadi M. Gut microbiota-associated metabolite trimethylamine N-Oxide and the risk of stroke: a systematic review and dose-response meta-analysis. Nutr J. 2020;19(1):76. doi:10.1186/s12937-020-00592-2
154. Sun T, Zhang Y, Yin J, et al. Association of gut microbiota-dependent metabolite trimethylamine N-oxide with first ischemic stroke. J Atheroscler Thromb. 2021;28(4):320–328. doi:10.5551/jat.55962
155. Wu C, Li C, Zhao W, et al. Elevated trimethylamine N-oxide related to ischemic brain lesions after carotid artery stenting. Neurology. 2018;90(15):e1283–e1290. doi:10.1212/WNL.0000000000005298
156. Ke Y, Li D, Zhao M, et al. Gut flora-dependent metabolite Trimethylamine-N-oxide accelerates endothelial cell senescence and vascular aging through oxidative stress. Free Radic Biol Med. 2018;116:88–100. doi:10.1016/j.freeradbiomed.2018.01.007
157. Wang M, Kaufman RJ. Protein misfolding in the endoplasmic reticulum as a conduit to human disease. Nature. 2016;529(7586):326–335. doi:10.1038/nature17041
158. Benson TW, Conrad KA, Li XS, et al. Gut microbiota-derived trimethylamine N-oxide contributes to abdominal aortic aneurysm through inflammatory and apoptotic mechanisms. Circulation. 2023;147(14):1079–1096. doi:10.1161/CIRCULATIONAHA.122.060573
159. Yong C, Huang G, Ge H, et al. Perilla frutescens L. alleviates trimethylamine N-oxide-induced apoptosis in the renal tubule by regulating ASK1-JNK phosphorylation. Phytother Res. 2023;37(4):1274–1292. doi:10.1002/ptr.7684
160. Liu X, Shao Y, Tu J, et al. Trimethylamine-N-oxide-stimulated hepatocyte-derived exosomes promote inflammation and endothelial dysfunction through nuclear factor-kappa B signaling. Ann Transl Med. 2021;9(22):1670. doi:10.21037/atm-21-5043
161. Wang Z, Wu F, Zhou Q, et al. Berberine improves vascular dysfunction by inhibiting trimethylamine-N-oxide via regulating the gut microbiota in angiotensin II-induced hypertensive mice. Front Microbiol. 2022;13:814855. doi:10.3389/fmicb.2022.814855
162. Chen S, Henderson A, Petriello MC, et al. Trimethylamine N-oxide binds and activates PERK to promote metabolic dysfunction. Cell Metab. 2019;30(6):1141–1151.e5. doi:10.1016/j.cmet.2019.08.021
163. Jiayu H, Jiaying L, Hanke Z, et al. Increased trimethylamine N-oxide contributes to metabolic dysfunction in a rat model of PCOS and decreases mitochondrial function. Researchgate. In press: 2020. doi:10.21203/rs.3.rs-122457/v1
164. Wu P, Chen J, Chen J, et al. Trimethylamine N-oxide promotes apoE(-/-) mice atherosclerosis by inducing vascular endothelial cell pyroptosis via the SDHB/ROS pathway. J Cell Physiol. 2020;235(10):6582–6591. doi:10.1002/jcp.29518
165. Singh GB, Zhang Y, Boini KM, et al. High mobility group box 1 mediates TMAO-induced endothelial dysfunction. Int J Mol Sci. 2019;20(14):3570. doi:10.3390/ijms20143570
166. Liu Y, Dai M. Trimethylamine N-oxide generated by the gut microbiota is associated with vascular inflammation: new insights into atherosclerosis. Mediators Inflamm. 2020;2020:4634172. doi:10.1155/2020/4634172
167. Seldin MM, Meng Y, Qi H, et al. Trimethylamine N-oxide promotes vascular inflammation through signaling of mitogen-activated protein kinase and nuclear factor-κB. J Am Heart Assoc. 2016;5(2). doi:10.1161/JAHA.115.002767
168. Liu X, Shao Y, Tu J, et al. TMAO-activated hepatocyte-derived exosomes impair angiogenesis via repressing CXCR4. Front Cell Dev Biol. 2021;9:804049. doi:10.3389/fcell.2021.804049
169. Gong B, Zhang LY, Pang CP, et al. Trimethylamine N-oxide alleviates the severe aggregation and ER stress caused by G98R alphaA-crystallin. Mol Vis. 2009;15:2829–2840.
170. Yang S, Dai H, Lu Y, et al. Trimethylamine N-oxide promotes cell proliferation and angiogenesis in colorectal cancer. J Immunol Res. 2022;2022:7043856. doi:10.1155/2022/7043856
171. Glick D, Barth S, Macleod KF. Autophagy: cellular and molecular mechanisms. J Pathol. 2010;221(1):3–12. doi:10.1002/path.2697
172. Shintani T, Klionsky DJ. Autophagy in health and disease: a double-edged sword. Science. 2004;306(5698):990–995. doi:10.1126/science.1099993
173. Duan X, Kong Z, Mai X, et al. Autophagy inhibition attenuates hyperoxaluria-induced renal tubular oxidative injury and calcium oxalate crystal depositions in the rat kidney. Redox Biol. 2018;16:414–425. doi:10.1016/j.redox.2018.03.019
174. Vanhorebeek I, Gunst J, Derde S, et al. Insufficient activation of autophagy allows cellular damage to accumulate in critically ill patients. J Clin Endocrinol Metab. 2011;96(4):E633–645. doi:10.1210/jc.2010-2563
175. Hu R, Wang MQ, Liu LY, et al. Calycosin inhibited autophagy and oxidative stress in chronic kidney disease skeletal muscle atrophy by regulating AMPK/SKP2/CARM1 signalling pathway. J Cell Mol Med. 2020;24(19):11084–11099. doi:10.1111/jcmm.15514
176. Nishida K, Yamaguchi O, Otsu K. Crosstalk between autophagy and apoptosis in heart disease. Circ Res. 2008;103(4):343–351. doi:10.1161/CIRCRESAHA.108.175448
177. Randow F, Youle RJ. Self and nonself: how autophagy targets mitochondria and bacteria. Cell Host Microbe. 2014;15(4):403–411. doi:10.1016/j.chom.2014.03.012
178. Dong F, Jiang S, Tang C, et al. Trimethylamine N-oxide promotes hyperoxaluria-induced calcium oxalate deposition and kidney injury by activating autophagy. Free Radic Biol Med. 2022;179:288–300. doi:10.1016/j.freeradbiomed.2021.11.010
179. Yue C, Yang X, Li J, et al. Trimethylamine N-oxide prime NLRP3 inflammasome via inhibiting ATG16L1-induced autophagy in colonic epithelial cells. Biochem Biophys Res Commun. 2017;490(2):541–551. doi:10.1016/j.bbrc.2017.06.075
180. Crea F. New challenges in vascular biology and medicine: from unravelling the mechanisms of neointima formation to the prevention of amputations and of ischaemic stroke. Eur Heart J. 2021;42(18):1715–1719. doi:10.1093/eurheartj/ehab224
181. Martinet W, De Bie M, Schrijvers DM, et al. 7-ketocholesterol induces protein ubiquitination, myelin figure formation, and light chain 3 processing in vascular smooth muscle cells. Arterioscler Thromb Vasc Biol. 2004;24(12):2296–2301. doi:10.1161/01.ATV.0000146266.65820.a1
182. Martinet W, Schrijvers DM, Timmermans J-P, et al. Interactions between cell death induced by statins and 7-ketocholesterol in rabbit aorta smooth muscle cells. Br J Pharmacol. 2008;154(6):1236–1246. doi:10.1038/bjp.2008.181
183. Zhang L, Wang J, Liang J, et al. Propofol prevents human umbilical vein endothelial cell injury from Ang II-induced apoptosis by activating the ACE2-(1-7)-Mas axis and eNOS phosphorylation. PLoS One. 2018;13(7):e0199373. doi:10.1371/journal.pone.0199373
184. Shi G, Zeng L, Shi J, et al. Trimethylamine N-oxide promotes atherosclerosis by regulating low-density lipoprotein-induced autophagy in vascular smooth muscle cells through PI3K/AKT/mTOR pathway. Int Heart J. 2023;64(3):462–469. doi:10.1536/ihj.22-603
185. Perrotta I. The use of electron microscopy for the detection of autophagy in human atherosclerosis. Micron. 2013;50:7–13. doi:10.1016/j.micron.2013.03.007
186. Roumenina LT, Rayes J, Frimat M, et al. Endothelial cells: source, barrier, and target of defensive mediators. Immunol Rev. 2016;274(1):307–329. doi:10.1111/imr.12479
187. Hong Q, Que D, Zhong C, et al. Trimethylamine-N-oxide (TMAO) promotes balloon injury-induced neointimal hyperplasia via upregulating Beclin1 and impairing autophagic flux. Biomed Pharmacother. 2022;155:113639. doi:10.1016/j.biopha.2022.113639
188. Levine B, Yuan J. Autophagy in cell death: an innocent convict. J Clin Invest. 2005;115(10):2679–2688. doi:10.1172/JCI26390
189. Ajoolabady A, Pratico D, Ren J. Endothelial dysfunction: mechanisms and contribution to diseases. Acta Pharmacol Sin. 2024;45(10):2023–2031. doi:10.1038/s41401-024-01295-8
190. Qiao X, Yin J, Zheng Z, et al. Endothelial cell dynamics in sepsis-induced acute lung injury and acute respiratory distress syndrome: pathogenesis and therapeutic implications. Cell Commun Signal. 2024;22(1):241. doi:10.1186/s12964-024-01620-y
191. Tang F, Zhao X-L, Xu L-Y, et al. Endothelial dysfunction: pathophysiology and therapeutic targets for sepsis-induced multiple organ dysfunction syndrome. Biomed Pharmacother. 2024;178:117180. doi:10.1016/j.biopha.2024.117180
192. Huveneers S, Phng L-K. Endothelial cell mechanics and dynamics in angiogenesis. Curr Opin Cell Biol. 2024;91:102441. doi:10.1016/j.ceb.2024.102441
193. Kesavardhana S, Malireddi R, Kanneganti TD. Caspases in cell death, inflammation, and pyroptosis. Annu Rev Immunol. 2020;38:567–595. doi:10.1146/annurev-immunol-073119-095439
194. Zhang Y, Chen X, Gueydan C, et al. Plasma membrane changes during programmed cell deaths. Cell Res. 2018;28(1):9–21. doi:10.1038/cr.2017.133
195. Yi ZY, Peng YJ, Hui BP, et al. Zuogui-Jiangtang-Yishen decoction prevents diabetic kidney disease: intervene pyroptosis induced by trimethylamine n-oxide through the mROS-NLRP3 axis. Phytomedicine. 2023;114:154775. doi:10.1016/j.phymed.2023.154775
196. Wu X, Zhang H, Qi W, et al. Nicotine promotes atherosclerosis via ROS-NLRP3-mediated endothelial cell pyroptosis. Cell Death Dis. 2018;9(2):171. doi:10.1038/s41419-017-0257-3
197. Xu YJ, Zheng L, Hu YW, et al. Pyroptosis and its relationship to atherosclerosis. Clin Chim Acta. 2018;476:28–37. doi:10.1016/j.cca.2017.11.005
198. Tian C, Liu Y, Li Z, et al. Mitochondria related cell death modalities and disease. Front Cell Dev Biol. 2022;10:832356. doi:10.3389/fcell.2022.832356
199. Mills EL, Kelly B, Logan A, et al. Succinate dehydrogenase supports metabolic repurposing of mitochondria to drive inflammatory macrophages. Cell. 2016;167(2):457–470.e13. doi:10.1016/j.cell.2016.08.064
200. Tsai HY, Hsu YJ, Lu CY, et al. Pharmacological activation of aldehyde dehydrogenase 2 protects against heatstroke-induced acute lung injury by modulating oxidative stress and endothelial dysfunction. Front Immunol. 2021;12:740562. doi:10.3389/fimmu.2021.740562
201. Ge P, Luo Y, Yang Q, et al. Ferroptosis in rat lung tissue during severe acute pancreatitis-associated acute lung injury: protection of Qingyi decoction. Oxid Med Cell Longev. 2023;2023:5827613. doi:10.1155/2023/5827613
202. Hu J, Yang L, Peng X, et al. ALDH2 hampers immune escape in liver hepatocellular carcinoma through ROS/Nrf2-mediated autophagy. Inflammation. 2022;45(6):2309–2324. doi:10.1007/s10753-022-01694-1
203. Chen CH, Ferreira J, Mochly-Rosen D. ALDH2 and cardiovascular disease. Adv Exp Med Biol. 2019;1193:53–67. doi:10.1007/978-981-13-6260-6_3
204. Long P, He M, Yan W, et al. ALDH2 protects naturally aged mouse retina via inhibiting oxidative stress-related apoptosis and enhancing unfolded protein response in endoplasmic reticulum. Aging. 2020;13(2):2750–2767. doi:10.18632/aging.202325
205. Li J, Lü H, Chen S, et al. Trimethylamine oxide induces pyroptosis of vascular endothelial cells through ALDH2/ROS/NLRP3/GSDMD pathway. Zhong Nan da Xue Xue Bao Yi Xue Ban. 2022;47(9):1171–1181. doi:10.11817/j.issn.1672-7347.2022.220086
206. Yu J, Li S, Qi J, et al. Cleavage of GSDME by caspase-3 determines lobaplatin-induced pyroptosis in colon cancer cells. Cell Death Dis. 2019;10(3):193. doi:10.1038/s41419-019-1441-4
207. Fan JX, Deng RH, Wang H, et al. Epigenetics-based tumor cells pyroptosis for enhancing the immunological effect of chemotherapeutic nanocarriers. Nano Lett. 2019;19(11):8049–8058. doi:10.1021/acs.nanolett.9b03245
208. Hu L, Chen M, Chen X, et al. Chemotherapy-induced pyroptosis is mediated by BAK/BAX-caspase-3-GSDME pathway and inhibited by 2-bromopalmitate. Cell Death Dis. 2020;11(4):281. doi:10.1038/s41419-020-2476-2
209. Rogers C, Fernandes-Alnemri T, Mayes L, et al. Cleavage of DFNA5 by caspase-3 during apoptosis mediates progression to secondary necrotic/pyroptotic cell death. Nat Commun. 2017;8(1):14128. doi:10.1038/ncomms14128
210. Wang Y, Gao W, Shi X, et al. Chemotherapy drugs induce pyroptosis through caspase-3 cleavage of a gasdermin. Nature. 2017;547(7661):99–103. doi:10.1038/nature22393
211. Wang L, Wang Y, Xu H, et al. Effect of dapagliflozin on ferroptosis through the gut microbiota metabolite TMAO during myocardial ischemia-reperfusion injury in diabetes mellitus rats. Sci Rep. 2024;14(1):13851. doi:10.1038/s41598-024-64909-5
212. Luo Z, Yu X, Wang C, et al. Trimethylamine N-oxide promotes oxidative stress and lipid accumulation in macrophage foam cells via the Nrf2/ABCA1 pathway. J Physiol Biochem. 2024;80(1):67–79. doi:10.1007/s13105-023-00984-y
213. González-Correa C, Moleón J, Miñano S, et al. Trimethylamine N-oxide promotes autoimmunity and a loss of vascular function in toll-like receptor 7-driven lupus mice. Antioxidants. 2021;11(1):84. doi:10.3390/antiox11010084
214. Ingold I, Berndt C, Schmitt S, et al. Selenium utilization by GPX4 is required to prevent hydroperoxide-induced ferroptosis. Cell. 2018;172(3):409–422.e21. doi:10.1016/j.cell.2017.11.048
215. Ge H, Wei Y, Zhang W, et al. Suyin Detoxification Granule alleviates trimethylamine N-oxide-induced tubular ferroptosis and renal fibrosis to prevent chronic kidney disease progression. Phytomedicine. 2024;135:156195. doi:10.1016/j.phymed.2024.156195
216. Dodson M, Castro-Portuguez R, Zhang DD. NRF2 plays a critical role in mitigating lipid peroxidation and ferroptosis. Redox Biol. 2019;23:101107. doi:10.1016/j.redox.2019.101107
217. Nguyen KT, Mun SH, Yang J, et al. The MARCHF6 E3 ubiquitin ligase acts as an NADPH sensor for the regulation of ferroptosis. Nat Cell Biol. 2022;24(8):1239–1251. doi:10.1038/s41556-022-00973-1
218. Hill C, Guarner F, Reid G, et al. Expert consensus document. The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat Rev Gastroenterol Hepatol. 2014;11(8):506–514. doi:10.1038/nrgastro.2014.66
219. Mahdavi-Roshan M, Salari A, Kheirkhah J, et al. The effects of probiotics on inflammation, endothelial dysfunction, and atherosclerosis progression: a mechanistic overview. Heart Lung Circ. 2022;31(5):e45–e71. doi:10.1016/j.hlc.2021.09.006
220. O’Morain VL, Ramji DP. The Potential of Probiotics in the Prevention and Treatment of Atherosclerosis. Mol Nutr Food Res. 2020;64(4):e1900797. doi:10.1002/mnfr.201900797
221. Katsimichas T, Theofilis P, Tsioufis K, et al. Gut microbiota and coronary artery disease: current therapeutic perspectives. Metabolites. 2023;13(2):256. doi:10.3390/metabo13020256
222. Dong Y, Gai Z, Han M, et al. Reduction in serum concentrations of uremic toxins driven by Bifidobacterium Longum Subsp. Longum BL21 is Associated with gut microbiota changes in a rat model of chronic kidney disease. Probiotics Antimicrob Proteins. 2024. doi:10.1007/s12602-024-10293-5
223. Wang X, Yang J, Qiu X, et al. Probiotics, pre-biotics and synbiotics in the treatment of pre-diabetes: a systematic review of randomized controlled trials. Front Public Health. 2021;9:645035. doi:10.3389/fpubh.2021.645035
224. Qiu L, Tao X, Xiong H, et al. Lactobacillus plantarum ZDY04 exhibits a strain-specific property of lowering TMAO via the modulation of gut microbiota in mice. Food Funct. 2018;9(8):4299–4309. doi:10.1039/c8fo00349a
225. Ramireddy L, Tsen HY, Chiang YC, et al. Molecular identification and selection of probiotic strains able to reduce the serum TMAO level in mice challenged with choline. Foods. 2021;10(12):2931. doi:10.3390/foods10122931
226. Qiu L, Yang D, Tao X, et al. Enterobacter aerogenes ZDY01 attenuates choline-induced trimethylamine N-oxide levels by remodeling gut microbiota in mice. J Microbiol Biotechnol. 2017;27(8):1491–1499. doi:10.4014/jmb.1703.03039
227. Tang J, Qin M, Tang L, et al. Enterobacter aerogenes ZDY01 inhibits choline-induced atherosclerosis through CDCA-FXR-FGF15 axis. Food Funct. 2021;12(20):9932–9946. doi:10.1039/d1fo02021h
228. Liu J, Li T, Wu H, et al. Lactobacillus rhamnosus GG strain mitigated the development of obstructive sleep apnea-induced hypertension in a high salt diet via regulating TMAO level and CD4(+) T cell induced-type I inflammation. Biomed Pharmacother. 2019;112:108580. doi:10.1016/j.biopha.2019.01.041
229. Tungsanga S, Katavetin P, Panpetch W, et al. Lactobacillus rhamnosus L34 attenuates chronic kidney disease progression in a 5/6 nephrectomy mouse model through the excretion of anti-inflammatory molecules. Nephrol Dial Transplant. 2022;37(8):1429–1442. doi:10.1093/ndt/gfac032
230. Moludi J, Saiedi S, Ebrahimi B, et al. Probiotics supplementation on cardiac remodeling following myocardial infarction: a single-center double-blind clinical study. J Cardiovasc Transl Res. 2021;14(2):299–307. doi:10.1007/s12265-020-10052-1
231. Ramos M, Felicidade I, de Oliveira Carlos L, et al. Effect of probiotic supplementation on plasma metabolite profile after Roux-Y gastric bypass: a prospective, randomized, double-blind, placebo-controlled clinical trial. Int J Obes Lond. 2022;46(11):2006–2012. doi:10.1038/s41366-022-01213-0
232. Liang X, Zhang Z, Lv Y, et al. Reduction of intestinal trimethylamine by probiotics ameliorated lipid metabolic disorders associated with atherosclerosis. Nutrition. 2020;79–80:110941. doi:10.1016/j.nut.2020.110941
233. Tripolt NJ, Leber B, Triebl A, et al. Effect of Lactobacillus casei Shirota supplementation on trimethylamine-N-oxide levels in patients with metabolic syndrome: an open-label, randomized study. Atherosclerosis. 2015;242(1):141–144. doi:10.1016/j.atherosclerosis.2015.05.005
234. Poesen R, Evenepoel P, de Loor H, et al. The influence of prebiotic arabinoxylan oligosaccharides on microbiota derived uremic retention solutes in patients with chronic kidney disease: a randomized controlled trial. PLoS One. 2016;11(4):e0153893. doi:10.1371/journal.pone.0153893
235. Chen ML, Yi L, Zhang Y, et al. Resveratrol attenuates trimethylamine-N-oxide (TMAO)-induced atherosclerosis by regulating TMAO synthesis and bile acid metabolism via remodeling of the gut microbiota. mBio. 2016;7(2):e02210–02215. doi:10.1128/mBio.02210-15
236. Xiong Q, Li L, Xiao Y, et al. The effect of inulin-type fructans on plasma trimethylamine N-oxide levels in peritoneal dialysis patients: a randomized crossover trial. Mol Nutr Food Res. 2023;67(9):e2200531. doi:10.1002/mnfr.202200531
237. Hsu CN, Hou CY, Chan J, et al. Hypertension programmed by perinatal high-fat diet: effect of maternal gut microbiota-targeted therapy. Nutrients. 2019;11(12):2908. doi:10.3390/nu11122908
238. Gibson GR, Hutkins R, Sanders ME, et al. Expert consensus document: the International Scientific Association for Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of prebiotics. Nat Rev Gastroenterol Hepatol. 2017;14(8):491–502. doi:10.1038/nrgastro.2017.75
239. Chalmers RA, Bain MD, Michelakakis H, et al. Diagnosis and management of trimethylaminuria (FMO3 deficiency) in children. J Inherit Metab Dis. 2006;29(1):162–172. doi:10.1007/s10545-006-0158-6
240. Kuka J, Videja M, Makrecka-Kuka M, et al. Metformin decreases bacterial trimethylamine production and trimethylamine N-oxide levels in db/db mice. Sci Rep. 2020;10(1):14555. doi:10.1038/s41598-020-71470-4
241. Aziz F, Tripolt NJ, Pferschy PN, et al. Alterations in trimethylamine-N-oxide in response to Empagliflozin therapy: a secondary analysis of the EMMY trial. Cardiovasc Diabetol. 2023;22(1):184. doi:10.1186/s12933-023-01920-6
242. Ge PX, Tai T, Jiang LP, et al. Choline and trimethylamine N-oxide impair metabolic activation of and platelet response to clopidogrel through activation of the NOX/ROS/Nrf2/CES1 pathway. J Thromb Haemost. 2023;21(1):117–132. doi:10.1016/j.jtha.2022.10.010
243. Li DY, Li XS, Chaikijurajai T, et al. Relation of statin use to gut microbial trimethylamine N-oxide and cardiovascular risk. Am J Cardiol. 2022;178:26–34. doi:10.1016/j.amjcard.2022.05.010
244. Xiong X, Zhou J, Fu Q, et al. The associations between TMAO-related metabolites and blood lipids and the potential impact of rosuvastatin therapy. Lipids Health Dis. 2022;21(1):60. doi:10.1186/s12944-022-01673-3
245. Li X, Su C, Jiang Z, et al. Berberine attenuates choline-induced atherosclerosis by inhibiting trimethylamine and trimethylamine-N-oxide production via manipulating the gut microbiome. NPJ Biofilms Microbiomes. 2021;7(1):36. doi:10.1038/s41522-021-00205-8
246. Qu H, Zhang Y, Shi JH, et al. Berberine decreases thrombosis potential induced by a high-choline diet by inhibiting CutC enzyme. Curr Med Chem. 2024;31(24):3844–3856. doi:10.2174/0929867330666230524142632
247. Ma SR, Tong Q, Lin Y, et al. Berberine treats atherosclerosis via a vitamine-like effect down-regulating Choline-TMA-TMAO production pathway in gut microbiota. Signal Transduct Target Ther. 2022;7(1):207. doi:10.1038/s41392-022-01027-6
248. Anwar S, Bhandari U, Panda BP, et al. Trigonelline inhibits intestinal microbial metabolism of choline and its associated cardiovascular risk. J Pharm Biomed Anal. 2018;159:100–112. doi:10.1016/j.jpba.2018.06.027
249. He Z, Zhu H, Liu J, et al. Mangiferin alleviates trimethylamine- N-oxide (TMAO)-induced atherogenesis and modulates gut microbiota in mice. Food Funct. 2023;14(20):9212–9225. doi:10.1039/d3fo02791k
250. Zhang J, Ou C, Chen M. Curcumin attenuates cadmium-induced atherosclerosis by regulating trimethylamine-N-oxide synthesis and macrophage polarization through remodeling the gut microbiota. Ecotoxicol Environ Saf. 2022;244:114057. doi:10.1016/j.ecoenv.2022.114057
251. Shi HH, Chen LP, Wang CC, et al. Docosahexaenoic acid-acylated curcumin diester alleviates cisplatin-induced acute kidney injury by regulating the effect of gut microbiota on the lipopolysaccharide- and trimethylamine-N-oxide-mediated PI3K/Akt/NF-κB signaling pathway in mice. Food Funct. 2022;13(11):6103–6117. doi:10.1039/d1fo04178a
252. Satheesh Babu AK, Petersen C, Iglesias-Carres L, et al. Blueberry intervention mitigates detrimental microbial metabolite trimethylamine N-oxide by modulating gut microbes. Biofactors. 2024;50(2):392–404. doi:10.1002/biof.2014
253. Yang JY, Zhang TT, Yu ZL, et al. Taurine alleviates trimethylamine N-oxide-induced atherosclerosis by regulating bile acid metabolism in ApoE(-/-) mice. J Agric Food Chem. 2022;70(18):5738–5747. doi:10.1021/acs.jafc.2c01376
254. Wu X, Chen L, Zeb F, et al. Regulation of circadian rhythms by NEAT1 mediated TMAO-induced endothelial proliferation: a protective role of asparagus extract. Exp Cell Res. 2019;382(1):111451. doi:10.1016/j.yexcr.2019.05.032
255. Zhang X, Wu Q, Zhao Y, et al. Decaisnea insignis seed oil inhibits trimethylamine-N-oxide formation and remodels intestinal microbiota to alleviate liver dysfunction in l-carnitine feeding mice. J Agric Food Chem. 2019;67(47):13082–13092. doi:10.1021/acs.jafc.9b05383
256. Gao X, Zhang H, Li K, et al. Sandalwood seed oil improves insulin sensitivity in high-fat/high-sucrose diet-fed rats associated with altered intestinal microbiota and its metabolites. Food Funct. 2021;12(20):9739–9749. doi:10.1039/d1fo02239c
257. Patel SS, Acharya A, Ray RS, et al. Cellular and molecular mechanisms of curcumin in prevention and treatment of disease. Crit Rev Food Sci Nutr. 2020;60(6):887–939. doi:10.1080/10408398.2018.1552244
258. Shehzad A, Wahid F, Lee YS. Curcumin in cancer chemoprevention: molecular targets, pharmacokinetics, bioavailability, and clinical trials. Arch Pharm. 2010;343(9):489–499. doi:10.1002/ardp.200900319
259. Jiang C, Wang S, Wang Y, et al. Polyphenols from hickory nut reduce the occurrence of atherosclerosis in mice by improving intestinal microbiota and inhibiting trimethylamine N-oxide production. Phytomedicine. 2024;128:155349. doi:10.1016/j.phymed.2024.155349
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms.php
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms.php
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.