")
Back to Journals » Journal of Inflammation Research » Volume 18
Metabolism-Related Adipokines and Metabolic Diseases: Their Role in Osteoarthritis
Authors Zhang Q, Zhao YX, Li LF, Fan QQ, Huang BB, Du HZ, Li C , Li W
Received 8 October 2024
Accepted for publication 31 December 2024
Published 25 January 2025 Volume 2025:18 Pages 1207—1233
DOI https://doi.org/10.2147/JIR.S499835
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Professor Ning Quan
Metabolism-related adipokines and diseases in osteoarthritisa – Video abstract [499835]
Views: 79
Qian Zhang,1,* Yi Xuan Zhao,1,* Long Fei Li,2 Qian Qian Fan,3 Bin Bin Huang,1 Hong Zhen Du,1 Chen Li,3 Wei Li3
1School of Special Education and Rehabilitation, Binzhou Medical University, Yantai, Shandong, People’s Republic of China; 2Cerebrovascular Disease Ward, The First People’s Hospital of Ping Ding Shan, Pingdingshan, Henan, People’s Republic of China; 3Department of Rehabilitation Medicine, Binzhou Medical University Hospital, Binzhou, Shandong, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Wei Li; Chen Li, Department of Rehabilitation Medicine Binzhou Medical University Hospital, No. 661 huanghe 2nd Road, Binzhou, Shandong, People’s Republic of China, Email [email protected]; [email protected]
Abstract: Osteoarthritis (OA) affects several joints but tends to be more prevalent in those that are weight-bearing, such as the knees, which are the most heavily loaded joints in the body. The incidence and disability rates of OA have continued to increase and seriously jeopardise the quality of life of middle-aged and older adults. However, OA is more than just a wear and tear disease; its aetiology is complex, and its pathogenesis is poorly understood. Metabolic syndrome (MetS) has emerged as a critical driver of OA development. This condition contributes to the formation of a distinct phenotype, termed metabolic syndrome-associated osteoarthritis (MetS-OA),which differs from other metabolically related diseases by its unique pathophysiological mechanisms and clinical presentation. As key mediators of MetS, metabolic adipokines such as leptin, lipocalin, and resistin regulate inflammation and bone metabolism through distinct or synergistic signaling pathways. Their modulation of inflammatory responses and bone remodeling processes plays a critical role in the pathogenesis and progression of OA. Due to their central role in regulating inflammation and bone remodeling, metabolic adipokines not only deepen our understanding of MetS-OA pathogenesis but also represent promising targets for novel therapeutic strategies that could slow disease progression and improve clinical outcomes in affected patients.
Keywords: osteoarthritis, metabolic syndrome, adipokines
Introduction
OA is a chronic degenerative disease characterised by pain, joint stiffness, and swelling, mostly in the knee joint and hip joint.1–4 There are gender differences in the prevalence and incidence of OA, with women generally being at higher risk than men, especially post-menopausal women around the age of 50; age is also an influencing factor in OA, with the incidence of OA of the knees and hips increasing with age for both men and women. Furthermore, the socio-economic level is an influencing factor in the incidence of OA, with poorer areas, such as rural areas, tending to have higher incidence rates than cities do.5–9 The prevalence and burden of OA are growing exponentially and are expected to affect 78.4 million people by 2040.10,11 As China gradually enters an ageing society, OA can seriously affect the quality of life of patients and impose a huge social and economic burden.12 Current treatments for OA are largely limited to steroidal or non-steroidal anti-inflammatory drugs, which only relieve pain and inflammatory symptoms.13 As a result, there is no effective treatment for the disease at this time.14
MetS is a multifaceted condition defined by a cluster of metabolic abnormalities, including obesity, hypertension, hyperglycemia, insulin resistance, and dyslipidemia.15 While traditionally recognized as a major risk factor for cardiovascular diseases and diabetes, emerging evidence highlights its critical role in the pathogenesis and progression of OA.15–17 MetS-related metabolic disturbances adversely affect multiple joint tissues, such as cartilage, bone, and synovium, through mechanisms that involve chronic low-grade inflammation, oxidative stress, and imbalances in adipokine regulation.15–17These interconnected processes collectively contribute to the initiation and exacerbation of OA, positioning MetS as a significant driver of disease development. The concept of metabolic syndrome-associated MetS-OA has recently been introduced, despite the high prevalence of MetS-OA, its underlying pathogenic mechanisms remain poorly understood.16 Metabolic syndrome has been shown to promote the release of inflammatory cytokines, particularly adipokines, which play a pivotal role in establishing a chronic low-grade inflammatory state, driving cartilage degeneration, and disrupting the balance of the intra-articular environment.16,18 These insights underscore the critical contribution of adipokines to the pathogenesis of MetS-OA.16
Adipokines, primarily secreted by white adipose tissue, are signaling molecules that regulate inflammation and metabolic processes, with dysregulation linked to various diseases, including OA.19–21 In OA, they are central mediators of pathogenesis, contributing to chronic inflammation, cartilage degradation, and bone remodeling imbalances through shared inflammatory and metabolic pathways such as NF-κB, PI3K/Akt, and MAPK.22,23 Recent clinical studies have highlighted their dual roles in OA progression, with some exacerbating the disease while others exhibit protective effects. This duality not only establishes adipokines as key contributors to OA pathophysiology but also underscores their potential as diagnostic biomarkers and therapeutic targets. Emerging targeted therapies for specific adipokines offer promising opportunities to modulate their activity, paving the way for personalized treatment strategies that address both inflammatory and metabolic components of OA and potentially transforming the management of this complex disease.
Further research is needed to confirm these relationships and investigate the role of adipose tissue in OA development. This review focuses on the impact and mechanisms of metabolic diseases and adipokines on OA onset and progression (Figure 1).
 |
Figure 1 Metabolic diseases and adipokines influence the progression of OA through regulating inflammation and matrix degradation. |
Metabolic Syndrome
MetS is a global health problem that is increasing globally and will account for approximately 1/3 of the world’s population by 2022, making it a major public health problem.24–26 It is usually made up of four components: hyperglycaemia, hypertension, dyslipidaemia, and obesity.27–29 The relationship between MetS and OA has been extensively studied (Figure 2).Some studies argue that metabolism minimally impacts OA, with one study showing no significant effect of MetS on OA incidence after adjusting for body mass index (BMI).30 Conversely, other research suggests that MetS contributes to OA by increasing systemic inflammatory mediators from adiposity.31 Moreover, in patients aged 18–78 years with OA, those with MetS develop OA earlier, have more extensive pathology, increased inflammation, and increased joint pain than do those without it.32,33 MetS manifests in a variety of ways, depending on the components that make up the syndrome,34 making its early recognition particularly important. In the context of economic development and improved living standards, the number of patients with MetS has increased dramatically worldwide.35,36 Therefore, there is a need to explore the links and mechanisms between MetS and OA to facilitate better prevention and treatment of OA.
 |
Figure 2 A Comprehensive Overview of OA Pathophysiology Linked to Metabolic Diseases: Mechanisms of Onset. This figure illustrates the mechanisms by which MetS contributes to OA development through shared pathways of inflammation, oxidative stress, and metabolic dysregulation: Obesity: Drives cartilage degradation and joint instability via pro-inflammatory cytokines (IL-6, TNF-α), ROS, and increased joint loading. DM: Amplifies oxidative stress and inflammation through AGEs and ROS while reduced H2S levels and impaired collagen synthesis exacerbate cartilage degeneration and joint instability. HT: Activates renin-angiotensin, endothelin, and Wnt-β-catenin pathways, leading to subchondral bone calcification and remodeling imbalance. DL: Induces chronic inflammation and oxidative stress, disrupting cartilage homeostasis and accelerating OA progression. OP: Weakens subchondral bone, resulting in cartilage calcification, biomechanical imbalance, and rapid OA progression. |
Relationship Between Obesity and OA
Obesity, a hallmark of MetS, is characterized by excessive fat accumulation. Over the past 50 years, its prevalence has risen steadily worldwide, with more than 2 billion individuals expected to be affected in the near future.37,38 As a major global health challenge, obesity has emerged as a significant risk factor for OA progression through adipose tissue-dependent inflammation.39–43 BMI, the primary clinical and research metric for measuring obesity, is positively correlated with OA risk. Specifically, individuals with a BMI greater than 30 face a two-thirds lifetime risk of OA and a doubled risk of asymptomatic OA.44,45 While mechanical joint overload caused by a high BMI has long been considered a primary driver of OA,46–49 it fails to fully explain the high prevalence of OA in non-weight-bearing joints, such as the hands.45 Recent evidence highlights the critical role of obesity-associated systemic inflammation and metabolic dysregulation in OA pathogenesis.50 In obesity, adipose tissue undergoes inflammatory remodeling, marked by a significant increase in pro-inflammatory macrophage infiltration. These macrophages release cytokines such as interleukin-6 (IL-6) and tumor necrosis factor-alpha (TNF-α), disrupting chondrocyte homeostasis and accelerating cartilage degradation.51–55 Concurrently, elevated leptin levels in synovial fluid enhance the interaction between chondrocytes and synovial fibroblasts, further amplifying IL-6 production and exacerbating local inflammatory responses.56,57 This adipose tissue-driven inflammatory cascade links systemic metabolic disturbances to localized joint pathology, establishing a critical connection between obesity and OA.
By elucidating the interplay between systemic inflammation, local joint damage, and metabolic dysregulation, this framework underscores the central role of adipose tissue-dependent inflammation in OA. These insights deepen our understanding of obesity-induced OA and provide a robust foundation for developing targeted interventions that address both the inflammatory and metabolic components of this complex disease.
Relationship Between Diabetes Mellitus and OA
Diabetes mellitus (DM) is a prevalent non-communicable disease characterized by systemic metabolic dysregulation caused by an imbalance between risk and protective factors.58,59 Studies have demonstrated a strong association between DM and the onset and progression of OA.58 Compared to non-diabetic OA patients, those with DM typically exhibit greater pain intensity and poorer physical and mental health.60 While numerous studies have identified a significant link between DM and OA—such as an increased risk of joint replacement surgery—the exact mechanisms underlying this relationship remain unclear and controversial.61,62
Emerging evidence indicates that DM exacerbates OA progression through three primary mechanisms: chronic inflammation, joint structure degeneration, and joint instability.63 Elevated levels of IL-6 and Progranulin(PGRN) in OA joint tissues of DM patients suggest that cartilage in these individuals is more susceptible to pro-inflammatory stress, leading to heightened inflammatory responses.64 Furthermore, increased reactive oxygen species (ROS) in DM-associated OA not only stimulate the production of inflammatory mediators such as IL-1β but also inhibit collagen synthesis in cartilage, accelerating cartilage degradation.65–67 This oxidative stress further disrupts cartilage homeostasis, potentially due to decreased levels of protective factors such as hydrogen sulfide (H2S) and nuclear factor erythroid 2-related factor 2 (Nrf-2).68 DM also contributes to neuromuscular deficits, which exacerbate joint instability and increase cartilage friction, thereby further advancing OA progression.68 These DM-induced structural changes amplify joint instability, accelerating disease development. However, some studies dispute the direct relationship between DM and OA, arguing that DM is not an independent risk factor for OA. Such discrepancies may arise from differences in study design, patient populations, and the multifactorial etiologies of both diseases.69–71
To resolve these controversies, future research should clarify the molecular mechanisms linking DM to OA, particularly through inflammatory and oxidative stress pathways. Studies should examine how DM differentially affects weight-bearing and non-weight-bearing joints and explore variations in OA progression across DM types. Addressing these gaps will enhance our understanding of the DM-OA relationship and facilitate the development of targeted therapies addressing shared metabolic and inflammatory pathways.
Relationship Between Hypertension and OA
Hypertension (HT) affects more than 1 billion adults worldwide, and its prevalence is on the rise.72 HT has been found to be significantly associated with the development of OA, and OA prevalence in patients with hypertension is approximately 40%.73,74 Gender is an influential factor in the relationship between HT and OA, with Lawrence suggesting that HT is associated with OA in women.75 Yang et al concluded that HT is linked to an increased OA risk in men, potentially due to joint structural degeneration and biochemical pathways.76 Increased subchondral bone calcification was found in hypertensive OA animals compared to normal OA animals,77 and a meta-analysis reported a stronger correlation between hypertension and knee OA on imaging, suggesting that hypertension is strongly associated with structural damage to the OA.78 Furthermore, HT activates the renin-angiotensin and endothelin systems, affecting the Wnt-β-catenin signalling pathway and potentially influencing joint disease.76 The causal link between OA and HT is unclear, and future research should clarify how HT impacts OA joints.
Relationship Between Dyslipidaemias and OA
Dyslipidaemias (DLs) are typically characterised by abnormal levels of serum cholesterol, triglycerides, or both, as well as abnormal levels of associated lipoprotein species.79 The link between DL and OA is debated. Several studies indicate DL may elevate OA risk,80–83 likely due to its impact on body homeostasis and association with chronic inflammation and oxidative stress.84,85 Conversely, Inoue found no significant of DL on hand OA risk.86
Relationship Between Osteoporosis and OA
Osteoporosis (OP) is a systemic skeletal disease increasingly prevalent with aging, marked by reduced bone strength and microarchitectural deterioration.87 Both OP and OA are complex, multifactorial disorders lacking a complete cure, significantly contributing to pain and socio-economic burdens worldwide. It is imperative to explore their interrelation and devise new therapeutic strategies.88–90 Recent studies highlight OA’s susceptibility to subchondral fragility, potentially leading to the emergent concept of OP-OA, meriting further attention.91 Although a negative correlation between OP and subsequent OA development is suggested by most research, including a two-sample MR analysis indicating that OP may lower OA incidence, this relationship remains debated.92,93 Further findings indicate that OP-OA patients experience faster OA progression due to abnormal subchondral bone remodelling, increased cartilage calcification and damage, and biomechanical deterioration.94–96 However, the exact mechanisms linking these conditions require more investigation.
The study of metabolic diseases associated with OA has transformed our understanding of its pathogenesis. It is now recognized that OA extends beyond mere joint “wear and tear” due to overloading; it is a chronic inflammatory condition influenced by systemic metabolism, inflicting both physical and psychological stress on patients, and imposing a significant economic burden on society. Consequently, exploring the link between metabolic diseases and OA to identify novel treatment strategies is a promising approach. The various roles of metabolic diseases in the context of OA have been discussed in Table 1.
 |
Table 1 Major Epidemiological Studies of Metabolic Diseases Associated with OA |
Metabolic Diseases and OA Pathogenesis
Obesity, DM, HT, DL, and OP collectively drive the onset and progression of OA through convergent mechanisms involving inflammation, oxidative stress, and metabolic dysregulation. Together, these MetS not only contribute to structural joint damage but also reveal the intricate, multifaceted complexity of OA pathogenesis.
Pro-inflammatory factors associated with obesity, such as IL-6, TNF-α, and leptin, aggravate cartilage degradation and local inflammation through the activation of NF-κB and PI3K/Akt signaling pathways.51–55 In DM, the accumulation of advanced glycation end-products (AGEs) and elevated levels of ROS amplify oxidative stress in cartilage while suppressing protective mechanisms mediated by nuclear factor Nrf-2 and H2S.68 This interplay accelerates cartilage deterioration and exacerbates joint instability. Hypertension contributes to OA-related structural damage by activating the Wnt-β-catenin pathway, which promotes subchondral bone calcification and remodeling imbalances.76,77 Although the precise role of dyslipidemia remains contested, chronic inflammation and oxidative stress likely mediate its contribution to OA progression.84,85 Osteoporosis introduces the concept of “OP-OA”, emphasizing how subchondral bone fragility exacerbates cartilage calcification and biomechanical imbalances, further driving OA advancement.93,95
Future research should focus on the differential effects of shared inflammatory and metabolic pathways in various joints and the synergistic impact of MetS components like obesity, DM, and HT. Targeting common molecular mechanisms through precision medicine approaches based on inflammatory and metabolic biomarkers can improve clinical outcomes and reduce OA’s socioeconomic burden.
Relationship Between Metabolism-Related Adipokines and OA and Their Pathways of Action
Adipokines
Adipokines are bioactive molecules secreted primarily by adipose tissue, particularly white adipose tissue, and include hormones, cytokines, and chemokines.19 These molecules function through autocrine, paracrine, and endocrine pathways to regulate a range of physiological and pathological processes, including energy metabolism, inflammatory responses, immune regulation, and insulin sensitivity.20 Beyond their critical role in maintaining metabolic homeostasis, adipokines are deeply involved in the pathogenesis of various diseases, such as metabolic syndrome, cardiovascular diseases, and OA.21 Adipokines play a central role in the pathogenesis of OA by modulating chronic inflammation, disrupting cartilage metabolic balance, and impairing bone remodeling.21 These interconnected mechanisms not only accelerate joint degeneration but also underscore the pivotal role of metabolic disturbances in driving OA progression.97–100 Furthermore, the unique contributions of adipokines, independent of mechanical loading, in obesity- and metabolic syndrome-associated OA highlight their potential as key targets for elucidating OA pathophysiology and developing innovative therapeutic strategies.101,102 This understanding offers a comprehensive perspective on the multifaceted roles of adipokines in OA and provides a robust foundation for future research focused on personalized interventions targeting adipokines and their associated signaling pathways.
Adipokines in Osteoarthritis: Clinical Evidence and Therapeutic Implications
Recent clinical studies have demonstrated that adipokines play a pivotal role in the onset and progression of OA, particularly in the regulation of inflammation, cartilage degradation, and metabolic dysregulation. Leptin levels are significantly associated with the severity of joint pain in OA patients, likely by exacerbating inflammation-induced pain.103,104 In contrast, adiponectin levels are reduced in late-stage OA and are inversely correlated with the severity of pain in OA patients, highlighting its potential protective role in joint health.103 Furthermore, leptin concentrations in synovial fluid are strongly correlated with BMI and waist circumference, suggesting that obesity and metabolic syndrome accelerate OA progression through leptin-mediated mechanisms.105 LCN2, a pro-catabolic adipokine, is markedly elevated in the synovial fluid and cartilage of OA patients. It enhances the activity of MMPs, promoting cartilage matrix degradation and intensifying inflammatory responses.106 Similarly, visfatin levels in serum and synovial fluid are strongly associated with inflammation severity and disease progression in OA, further exacerbating joint tissue damage.103 Resistin levels are significantly increased in the synovial fluid and serum of OA patients, accelerating OA pathogenesis by promoting extracellular matrix degradation and the release of pro-inflammatory cytokines.103,107 Additionally, adipsin levels are significantly associated with lateral cartilage volume loss in the knee, suggesting its role in structural joint damage.108 OPN levels in serum are strongly correlated with OA severity, particularly in the early stages of the disease, highlighting its potential as a biomarker for early diagnosis and intervention.104 RBP4 is highly expressed in the serum and synovial fluid of OA patients, with levels significantly associated with matrix metalloproteinase (MMP) activity and pro-inflammatory cytokines, underscoring its critical role in cartilage degradation and inflammation.109 Omentin-1 levels are significantly reduced in the plasma and synovial fluid of OA patients. This reduction is closely linked to joint pain, stiffness, and advanced radiographic severity, indicating a potential protective role for omentin-1 in joint health.110 Metrnl levels are notably lower in late-stage OA patients compared to those in earlier stages, with higher Metrnl levels in synovial fluid inversely correlated with MMP-13, a key marker of cartilage degradation.111 These findings suggest that Metrnl may protect cartilage and mitigate inflammation. Conversely, nesfatin-1 levels are significantly elevated in the serum of OA patients, and its synovial fluid concentrations are positively correlated with pro-inflammatory cytokine IL-18, further supporting its role as a pro-inflammatory mediator.112
These findings collectively underscore that adipokines are not only critical regulators of inflammation and metabolic processes in OA progression but also represent promising diagnostic and therapeutic targets. This evidence provides a robust foundation for developing personalized treatment strategies based on adipokine levels, offering new avenues for precision medicine in OA management.
Therapeutic Potential of Adipokines in OA Treatment
Adipokines have recently emerged as promising therapeutic targets for OA, offering innovative strategies to mitigate disease progression by modulating inflammation, cartilage degradation, and metabolic dysregulation. Leptin, a pro-inflammatory adipokine, has been targeted through therapeutic approaches such as leptin receptor antagonists (eg, Allo-aca) and leptin analogs (eg, Metreleptin), which have shown potential in alleviating inflammation and addressing metabolic disorders by modulating leptin signaling pathways.113,114 In contrast, adiponectin exhibits anti-inflammatory and chondroprotective properties. Its receptor agonist, AdipoRon, demonstrated efficacy in preclinical models by significantly reducing inflammation and preserving cartilage integrity.115 Similarly, visfatin, another pro-inflammatory mediator associated with OA, has been targeted using the specific inhibitor FK866, which has shown promise in suppressing inflammatory responses and represents a potential therapeutic target.116 Additionally, RBP4 exacerbates metabolic dysregulation in OA. The RBP4 inhibitor Fenretinide has demonstrated efficacy in mitigating metabolic disturbances, highlighting its potential as a viable treatment option for OA.117
These findings underscore the pivotal role of adipokines in OA pathophysiology and highlight their targeted therapeutic potential. Future research should prioritize validating these therapeutic strategies through rigorous clinical trials and evaluating their integration into precision medicine frameworks.
Leptin’s Role in OA
Leptin was first identified as a product of the ob gene in 1994;118 it is one of the most intensively studied adipokines, exhibiting pleiotropic properties and being mainly involved in both pro-inflammatory and bone metabolism in the pathomechanisms of OA.119–121 Inflammatory factors play an important role in the developmental process of inflammation in OA, and IL-1, IL-6, TNFα, and IL-17 are the major cytokines involved in the pathogenesis of OA.122 Furthermore, some studies have found that the levels of IL-6, IL-18, and leptin are significantly correlated with the severity of post-traumatic osteoarthritis (PTOA), with the combination of IL-6 and leptin being the most discriminatory biomarker of PTOA.123 IL-1β is the factor most correlated with leptin and has been extensively studied, with reports indicating that levels of both leptin and IL-1β correlate with OA.124 Both leptin and IL-6 significantly contribute to the development of OA through the JAK-STAT pathway, a crucial element in OA pathogenesis.125 Both molecules operate via the JAK-STAT3 pathway and play roles in OA-associated regulatory mechanisms; notably, leptin not only activates the OBRl receptor, which subsequently activates the IRS-1, PI3K, Akt, and AP-1 pathways enhancing IL-6 expression.126 Specifically, in fibroblasts of the temporomandibular joint (TMJ-SFs), leptin engages the JAK2/STAT3 or p38 MAPK or PI3K/Akt signalling pathway and binds to the leptin-specific receptor (Ob-Rb) in the TMJ-SFs to regulate IL-6 production in vitro.127 Furthermore, leptin, in synergy with IL-1β, prompts chondrocytes to secrete pro-inflammatory agents such as IL-6, IL-8, nitric oxide, and cyclooxygenase-2 and modulates IL-6 and IL-8 production through CD4+ T cells.128–130 Furthermore, leptin regulates bone metabolism,125,131 in correlation with the extent of cartilage destruction;132 bone metabolism is also partially affected. Inflammatory factors such as TNF-α, IL-1, and IL-6 notably induce MMP and prostaglandin production and inhibit proteoglycan and type II collagen synthesis, therefore, they play a key role in cartilage matrix degradation and bone resorption in OA.133 Leptin and its downstream factors influence bone metabolism via multiple pathways, these include the induction of human ADAMTS-4 in chondrocytes through the mitogen-activated protein kinase and NF-kB signalling pathways; similarly, leptin promotes the expression of ADAMTS-4 and ADAMTS-5 in human chondrocytes, which are implicated in joint damage and the onset of OA.134 Furthermore, leptin induces cellular senescence in OA chondrocytes by activating the mTOR pathway. Additionally, a high level of Ob-Rb expression accelerates chondrocyte senescence through the leptin pathways in OA.135 Moreover, leptin enhances VCAM-1 expression in cartilage cells via the kinase kinases JAK2, PI3K, and AMPK, leading to accelerated cartilage degradation by promoting leukocyte and monocyte infiltration in inflamed joints.136 Leptin increases MMP production via the JAK2 signal transducer and activator of STAT3 signalling pathway, which has a catabolic effect on OA cartilage and promotes apoptosis.137 Finally, leptin acts through the JAK2/STAT3 signalling pathway to inhibit chondrogenicity and prevent chondrocyte apoptosis.138 Factors downstream of leptin also play a partial role in bone metabolism, with DUSP19 downstream inhibiting chondrocyte apoptosis by dephosphorylating JNK.139 Furthermore, LOXL3, downstream of leptin, stimulates chondrocyte apoptosis and inhibits chondrocyte autophagy.137 Taken together, the evidence increasingly supports the potential role of leptin in OA.122,140,141 Leptin and its receptors are critical targets for intervention in OA.142
Lipocalin’s Role in OA
Lipocalin is a metabolism-related adipokine that regulates lipid metabolism, bone metabolism, and glucose homeostasis.16,143 It has been found that in joints, joint adipose tissue and synovium in patients with inflammatory joint diseases are important sources of lipocalin.144,145 Current research highlights the significant relationship between lipocalin, adipose tissue, and synovium in patients with inflammatory joint diseases. The association between lipocalin and OA remains a subject of debate, with various studies indicating that lipocalin levels are positively correlated with the development of OA and associated with OA joint pain.16,142,146,147 However, the underlying mechanisms by which lipocalin contributes to OA are still not fully understood and warrant further investigation.148 Notably, lipocalin has been shown to correlate positively with IL-6 levels and enhance IL-6 production in synovial fibroblasts via the AdipoR1 receptor/AMPK/p38/IKK alpha beta and NF-κB signalling pathways.149 This process is crucial for the pathogenesis of OA.150 Lipocalin has also been found to increase VCAM-1 expression in chondrocytes through kinases such as JAK2, PI3K, and AMPK, accelerating chondrocyte degradation by inducing infiltration of leukocytes and monocytes in the inflamed joints.136 However, other studies have indicated that lipocalin levels are negatively correlated with the severity of OA and exhibit an anti-inflammatory role in its development. Notably, lipocalin levels were observed to decrease significantly with the increasing severity of Kellgren-Lawrence OA; furthermore, the concentrations of lipocalin in blood and synovial fluid were significantly and negatively correlated with the grading of OA, leading researchers to suggest that lipocalin may serve a protective role in OA.151 Feng et al reviewed the protective mechanisms of lipocalin in OA in terms of both apoptosis and autophagy.143 Liu et al found that lipocalcin activated the AdipoR1/AMPK/PKC pathway to reduce endoplasmic reticulum stress-induced apoptosis and reduced apoptosis by regulating anti-apoptotic proteins, such as Bcl-2, in mouse adipose tissues, thereby reducing the severity of OA.152 He and Duan discovered that LipocalinRon induces autophagy to mitigate cartilage calcification in OA, where He demonstrated that lipocalcin activates autophagy by mediating the AMPK-mTOR signalling pathway.27 It has been proposed that lipocalcin levels do not correlate with OA development.50 Lipofuscin-2 (Lipofectin) also plays a critical role in OA development. Lipocalin-2 (LCN2), also known as neutrophil gelatinase-associated lipocalin, is identified as a novel adipokine with catabolic functions in OA.153,154 Most studies indicate that LCN2 serves as a biomarker for cartilage degradation in OA, and complexes comprising LCN2 and MMP9 are prevalent in synovial fluid from patients with OA, contributing to matrix degradation and OA exacerbation.155,156 However, other research has shown that while LCN2 expression is elevated in OA chondrocytes and cartilage, its overexpression does not change the expression levels of metabolic enzymes involved in matrix degradation, such as catabolic MMP3 or anabolic chondrogenic matrix molecules; thus, its increased levels are neither sufficient nor necessary for cartilage destruction in mouse OA.157 These conflicting findings indicate that lipocalin and LCN2 have dual or complex roles in OA, highlighting the uncertainty of their impact on OA and underscoring the need for further investigations.
Resistin’s Role in OA
The primary source of resistin is white adipose tissue, which can be produced by macrophages as well as by cartilage itself.96,157 Resistin plays an important role in synovial inflammation and cartilage degradation.158 It is a highly potent pro-inflammatory cytokine that elevates the production of various inflammatory factors, including IL-1, TNF, and other inflammatory factors.159,160 It has been observed that resistin facilitates the release of pro-inflammatory factors through multiple pathways.159 It suppresses the expression of miR-149 and boosts the expression of TNF-α and IL-1β via MEK and ERK signalling.161 Nirupama et al discovered that the enhanced secretion of pro-inflammatory cytokines could also be mediated by NF-κB;162 resistin can further activate the p38-MAPK and NF-κB signalling pathways in human OA chondrocytes by binding to CAP1, fostering the expression of pro-inflammatory cytokines (CCL3, CCL4) and matrix-degrading enzymes (MMP-13, ADAMTS-4), and the release of these substances disrupts intra-articular homeostasis, leading to synovitis in knee joints and cartilage degeneration.163 Moreover, resistin induces the expression of pro-inflammatory cytokines as well as miR-34a and miR-146a, which mediate mucin-induced oxidative stress in OA through the NF-κB pathway.159 Chen et al also noted that resistin inhibits the synthesis of miR-381 via the PKCα, p38, and JNK signalling pathways, thereby influencing the expression of VCAM-1 and the adherence of monocytes to OASFs, in turn, impacts OA progression.164 Some studies have found that resistin levels are positively associated with cartilage defects in OA,165,166 promoting the generation of bone nodules.167 Moreover, some researchers have found that resistin in OA bone fragments are mediated by p38 MAPK, which increases the release of pro-inflammatory mediators from osteoclasts and chondrocytes, aggravating the process of OA.21 Additionally, resistin is known to be a key component in the development and growth of bone mineralisation. Resistin inhibits cartilage synthesis by inducing the expression of pro-inflammatory factors such as degradative enzymes through the binding to Toll-like receptor 4 and adenylyl cyclase-associated protein 1 receptor;168 moreover, resistin stimulates the significant overexpression of miR-34a, leading to apoptosis in OA chondrocytes and limiting proliferation.169 Additionally, resistin promotes the expression of MMP-1 and MMP-13 in chondrocytes and increases Col2a1 mRNA, the primary collagen synthesized by these cells, thereby contributing to cartilage degradation.169 Resistin has also been associated with OA and joint pain.142,170,171 Notably, it has been found that garlic supplementation can reduce the severity of pain in women who are overweight or women with obesity suffering from OA, possibly through a reduction in resistin.172 Therefore, resistin is a critical factor in the severity of OA and cartilage degeneration of the knee joint. Resistin, as a potential biomarker of knee OA disease severity and cartilage degeneration,173,174 is closely related to the course of OA,146,175,176 and further in-depth studies are needed to elucidate its effect on disease outcome.
Osteopontin’s Role in OA
Osteopontin (OPN) is an extracellular matrix glycoprotein that plays an important role in the release of inflammatory factors and bone remodelling in OA.177,178 The expression of OPN is regulated by the β-catenin/TCF-4 pathway and miRNA-127-5p.179,180 Currently, several studies have concluded that OPN is significantly associated with the level of severity of OA.104,181 OPN can promote the expression of metalloproteinase 13 (MMP13) in OA through the NF-kB signalling pathway,182 and its deficiency induces the secretion of pro-inflammatory cytokines, such as COL10A1, IL-1β, IL-6, IL-8, and TNF- β, which exacerbate the progression of OA.183,184 Recent studies have shown that OPN plays a crucial role in bone metabolism.185 Moreover, the overexpression of many inflammatory factors also causes an imbalance in bone metabolism. Overexpression of MMP-1 degrades the main component of type II collagen of cartilage matrix proteins in mice;186 the release of TNF-α, IL-6, and IL-1β induces apoptosis of chondrocytes.187 Furthermore, IL-1β, OPN, p53 upregulation, and COL1A1 and COL2A1 overexpression significantly inhibit chondrocyte viability and migration, enhance apoptosis, and induce cartilage damage.188 Notably, OPN promotes NF-κB signalling, accelerates chondrocyte proliferation, and thus induces OA in rats.189 Additionally, OPN-induced expression of VEGF in articular cartilage causes severe vascular invasion of cartilage and exacerbates the process of OA.190
Visfatin’s Role in OA
Visfatin, an adipokine, is negatively correlated with the severity of OA.169,191 It enhances inflammation in OA, as demonstrated by Yang, who reported increased levels of inflammatory markers such as MMP3 and MMP13 in chondrocytes.192 Furthermore, Laiguillon observed that visfatin was implicated in pro-inflammatory activation between chondrocytes and osteoblasts, significantly inducing IL-6 release.193 Moreover, Cheleschi noted that visfatin substantially upregulated the expression of inflammatory factors, including IL-1β, IL-6, TNF-α, MMP-1, and MMP-13.159 In this regard, Han and other scholars have provided a more comprehensive summary of the role of visfatin in OA: visfatin affects the differentiation of mesenchymal stem cells (MSCs) to adipocytes or osteoblasts by increasing the production of MMPs and ADAMTS, leading to alterations in cartilage and bone tissue; induces the production of inflammatory factors, such as IL-6 and TNF-α, and promotes an inflammatory state; and inhibits the production of osteoblasts. Osteoclastogenesis, which may promote bone regrowth formation in the context of inflammatory diseases.194
Adipsin’s Role in OA
Adipsin, discovered in 1987, is produced by adipocytes through activation of PPAR.195–197 Notably, lipocalin deficiency protects joint tissues from the progression of OA.198 Adipsin levels are significantly elevated in serum, SF, and cartilage in patients with OA.199 However, the pathway of action between adipsin and OA remains unclear. Adipsin levels have been significantly associated with cartilage volume loss in the lateral compartment of the knee and correlated with the incidence of Total Knee Arthroplasty.140,200
Fatty Acid Binding Protein 4’s Role in OA
Fatty acid binding protein 4 (FABP4), also known as adipocyte FABP, is involved in lipolysis and is secreted by macrophages and adipocytes.93,201 In patients with OA, FABP4 levels negatively correlate with cartilage thickness and have been identified as playing a role in cartilage degradation.202–205 Furthermore, FABP4 has been found to activate the NF-κB signalling pathway via PPARγ, enhancing IL-1β-induced inflammation, oxidative stress, apoptosis, and extracellular matrix degradation in chondrocytes;thus, FABP4 is implicated in promoting chondrocyte degeneration and plays a significant role in the progression of OA.206,207
Nesfatin-1’s Role in OA
Nesfatin-1, an adipokine, is pivotal in OA development by modulating inflammatory mediators and chondrocytes, though its role in inflammation is debated. This adipokine notably enhances rat chondrocytes’ collagen type II alpha-1 chain (Col2a1) expression and diminishes several inflammatory agents, such as MMPs, cyclooxygenase-2, nitric oxide, prostaglandin E2, and IL-6; additionally, reduces chondrocyte apoptosis, thus safeguarding against OA.112,208 Conversely, most studies have demonstrated a significant positive correlation between nesfatin-1 levels and OA severity.209,210 Furthermore, Lee reported that nesfatin-1 contributes to the production of inflammatory cytokines, particularly promoting IL-1β production in osteoarthritic synovial fibroblasts by suppressing miR-204-5p synthesis through the AP-1 and NF-κB pathway.211 In contrast, some studies suggest that Nesfatin-1, by blocking the activation of the RhoA/ROCK pathway, prevents excessive autophagy in OA cartilage and enhances chondrocyte cytoskeletal integrity.212 Thus, Nesfatin-1 is pivotal in OA progression, yet its interactions with inflammatory factors are underexplored, warranting further investigation as a potential therapeutic target for OA.
Serpin Peptidase Inhibitor, Clade E, Member 2’s Role in OA
Serpin peptidase inhibitor, clade E, member 2 (Serpin E2) exhibits beneficial effects on the progression of OA, offering chondroprotective benefits, inhibiting cartilage degradation, and preventing joint destruction in rabbits.213,214 Crucially, the efficacy of Serpin E2 hinges on its interaction with MMPs.215,216 Among these, MMP-13 emerges as a critical collagenase involved in cartilage catabolism in OA. Santoro et al demonstrated that Serpin E2 obstructs IL-1-induced MMP-13 expression in chondrocytes via pathways involving ERK, NF-κB, and AP-1, thereby mitigating cartilage catabolism.215
Progranulin’s Role in OA
PGRN is an adipokine with multifaceted functions, contributing to chondrogenesis and anti-inflammation in OA and preventing further OA deterioration.217,218 Atsttrin, a derivative of PGRN, protects against early OA.219 Autophagy serves as a protective mechanism for normal cartilage,220 and its loss is linked to pathological changes in OA.221 PGRN regulates chondrocyte autophagy by modulating the interaction with the ATG5-ATG12 complex, thus providing a protective effect on cartilage.222 Furthermore, PGRN interacts with the inositol-requiring enzyme 1α, an inositol-requiring enzyme, enhancing the expression of collagen type 2 and maintaining collagen homeostasis to protect cartilage.223 As an antagonist of TNF-α signalling, PGRN plays a crucial role in the pathogenesis of inflammatory arthritis in mice, antagonizes TNF-α, and protects against cartilage destruction in OA.224,225 Additionally, PGRN restores chondrocyte metabolic homeostasis by activating the ERK1/2 signalling pathway and elevating anabolic biomarkers, including collagen type 2 and Aggrecan.226
Chemerin’s Role in OA
Chemerin exhibits a pro-inflammatory effect, and in vitro studies suggest its role in inflammatory lesions and cartilage degeneration in OA.17,227,228 This protein is pro-inflammatory in OA; notably, chemerin induces the release of inflammatory mediators such as IL-6, TNF-a, and metalloproteinases from macrophages and chondrocytes.229 Furthermore, chemerin elevates TLR4 expression and triggers CCL2 release from synovial fibroblasts, enhancing leukocyte migration to inflammation sites and amplifying inflammatory signalling in chondrocytes.155 Moreover, chemerin intensifies inflammatory signals in macrophages and chondrocytes,230 influences chondrocyte metabolism by boosting AKT/ERK phosphorylation, and decreases chondrocyte proliferation, worsening OA symptoms.231 Therefore, while chemerin’s role in OA development is acknowledged, its precise pathways and mechanisms warrant further study.
Wnt-1-Induced Signalling Pathway Protein 2’s Role in OA
Emerging adipokines, such as Wnt-1-induced signalling pathway protein 2 (WISP-2) or CCN5, have been linked to OA progression. Found in the chondrocytes and synovial membranes of OA patients, WISP-2 significantly influences OA pathogenesis.232–234 Studies indicate that WISP-2 counters the effects of IL-1β on MMP-13 and ADAMTS-5, and decreases IL-6 and IL-8 levels in OA chondrocytes through the WNT/β-catenin pathway, thus potentially slowing OA progression.235
Visceral Adipose Tissue-Derived Serpin’s Role in OA
Visceral adipose tissue-derived Serpin (vaspin), which is expressed at multiple sites and possesses pleiotropic properties, has also emerged.236 Serum vaspin levels are lower in OA patients than in healthy controls.237 Vaspin may inhibit the release of pro-inflammatory factors and certain adipokines such as leptin and resistin,238 influencing the inflammatory process in OA and promoting the development of bone mesenchymal stem cells through activation of the PI3K/AKT pathway.239,240 Reduced expression of vaspin inhibits cholesterol synthesis via the miR155/LXRα efflux pathway, contributing to the accumulation of cholesterol in the cartilage and the development of OA.241
Serum Amyloid A’s Role in OA
Serum Amyloid A (SAA) is a newly discovered adipokine; studies have shown that the level of SAA in serum and synovial fluid (SF) of OA patients was higher than normal, and its expression level was positively correlated with the severity of OA.242,243 Furthermore, it was found that SAA induces the release of pro-inflammatory cytokines,244,245 which plays a key role in the inflammatory process of OA and induces the secretion of MMPs by chondrocytes under the control of TGF-b, exacerbating the progression of OA.246
Omentin-1’s Role in OA
Omentin-1 (also known as Intelectin-1) is a newly discovered adipokine, and multiple studies have found that its level is negatively correlated with OA severity.247,248 Omentin-1 displays anti-inflammatory effects in OA, with both omentin-1 and IL-4 levels significantly reduced in OA patients relative to controls. Studies have shown that omentin-1 triggers IL-4-dependent anti-inflammatory responses and M2 macrophage polarization in OA synovial fibroblasts via PI3K, ERK, and AMPK pathways, thereby preventing cartilage degradation and bone erosion.249 Additionally, omentin-1 has been described as a pleiotropic protective adipokine that offers a repair mechanism for chondrocytes in joint tissue. This is done by attenuating IL-1β-induced G1-phase cell cycle block and inhibiting IL-1-induced cellular senescence, thus protecting chondrocytes from senescence.250
Metrnl’s Role in OA
Metrnl, a newly discovered adipokine, has been linked to the pathogenesis of OA. Studies have shown that higher levels of Metrnl correlate with a reduced likelihood of developing OA.251–253 It has also been shown that Metrnl can regulate IL-4 and IL-13 expression levels to exert anti-inflammatory effects;Liu has noted that its anti-inflammatory effects are mediated by inhibiting the PI3K/Akt/NF-κB pathway.254,255
Adipokines as Key Mediators in the Pathogenesis of OA
Adipokines play a pivotal role in the pathogenesis of OA through complex interactions within shared inflammatory and metabolic signaling pathways, including NF-κB, PI3K/Akt, MAPK, and others. These molecules influence critical processes such as chronic inflammation, cartilage metabolism, and bone remodeling, contributing to the progression of OA.
Firstly, Chronic Inflammation Driven by NF-κB and JAK/STAT Pathways: Adipokines such as leptin, resistin, and lipocalin establish a positive feedback loop by co-activating NF-κB and JAK/STAT signaling pathways.21,113,146 This significantly amplifies the release of pro-inflammatory cytokines (eg, IL-6, TNF-α), maintaining a chronic low-grade inflammatory microenvironment in the joint.21,113,146 Such sustained inflammation exacerbates cartilage degradation, synovial inflammation, and overall OA progression. Secondly, Cartilage Metabolic Imbalance via PI3K/Akt and MAPK Pathways: Leptin, Visfatin, and Nesfatin-1 also converge on the PI3K/Akt and MAPK pathways to regulate chondrocyte metabolic activity.104,112,194,197 This disrupts the balance between cartilage matrix synthesis and degradation, accelerating cartilage degeneration and further driving OA progression.
Thirdly, Bone Remodeling Imbalance via ERK and mTOR Pathways: adipokines such as leptin and osteopontin modulate osteoclast and osteoblast activity through ERK and mTOR signaling pathways.118,119,177 This results in increased bone resorption, inhibited bone formation, and progressive structural deterioration of bone tissue, destabilizing joint integrity and worsening OA outcomes. Lastly, Insufficient Anti-Inflammatory Protection via PPARγPathways: anti-inflammatory adipokines, including metrnl, and FABP4, exhibit protective effects by mitigating inflammation and preserving joint structures through Wnt/β-catenin and PPARγ pathways.116,239 However, these protective mechanisms are often insufficient to counteract the strong pro-inflammatory signals driven by other adipokines, limiting their ability to effectively slow OA progression.
Chronic low-grade inflammation and bone destruction, mediated by key adipokines, are fundamental to OA pathogenesis within the framework of MetS. Pro-inflammatory adipokines, including leptin, resistin, and osteopontin, exacerbate cartilage degradation and synovial inflammation by activating signaling pathways such as NF-κB, PI3K/Akt, and WNT/β-catenin, leading to chondrocyte catabolism and inflammatory cascades.21,128,183,256 Conversely, protective adipokines like Serpin E2, PGRN, and WISP-2 counteract these processes by suppressing cartilage degradation and enhancing anti-inflammatory mechanisms, promoting tissue homeostasis.213,215,217 Notably, adipokines such as lipocalin, adipsin, and nesfatin-1 exhibit context-dependent dual roles, either promoting inflammation or supporting cartilage stability, reflecting their complex contributions to OA progression.112,152,200 Understanding these diverse functional phenotypes and underlying molecular mechanisms is critical for identifying therapeutic targets and advancing treatments for metabolic OA. Epidemiological studies strongly reinforce the link between adipokines and OA, underscoring their pivotal roles in disease initiation and progression. Tables 2 and 3, along with Figure 3, delineate the functional and clinical relevance of various adipokines, illustrating both shared and distinct mechanisms underlying OA pathophysiology. Pro-inflammatory adipokines, such as SAA and chemerin, amplify cytokine cascades, serving as critical markers of OA progression.217,229 In contrast, anti-inflammatory adipokines like omentin-1 and Metrnl show promise in cartilage repair and symptom alleviation by inhibiting NF-κB signaling and promoting anti-inflammatory pathways.247,255,
 |
Table 2 The Role of Different Adipokines in the Pathogenesis of OA |
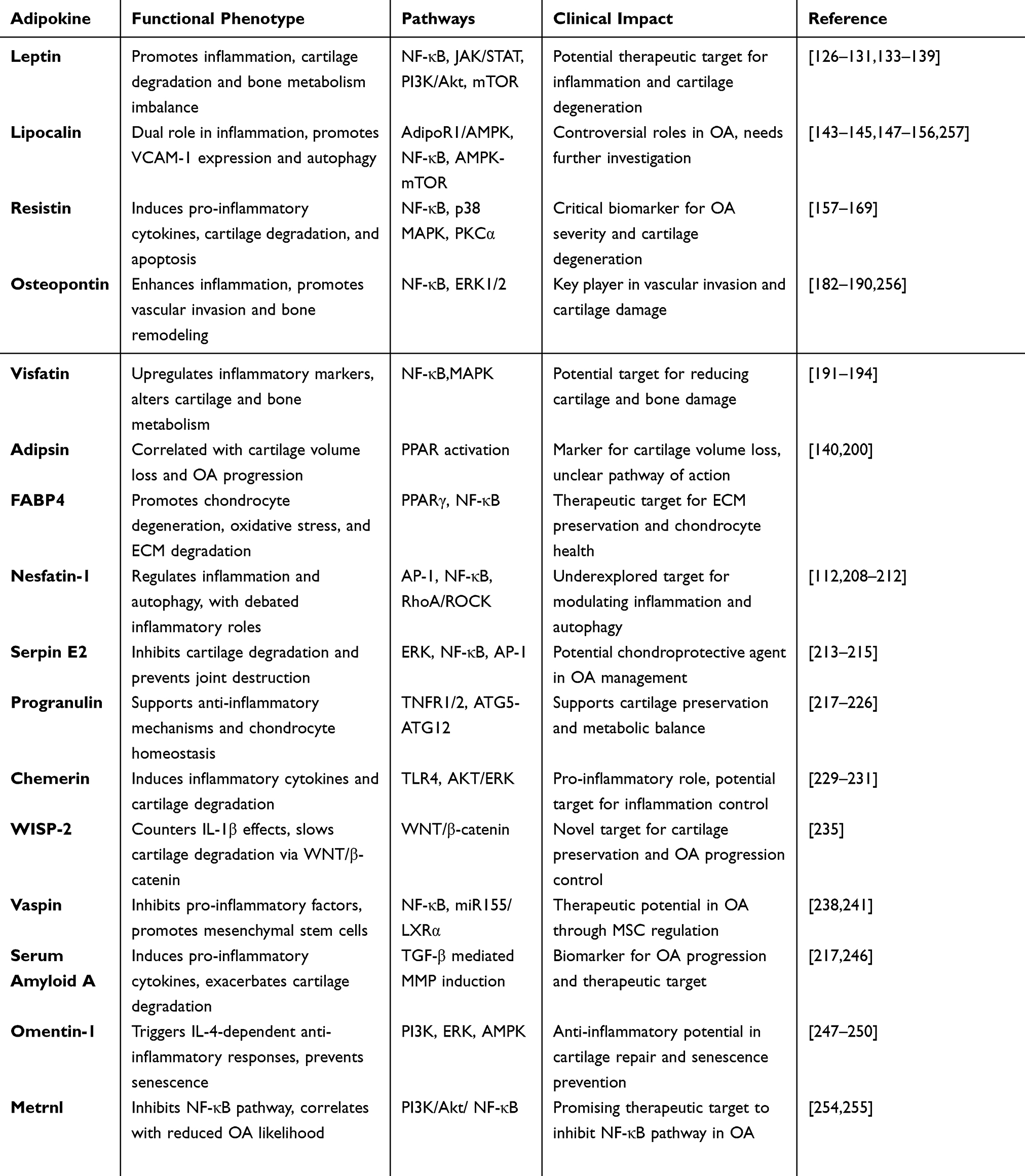 |
Table 3 Adipokines in Osteoarthritis: Multifaceted Roles, Mechanistic Pathways, and Clinical Implications |
 |
Figure 3 Pathways of action of multiple adipokines triggering OA. |
Conclusion
OA is a complex degenerative joint disease significantly influenced by metabolic dysregulation, with metabolism-related adipokines playing a pivotal role in the pathogenesis of MetS-OA. While previous studies have highlighted the crucial roles of metabolic dysfunction and adipokines in OA progression, the precise mechanisms and interactions remain incompletely understood. The relationship between adipokines and OA is multifaceted, with these molecules exerting varying effects at different stages of the disease, from promoting inflammation and cartilage degradation to potentially modulating repair processes. This complexity underscores the importance of further investigating how adipokines influence OA progression and their roles in both early and advanced stages of the disease.
Future research should focus on elucidating the diverse functions of adipokines at different stages of OA, particularly their dual roles in inflammation, metabolism, and tissue remodeling. A deeper understanding of the complex mechanisms underlying adipokine actions is essential for the development of targeted therapies. Targeting specific adipokines or their receptors could not only alleviate symptoms but also slow disease progression, providing long-term therapeutic benefits for OA patients. Moreover, combining adipokine-targeted treatments with other therapeutic approaches, such as cartilage repair or immune modulation, may offer a more comprehensive and effective disease management strategy.
Abbreviations
OA, Osteoarthritis; MetS, Metabolic syndrome; MetS-OA, Metabolic syndrome-associated osteoarthritis; BMI, Body mass index; IL-6, Interleukin-6; TNFα, Tumour necrosis factor alpha; DM, Diabetes mellitus; PGRN, Progranulin; ROS, reactive oxygen species; HT, Hypertension; DLs, Dyslipidaemias; OP, Osteoporosis ; Nrf-2, nuclear factor erythroid 2-related factor 2; AGEs, advanced glycation end-products; PTOA, post-traumatic osteoarthritis; TMJ-SFs, temporomandibular joint; Ob-Rb, leptin-specific receptor; MMP, metalloproteinase; LCN2, Lipocalin-2; OPN, Osteopontin; MMP13, Metalloproteinase 13; MSCs, Mesenchymal stem cells; FABP4, Fatty acid binding protein 4; Col2a1, Collagen type II alpha-1 chain; Serpin E2, Serpin peptidase inhibitor, clade E, member 2; WISP-2, Wnt-1-induced signalling pathway protein 2; Vaspin, Visceral adipose tissue-derived Serpin; SAA, Serum Amyloid A.
Acknowledgments
The figures in this manuscript were created by Figdraw. We thank Figdraw for providing this professional drawing platform.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This study was funded by the Shandong Natural Science Foundation (ZR2022MH063 to Wei Li), supported by the National Natural Science Foundation of China (82171521, 82371539 to Chen Li), and received support from the Special Funds of the Taishan Scholars Project of Shandong Province (No. tsqn202211368 to Chen Li).
Disclosure
The authors declare that they have no competing interests.
References
1. Liu S, Pan Y, Li T. et al. The role of regulated programmed cell death in osteoarthritis: from Pa thogenesis to therapy. Int J Mol Sci. 2023;24(6):5364. doi:10.3390/ijms24065364
2. Colletti A, Cicero AFG. Nutraceutical approach to chronic osteoarthritis: from molecular resea rch to clinical evidence. Int J Mol Sci. 2021;22(23):12920. doi:10.3390/ijms222312920
3. Rim YA, Nam Y, Ju JH. The role of chondrocyte hypertrophy and senescence in osteoarthritis i nitiation and progression. Int J Mol Sci. 2020;21(7):2358. doi:10.3390/ijms21072358
4. Kolasinski SL, Neogi T, Hochberg MC, et al. American college of rheumatology/arthritis foundation guideline f or the management of osteoarthritis of the hand, hip, and knee. Arthritis Care Res. 2019;72(2):149–162. doi:10.1002/acr.24131
5. Srikanth VK, Fryer JL, Zhai G, et al. A meta-analysis of sex differences prevalence, incidence and severity of osteoarthritis. Osteoarthritis Cartilage. 2005;13(9):769–781. doi:10.1016/j.joca.2005.04.014
6. de Klerk BM, Schiphof D, Groeneveld FPMJ, et al. No clear association between female hormonal aspects and osteoarthriti s of the hand, hip and knee: a systematic review. Rheumatology. 2009;48(9):1160–1165. doi:10.1093/rheumatology/kep194
7. Prieto-Alhambra D, Judge A, Javaid MK, et al. Incidence and risk factors for clinically diagnosed knee, Hip and hand osteoarthritis: influences of age, gender and osteoarthritis affectin g other joints. Ann Rheum Dis. 2014;73(9):1659–1664. doi:10.1136/annrheumdis-2013-203355
8. Li D, Li S, Chen Q, et al. The prevalence of symptomatic knee osteoarthritis in relation to age, sex, area, region, and body mass index in China: a systematic review a nd meta-analysis. Front Med. 2020;7:304. doi:10.3389/fmed.2020.00304
9. Reyes C, Garcia-Gil M, Elorza JM, et al. Socio-economic status and the risk of developing hand, hip or knee ost eoarthritis: a region-wide ecological study. Osteoarthritis Cartilage. 2015;23(8):1323–1329. doi:10.1016/j.joca.2015.03.020
10. Hunter DJ, Schofield D, Callander E. The individual and socioeconomic impact of osteoarthritis. Nat Rev Rheumatol. 2014;10(7):437–441. doi:10.1038/nrrheum.2014.44
11. Hootman JM, Helmick CG, Barbour KE, et al. Updated projected prevalence of self-reported doctor-diagnosed arthrit is and arthritis-attributable activity limitation among US adults, 2015–2040. Arthritis Rheumatol. 2016;68(7):1582–1587. doi:10.1002/art.39692
12. Hunter DJ, March L, Chew M. Osteoarthritis in 2020 and beyond: a lancet commission. Lancet. 2020;396(10264):1711–1712. doi:10.1016/S0140-6736(20)32230-3
13. Crofford LJ. Use of NSAIDs in treating patients with arthritis. Arthritis Res Ther. 2013;3(Suppl 3):S2. doi:10.1186/ar4174
14. Zhao SS, Karhunen V, Morris AP, et al. ADAMTS5 as a therapeutic target for osteoarthritis: Mendelian randomis ation study. Ann Rheum Dis. 2022;81(6):903–904. doi:10.1136/annrheumdis-2021-222007
15. Sampath SJP, Venkatesan V, Ghosh S, et al. Obesity, metabolic syndrome, and osteoarthritis-an updated review. 2023;Curr Obes Rep. 12(3):308–331. doi:10.1007/s13679-023-00520-5
16. Fan J, Zhu J, Sun L, et al. Causal association of adipokines with osteoarthritis: a Mendelian randomization study. Rheumatology. 2021;60(6):2808–2815. doi:10.1093/rheumatology/keaa719
17. Huang X, Liu J, Huang W. Identification of S100A8 as a common diagnostic biomarkers and explori ng potential pathogenesis for osteoarthritis and metabolic syndrome. Front Immunol. 2023. 14;(14):1185275. doi:10.3389/fimmu.2023.1185275
18. Jiménez-Muro M, Soriano-Romaní L, Mora G, et al. The microbiota-metabolic syndrome axis as a promoter of metabolic oste oarthritis. Life Sci. 2023;329:121944. doi:10.1016/j.lfs.2023.121944
19. Derosa G, Catena G, Gaudio G, et al. Adipose tissue dysfunction and metabolic disorders: is it possible to predict who will develop type 2 diabetes mellitus? Role of markErs in the progreSsion of dIabeteS in obese paTIeNts (The RESISTIN trial). Cytokine. 2020;127:154947. doi:10.1016/j.cyto.2019.154947
20. Landecho MF, Tuero C, Valentí V, et al. Relevance of leptin and other adipokines in obesity-associated cardiov ascular risk. Nutrients. 2019;11(11):2664. doi:10.3390/nu11112664
21. Xie C, Chen Q. Adipokines: new therapeutic target for osteoarthritis? Curr Rheumatol Rep. 2019;21(12):71. doi:10.1007/s11926-019-0868-z
22. Azamar-Llamas D, Hernández-Molina G, Ramos-ávalos B, et al. Adipokine contribution to the pathogenesis of osteoarthritis. Mediators Inflamm. 2017;2017:5468023. doi:10.1155/2017/5468023
23. Tu C, He J, Wu B, et al. An extensive review regarding the adipokines in the pathogenesis and p rogression of osteoarthritis. Cytokine. 2019;113:1–12. doi:10.1016/j.cyto.2018.06.019
24. Lotti F, Marchiani S, Corona G, et al. Metabolic syndrome and reproduction. Int J Mol Sci. 2021. 22;(4):1988. doi:10.3390/ijms22041988
25. Vrdoljak J, Kumric M, Vilovic M, et al. Can fasting curb the metabolic syndrome epidemic? Nutrients. 2022;14(3):456. doi:10.3390/nu14030456
26. Bouillon-Minois J-B, Dutheil F. Biomarker of stress. Metabolic Syndrom Human Health Nutri. 2022;14(14):2935. doi:10.3390/nu14142935
27. He K, Nie L, Ali T, et al. Adiponectin alleviated Alzheimer‐like pathologies via autophagy‐lysosomal activation. Aging Cell. 2021;20(12):e13514. doi:10.1111/acel.13514
28. Rojas M, Chávez-Castillo M, Pirela D, et al. Metabolic syndrome: is it time to add the central nervous system? Nutrients. 2021;13(7):2254. doi:10.3390/nu13072254
29. Weihe P, Weihrauch-Blüher S. Metabolic syndrome in children and adolescents: diagnostic criteria, T herapeutic options and perspectives. Curr Obes Rep. 2019;8(4):5075–5086. doi:10.2147/DDDT.S340410
30. Niu J, Clancy M, Aliabadi P, et al. Metabolic syndrome, its components, and knee osteoarthritis: the frami ngham osteoarthritis study. Arthritis Rheumatol. 2017;69(6):1194–1203. doi:10.1002/art.40087
31. Zheng L, Zhang Z, Sheng P, et al. The role of metabolism in chondrocyte dysfunction and the progression of osteoarthritis. Ageing Res Rev. 2021;66:101249. doi:10.1016/j.arr.2020.101249
32. Puenpatom RA, Victor TW. Increased prevalence of metabolic syndrome in individuals with osteoar thritis: an analysis of NHANES III data. Postgrad Med. 2009;121(6):9–20. doi:10.3810/pgm.2009.11.2073
33. Engström G, Gerhardsson de Verdier M, Rollof J, et al. C-reactive protein, metabolic syndrome and incidence of severe Hip and knee osteoarthritis. A population-based cohort study. Osteoarthritis Cartilage. 2009;17(2):168–173. doi:10.1016/j.joca.2008.07.003
34. Eckel RH, Grundy SM, Zimmet PZ. The metabolic syndrome. Lancet. 2005;365(9468):1415–1428. doi:10.1016/S0140-6736(05)66378-7
35. Xu H, Li X, Adams H, et al. Etiology of metabolic syndrome and dietary intervention. Int J Mol Sci. 2018;20(1):128. doi:10.3390/ijms20010128
36. Spahis S, Borys J-M, Levy E. Metabolic syndrome as a multifaceted risk factor for oxidative stress. Antioxid Redox Signal. 2017;26(9):445–461. doi:10.1089/ars.2016.6756
37. Blüher M. Obesity: global epidemiology and pathogenesis. Nat Rev Endocrinol. 2019;15(5):288–298. doi:10.1038/s41574-019-0176-8
38. Caballero B. Humans against obesity: who will win? Adv Nutr. 2019;10(suppl_1):S4–S9. doi:10.1093/advances/nmy055
39. Piché M-E, Tchernof A, Després J-P. Obesity phenotypes, diabetes, and cardiovascular diseases. Circ Res. 2020;126(11):1477–1500. doi:10.1161/CIRCRESAHA.120.316101
40. Felson DT, Anderson JJ, Naimark A, et al. Obesity and knee osteoarthritis. The Framingham study. Ann Intern Med. 1988 109(1):18–24. doi:10.7326/0003-4819-109-1-18
41. Harasymowicz NS, Clement ND, Azfer A, et al. Regional differences between perisynovial and infrapatellar adipose ti ssue depots and their response to class II and class III obesity in pa tients with osteoarthritis. Arthritis Rheumatol. 2017;69(7):1396–1406. doi:10.1002/art.40102
42. Reyes C, Leyland KM, Peat G, et al. Association between overweight and obesity and risk of clinically diag nosed knee, hip, and hand osteoarthritis: a population-based cohort study. Arthritis Rheumatol. 2016;68(8):1869–1875. doi:10.1002/art.39707
43. Gambari L, Cellamare A, Grassi F, et al. Targeting the inflammatory hallmarks of obesity-associated osteoarthri tis: towards nutraceutical-oriented preventive and complementary thera peutic strategies based on n-3 polyunsaturated fatty acids. Int J Mol Sci. 2023;24(11):9340. doi:10.3390/ijms24119340
44. Murphy L, Schwartz TA, Helmick CG, et al. Lifetime risk of symptomatic knee osteoarthritis. Arthritis Rheum. 2008;59(9):1207–1213. doi:10.1002/art.24021
45. Nedunchezhiyan U, Varughese I, Sun AR, et al. Obesity, inflammation, and immune system in osteoarthritis. Front Immunol. 2022;13(13):907750. doi:10.3389/fimmu.2022.907750
46. Harding GT, Hubley-Kozey CL, Dunbar MJ, et al. Body mass index affects knee joint mechanics during gait differently w ith and without moderate knee osteoarthritis. Osteoarthritis Cartilage. 2012;20(11):1234–1242. doi:10.1016/j.joca.2012.08.004
47. Batushansky A, Zhu S, Komaravolu RK, et al. Fundamentals of OA. an initiative of osteoarthritis and cartilage. obe sity and metabolic factors in OA. Osteoarthritis Cartilage. 2022;30(4):501–515. doi:10.1016/j.joca.2021.06.013
48. Powell A, Teichtahl AJ, Wluka AE, et al. Obesity: a preventable risk factor for large joint osteoarthritis whic h may act through biomechanical factors. Br J Sports Med. 2005;39(1):4–5. doi:10.1136/bjsm.2004.011841
49. Fang T, Zhou X, Jin M, et al. Molecular mechanisms of mechanical load-induced osteoarthritis. Int Ortho. 2021;45:1125–1136. doi:10.1007/s00264-021-04938-1
50. Kroon FPB, Veenbrink AI, de Mutsert R, et al. The role of leptin and adiponectin as mediators in the relationship be tween adiposity and hand and knee osteoarthritis. Osteoarthritis Cartilage. 2019;27(12):1761–1767. doi:10.1016/j.joca.2019.08.003
51. Killock D. Osteoarthritis: the influence of obesity on OA-does size matter or is metabolic dysfunction more important? Nat Rev Rheumatol. 2012;8(2):61. doi:10.1038/nrrheum.2011.221
52. Yao Z, Qi W, Zhang H, et al. Down-regulated GAS6 impairs synovial macrophage efferocytosis and prom otes obesity-associated osteoarthritis. Elife. 2023;12:e83069. doi:10.7554/eLife.83069
53. Klein-Wieringa IR, Kloppenburg M, Bastiaansen-Jenniskens YM, et al. The infrapatellar fat pad of patients with osteoarthritis has an infla mmatory phenotype. Ann Rheumatic Dis. 2011;70(5):851–857. doi:10.1136/ard.2010.140046
54. de Jong AJ, Klein-Wieringa IR, Andersen SN, et al. Lack of high BMI-related features in adipocytes and inflammatory cells in the infrapatellar fat pad (IFP). Arthritis Res. Ther.2017;19:1–12.
55. Sun AR, Panchal SK, Friis T, et al. Obesity-associated metabolic syndrome spontaneously induces infiltration of pro-inflammatory macrophage in synovium and promotes osteoarthritis. PLoS One. 2017;12(8):e0183693. doi:10.1371/journal.pone.0183693
56. Pearson MJ, Herndler-Brandstetter D, Tariq MA, et al. IL-6 secretion in osteoarthritis patients is mediated by chondrocyte-synovial fibroblast cross-talk and is enhanced by obesity. Sci Rep. 2017;7(1):3451. doi:10.1038/s41598-017-03759-w
57. Liu H, Rosol TJ, Sathiaseelan R, et al. Cellular carbon stress is a mediator of obesity-associated osteoarthritis development. Osteoar Cartil. 2021;29(9):1346–1350. doi:10.1016/j.joca.2021.04.016
58. Seow SR, Mat S, Azam AA, et al. Impact of diabetes mellitus on osteoarthritis: a scoping review on biomarkers. Expert Rev Mol Med. 2024;26:e8.
59. Yu MG, Gordin D, Fu J, et al. Protective factors and the pathogenesis of complications in diabetes. Endocr Rev. 2024;45(2):227–252. doi:10.1210/endrev/bnad030
60. Eitner A, Culvenor AG, Wirth W, et al. Impact of diabetes mellitus on knee osteoarthritis pain and physical and mental status: data from the osteoarthritis initiative. Arthr Care Res. 2021;73(4):540–548. doi:10.1002/acr.24173
61. Schett G, Kleyer A, Perricone C, et al. Diabetes is an independent predictor for severe osteoarthritis: results from a longitudinal cohort study. Diabetes Care. 2013;36(2):403–409. doi:10.2337/dc12-0924
62. Eitner A, Pester J, Vogel F, et al. Pain sensation in human osteoarthritic knee joints is strongly enhanced by diabetes mellitus. Pain. 2017;158(9):1743–1753. doi:10.1097/j.pain.0000000000000972
63. Berenbaum F. Diabetes-induced osteoarthritis: from a new paradigm to a new phenotype. Postgraduate Medl j. 2012;88(2012):240–242. doi:10.1136/pgmj.2010.146399rep
64. Laiguillon M-C, Courties A, Houard X, et al. Characterization of diabetic osteoarthritic cartilage and role of high glucose environment on chondrocyte activation: toward pathophysiological delineation of diabetes mellitus-related osteoarthritis. Osteoa Cartil. 2015;23(9):1513–1522. doi:10.1016/j.joca.2015.04.026
65. Piva SR, Susko AM, Khoja SS, et al. Links between osteoarthritis and diabetes: implications for management from a physical activity perspective. Clin Geria Med. 2015;31(1):67–87. doi:10.1016/j.cger.2014.08.019
66. Umpierrez GE, Zlatev T, Spanheimer RGJM. Correction of altered collagen metabolism in diabetic animals with insulin therapy3. Matrix. 1989;9(4):336–342. doi:10.1016/s0934-8832(89)80010-1
67. Neumann J, Hofmann FC, Heilmeier U, et al. Type 2 diabetes patients have accelerated cartilage matrix degeneration compared to diabetes free controls: data from the osteoarthritis initiative. Osteoarth Cartil. 2018;26(6):751–761. doi:10.1016/j.joca.2018.03.010
68. Piñeiro-Ramil M, Burguera EF, Hermida-Gómez T, et al. Reduced levels of H2S in diabetes-associated osteoarthritis are linked to hyperglycaemia, Nrf-2/HO-1 signalling downregulation and chondrocyte dysfunction. Antioxidants. 2022;11(4):628. doi:10.3390/antiox11040628
69. Frey N, Hügle T, Jick S, et al. cartilage. Type II diabetes mellitus and incident osteoarthritis of the hand: a population-based case–control analysis. Osteoarth Cartil. 2016;24(9):1535–1540. doi:10.1016/j.joca.2016.04.005
70. Kuusalo L, Felson DT, Wang N, et al. Metabolic osteoarthritis–relation of diabetes and cardiovascular disease with knee osteoarthritis. Osteoarth Cartil. 2021;29(2):230–234. doi:10.1016/j.joca.2020.09.010
71. Nielen JT, Emans PJ, van den Bemt B, et al. Association of type 2 diabetes mellitus with self-reported knee pain and clinical knee osteoarthritis: the Maastricht study. Diabetes Metab. 2018;44(3):296–299. doi:10.1016/j.diabet.2018.01.013
72. Carey RM, Moran AE, Whelton PKJJ. Treatment of hypertension: a review. JAMA. 2022;328(18):1849–1861. doi:10.1001/jama.2022.19590
73. Yoshimura N, Muraki S, Oka H, et al. Accumulation of metabolic risk factors such as overweight, hypertension, dyslipidaemia, and impaired glucose tolerance raises the risk of occurrence and progression of knee osteoarthritis: a 3-year follow-up of the ROAD study. Osteoarth Cartil. 2012;20(11):1217–1226. doi:10.1016/j.joca.2012.06.006
74. Verdecchia P, Angeli F, Mazzotta G, et al. Treatment strategies for osteoarthritis patients with pain and hypertension. Therap Adv Musculoskelet Dis. 2010;2(4):229–240. doi:10.1177/1759720X10376120
75. Lawrence J. Hypertension in relation to musculoskeletal disorders. Ann. Rheum. Dis.1975;34(5):451–456.
76. Ching K, Houard X, Berenbaum F, et al. Hypertension meets osteoarthritis—revisiting the vascular aetiology hypothesis. Nat Rev Rheumatol. 2021;17(9):533–549. doi:10.1038/s41584-021-00650-x
77. Yeater TD, Griffith JL, Cruz CJ, et al. Hypertension contributes to exacerbated osteoarthritis pathophysiology in rats in a sex-dependent manner. Arth Res Ther. 2023;25(1):7. doi:10.1186/s13075-022-02966-9
78. Lo K, Au M, Ni J, et al. Association between hypertension and osteoarthritis: a systematic review and meta-analysis of observational studies. J Orthop Transl. 2022;32:12–20. doi:10.1016/j.jot.2021.05.003
79. Berberich AJ, Hegele RA. A modern approach to dyslipidemia. Endocr Rev. 2022;43(4):611–653. doi:10.1210/endrev/bnab037
80. Xiong J, Long J, Chen X, et al. Dyslipidemia might be associated with an increased risk of osteoarthritis. Biomed Res. Int2020;2020.
81. Frey N, Hügle T, Jick S, et al. Hyperlipidaemia and incident osteoarthritis of the hand: a population-based case-control study. Osteoarth Cartil. 2017;25(7):1040–1045. doi:10.1016/j.joca.2017.01.014
82. D-x X, Wei J, Zeng C, et al. Association between metabolic syndrome and knee osteoarthritis: a cross-sectional study. BMC Musculoskelet. Disord. 2017;18:1–7.
83. Zhou M, Guo Y, Wang D, et al. The cross-sectional and longitudinal effect of hyperlipidemia on knee osteoarthritis: results from the Dongfeng-Tongji cohort in China. Sci Rep. 2017;7(1):9739. doi:10.1038/s41598-017-10158-8
84. Esteve E, Ricart W, Fernández-Real JM. Dyslipidemia and inflammation: an evolutionary conserved mechanism. Clin Nutr). 2005;24(1):16–31. doi:10.1016/j.clnu.2004.08.004
85. Kontush A, de Faria EC, Chantepie S, et al. A normotriglyceridemic, low HDL-cholesterol phenotype is characterised by elevated oxidative stress and HDL particles with attenuated antioxidative activity. Atherosclerosis. 2005;182(2):277–285. doi:10.1016/j.atherosclerosis.2005.03.001
86. Inoue R, Ishibashi Y, Tsuda E, et al. Medical problems and risk factors of metabolic syndrome among radiographic knee osteoarthritis patients in the Japanese general population. J. Orthop. Sci. 2011;16:704–709. doi:10.1007/s00776-011-0157-9
87. Williams SA, Daigle SG, Weiss R, et al. Economic burden of osteoporosis-related fractures in the US Medicare population. Ann Pharmacother. 2021;55(7):821–829. doi:10.1177/1060028020970518
88. Guan Z, Luo L, Liu S, et al. The role of depletion of gut microbiota in osteoporosis and osteoarthritis: a narrative review. Front Endocrinol. 2022;13:847401. doi:10.3389/fendo.2022.847401
89. Visconti VV, Cariati I, Fittipaldi S, et al. DNA methylation signatures of bone metabolism in osteoporosis and osteoarthritis aging-related diseases: an updated review. Int J Mol Sci. 2021;22(8):4244. doi:10.3390/ijms22084244
90. Riegger J, Schoppa A, Ruths L, et al. Oxidative stress as a key modulator of cell fate decision in osteoarthritis and osteoporosis: a narrative review. Cell Mol Biol Letters. 2023;28(1):76. doi:10.1186/s11658-023-00489-y
91. Yokota S, Ishizu H, Miyazaki T, et al. Osteoporosis, osteoarthritis, and subchondral insufficiency fracture. Recent Insights. 2024;12(4):843.
92. Cooper C, Cook P, Osmond C, et al. Osteoarthritis of the Hip and osteoporosis of the proximal femur. Ann Rheum Dis. 1991;50(8):540–542. doi:10.1136/ard.50.8.540
93. Guo D, Lin C, Lu Y, et al. FABP4 secreted by M1-polarized macrophages promotes synovitis and angiogenesis to exacerbate rheumatoid arthritis. Bone Res. 2022;10(1):45. doi:10.1038/s41413-022-00211-2
94. Bellido M, Lugo L, Roman-Blas J, et al. Improving subchondral bone integrity reduces progression of cartilage damage in experimental osteoarthritis preceded by osteoporosis. Osteoarth Cartilage. 2011;19(10):1228–1236. doi:10.1016/j.joca.2011.07.003
95. Chu L, Liu X, He Z, et al. Articular cartilage degradation and aberrant subchondral bone remodeling in patients with osteoarthritis and osteoporosis. J Bone Min Res. 2020;35(3):505–515. doi:10.1002/jbmr.3909
96. Wada H, Aso K, Izumi M, et al. The effect of postmenopausal osteoporosis on subchondral bone pathology in a rat model of knee osteoarthritis. Sci Rep. 2023;13(1):2926. doi:10.1038/s41598-023-29802-7
97. Unamuno X, Gómez‐Ambrosi J, Rodríguez A, et al. Adipokine dysregulation and adipose tissue inflammation in human obesity. Eur J Clin Invest. 2018;48(9):e12997. doi:10.1111/eci.12997
98. Barchetta I, Cimini F, Ciccarelli G, et al. Sick fat: the good and the bad of old and new circulating markers of adipose tissue inflammation. J. Endocrinol. Invest. 2019;42(11):1257–1272. doi:10.1007/s40618-019-01052-3
99. Liu L, Shi Z, Ji X, et al. Adipokines, adiposity, and atherosclerosis. Cell Mol Life Sci. 2022;79(5):272. doi:10.1007/s00018-022-04286-2
100. Kirk B, Feehan J, Lombardi G, et al. Muscle, bone, and fat crosstalk: the biological role of myokines, osteokines, and adipokines. Curr. Osteoporos. Rep. 2020;18(4):388–400. doi:10.1007/s11914-020-00599-y
101. Zhang C, Lin Y, Yan CH, et al. Adipokine signaling pathways in osteoarthritis. Front. Bioeng. Biotechnol. 2022;10:865370. doi:10.3389/fbioe.2022.865370
102. Shumnalieva R, Kotov G, Monov SJL. Obesity-related knee osteoarthritis—current concepts. Life. 2023;13(8):1650. doi:10.3390/life13081650
103. Economou A, Mallia I, Fioravanti A, et al. The role of adipokines between genders in the pathogenesis of osteoarthritis. Int J Mol Sci. 2024;25(19):10865. doi:10.3390/ijms251910865
104. Abdelnaby R, Sonbol YT, Dardeer KT, et al. Could osteopontin be a useful biomarker in the diagnosis and severity assessment of osteoarthritis? A systematic review and meta-analysis of recent evidence. Clin Immunol. Clin Immunol. 2023;246:109187.
105. Gandhi R, Takahashi M, Syed K, et al. Relationship between body habitus and joint leptin levels in a knee osteoarthritis population. J Orthop Res. 2010;28(3):329–333. doi:10.1002/jor.21000
106. Conde-Aranda J, Scotece M, Varela-García M, et al. Lipocalin-2 serum levels in rheumatoid arthritis patients treated with adalimumab and its correlation with proinflammatory factors. Mediators Inflamm. 2024;2024:7264704. doi:10.1155/2024/7264704
107. Lambova SN, Batsalova T, Moten D, et al. Serum leptin and resistin levels in knee osteoarthritis-clinical and radiologic links: towards precise definition of metabolic type knee osteoarthritis. Biomedicines. 2021;9(8):1019. doi:10.3390/biomedicines9081019
108. Zhu J, Ruan G, Cen H, et al. Association of serum levels of inflammatory markers and adipokines with joint symptoms and structures in participants with knee osteoarthritis. Rheumatology. 2022;61(3):1044–1052. doi:10.1093/rheumatology/keab479
109. Scotece M, Koskinen-Kolasa A, Pemmari A, et al. Novel adipokine associated with OA: retinol binding protein 4 (RBP4) is produced by cartilage and is correlated with MMPs in osteoarthritis patients. Inflamm Res. 2020;69(4):415–421. doi:10.1007/s00011-020-01326-0
110. Chen R, Zhang Y, Xu H, et al. Val109Asp polymorphism of the omentin-1 gene and incidence of knee osteoarthritis in a Chinese han population: a correlation analysis. Drug Des Devel Ther. 2021;15:5075–5086. doi:10.2147/DDDT.S340410
111. Zhang S, Lei Y, Sun T, Gao Z, Li Z, Shen H. Elevated levels of metrnl in rheumatoid arthritis: association with disease activity. Cytokine. 2022;159:156026. doi:10.1016/j.cyto.2022.156026
112. Jiang L, Xu K, Li J, et al. Nesfatin-1 suppresses interleukin-1β-induced inflammation, apoptosis, and cartilage matrix destruction in chondrocytes and ameliorates osteoarthritis in rats. Aging. 2020;12(2):1760–1777. doi:10.18632/aging.102711
113. Wang Y, Wan R, Hu C. Leptin/obR signaling exacerbates obesity-related neutrophilic airway inflammation through inflammatory M1 macrophages. Mol Med. 2023;29(1):100. doi:10.1186/s10020-023-00702-w
114. Chakhtoura M, Haber R, Ghezzawi M, Rhayem C, Tcheroyan R, Mantzoros CS. Pharmacotherapy of obesity: an update on the available medications and drugs under investigation. EClinicalMedicine. 2023;58:101882. doi:10.1016/j.eclinm.2023.101882
115. Duan ZX, Tu C, Liu Q, et al. Adiponectin receptor agonist AdipoRon attenuates calcification of osteoarthritis chondrocytes by promoting autophagy. J Cell Biochem. 2020;121(5–6):3333–3344. doi:10.1002/jcb.29605
116. Yang S, Ryu JH, Oh H, et al. NAMPT (visfatin), a direct target of hypoxia-inducible factor-2α, is an essential catabolic regulator of osteoarthritis. Ann Rheum Dis. 2015;74(3):595–602. doi:10.1136/annrheumdis-2013-204355
117. Thompson D, Mahmood S, Morrice N, et al. Fenretinide inhibits obesity and fatty liver disease but induces Smpd3 to increase serum ceramides and worsen atherosclerosis in LDLR-/- mice. Sci Rep. 2023;13(1):3937. doi:10.1038/s41598-023-30759-w
118. Joffin N, Niang F, Forest C, et al. Is there NO help for leptin? Biochimie. 2012;94(10):2104–2110. doi:10.1016/j.biochi.2012.06.017
119. Yan M, Zhang J, Yang H, et al. The role of leptin in osteoarthritis. Medicine. 2018;97(14):e0257. doi:10.1097/MD.0000000000010257
120. Cordero-Barreal A, González-Rodríguez M, Ruiz-Fernández C, et al. An update on the role of leptin in the immuno-metabolism of cartilage. Inter J Mol Sci. 2021;22(5):2411. doi:10.3390/ijms22052411
121. Ait Eldjoudi D, Cordero Barreal A, Gonzalez-Rodríguez M, et al. Leptin in osteoarthritis and rheumatoid arthritis: player or bystander? Inter J Mol Sci. 2022;23(5):2859. doi:10.3390/ijms23052859
122. Min S, Shi T, Han X, et al. Serum levels of leptin, osteopontin, and sclerostin in patients with and without knee osteoarthritis. Clin Rheumatol. 2021;40:287–294. doi:10.1007/s10067-020-05150-z
123. Panina SB, Krolevets IV, Milyutina NP, et al. Circulating levels of proinflammatory mediators as potential biomarkers of post-traumatic knee osteoarthritis development. J Orthop Traumatol. 2017;18:349–357. doi:10.1007/s10195-017-0473-8
124. Mai NTT, Hang NT, Hanh DH, et al. Leptin and interleukin-1β levels associated with osteoarthritis in Vietnamese patients: a cross-sectional analysis. Braz J Med Biol Res. 2023;56:e12746. doi:10.1590/1414-431x2023e12746
125. Fu Y, Batushansky A, Kinter M, et al. Effects of leptin and body weight on inflammation and knee osteoarthritis phenotypes in female rats. JBMR Plus. 2023;7(7):e10754. doi:10.1002/jbm4.10754
126. Yang W-H, Liu S-C, Tsai C-H, et al. Leptin induces IL-6 expression through OBRl receptor signaling pathway in human synovial fibroblasts. PLoS One. 2013;8(9):e75551. doi:10.1371/journal.pone.0075551
127. Xiong H, Li W, Li J, et al. Elevated leptin levels in temporomandibular joint osteoarthritis promote proinflammatory cytokine IL‐6 expression in synovial fibroblasts. J. Oral Pathol. 2019;48(3):251–259. doi:10.1111/jop.12819
128. Scotece M, Mobasheri A. Leptin in osteoarthritis: focus on articular cartilage and chondrocytes. Life Sci. 2015;140:75–78. doi:10.1016/j.lfs.2015.05.025
129. Gualillo OJO. Are we approaching a holistic view of leptin’s role in arthritis? Osteoarthritis Cartilage2023;31(1):21–22.
130. Griffin TM, Huebner JL, Kraus VB, et al. Extreme obesity due to impaired leptin signaling in mice does not cause knee osteoarthritis. Arth Rheumat. 2009;60(10):2935–2944. doi:10.1002/art.24854
131. Deis J, Lin T-Y, Bushman T, et al. Lipocalin 2 deficiency alters prostaglandin biosynthesis and mTOR signaling regulation of thermogenesis and lipid metabolism in adipocytes. Cells. 2022;11(9):1535. doi:10.3390/cells11091535
132. Xing X, Wang Y, Pan F, et al. Osteoarthritis and risk of type 2 diabetes: a two‐sample Mendelian randomization analysis. J Diabet. 2023;15(11):987–993. doi:10.1111/1753-0407.13451
133. Wang T, He CJC. Pro-inflammatory cytokines: the link between obesity and osteoarthritis. Cytokine Growth Factor Rev. 2018;44:38–50. doi:10.1016/j.cytogfr.2018.10.002
134. Al Jaberi S, Cohen A, D’Souza C, et al. Lipocalin-2: structure, function, distribution and role in metabolic disorders. Biomed Pharmacother. 2021;142:112002. doi:10.1016/j.biopha.2021.112002
135. Zhao X, Huang P, Li G, et al. Activation of the leptin pathway by high expression of the long form of the leptin receptor (Ob-Rb) accelerates chondrocyte senescence in osteoarthritis. Bone Joint Res. 2019;8(9):425–436. doi:10.1302/2046-3758.89.BJR-2018-0325.R2
136. Conde J, Scotece M, López V, et al. Adiponectin and leptin induce VCAM-1 expression in human and murine chondrocytes. PLoS One. 2012;7(12):e52533. doi:10.1371/journal.pone.0052533
137. Zhang ZM, Shen C, Li H, et al. Leptin induces the apoptosis of chondrocytes in an in vitro model of osteoarthritis via the JAK2‑STAT3 signaling pathway. Mol Med Rep. 2016;13(4):3684–3690. doi:10.3892/mmr.2016.4970
138. Garner M, Alshameeri Z, Khanduja V. Osteoarthritis: genes, nature–nurture interaction and the role of leptin. Int Orthop. 2013;37(12):2499–2505. doi:10.1007/s00264-013-2088-x
139. Wang Y, Xu Z, Wang J, et al. DUSP19, a downstream effector of leptin, inhibits chondrocyte apoptosis via dephosphorylating JNK during osteoarthritis pathogenesis. Mol BioSys. 2016;12(3):721–728. doi:10.1039/c5mb00776c
140. Martel-Pelletier J, Raynauld J-P, Dorais M, et al. The levels of the adipokines adipsin and leptin are associated with knee osteoarthritis progression as assessed by MRI and incidence of total knee replacement in symptomatic osteoarthritis patients: a post hoc analysis. Rheumatology. 2016;55(4):680–688. doi:10.1093/rheumatology/kev408
141. MacDonald IJ, Liu S-C, Huang -C-C, et al. Associations between adipokines in arthritic disease and implications for obesity. Int J Mol Sci. 2019;20(6):1505. doi:10.3390/ijms20061505
142. Gao Y-H, Zhao C-W, Liu B, et al. An update on the association between metabolic syndrome and osteoarthritis and on the potential role of leptin in osteoarthritis. Cytokine. 2020;129:155043. doi:10.1016/j.cyto.2020.155043
143. Feng X, Xiao J, LJFiC B, et al. Role of adiponectin in osteoarthritis. Front Cell Dev Biol. 2022;10:992764. doi:10.3389/fcell.2022.992764
144. Ehling A, Schäffler A, Herfarth H, et al. The potential of adiponectin in driving arthritis. J Immunol. 2006;176(7):4468–4478. doi:10.4049/jimmunol.176.7.4468
145. Schäffler A, Angela E, Elena N, et al. Adipocytokines in synovial fluid. JAMA. 2003;290(13):1709–1710. doi:10.1001/jama.290.13.1709-c
146. Askari A, Arasteh P, Homayounfar R, et al. The role of adipose tissue secretion in the creation and pain level in osteoarthritis. Endocr Regulat. 2020;54(1):6–13. doi:10.2478/enr-2020-0002
147. Yusuf E, Ioan-Facsinay A, Bijsterbosch J, et al. Association between leptin, adiponectin and resistin and long-term progression of hand osteoarthritis. Ann. Rheum. Dis2011;70(7):1282–1284. doi:10.1136/ard.2010.146282
148. Ilia I, Nitusca D, Marian CJD. Adiponectin in osteoarthritis: pathophysiology. Relat Obesity Presu Diagnostic Biomar Pot. 2022;12(2):455.
149. Orellana C, Calvet J, Berenguer-Llergo A, et al. Synovial adiponectin was more associated with clinical severity than synovial leptin in women with knee osteoarthritis. Cartilage. 2021;13(1_suppl):1675S–83S. doi:10.1177/1947603520904776
150. Tang CH, Chiu YC, Tan TW, et al. Adiponectin enhances IL-6 production in human synovial fibroblast via an AdipoR1 receptor, AMPK, p38, and NF-kappa B pathway. J Immunol. 2007;179(8):5483–5492. doi:10.4049/jimmunol.179.8.5483
151. Honsawek S, Chayanupatkul M. Correlation of plasma and synovial fluid adiponectin with knee osteoarthritis severity. Arch Med Res. 2010;41(8):593–598. doi:10.1016/j.arcmed.2010.11.007
152. Liu Z, Gan L, Wu T, et al. Adiponectin reduces ER stress-induced apoptosis through PPARα transcriptional regulation of ATF2 in mouse adipose. Cell Death Dis. 2016;7(11):e2487–e. doi:10.1038/cddis.2016.388
153. Czerewaty M, Łączna M, Kiełbowski K, et al. The effect of plasma cytokines on the expression of adiponectin and its receptors in the synovial membrane of joints and the infrapatellar fat pad in patients with rheumatoid arthritis and osteoarthritis. Prostaglandins Other Lipid Mediat. 2024;172:106824. doi:10.1016/j.prostaglandins.2024.106824
154. Abella V, Scotece M, Conde J, et al. The potential of lipocalin-2/NGAL as biomarker for inflammatory and metabolic diseases. Biomarkers. 2015;20(8):565–571. doi:10.3109/1354750X.2015.1123354
155. Wilson R, Belluoccio D, Little CB, et al. Proteomic characterization of mouse cartilage degradation in vitro. Arthritis Rheumat. 2008;58(10):3120–3131. doi:10.1002/art.23789
156. Gupta K, Shukla M, Cowland JB, et al. Neutrophil gelatinase–associated lipocalin is expressed in osteoarthritis and forms a complex with matrix metalloproteinase 9. Arthritis Rheumatol. 2007;56(10):3326–3335. doi:10.1002/art.22879
157. Choi W-S, Chun J-SJO. Upregulation of lipocalin-2 (LCN2) in osteoarthritic cartilage is not necessary for cartilage destruction in mice. Osteoarthritis Cartilage. 2017;25(3):401–405. doi:10.1016/j.joca.2016.07.009
158. Lee J, Ort T, Ma K, et al. Resistin is elevated following traumatic joint injury and causes matrix degradation and release of inflammatory cytokines from articular cartilage in vitro. Osteoarthritis Cartilage. 2009;17(5):613–620. doi:10.1016/j.joca.2008.08.007
159. Cheleschi S, Gallo I, Barbarino M, et al. MicroRNA mediate visfatin and resistin induction of oxidative stress in human osteoarthritic synovial fibroblasts via NF-κB pathway. Int J Mol Sci. 2019;20(20):5200. doi:10.3390/ijms20205200
160. Chen WC, Lu YC, Kuo SJ, et al. Resistin enhances IL‐1β and TNF‐α expression in human osteoarthritis synovial fibroblasts by inhibiting miR‐149 expression via the MEK and ERK pathways. FASEB J. 2020;34(10):13671–13684. doi:10.1096/fj.202001071R
161. Zhang Z, Zhang Z, Kang Y, et al. Resistin stimulates expression of chemokine genes in chondrocytes via combinatorial regulation of C/EBPβ and NF-κB. Int J Mol Sci. 2014;15(10):17242–17255. doi:10.3390/ijms151017242
162. Silswal N, Singh AK, Aruna B, et al. Human resistin stimulates the pro-inflammatory cytokines TNF-α and IL-12 in macrophages by NF-κB-dependent pathway. Biochem Biophys Res Commun. 2005;334(4):1092–1101. doi:10.1016/j.bbrc.2005.06.202
163. Zhao CW, Song WX, Liu B, et al. Resistin induces chemokine and matrix metalloproteinase production via CAP1 receptor and activation of p38-MAPK and NF-κB signalling pathways in human chondrocytes. Clin Exp Rheumatol. 2022;40(3):501–513. doi:10.55563/clinexprheumatol/avcj31
164. Chen W-C, Lin C-Y, Kuo S-J, et al. Resistin enhances VCAM-1 expression and monocyte adhesion in human osteoarthritis synovial fibroblasts by inhibiting MiR-381 expression through the PKC, p38, and JNK signaling pathways. Cells. 2020;9(6):1369. doi:10.3390/cells9061369
165. Han W, Aitken D, Zheng S, et al. Higher serum levels of resistin are associated with knee synovitis and structural abnormalities in patients with symptomatic knee osteoarthritis. J Am Med Dir Assoc. 2019;20(10):1242–1246. doi:10.1016/j.jamda.2019.07.001
166. Wang K, Xu J, Cai J, et al. Serum levels of resistin and interleukin-17 are associated with increased cartilage defects and bone marrow lesions in patients with knee osteoarthritis. Mod Rheumatol. 2017;27(2):339–344. doi:10.1080/14397595.2016.1205777
167. Philp AM, Collier RL, Grover LM, et al. Resistin promotes the abnormal Type I collagen phenotype of subchondral bone in obese patients with end stage hip osteoarthritis. Sci Rep. 2017;7(1):4042. doi:10.1038/s41598-017-04119-4
168. Zhao C-W, Gao Y-H, Song W-X, et al. An update on the emerging role of resistin on the pathogenesis of osteoarthritis. 2019.
169. Cheleschi S, Giordano N, Volpi N, et al. A complex relationship between visfatin and resistin and microRNA: an in vitro study on human chondrocyte cultures. Int J Mol Sci. 2018;19(12):3909. doi:10.3390/ijms19123909
170. Calvet J, Orellana C, Gratacós J, et al. Synovial fluid adipokines are associated with clinical severity in knee osteoarthritis: a cross-sectional study in female patients with joint effusion. Arthritis Res. Ther.2016;18:1–11.
171. Perruccio AV, Mahomed NN, Chandran V, et al. Plasma adipokine levels and their association with overall burden of painful joints among individuals with hip and knee osteoarthritis. J Rheumatol. 2014;41(2):334–337. doi:10.3899/jrheum.130709
172. Dehghani S, Alipoor E, Salimzadeh A, et al. The effect of a garlic supplement on the pro-inflammatory adipocytokines, resistin and tumor necrosis factor-alpha, and on pain severity, in overweight or obese women with knee osteoarthritis. Phytomedicine. 2018;48:70–75. doi:10.1016/j.phymed.2018.04.060
173. Song Y, Guan J, Wang H, et al. Possible involvement of serum and synovial fluid resistin in knee osteoarthritis: cartilage damage, clinical, and radiological links. J Clin Laboratory Analysis. 2016;30(5):437–443. doi:10.1002/jcla.21876
174. Alissa EM, Alzughaibi LS, Marzouki ZMJ. Relationship between serum resistin, body fat and inflammatory markers in females with clinical knee osteoarthritis. Knee. 2020;27(1):45–50. doi:10.1016/j.knee.2019.12.009
175. Karvonen-Gutierrez CA, Zheng H, Mancuso P, et al. Higher leptin and adiponectin concentrations predict poorer performance-based physical functioning in midlife women. J Gerontol A Biol Sci Med Sci. 2016;71(4):508–514.
176. Zhao X, Yang Z, Zhang Z, et al. CCL3 serves as a potential plasma biomarker in knee degeneration (osteoarthritis). Osteoarthritis Cartilage. 2015;23(8):1405–1411. doi:10.1016/j.joca.2015.04.002
177. Martín-Márquez BT, Sandoval-García F, Corona-Meraz FI, et al. Osteopontin: a bone-derived protein involved in rheumatoid arthritis and osteoarthritis immunopathology. Biomolecules. 2023;13(3):502. doi:10.3390/biom13030502
178. Hasegawa M, Segawa T, Maeda M, et al. Thrombin-cleaved osteopontin levels in synovial fluid correlate with disease severity of knee osteoarthritis. J Rheumatol. 2011;38(1):129–134. doi:10.3899/jrheum.100637
179. Tian J, Gao S-G, Li Y-S, et al. The β-catenin/TCF-4 pathway regulates the expression of OPN in human osteoarthritic chondrocytes. J. Orthop. Surg. Res.2020;15:1–11.
180. Tu M, Li Y, Zeng C, et al. MicroRNA-127-5p regulates osteopontin expression and osteopontin-mediated proliferation of human chondrocytes. Sci Rep. 2016;6(1):25032. doi:10.1038/srep25032
181. Gao S, Li K, Zeng K, et al. Elevated osteopontin level of synovial fluid and articular cartilage is associated with disease severity in knee osteoarthritis patients. Osteoarthritis Cartilage. 2010;18(1):82–87. doi:10.1016/j.joca.2009.07.009
182. Li Y, Jiang W, Wang H, et al. Osteopontin promotes expression of matrix metalloproteinase 13 through NF-κB signaling in osteoarthritis. Biomed Res. Int. 2016;2016.
183. Molnar V, Matišić V, Kodvanj I, et al. Cytokines and chemokines involved in osteoarthritis pathogenesis. Int J Mol Sci. 2021;22(17):9208. doi:10.3390/ijms22179208
184. Dai G, Xiao H, Liao J, et al. Osteocyte TGFβ1‑Smad2/3 is positively associated with bone turnover parameters in subchondral bone of advanced osteoarthritis. Int J Mol Med. 2020;46(1):167–178. doi:10.3892/ijmm.2020.4576
185. Bai R-J, Li Y-S, Zhang F. Osteopontin, a bridge links osteoarthritis and osteoporosis. Front Endocrinol. 2022;13:1012508. doi:10.3389/fendo.2022.1012508
186. Matsui Y, Iwasaki N, Kon S, et al. Accelerated development of aging‐associated and instability‐induced osteoarthritis in osteopontin‐deficient mice. Arthritis Rheumatism. 2009;60(8):2362–2371. doi:10.1002/art.24705
187. Gao SG, Yu Y, Zeng C, et al. Phosphorylation of osteopontin has proapoptotic and proinflammatory effects on human knee osteoarthritis chondrocytes. Exp Ther Med. 2016;12(5):3488–3494. doi:10.3892/etm.2016.3784
188. Li S, Liu J, Liu S, et al. Chitosan oligosaccharides packaged into rat adipose mesenchymal stem cells-derived extracellular vesicles facilitating cartilage injury repair and alleviating osteoarthritis. Chin J Nat Med. 2021;19:1–19. doi:10.1016/S1875-5364(21)60001-8
189. Sun P-F, Kong W-K, Liu L, et al. Osteopontin accelerates chondrocyte proliferation in osteoarthritis rats through the NF-κb signaling pathway. Eur Rev Med Pharmacol Sci. 2020;24(6):2836–2842. doi:10.26355/eurrev_202003_20647
190. Xu J, Yi Y, Li L, et al. Osteopontin induces vascular endothelial growth factor expression in articular cartilage through PI3K/AKT and ERK1/2 signaling. Mol Med Rep. 2015;12(3):4708–4712. doi:10.3892/mmr.2015.3975
191. Chen W-P, Bao J-P, Feng J, et al. Increased serum concentrations of visfatin and its production by different joint tissues in patients with osteoarthritis. Clin Chem Lab Med. 2010;48(8):1141–1145. doi:10.1515/CCLM.2010.230
192. Oh H, Kwak J-S, Yang S, et al. Reciprocal regulation by hypoxia-inducible factor-2α and the NAMPT-NAD + -SIRT axis in articular chondrocytes is involved in osteoarthritis. Osteoarthritis Cartilage. 2015;23(12):2288–2296. doi:10.1016/j.joca.2015.07.009
193. Laiguillon M-C, Houard X, Bougault C, et al. Expression and function of visfatin (Nampt), an adipokine-enzyme involved in inflammatory pathways of osteoarthritis. Arthritis Res. Ther. 2014;16:1–12.
194. Han D-F, Li Y, Xu H-Y, et al. An update on the emerging role of visfatin in the pathogenesis of osteoarthritis and pharmacological intervention. Evid Based Complement Alternat Med. 2020;2020. doi:10.1155/2020/8303570
195. Cook KS, Min HY, Johnson D, et al. Adipsin: a circulating serine protease homolog secreted by adipose tissue and sciatic nerve. Science. 1987;237(4813):402–405. doi:10.1126/science.3299705
196. Choy LN, Rosen B, Spiegelman BMJJo BC. Adipsin and an endogenous pathway of complement from adipose cells. J. Biol. Chem. 1992;267(18):12736–12741.
197. Tontonoz P, Hu E, Spiegelman BMJC. Stimulation of adipogenesis in fibroblasts by PPARγ2, a lipid-activated transcription factor. Cell. 1994;79(7):1147–1156. doi:10.1016/0092-8674(94)90006-x
198. Paré F, Tardif G, Fahmi H, et al. In vivo protective effect of adipsin-deficiency on spontaneous knee osteoarthritis in aging mice. Aging. 2020;12(3):2880. doi:10.18632/aging.102784
199. Valverde-Franco G, Tardif G, Mineau F, et al. High in vivo levels of adipsin lead to increased knee tissue degradation in osteoarthritis: data from humans and animal models. Rheumatology2018;57(10):1851–1860. doi:10.1093/rheumatology/key181
200. Martel-Pelletier J, Tardif G, Trépanier JR, et al. The ratio adipsin/MCP-1 is strongly associated with structural changes and CRP/MCP-1 with symptoms in obese knee osteoarthritis subjects: data from the osteoarthritis initiative. Osteoarthritis Cartilage. 2019;27(8):1163–1173. doi:10.1016/j.joca.2019.04.016
201. Prentice KJ, Saksi J, Robertson LT, et al. A hormone complex of FABP4 and nucleoside kinases regulates islet function. Nature. 2021;600(7890):720–6.
202. Zhang C, Li T, Chiu KY, et al. FABP4 as a biomarker for knee osteoarthritis. Biomarker med2018;12(2):107–118. doi:10.2217/bmm-2017-0207
203. Schadler P, Lohberger B, Thauerer B, et al. Fatty acid–binding protein 4 (FABP4) is associated with cartilage thickness in end-stage knee osteoarthritis. Cartilage. 2021;13(2_suppl):1165S–73S. doi:10.1177/19476035211011520
204. Yang K, Xie Q, Liang J, et al. Identification of andrographolide as a novel FABP4 inhibitor for osteoarthritis treatment. Phytomedicine. 2023;118:154939. doi:10.1016/j.phymed.2023.154939
205. Zhang C, Chiu K, Chan B, et al. Knocking out or pharmaceutical inhibition of fatty acid binding protein 4 (FABP4) alleviates osteoarthritis induced by high-fat diet in mice. Osteoarthritis Cartilage. 2018;26(6):824–833. doi:10.1016/j.joca.2018.03.002
206. Mao H, Han B, Li H, et al. FABP4 knockdown suppresses inflammation, apoptosis and extracellular matrix degradation in IL-1β-induced chondrocytes by activating PPARγ to regulate the NF-κB signaling pathway. Mol. Med. Rep2021;24(6):1–10.
207. Zhang C, Lin Y, Li H, et al. Fatty acid binding protein 4 (FABP4) induces chondrocyte degeneration via activation of the NF‐κb signaling pathway. FASEB J. 2024;38(1):e23347. doi:10.1096/fj.202301882R
208. Scotece M, Conde J, Abella V, et al. NUCB2/nesfatin‐1: a new adipokine expressed in human and murine chondrocytes with pro‐inflammatory properties, an in vitro study. J Orthopaedic Res. 2014;32(5):653–660. doi:10.1002/jor.22585
209. Jiang L, Bao J, Zhou X, et al. Increased serum levels and chondrocyte expression of nesfatin-1 in patients with osteoarthritis and its relation with BMI, hsCRP, and IL-18. Mediators of Inflamm. 2013;2013:1–9. doi:10.1155/2013/631251
210. Zhang Y, Shui X, Lian X, et al. Serum and synovial fluid nesfatin-1 concentration is associated with radiographic severity of knee osteoarthritis. Med Sci Monit. 2015;21:1078. doi:10.12659/MSM.892875
211. Lee K-T, Chen B-C, Liu S-C, et al. Nesfatin-1 facilitates IL-1β production in osteoarthritis synovial fibroblasts by suppressing miR-204-5p synthesis through the AP-1 and NF-κB pathways. Aging. 2021;13(18):22490. doi:10.18632/aging.203559
212. Jiang L, Moqbel SAA, Zhu J, et al. Nesfatin-1 suppresses autophagy of chondrocytes in osteoarthritis via remodeling of cytoskeleton and inhibiting RhoA/ROCK signal pathway. J. Orthop. Surg. Res. 2023;18(1):153. doi:10.1186/s13018-023-03539-5
213. DJJBST W. Serpins in cartilage and osteoarthritis: what do we know? Bio Soc Trans. 2021;49(2):1013–1026. doi:10.1042/BST20201231
214. Shen S, Wu Y, Chen J, et al. CircSERPINE2 protects against osteoarthritis by targeting miR-1271 and ETS-related gene. Ann. Rheum. Dis. 2019;78(6):826–836. doi:10.1136/annrheumdis-2018-214786
215. Santoro A, Conde J, Scotece M, et al. SERPINE2 inhibits IL-1α-induced MMP-13 expression in human chondrocytes: involvement of ERK/NF-κB/AP-1 pathways. PLoS One. 2015;10(8):e0135979. doi:10.1371/journal.pone.0135979
216. Wu S, Yang Y, Zhang M, et al. Serpin peptidase inhibitor, clade E, member 2 in physiology and pathology: recent advancements. Front Mol Biosci. 2024;11:1334931. doi:10.3389/fmolb.2024.1334931
217. Wei J, Hettinghouse A, Liu C. The role of progranulin in arthritis. Ann. N. Y. Acad. Sci. 2016;1383(1):5–20. doi:10.1111/nyas.13191
218. He Z, Ong CH, Halper J, et al. Progranulin is a mediator of the wound response. Nat Med. 2003;9(2):225–229. doi:10.1038/nm816
219. J-l W, Fu W, Y-j D, et al. Progranulin derivative atsttrin protects against early osteoarthritis in mouse and rat models. Arthritis Res. Ther. 2017;19:1–14.
220. Caramés B, Olmer M, Kiosses WB, et al. The relationship of autophagy defects to cartilage damage during joint aging in a mouse model. Arthritis amp. 2015;67(6):1568–1576. doi:10.1002/art.39073
221. Loeser RF, Collins JA, Diekman BO. Ageing and the pathogenesis of osteoarthritis. Nat Rev. Rheumatol. 2016;12(7):412–420. doi:10.1038/nrrheum.2016.65
222. Pan Y, Yang Y, Fan M, et al. Progranulin regulation of autophagy contributes to its chondroprotective effect in osteoarthritis. Genes amp. 2023;10(4):1582–1595. doi:10.1016/j.gendis.2022.05.031
223. Liang L, Zhang F, Feng N, et al. IRE1α protects against osteoarthritis by regulating progranulin-dependent XBP1 splicing and collagen homeostasis. Experimental amp. 2023;55(11):2376–2389. doi:10.1038/s12276-023-01106-w
224. Zhao Y-P, Tian Q-Y, Frenkel S, et al. The promotion of bone healing by progranulin, a downstream molecule of BMP-2, through interacting with TNF/TNFR signaling. Biomaterials. 2013;34(27):6412–6421. doi:10.1016/j.biomaterials.2013.05.030
225. Tang W, Lu Y, Tian Q-Y, et al. The growth factor progranulin binds to TNF receptors and is therapeutic against inflammatory arthritis in mice. Science. 2011;332(6028):478–484. doi:10.1126/science.1199214
226. Zhao Y-P, Liu B, Tian Q-Y, et al. Progranulin protects against osteoarthritis through interacting with TNF-α and β-Catenin signalling. Ann. Rheum. Dis. 2015;74(12):2244–2253. doi:10.1136/annrheumdis-2014-205779
227. Ma J, Niu D-S, Wan N-J, et al. Elevated chemerin levels in synovial fluid and synovial membrane from patients with knee osteoarthritis. Int JClinl Exp Patholo. 2015;8(10):13393.
228. Valcamonica E, Chighizola C, Comi D, et al. Levels of chemerin and interleukin 8 in the synovial fluid of patients with inflammatory arthritides and osteoarthritis. Clin and Exp Rheumatol. 2014;32(2):243–250.
229. Santana LJC, Herrera FR, Rojas AP, et al. Serum chemerin in a cohort of Colombian patients with primary osteoarthritis. Reumatolog. 2021;17(9):530–535.
230. Eisinger K, Bauer S, Schäffler A, et al. Chemerin induces CCL2 and TLR4 in synovial fibroblasts of patients with rheumatoid arthritis and osteoarthritis. Exp Mol Patholo. 2012;92(1):90–96. doi:10.1016/j.yexmp.2011.10.006
231. Ma J, Ren L, Guo C-J, et al. Chemerin affects the metabolic and proliferative capabilities of chondrocytes by increasing the phosphorylation of AKT/ERK. Eur Rev Med Pharmacol Sci. 2018;22(12).
232. Dahlman I, Elsen M, Tennagels N, et al. Functional annotation of the human fat cell secretome. Arch. Physiol. Biochem. 2012;118(3):84–91. doi:10.3109/13813455.2012.685745
233. Conde J, Scotece M, Abella V, et al. Identification of novel adipokines in the joint. differential expression in healthy and osteoarthritis tissues. PLoS One. 2015;10(4):e0123601. doi:10.1371/journal.pone.0123601
234. Kumar S, Hand AT, Connor JR, et al. Identification and cloning of a connective tissue growth factor-like cDNA from human osteoblasts encoding a novel regulator of osteoblast functions. J Bio Chem. 1999;274(24):17123–17131. doi:10.1074/jbc.274.24.17123
235. Ruiz-Fernández C, González-Rodríguez M, Abella V, et al. WISP-2 modulates the induction of inflammatory mediators and cartilage catabolism in chondrocytes. Lab Invest. 2022;102(9):989–999. doi:10.1038/s41374-022-00793-9
236. Blüher MJE. Vaspin in obesity and diabetes: pathophysiological and clinical significance. Endocrine 2012;41(2):176–182.
237. Bao J-P, Jiang L-F, Chen W-P, et al. Expression of vaspin in the joint and the levels in the serum and synovial fluid of patients with osteoarthritis. Int J Clin Exp Med. 2014;7(10):3447.
238. Pilarski Ł, Pelczyńska M, Koperska A, et al. Association of serum vaspin concentration with metabolic disorders in obese individuals. Biomolecules. 2023;13(3):508. doi:10.3390/biom13030508
239. Wang J, Zhang K, Zhang S, et al. Vaspin promotes chondrogenic differentiation of BMSCs via Akt activation in osteoarthritis. BMC Musculoskelet. Disord. 2022;23(1):344. doi:10.1186/s12891-022-05295-9
240. Wang J, Chen Z, Guan ZJIJo RD. Vaspin deficiency failed to promote the proliferation of BMSCs in osteoarthritis. Int. J. Rheum. Dis. 2021;24(1):90–95. doi:10.1111/1756-185X.14010
241. He H, Lu M, Shi H, et al. Vaspin regulated cartilage cholesterol metabolism through miR155/LXRα and participated in the occurrence of osteoarthritis in rats. Life Sci. 2021;269:119096. doi:10.1016/j.lfs.2021.119096
242. Sukenik S, Henkin J, Zimlichman S, et al. Serum and synovial fluid levels of serum amyloid A protein and C-reactive protein in inflammatory and noninflammatory arthritis. J Rheumatol. 1988;15(6):942–945.
243. Kellgren JH, Lawrence JJ. Radiological assessment of osteo-arthrosis. Ann. Rheum. Dis. 1957;16(4):494–502. doi:10.1136/ard.16.4.494
244. Koga T, Torigoshi T, Motokawa S, et al. Serum amyloid A-induced IL-6 production by rheumatoid synoviocytes. FEBS Letters. 2008;582(5):579–585. doi:10.1016/j.febslet.2008.01.022
245. Connolly M, Marrelli A, Blades M, et al. Acute serum amyloid A induces migration, angiogenesis, and inflammation in synovial cells in vitro and in a human rheumatoid arthritis/SCID mouse chimera model. J Immunol. 2010;184(11):6427–6437. doi:10.4049/jimmunol.0902941
246. de Seny D, Cobraiville G, Charlier E, et al. Acute-phase serum amyloid a in osteoarthritis: regulatory mechanism and proinflammatory properties. PLoS One. 2013;8(6):e66769. doi:10.1371/journal.pone.0066769
247. Li Z-G, Zhao D-W, Xia C-J, et al. Decreased synovial fluid omentin-1 concentrations reflect symptomatic severity in patients with knee osteoarthritis. Scand J Clin Lab Invest. 2012;72(8):623–628. doi:10.3109/00365513.2012.726370
248. Xu L, Zhu G-B, Wang L, et al. Synovial fluid omentin-1 levels are inversely correlated with radiographic severity of knee osteoarthritis. J. Investig. Med. 2012;60(3):583–586. doi:10.2310/JIM.0b013e31824443cb
249. Ko C-Y, Lin -Y-Y, Achudhan D, et al. Omentin-1 ameliorates the progress of osteoarthritis by promoting IL-4-dependent anti-inflammatory responses and M2 macrophage polarization. Int. J. Biol. Sci. 2023;19(16):5275. doi:10.7150/ijbs.86701
250. Chai B, Zheng Z-H, Liao X, et al. The protective role of omentin-1 in IL-1β-induced chondrocyte senescence. Artif Cells Nanomed Biotechnol. 2020;48(1):8–14. doi:10.1080/21691401.2019.1699803
251. Bridgewood C, Russell T, Weedon H, et al. The novel cytokine Metrnl/IL-41 is elevated in psoriatic arthritis synovium and inducible from both entheseal and synovial fibroblasts. Clin Immunol. 2019;208:108253. doi:10.1016/j.clim.2019.108253
252. Sobieh BH, El-Mesallamy HO, Kassem DH. Beyond mechanical loading: the metabolic contribution of obesity in osteoarthritis unveils novel therapeutic targets. Heliyon2023;9:e15700. doi:10.1016/j.heliyon.2023.e15700
253. Sobieh BH, Kassem DH, Zakaria ZM, et al. Potential emerging roles of the novel adipokines adipolin/CTRP12 and meteorin-like/METRNL in obesity-osteoarthritis interplay. Cytokine. 2021;138:155368. doi:10.1016/j.cyto.2020.155368
254. Zheng S-L, Z-y L, Song J, et al. Metrnl: a secreted protein with new emerging functions. Acta Pharmacol. Sin. 2016;37(5):571–579. doi:10.1038/aps.2016.9
255. Liu J, Jia S, Yang Y, et al. Exercise induced meteorin-like protects chondrocytes against inflammation and pyroptosis in osteoarthritis by inhibiting PI3K/Akt/NF-κB and NLRP3/caspase-1/GSDMD signaling. Bio amp. 2023;158:114118. doi:10.1016/j.biopha.2022.114118
256. Warner SC, Nair A, Marpadga R, et al. IL-15 and IL15RA in osteoarthritis: association with symptoms and protease production, but not structural severity. Front. Immunol. 2020;11:1385. doi:10.3389/fimmu.2020.01385
257. Villalvilla A, García-Martín A, Largo R, et al. The adipokine lipocalin-2 in the context of the osteoarthritic osteochondral junction. Sci Rep. 2016;6(1):29243. doi:10.1038/srep29243
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms.php
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms.php
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.