")
Back to Journals » International Journal of Nanomedicine » Volume 20
Mesenchymal Stem Cell-Derived Exosomes Hold Promise in the Treatment of Diabetic Foot Ulcers
Authors Wang H, Wu S, Bai X, Pan D, Ning Y, Wang C, Guo L, Guo J, Gu Y
Received 9 January 2025
Accepted for publication 11 April 2025
Published 6 May 2025 Volume 2025:20 Pages 5837—5857
DOI https://doi.org/10.2147/IJN.S516533
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 5
Editor who approved publication: Dr Kamakhya Misra
Hui Wang,1,* Sensen Wu,1,* Xinyu Bai,2 Dikang Pan,1 Yachan Ning,1 Cong Wang,1 Lianrui Guo,1 Jianming Guo,1 Yongquan Gu1
1Department of Vascular Surgery, Xuanwu Hospital, Capital Medical University, Beijing, 100053, People’s Republic of China; 2Department of Ophthalmology, The Second Hospital of Jilin University, Changchun, Jilin Province, 130033, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Yongquan Gu; Jianming Guo, Department of Vascular Surgery, Xuanwu Hospital, Capital Medical University, No. 45, Changchun Street, Beijing, 100053, People’s Republic of China, Tel +86 15311614369, Email [email protected]; [email protected]
Abstract: Diabetic foot ulcers (DFU) represent one of the most common side effects of diabetes, significantly impacting patients’ quality of life and imposing considerable financial burdens on families and society at large. Despite advancements in therapies targeting lower limb revascularization and various medications and dressings, outcomes for patients with severe lesions remain limited. A recent breakthrough in DFU treatment stems from the development of mesenchymal stem cells (MSCs). MSCs have shown promising results in treating various diseases and skin wounds due to their ability for multidirectional differentiation and immunomodulation. Recent studies highlight that MSCs primarily repair tissue through their paracrine activities, with exosomes playing a crucial role as the main biologically active components. These exosomes transport proteins, mRNA, DNA, and other substances, facilitating DFU treatment through immunomodulation, antioxidant effects, angiogenesis promotion, endothelial cell migration and proliferation, and collagen remodeling. Mesenchymal stem cell-derived exosomes (MSC-Exo) not only deliver comparable therapeutic effects to MSCs but also mitigate adverse reactions like immune rejection associated with MSCs transplantation. This article provides an overview of DFU pathophysiology and explores the mechanisms and research progress of MSC-Exo in DFU therapy.
Keywords: diabetic foot ulcer, DFU, mesenchymal stem cells, MSCs, exosomes, skin repair, regeneration, angiogenesis
Graphical Abstract:

Introduction
Diabetes stands out as one of the most significant and widespread chronic diseases globally. The number of people with diabetes increased to 536.6 million in 2021 alone, and estimates show that figure will rise to an astounding 783.2 million by 2045.1,2 The metabolic imbalances linked to diabetes give rise to a myriad of complications, spanning from cardiovascular and cerebrovascular disorders to neuropathy, retinopathy, and nephropathy.3,4 One of the notable complications is diabetic foot ulcer (DFU), with estimates suggesting that approximately 15% of patients with diabetes experience DFU at some point in their lives.5,6 DFU can significantly contribute to pain, deteriorate quality of life, impede mobility, and, in severe cases, culminate in amputation. Global health-related costs associated with diabetes were expected to be 966 billion USD in 2021; estimates suggest that by 2045, these costs will have increased to 1.054 trillion USD.7 Hence, diabetes mellitus and its associated complications, such as foot ulceration, impose substantial human, societal, and economic burdens.
DFU typically manifests alongside symptoms of trophic disorders, such as ulceration or gangrene of the foot tissues, as well as neuropathy and/or vascularization issues affecting the lower extremities. The underlying pathology is characterized by chronic inflammation.8,9 Diabetic foot infection (DFI) can be caused by a wide range of microorganisms, either alone or in combination. However, gram-positive cocci (GPC), especially staphylococci, are the most common. Other common pathogens encompass Enterobacteriales, Enterococci, and Streptococci.10,11 Surgical debridement, pressure reduction by weight-bearing management, treatment of lower extremity ischemia and foot infection, and timely referral for multidisciplinary care are commonly used as first-line therapy for DFU.12
Endovascular therapy has gained considerable traction in recent years as a method to enhance the quality of life for DFU patients and preserve limbs by reinstating blood flow to the arteries of the lower extremities. The primary objective is to alleviate rest pain and salvage limbs, while the secondary goal is to uphold the long-term patency of targeted blood vessels.13,14 Nevertheless, due to the intricate nature of vasculopathy in diabetic foot patients, attaining long-term vascular patency is often deemed exceedingly challenging.15 Therefore, a unique strategy to speed up diabetic wound healing is desperately needed. For the treatment of ischemia in DFU, mesenchymal stem cells (MSCs) therapy has become a viable substitute for conventional surgical and/or endovascular revascularization throughout the last 20 years.16–18
MSCs are a kind of pluripotent stem cells that have the amazing capacity to differentiate into multiple lineages, such as adipocytes, chondrocytes, osteoblasts, and vascular endothelial cells, as well as to self-renew, promoting angiogenesis and tissue repair17 Moreover, MSCs can secrete various bioactive substances, such as growth factors, cytokines, and exosomes, exerting immunomodulatory and anti-inflammatory effects. This endows them with broad application prospects in the complex pathological environments of diabetes and cardiovascular diseases.19,20 Multiple clinical studies have shown that transplantation of MSCs is relatively safe, with no significant immune rejection reactions or malignant proliferation observed.21–23 Due to their inherent immunosuppressive properties, MSCs can evade attacks from the host immune system, reducing adverse reactions. Furthermore, exosomes derived from MSCs, as a cell-free therapeutic approach, further reduce potential safety risks. In addition, in the treatment of diabetes, MSCs have demonstrated positive effects by regulating glucose and lipid metabolism, improving islet function, and promoting the healing of diabetic foot ulcers.24,25
Recent studies indicate that the effects of stem cells are largely mediated by exosomes secreted by MSCs. The use of exosomes circumvents several challenges associated with MSCs transplantation, such as cell survival rates, homing abilities, and immune rejection.26–28 Exosomes can deliver the bioactive molecules of MSCs without the complex manipulations required for live cells, reducing the technical difficulty and risks of treatment. Additionally, the nanoparticle characteristics of exosomes grant them superior biodistribution and targeting capabilities. They can traverse biological barriers within the body to precisely reach damaged tissues and achieve targeted release of therapeutic factors, thereby enhancing therapeutic efficacy.29 Moreover, exosomes possess strong anti-inflammatory and pro-healing properties; they can modulate the local immune microenvironment, alleviate chronic inflammatory responses, and promote angiogenesis and cellular regeneration. This regulatory effect is more efficient than treatment with MSCs alone.30,31 So this review aims to clarify the pathophysiology of DFU and explore the mechanisms and recent advancements in mesenchymal stem cell-derived exosome (MSC-Exo) therapy for DFU.
The Pathophysiology of DFU
The pathophysiology of the diabetic foot encompasses polyneuropathy and angiopathy, with a distinction between microangiopathy and macroangiopathy. Approximately 60% of diabetic feet are primarily neuropathic, around 10% exhibit solely peripheral arterial changes, and roughly 30% present a combination of both neuropathic and arterial components.32–34
Vascular Changes in DFU
Peripheral artery disease (PAD) contributes to approximately 50% of foot ulcers in diabetic patients, with atherosclerosis emerging as the leading cause of peripheral vascular disease.35 Irreversible non-enzymatic glycation, oxidative stress, heightened inflammatory responses, endothelial dysfunction, and a hypercoagulable state are all factors that promote atherosclerosis and exert a detrimental effect on wound healing.36,37
Small and microvessels are the primary target of vascular lesions in diabetes patients, with lower limb lesions predominantly affecting vessels below the knee, such as the tibial and peroneal arteries. Structural alterations in lower limb vessels impede the delivery of nutrients and active leukocytes to lesion sites, heightening susceptibility to infection and its severity. This dynamic accelerates the onset and progression of DFU.38 Moreover, diabetic patients display a hypercoagulable state, predisposing them to micro thrombosis, particularly in smaller diameter end vessels. This phenomenon exacerbates local ischemia and hypoxia within diabetic wound tissues, compounding the challenges of wound healing.39
Neuropathy of DFU
Research suggests that high blood glucose levels may interfere with the way important molecules involved in nervous system plasticity function. In the dorsal root ganglion (DRG), this disturbance alters the expression patterns of heat shock proteins, neuroregulatory proteins, β-tubulin proteins, and poly-ADP-ribose polymerase, leading to an elevated stress response and mitochondrial malfunction. Consequently, axonal deformation occurs, causing permanent damage to the nervous system.40–42 Diabetic foot ulcer neuropathy encompasses diabetic sensory, motor, and autonomic neuropathy.38,43,44 Sensory neuropathy reduces the perception of pain, leading to a diminished ability to detect injuries or wounds. Consequently, ulcerations may go unnoticed by both patients and healthcare providers, increasing the risk of complications.45,46 Motor neuropathy contributes to foot deformities and biomechanical abnormalities47, while autonomic neuropathy results in skin changes such as dryness and increased susceptibility to fissures. These alterations compromise the skin’s barrier function, making it more prone to microbial invasion and subsequent dermal infections, including cellulitis.38
Molecular Biological Mechanisms of DFU
Oxidative Stress Pathway (ROS-Nrf2 Pathway)
Patients with diabetes often experience prolonged hyperglycemia, which induces oxidative stress by increasing the production of reactive oxygen species (ROS).48 The accumulation of ROS can damage cell membranes, proteins, and DNA, leading to cellular dysfunction and necrosis. Under normal circumstances, Nuclear Factor E2-related Factor 2 (Nrf2) is the main regulator of antioxidant stress.49 When oxidative stress increases, Nrf2 is activated to initiate the expression of antioxidant genes such as superoxide dismutase (SOD) and glutathione peroxidase (GPx).50,51 However, in diabetic patients, the Nrf2 pathway is often impaired, resulting in decreased cellular antioxidant capacity, further exacerbating tissue damage and ulcer formation.
Nuclear Factor κB (NF-κB) Pathway
NF-κB is a key transcription factor regulating inflammatory responses. In a hyperglycemic environment, the NF-κB signaling pathway is activated, leading to the release of a series of pro-inflammatory factors such as tumor necrosis factor-alpha (TNF-α), interleukin-6 (IL-6), and interleukin-1β (IL-1β).52,53 These inflammatory factors initiate and sustain chronic inflammatory responses in local tissues, delaying wound healing. Activation of NF-κB also affects the polarization state of macrophages, keeping them in a pro-inflammatory M1 state, which further deteriorates the local inflammatory environment.54
TGF-β/Smad Signaling Pathway
Transforming Growth Factor-beta (TGF-β) is an important regulatory factor in the wound healing process, responsible for regulating cell proliferation, differentiation, and matrix remodeling.55,56 TGF-β transmits signals through the Smad protein family, regulating the functions of fibroblasts and vascular endothelial cells. In diabetic foot ulcers, the TGF-β/Smad pathway is often inhibited, leading to insufficient matrix remodeling, affecting the synthesis of collagen and extracellular matrix (ECM), and delaying tissue repair and wound closure.57–59
Phosphatidylinositol 3-Kinase/Protein Kinase B (PI3K/Akt) Pathway
The PI3K/Akt signaling pathway plays a central role in regulating cell survival, proliferation, and metabolism. Especially in the repair process of diabetic foot ulcers, this pathway promotes angiogenesis and cell proliferation, regulating cellular stress responses.60 However, hyperglycemia inhibits the PI3K/Akt signaling pathway, reducing the proliferation and migration abilities of endothelial cells and fibroblasts, decreasing angiogenesis around the wound, and delaying the healing process61 (Figure 1).
 |
Figure 1 Pathogenesis of Diabetic Ulcers. (1) In a high-glucose environment, the increased production of reactive oxygen species (ROS) induces oxidative stress, leading to mitochondrial damage and activation of the NF-κB signaling pathway. This promotes the differentiation of neutrophils and M1 macrophages, while reducing Nuclear factor erythroid 2-related factor 2(Nrf2) activation, resulting in decreased levels of superoxide dismutase (SOD), glutathione peroxidase (GPx), and other antioxidants, ultimately reducing cellular antioxidant capacity.(2) The TGF-β/Smad pathway is often inhibited, delaying tissue repair and wound closure.(3) Moreover, the activation of ROS and inhibition of the PI3K/Akt signaling pathway reduce the proliferation and migration ability of endothelial cells and fibroblasts, thereby decreasing angiogenesis around the wound.Yellow arrows represent the antagonistic effects of high glucose, and red arrows represent the inhibitory effects of high glucose. Abbreviations|: AGE, advanced glycation end product, TNF, tumor necrosis factor. |
The Healing Process of Diabetic Foot Ulcers (DFU)
Hemostasis, inflammation, proliferation, and maturation are the four sequential but related stages of the wound-healing process (Figure 2). Hemostasis, the initial and critical stage of healing, is dedicated to promptly halting bleeding upon skin injury. This phase involves platelet activation, which takes place at the interface between platelets and the matrix, facilitating their aggregation.45,46 Ultimately, under the influence of thrombin, platelet clots transition into stable clots, significantly reducing the creation of a fibrin network. Inflammation is the next stage of wound healing, and its main goal is to get rid of debris and bacteria and establish the optimal wound bed for the formation of fresh tissue.38,47 The activation of neutrophils and macrophages, which are essential for limiting bleeding and avoiding infection either directly or by the release of different soluble mediators, is critical to these processes.48 During the proliferative phase of wound repair, several vital processes occur, including wound filling, the traction of wound edges facilitated by myofibroblasts, and the covering of the wound, also referred to as epithelialization.49,50 At the same time, it is crucial to establish new vascular networks to ensure an adequate supply of oxygen and nutrients to the developing granulation tissue.51 During the remodeling phase, the inflammation gradually diminishes, collagen deposition takes place, and the injured area becomes fully enveloped by new tissue, ultimately forming scar tissue. Collagen type III gradually changes into collagen type I as the newly created tissue grows stronger and more flexible over time, leading to the wound’s full closure.52
 |
Figure 2 The healing process of ulcer wounds. The process of diabetic foot ulcer healing generally goes through four stages: thrombosis, inflammation, proliferation, and matrix remodeling. |
MSCs
MSCs Therapy for DFU
MSCs are a subset of adult stem cells that can be obtained from the bone marrow, brain, adipose tissue, and umbilical cord, among other biological sources. What sets them apart is their capacity to differentiate into cell lines that are not part of the hematopoietic system.62–64 MSCs possess an exceptional ability to differentiate into various cell types, including osteocytes, chondrocytes, and adipocytes. Moreover, they can also give rise to neural cells derived from both the inner and outer layers of the embryo.65 Three main processes underlie the effects of MSCs transplantation on diabetic wound healing: the release of angiogenic agents and cytokines, immune system modulation, and the cell transplantation and differentiation. MSCs can specifically increase local angiogenic factor and cytokine expression and release, assisting in improving diabetes and PAD while also accelerating wound healing at various phases.66,67
Vascular damage and dysfunction are significant contributing factors to DFU. To support the creation of granulation tissue and facilitate the removal of metabolic wastes, which are critical for decreasing ulcer size and accelerating healing, the production and regeneration of new blood vessels in the DFU region are vital. Studies show that MSCs can release a range of cytokines, such as placental growth factor, angiopoietin-1, vascular endothelial growth factor (VEGF), essential fibroblast growth factor (bFGF), stromal cell-derived factor-1 (SDF-1), keratinocyte growth factor-2, insulin-like growth factor-1, and epidermal growth factor (EGF).68–70 These elements are essential for wound healing, angiogenesis promotion, and enhanced microcirculation dynamics. Of them, VEGF is one that is particularly important for vascular regeneration and is essential for the healing of wounds.71
Additionally, MSCs contribute to immune modulation by secreting immunoglobulins IgM and IgG, which help alleviate inflammation in DFU wounds. They also prevent the release of certain inflammatory cytokines by macrophages, including as IL-6, interferon-alpha (IFN-α), and tumor necrosis factor-alpha (TNF-α), while encouraging the release of anti-inflammatory cytokines, such as IL-10 and IL-12, which stimulate immune cells.72,73 Additionally, inflammatory substances such as prostaglandin E2 (PGE2), nitric oxide (NO), tryptophan 2.3-dioxygenase (TDO), transforming growth factor-gamma (TGF-γ), and IL-10 are released by both MSCs and cytotoxic T cells, and they aid in reducing local inflammation brought on by wounds. Furthermore, MSCs have anti-inflammatory properties and promote the resolution of chronic inflammation by preventing the activation of M1 macrophages, which are inflammatory, and increasing the activation of M2 macrophages, which are anti-inflammatory macrophages.74,75
Lastly, by actively differentiating into epithelial and endothelial cells, MSCs can stimulate angiogenesis through paracrine signaling and aid in the repair of DFU. One of the key variables in promoting MSCs’ VEGF-induced differentiation into endothelial cells is the MAPK/ERK signaling pathway.76 Additionally, MSCs are essential for increasing collagen synthesis and helping DFU produce collagen.77 Furthermore, by boosting fibroblast proliferation and functional recovery, MSCs aid in the formation of granulation tissue in DFU. More growth factors and extracellular matrix (ECM) are released because of this process, which eventually aids in tissue repair.78
Characteristics of MSC-Exo
The discovery of extracellular vehicles (EVs) or exosomes has its roots in the 1940s, a period during which these minute vesicles were largely ignored and dismissed as cellular debris. However, it was not until the mid-2000s that exosomes were rediscovered as messengers facilitating intercellular communication, sparking considerable interest and attention in the scientific community.79–81 However, it was not until the mid-2000s that exosomes were rediscovered as messengers facilitating intercellular communication, sparking considerable interest and attention in the scientific community.82–84 Apoptotic bodies, microvesicles, exosomes, and other vesicles are all included in the category of “extracellular vesicles” under this umbrella term. On the other hand, “exosomes” refer especially to vesicles that are produced inside the cell multivesicular bodies (MVBs). Membrane-bound extracellular vesicles, also known as exosomes, typically range in diameter from 30 to 150 nm. However, some studies report slightly different ranges, such as 40–160 nm or 50–200 nm, though they are generally smaller than 200 nm. Numerous cell types secrete them, including brain cells, endothelial cells, epithelial cells, cancer cells, dendritic cells, immunological cells (such as B, T, mast, and dendritic cells), and MSCs.85–89 Exosomes play a crucial function in intercellular communication by acting as carriers of essential information and biomolecules that originate from cellular sources. These large biomolecules are present on the surface of exosomes both internally and externally and consist of a diverse range of constituents such as proteins, enzymes, transcription factors, lipids, extracellular matrix proteins, receptors, DNA, and mRNA (Figure 3). Exosomes can be found in almost all body fluids, including blood, serum, urine, breast milk, amniotic fluid, cerebrospinal fluid, malignant ascites, saliva, bile, and lymphatic fluids, in both healthy and sick states.90–95
 |
Figure 3 The formation process and molecular structure of mesenchymal stem (MSC) cell-derived exosomes. This figure illustrates the biogenesis and composition of exosomes derived from mesenchymal stem cells (MSCs). Exosome formation begins with the generation of early endosomes through endocytosis within MSCs. These endosomes mature into late endosomes and multivesicular bodies (MVBs), which release exosomes via exocytosis. The detailed structure of an MSC-derived exosome is shown on the right, highlighting surface molecules like integrins for cell adhesion and tetraspanins essential for membrane transport. The exosome membrane also includes lipid anchors and surface proteoglycans, which enhance stability. Internally, MSC-derived exosomes carry various biomolecules, including proteins, RNA species (such as mRNA, miRNA, circRNA), and DNA (mtDNA, ssDNA, dsDNA). These components contribute to diverse functional roles, such as immunomodulation, membrane transport, and intercellular communication, making exosomes promising therapeutic agents for regenerative medicine. |
Currently, the most widely used method for exosome isolation is ultracentrifugation, which separates exosomes from culture media or biological fluids using high-speed centrifugation. Although this technique is cost-effective and relatively simple, it has significant drawbacks, including low purity and the potential contamination with cellular debris.90,96 To improve the purity of the separation, density gradient centrifugation is often used. This method forms a gradient in solutions of different densities, allowing for more precise separation of exosomes from other extracellular vesicles.97 While it improves purity, density gradient centrifugation is more time-consuming and yields lower quantities compared to ultracentrifugation98 Immunoaffinity-based methods are also promising, utilizing antibodies that specifically recognize and capture exosome surface markers. This method offers high specificity and can isolate exosomes with minimal contamination.99–101 However, its high cost and the need for specific antibodies limit its widespread application. Additionally, polymer-based precipitation methods, such as using polyethylene glycol (PEG) for isolation, have gained attention due to their simplicity and relatively low cost. These methods can isolate large quantities of exosomes, but they may affect the purity of the exosome preparation. Genetic engineering approaches have also been explored to enhance the functionality of exosomes. By modifying the cells that produce exosomes, exosomes with better therapeutic properties can be generated, and this technology holds great potential in the development of exosome-based therapies for specific clinical applications.102,103
Despite significant progress in the development of exosome preparation methods, challenges remain in ensuring consistency, purity, and scalability of exosome production. Furthermore, the choice of exosome isolation method may depend on the specific application and the source of the exosomes. For example, exosomes derived from different cell types have distinct functional characteristics, and their therapeutic effects may vary based on their composition and bioactivity. Further research has revealed that MSCs have a low survival rate in areas of injured tissue and limited potential for transplantation despite early studies suggesting that they may play a critical role in tissue repair.104 Moreover, MSCs primarily use paracrine signaling to carry out their restorative and regenerative functions. Hence, exosomes derived from mesenchymal sources appear to provide a more stable alternative. These nano-sized exosomes can more efficiently transfer between cells and exert their effects. The bilayer structure of exosomes effectively shields the bioactive substances inside from degradation. Additionally, surface proteins and signaling molecules on exosomes can be modified to target specific organs, tissues, and cells.92,105–107 Exosomes help mitigate potential safety concerns associated with MSCs transplantation, including unintended differentiation, and the risk of malignant transformation. MSCs almost do not express MHC-I, or only express negligible levels of MHC-II molecules. In addition, MSCs do not express cell surface glycoproteins such as Fas ligand, nor do they express co-stimulatory molecules, including CD40/CD40L, B7/CD28, ICOS, 4–1BB, OX40/OX40L, which are essential for the activation of T and B lymphocytes. Therefore, MSCs lacking these molecules do not activate allogeneic T and B lymphocytes, demonstrating their low immunogenicity. Similarly, mesenchymal stem cell-derived exosomes also exhibit low immunogenicity, and the various small interfering RNAs they express can effectively reduce immune responses.108–111 Studies have shown that exosomes derived from human bone marrow mesenchymal stem cells can silence Fas and miR-375 in human islets. These exosomes suppress immune responses by inhibiting the proliferation of peripheral blood mononuclear cells and enhancing the function of regulatory T cells (Tregs).112
MSC-Exo Therapy for DFU
Due to inheriting most of the biological functions of MSCs, MSC-Exo has found widespread application in the treatment of autoimmune diseases, metabolic disorders, ischemic injuries, and tumor-related diseases.113–116 Owing the complexity of DFU’s pathophysiology and clinical symptoms, recovery requires multidisciplinary collaboration and careful, long-term care from medical professionals. Presently, a great deal of research has been published on the use of MSC-Exo to treat DFU. MSC-Exo primarily promotes the healing of DFU (Table 1) and diabetic wounds (Figure 4) by stimulating angiogenesis, extracellular matrix remodeling, cell migration and proliferation, and anti-inflammatory actions.117,118
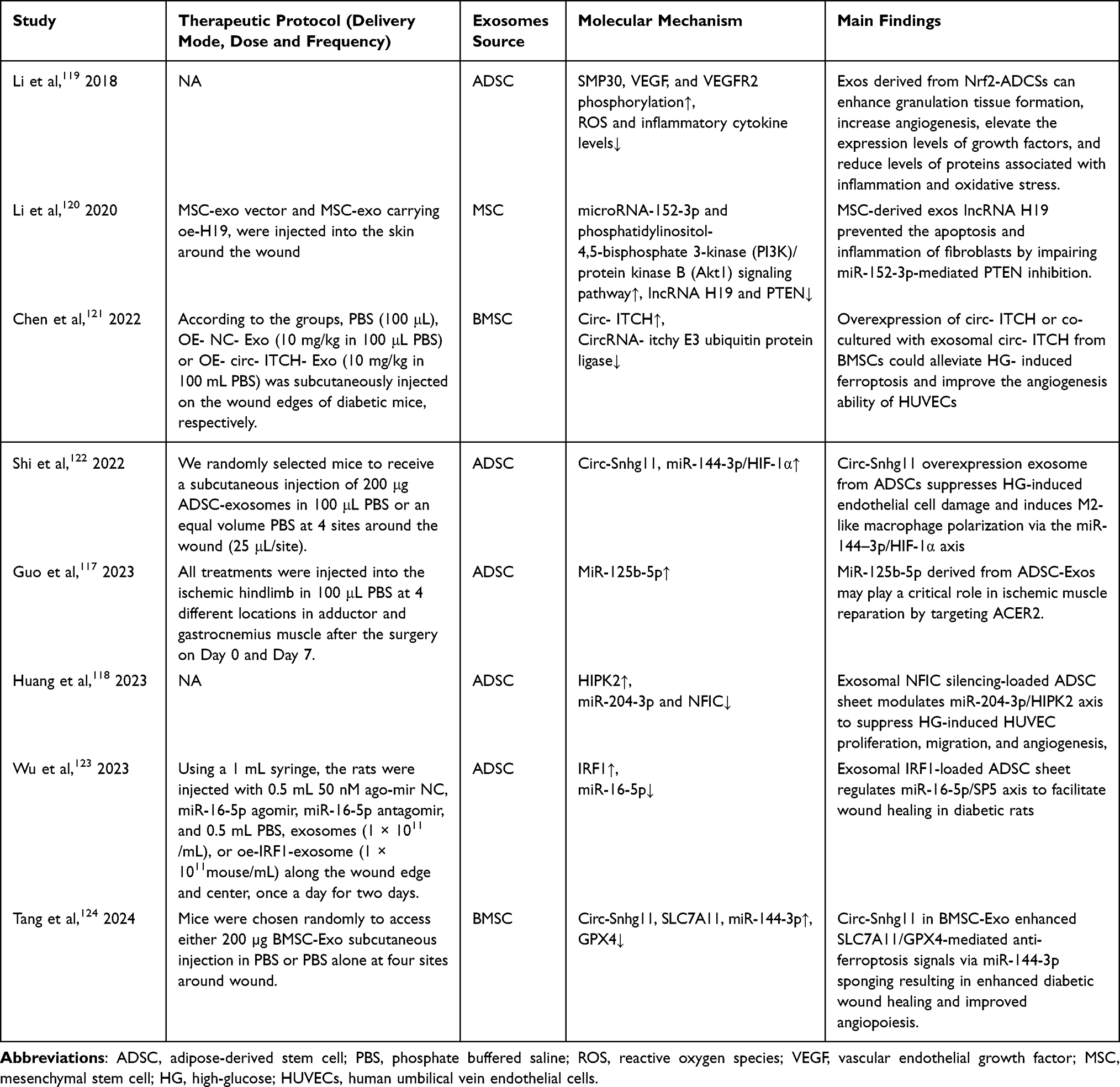 |
Table 1 Studies Related to the Treatment of Diabetic Foot Ulcer with Mesenchymal Stem Cell-Derived Exosomes |
 |
Figure 4 The upregulation and downregulation of active factors, mRNA, and signaling pathways during the ulcer healing process. This figure illustrates the regulatory effects of mesenchymal stem cell-derived exosomes (MSC-Exo) on wound healing, focusing on inflammation, cell proliferation/migration, angiogenesis, and matrix remodeling. The figure depicts the promotion of wound healing by upregulating or inhibiting certain key signaling pathways and molecular mediators. |
Inflammation
The essence of DFU is viewed as a type of chronic inflammatory injury, and mitigating inflammation is crucial for improving DFU outcomes. Macrophages, serving as the primary effectors of the inflammatory process, selectively polarize into M1 or M2 macrophage cells in response to signals from the microenvironment.125 Toll-such receptors (TLRs) and NFκB activation are the main drivers of M1 macrophages’ pro-inflammatory response, which triggers actions such as pathogen phagocytosis, oxidative stress, and intracellular perish. In contrast, the actions of M2 macrophages involve the recruitment of transcription factors like STAT3, which function to suppress inflammation and encourage tissue remodeling.126–128 Studies suggest that in a high glucose environment, there is an enhancement in polarization toward M1 macrophages, which oppresses the polarization of M2 macrophages. This over-polarization of M1 macrophages can lead to the production of inducible nitric oxide synthase (iNOS), IL-1, IL-6, and IL-23, further inhibiting endothelial cell proliferation, migration, and angiogenesis.129–131 It was shown in a study by He et al132 that MSC-Exo infusion into a mouse wound model promoted M2 macrophage polarization. Further investigations revealed that MSC-Exo contained miR-223, which not only inhibited the classical pro-inflammatory pathway but also enhanced alternative anti-inflammatory responses, serving as a novel regulatory factor in macrophage polarization This primarily promoted changes in the protein expression of pknox1 in macrophages, thereby regulating M2 macrophage polarization. Similarly, ADSC-Exosomes achieved comparable effects through the 144-3p/HIF-1α axis.122,133,134 Moreover, miR-146a-5p expression is significantly downregulated in diabetes wounds. MiR-146a-5p has been demonstrated to increase M1 and decrease M2 macrophage polarization when electroporated into bone mesenchymal stem cell-derived exosomes (BMSCs-Exo), thereby facilitating diabetic wound healing.135,136 Lipopolysaccharide (LPS)-treated mesenchymal stem cells (LPS pre-MSCs-Exo) produce exosomes that exhibit enhanced M2 macrophage polarization and anti-inflammatory factor expression. Microarray analysis of LPS pre-MSCs-Exo reveals an upregulation in the expression of the lsa-let-7b molecule, which acts to regulate Toll-like Receptor 4 (TLR4) expression negatively. This mechanism finely tunes the inflammatory response, ultimately promoting wound healing.126,137
The oxidative stress response is another contributing factor to chronic inflammatory damage, complementing the inflammatory response. In a high-glucose environment, glucose and proteins undergo non-enzymatic reactions, resulting in the generation of AGEs. ROS play an important role in the pathophysiology of diabetic foot. They can damage endothelial cells and increase vascular permeability, thereby affecting blood flow supply and increasing the risk of diabetic foot138 Additionally, an increase in ROS can lead to neuronal cell damage, triggering diabetic neuropathy, reducing sensory ability in the feet, and making patients more susceptible to injuries and ulcers. ROS can also activate inflammatory signaling pathways, promoting the release of inflammatory factors, leading to local chronic inflammation and thus delaying wound healing. While an appropriate amount of ROS can promote wound healing, excessive ROS causes cellular damage and hinders healing.139 Therefore, the reasonable regulation of ROS production and clearance is of great significance for improving foot health in diabetic patients.48,140 Research has shown that using immunomodulatory hydrogels to achieve self-cascading glucose depletion and ROS clearance lowers the M1/M2 macrophage ratio, thereby improving the therapeutic effects on the dorsal skin and DFU wounds in diabetic rats.141 Additionally, mitochondria and other cellular compartments produce elevated levels of matrix metalloproteinases (MMPs), ROS, and reactive nitrogen species (RNS), collectively inducing an oxidative stress response. This, in turn, promotes the onset and maintenance of inflammation.39,142 Studies have indicated that adipose-derived stem cell exosomes (ADSCs-Exo) can mitigate MMP production, precise accumulated ROS, and decrease the expression of inflammatory cytokines and adhesion molecules. Moreover, they can enhance the expression of SIRT3 protein in DFU wounds, thereby inhibiting mitochondrial ROS production. Additionally, ADSCs-Exo enhances the activity of superoxide dismutase 2 (SOD2) and reduces ROS accumulation in vascular endothelial cells under high-glucose conditions.143 Furthermore, Nrf2 represents a promising avenue for diabetic foot ulcer treatment. Typically, Nrf2 binds to Kelch-like ECH-associated protein 1 (Keap1). However, during cellular oxidative stress, the binding between Nrf2 and Keap1 is disrupted, allowing Nrf2 to dissociate and translocate into the nucleus. The expression of antioxidant and cytoprotective genes, such as heat shock proteins, detoxification enzymes, and antioxidant enzymes, is then activated when Nrf2 binds to antioxidant response elements (AREs). This process aids in clearing free radicals, alleviating oxidative stress, mitigating cell damage, and reducing inflammation. Overexpression of Nrf2 in ADSC-Exo applied to diabetic ulcer mouse models has demonstrated a significant reduction in inflammation and oxidative stress-related protein levels.119,121,144,145
Proliferation
During the proliferation phase, the resolution of inflammation gives way to the gradual proliferation of tissue repair cells. This phase is predominantly characterized by the physiological process of granulation tissue formation and the proliferation and migration of epidermal cells. Histologically, this phase is distinguished by the formation of capillary buds and fibroblast proliferation, accompanied by the production of an abundant extracellular matrix. Research suggests that mRNA, microRNA, and various proteins within exosomes play crucial roles in angiogenesis during this phase. Tang et al124 demonstrated that the expression of circ-Snhg11 in diabetic wounds was upregulated after treatment with BMSCs-Exo. This upregulation alleviated the inhibition of endothelial progenitor cell (EPC) mobilization caused by miR-144-3P overexpression and enhanced the SLC7A11/GPX4-mediated anti-ferroptosis signal. Conversely, overexpression of miR-16-5p downregulated the expression of SP5, thereby promoting fibroblast proliferation and migration.123 Fibroblasts play a crucial role in tissue repair, from the late stage of inflammation to the final process of extracellular matrix remodeling. They interact with various cell types, including keratinocytes, endothelial cells, adipocytes, mast cells, and extracellular matrix components like collagen, to promote proliferation. Studies suggest that MSC-Exo encourage fibroblast proliferation and migration and inhibit apoptosis by activating signaling pathways such as Notch, ERK, AKT, STAT-3, and Wnt/β-catenin. Additionally, heat shock protein 90 (HSP90) on the exosomal membrane can bind to the low-density lipoprotein receptor-related protein 1 (LRP1) receptor on recipient cell membranes, enhancing the AKT and ERK signaling pathways in fibroblasts.146–148 Moreover, studies have demonstrated that ADSCs-Exo can stimulate monocytes/macrophages to secrete more TGF-β1, thereby activating the TGF-β/Smad3 signaling pathway. Once activated by exosomes containing TGF-β1 from ASCs, fibroblasts then secrete TGF-β1 protein in an autocrine manner, thereby promoting fibroblast proliferation and activation.149
Angiogenesis plays a pivotal role in cell proliferation, migration, wound healing, and tissue engineering during the process of proliferation. It serves as an essential requirement for the formation of granulation tissue.150 In their study, Chen et al151 identified a protein known as deleted in malignant brain tumors 1 (DMBT1) by clustering and enriching proteins from human urine-derived stem cell exosomes (HUSC-Exo). DMBT1 was found to promote angiogenesis by enhancing endothelial cell proliferation, migration, and adhesion, resulting in pronounced angiogenesis observed in DFU mouse models. DMBT1 also triggers an angiogenic response in lung alveolar tissue by stimulating the production of VEGFA. Moreover, both VEGFA and the PI3K-Akt signaling pathway are downstream targets influenced by DMBT1. Furthermore, MSC-Exo harbor a diverse array of pro-angiogenic proteins, including VEGFA, SRPX2, and ANGPTL4, which play pivotal roles in neovascularization.152–154 MSC-Exo treated with atorvastatin activates the AKT/eNOS pathway and enhances the expression of miR-221-3p. This activation facilitates the proliferation, migration, and tube formation of endothelial cells while elevating vascular endothelial growth factor levels. Similarly, MSC-Exo treated with pioglitazone stimulate angiogenesis via the same signaling pathway while suppressing the expression of the PTEN protein.155,156 ADSC-Exo overexpressing circular RNA mmu_circ_0000250 has been confirmed to exert pro-angiogenic effects by upregulating SIRT1 and miR-128-3p in DFU.157
Matrix Remodeling
The extracellular matrix (ECM) is restructured during the crucial last stage of wound healing, known as matrix remodeling, to improve the stability and strength of the injured area. PI3K/Akt is a key component in mechanistic remodeling. Exosomes derived from MSCs in DFU contain long non-coding RNA H19 (lncRNA H19), which activates the PI3K/Akt signaling pathway to promote the proliferation and migration of fibroblasts, while inhibiting apoptosis and inflammation. This process ultimately accelerates wound healing in DFU mice.120 Wang et al found that exosomes derived from hypoxic adipose-derived stem cells (HypADSCs) exhibit downregulation of miRNA-99b and miRNA-146a, as well as upregulation of miRNA-21-3p, miRNA-126-5p, and miRNA-31-5p, compared to exosomes derived from ADSCs. These miRNAs activate the PI3K/Akt signaling pathway, promoting the proliferation and migration of fibroblasts, enhancing the secretion of VEGF and extracellular matrix, thereby improving the healing quality of diabetic wounds.158,159
According to Wang et al160 mice’s incision scars shrank in size when ADSC-Exo was injected intravenously, and the ratio of collagen III to collagen I rose. Fibroblasts also can absorb and internalize ADSCs-Exo, which in a dose-dependent way promotes collagen synthesis, cell migration, and proliferation. It boosts the ratio of transforming TGF-β3 to TGF-β1 in vivo, inhibits fibroblasts from developing into myofibroblasts, and promotes the expression of the N-cadherin and Cyclin D1 genes. Additionally, ADSCs-Exo upregulates the expression of skin MMP3, leading to an increased ratio of MMP3 to tissue inhibitor of TIMP1, thus promoting ECM remodeling. Extracellular vesicles from human-induced pluripotent stem cell-derived mesenchymal stem cells also promote skin healing by enhancing collagen synthesis.161,162
However, it has been reported that miR-21 can suppress the expression of TIMP3 and TIAM1 and is essential for the keratinocyte migration induced by TGF-β.163 In contrast, According to separate findings by Lv et al164 different results that the expression of miR-21-5P contained in MSC-Exo can activate the Wnt/β-catenin signaling pathway and upregulate the expression of MMP-7, which would greatly enhance keratinocyte migration and aid in the re-epithelialization of diabetic wounds. The Wnt/β-catenin signaling pathway is closely associated with cell adhesion, intercellular communication, cell proliferation, and cell migration. Previous research has shown that ADSCs-Exo significantly increases wound re-epithelialization and collagen deposition, thus accelerating wound healing through the AKT and extracellular signal-regulated kinase (ERK) signaling pathways. This has also been validated in a rabbit model of DFU.165,166 Furthermore, BMSCs-Exo can accelerate re-epithelialization and enrich the expression of cytokeratin 19 (CK19), proliferating cell nuclear antigen (PCNA), and collagen I in vivo.167 Umbilical cord blood mesenchymal stem cells-derived Exosomes (UCB-MSCs-Exo) expedite wound closure, reduce scar formation, improve the regeneration of skin appendages, nerves, and blood vessels, and regulate the natural distribution of collagen fibers during wound healing. Functional analysis suggests that miRNAs derived from UCB-MSCs are closely associated with the transforming TGF-β signaling pathway, which can induce the differentiation of myofibroblasts. Furthermore, UCB-MSCs-Exo promotes skin cell migration and proliferation both in vivo and in vitro while suppressing the overproduction of myofibroblasts and collagen I.168
Biological Engineering Applications of MSC-Exo
Despite the significant advantages exhibited by MSC-Exo in treating DFU, challenges persist. Common administration routes for exosomes include intravenous injection, local injection, and intraperitoneal injection. However, DFU healing cycles are lengthy, while exosomes have a short half-life and rapid clearance rates, significantly limiting their biological function. Therefore, there has been considerable interest in combining exosomes with convenient biocompatible materials to preserve their biological activity. Studies have shown that adding exosomes to hydrogels can enhance and maintain the stability of proteins, mRNA, DNA, and other biological components within the exosomes.169 Hydrogels are highly hydrophilic three-dimensional networks formed by physical or chemical crosslinking of hydrophilic polymers. They possess properties such as anti-infection, absorption of wound exudates, maintenance of moisture balance and gas exchange, as well as the ability to encapsulate, protect, and deliver bioactive molecules. Therefore, hydrogels are considered ideal skin substitutes and wound dressings. Additionally, locally injecting hydrogels containing exosomes into injured tissues is a commonly used administration method. This approach allows for sustained local release of exosomes, promoting the repair and regeneration of damaged tissues.170,171
 |
Table 2 Studies Related to the Treatment of Diabetic Wounds with Mesenchymal Stem Cell-Derived Exosomes in Combination with Hydrogels |
Currently, research on MSC-Exo for treating DFU is limited, with most studies focused primarily on diabetic wounds. The combination of MSC-Exo with hydrogels shows promising therapeutic effects in the treatment of diabetic wound173,174 (Table 2). Zhao et al176 incorporated HUVECs-Exo into GelMA hydrogels, creating an exosome-loaded hydrogel dressing for wound management. Fluorescence microscopy and three-dimensional reconstruction images revealed a uniform distribution of Exosomes within the hydrogel with sustained release. Moreover, compared to the sole Exo group, mice diabetic wound models treated with this composite showed accelerated wound healing. Interestingly, the hydrogel itself seemed to play a role in promoting diabetic wound healing. Li et al172 encapsulated exosomes derived from synovial mesenchymal stem cells (SMSCs) with high expression of miR-126-3p in hydroxyapatite/chitosan (HAP-CS) composite hydrogel (HAP-CS-SMSCs-126-Exos). Results demonstrated that HAP-CS hydrogel alone promoted angiogenesis, increased wound closure rate, enhanced re-epithelialization, maturation of granulation tissue, collagen arrangement, and deposition, exhibiting synergistic effects with exosomes. Additionally, research has utilized human adipose-derived stem cells (HASCs) to prepare hydrogel scaffolds. Prior to gelation, it exists as a liquid solution, facilitating cell encapsulation. Its sol-gel transition properties favor cell delivery and site adherence. Derived from adipose tissue, it contains native ECM, promoting interaction and dynamic communication with stem cells. It plays a vital role in tissue repair and regeneration by recruiting endogenous stem and progenitor cells, exhibiting antimicrobial activity, and modulating macrophage polarization.175,177
Local administration of MSC-Exo indeed offers numerous advantages, such as increasing local concentrations, reducing systemic side effects, and ease of use. However, local delivery also faces the issue of exosome loss. Due to their poor stability, exosomes are susceptible to enzyme degradation, diffusion loss, and other factors. Direct local application may lead to the degradation of some exosomes in the wound environment or their diffusion to non-target areas, thereby reducing their bioavailability and therapeutic effects. This issue is particularly prominent in DFU wounds, which have high permeability and a chronic inflammatory environment. Therefore, finding suitable delivery carriers to enhance exosome retention time and bioactivity is essential. Combining MSC-Exo with advanced biomaterials, especially hydrogels, nanoparticle carriers, and biodegradable scaffolds, can not only prolong the action time of exosomes in the body but also enhance their stability and targeting. Several studies have explored bioengineering-based MSC-Exo delivery systems, showing promising applications in regenerative medicine and wound healing. For example, nano-hydrogel combined with exosomes can provide controlled release, offering precise temporal and spatial delivery in DFU treatment. 3D-printed biomaterial scaffolds can load exosomes, providing mechanical support and improving the wound microenvironment. Microneedle patch technology combined with exosomes can penetrate the skin barrier for local precise delivery, improving the treatment outcomes of diabetic chronic wounds. Unfortunately, there is still a lack of studies comparing MSC-Exo with current standard treatments for DFU, such as wound dressings, antibiotics, and hyperbaric oxygen therapy.
Limitations and Clinical Application Prospects
Although mesenchymal stem cell-derived exosomes have been widely validated for their efficacy in diabetic foot ulcers, there are still unavoidable limitations. Firstly, the preparation and purification techniques of exosomes face certain technical bottlenecks. How to increase yield while ensuring the activity of exosomes is an urgent problem that needs to be addressed. Additionally, the heterogeneity of exosomes makes standardized production more difficult. The exosome preparations used in different studies vary greatly, leading to lower reproducibility of clinical trial results. Secondly, among the studies included in this research, there are certain differences in the injection doses of exosomes. Currently, there are no clear guidelines to guide standard usage doses, and little is known about their metabolic pathways, distribution characteristics, and excretion methods after entering the body. Understanding their pharmacokinetic properties is crucial for evaluating the safety and effectiveness of exosomes, but related research is currently lacking. Moreover, although MSC exosomes have shown good therapeutic effects in animal models, their long-term safety and efficacy in humans remain unclear. Whether exosome therapy can maintain efficacy during long-term use and whether there are potential side effects need to be verified through more clinical trials. Finally, since exosome therapy is an emerging biotechnology, existing drug regulatory policies are not yet well-developed for its standardization. Exosome products used for the treatment of human diseases and conditions in the United States are regulated as drugs under the Federal Food, Drug, and Cosmetic Act (FD&C Act) and as biological products under the Public Health Service Act (PHS Act), requiring pre-market review and approval. According to European directives (Directive 2001/83/EC and Regulation 1394/2007/EC), exosomes purified directly from cells or extracted from transcriptional RNA are not considered advanced therapy medicinal products (ATMPs), but are classified as biological products. In Japan, exosomes are classified as biological products and are subject to the same regulatory requirements as vaccines, blood products, and other biological products. For medical topical formulations, non-commercial clinical studies are managed by physicians on a case-by-case basis, as these formulations lack live cell components and are not considered specifically processed cells. Exosomes are typically derived from human or animal cells, particularly MSCs, so their donor source, method of collection, and ethical approval need to be strictly regulated to ensure informed consent from donors and compliance with ethical standards. Ensuring the quality, safety, and effectiveness of exosome preparations in clinical applications is a challenge faced by regulatory authorities, necessitating the development of further policies and standards in the future.
To overcome the above limitations, future research can proceed from the following aspects: (1) In-depth exploration of mechanisms of action: Utilize high-throughput omics technologies (such as proteomics, metabolomics, and miRNA omics) to systematically analyze the composition and mechanisms of exosomes, providing a theoretical basis for optimizing exosome therapy protocols. (2) Optimize preparation and purification processes: Enhance exosome purification technology to ensure their stability and biological activity, while establishing unified production standards to promote the widespread clinical application of exosome preparations.178 (3) Conduct large-scale clinical trials: Under the premise of ensuring the safety of exosome therapy, carry out multicenter, large-sample randomized controlled clinical trials to verify its efficacy and long-term safety.
MSC-Exo therapy can reduce the consumption of biological materials. Traditional wound healing treatments often rely on disposable dressings, antibiotics, and other medical resources, whereas exosome therapy has the potential to minimize the use of these consumables, thereby reducing medical waste and alleviating environmental burdens. Additionally, by accelerating wound healing, this therapy shortens hospital stays and reduces the number of treatments required, indirectly lowering energy consumption and carbon emissions within the healthcare system. This advantage in reducing medical resource consumption makes MSC-Exo therapy highly promising for sustainable healthcare applications.
Conclusion
In summary, MSC-Exo, a crucial type of extracellular vesicle, plays a significant role in the anti-inflammatory process during DFU healing. It promotes the proliferation and migration of endothelial cells, fibroblasts, and other cells involved in angiogenesis and extracellular matrix remodeling through various cytokines, mRNA, proteins, and related signaling pathways. MSC-Exo exhibits favorable biocompatibility and stability, offering a promising new avenue for DFU treatment. However, efforts to enhance its efficacy through various hydrogels and develop industrial-scale production methods for extracting exosomes, as well as establishing standardized processes for storage, transportation, and maintaining biological activity, are necessary. Combining exosomes with other biotechnologies and formulating standardized therapeutic regimens and dosages tailored to DFU patients, along with assessing synergistic effects with other treatment modalities, require further extensive research. Additionally, while much of the current research is limited to in vitro and rodent studies, advancing studies in primates and human clinical trials is essential to evaluate the therapeutic efficacy of MSC-Exo further. Nevertheless, as a promising therapeutic modality for DFU, MSC-Exo holds vast potential and deserves further investigation.
Acknowledgment
Hui Wang and Sensen Wu are co-first authors for this study. We sincerely thank to all the authors for their hard work on this manuscript. The authors declare that they have not use AI-generated work in this manuscript.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This research was funded by the National Key Research and Development Program of China (2021YFC2500500) and Beijing Municipal Science & Technology Commission (No. Z241100009024028).
Disclosure
No, there is no conflict of interest.
References
1. Du S, Zeugolis DI, O’Brien T. Scaffold-based delivery of mesenchymal stromal cells to diabetic wounds. Stem Cell Research & Therapy. 2022;13(1):426. doi:10.1186/s13287-022-03115-4
2. Ogurtsova K, Guariguata L, Barengo NC, et al. IDF diabetes Atlas: global estimates of undiagnosed diabetes in adults for 2021. Diabetes Research and Clinical Practice. 2022;183:109118. doi:10.1016/j.diabres.2021.109118
3. Zheng Y, Ley SH, Hu FB. Global aetiology and epidemiology of type 2 diabetes mellitus and its complications. Nature Reviews Endocrinology. 2018;14(2):88–98. doi:10.1038/nrendo.2017.151
4. Ali MK, Pearson-Stuttard J, Selvin E, Gregg EW. Interpreting global trends in type 2 diabetes complications and mortality. Diabetologia. 2022;65(1):3–13. doi:10.1007/s00125-021-05585-2
5. Yazdanpanah L, Nasiri M, Adarvishi S. Literature review on the management of diabetic foot ulcer. World Journal of Diabetes. 2015;6(1):37–53. doi:10.4239/wjd.v6.i1.37
6. Jhamb S, Vangaveti VN, Malabu UH. Genetic and molecular basis of diabetic foot ulcers: clinical review. Journal of Tissue Viability. 2016;25(4):229–236. doi:10.1016/j.jtv.2016.06.005
7. Sun H, Saeedi P, Karuranga S, et al. IDF Diabetes Atlas: global, regional and country-level diabetes prevalence estimates for 2021 and projections for 2045. Diabetes Research and Clinical Practice. 2022;183:109119. doi:10.1016/j.diabres.2021.109119
8. Kalan LR, Meisel JS, Loesche MA, et al. Strain- and Species-Level Variation in the Microbiome of Diabetic Wounds Is Associated with Clinical Outcomes and Therapeutic Efficacy. Cell Host & Microbe. 2019;25(5):641–655.e645. doi:10.1016/j.chom.2019.03.006
9. Wang H, Xu Z, Zhao M, Liu G, Wu J. Advances of hydrogel dressings in diabetic wounds. Biomaterials Science. 2021;9(5):1530–1546. doi:10.1039/D0BM01747G
10. Álvaro-Afonso FJ, García-álvarez Y, Tardáguila-García A, García-Madrid M, López-Moral M, Lázaro-Martínez JL. Bacterial Diversity and Antibiotic Resistance in Patients with Diabetic Foot Osteomyelitis. Antibiotics. 2023;12(2):212. doi:10.3390/antibiotics12020212
11. Cortes-Penfield NW, Armstrong DG, Brennan MB, et al. Evaluation and Management of Diabetes-related Foot Infections. Clinical Infectious Diseases: an Official Publication of the Infectious Diseases Society of America. 2023;77(3):e1–e13. doi:10.1093/cid/ciad255
12. Armstrong DG, Tan TW, Boulton AJM, Bus SA. Diabetic Foot Ulcers: a Review. JAMA. 2023;330(1):62–75. doi:10.1001/jama.2023.10578
13. Dinoto E, Ferlito F, La Marca MA, et al. The Role of Early Revascularization and Biomarkers in the Management o f Diabetic Foot Ulcers: a Single Center Experience. Diagnostics (Basel). 2022;12(2):538. doi:10.3390/diagnostics12020538
14. Kostanyan GM, Khramykh TP, Gaygiev TI, Ermolaev PA, Kolbina MV, Memus KP. Endovascular treatment for critical limb ischemia in patients with diabetes mellitus: new opportunities and prospects. Khirurgiia. 2022;2022(10):99.
15. Spiliopoulos S, Festas G, Paraskevopoulos I, Mariappan M, Brountzos E. Overcoming ischemia in the diabetic foot: minimally invasive treatment options. World Journal of Diabetes. 2021;12(12):2011–2026. doi:10.4239/wjd.v12.i12.2011
16. Shafiee S, Heidarpour M, Sabbagh S, et al. Stem cell transplantation therapy for diabetic foot ulcer: a narrative review. Asian Biomedicine: Research, Reviews and News. 2021;15(1):3–18. doi:10.2478/abm-2021-0002
17. Mohamad Yusoff F, Higashi Y. Mesenchymal Stem/Stromal Cells for Therapeutic Angiogenesis. Cells. 2023;12(17):2162. doi:10.3390/cells12172162
18. Mikłosz A, Chabowski A. Efficacy of adipose-derived mesenchymal stem cell therapy in the treatment of chronic micro- and macrovascular complications of diabetes. Diabetes, Obesity & Metabolism. 2024;26(3):793–808. doi:10.1111/dom.15375
19. Huang Y, Wu Q, Tam PKH. Immunomodulatory Mechanisms of Mesenchymal Stem Cells and Their Potent ial Clinical Applications. International Journal of Molecular Sciences. 2022;23(17):10023. doi:10.3390/ijms231710023
20. Zhao L, Guo Z, Chen K, et al. Combined Transplantation of Mesenchymal Stem Cells and Endothelial Col ony-Forming Cells Accelerates Refractory Diabetic Foot Ulcer Healing. Stem Cells Int. 2020;2020:8863649. doi:10.1155/2020/8863649
21. Zhang C, Huang L, Wang X, et al. Topical and intravenous administration of human umbilical cord mesenchymal stem cells in patients with diabetic foot ulcer and peripheral arterial disease: a Phase I pilot study with a 3-year follow-up. Stem Cell Research & Therapy. 2022;13(1):451. doi:10.1186/s13287-022-03143-0
22. Arango-Rodríguez ML, Mateus LC, Sossa CL, et al. A novel therapeutic management for diabetes patients with chronic limb-threatening ischemia: comparison of autologous bone marrow mononuclear cells versus allogenic Wharton jelly-derived mesenchymal stem cells. Stem Cell Research & Therapy. 2023;14(1):221. doi:10.1186/s13287-023-03427-z
23. Huerta CT, Voza FA, Ortiz YY, Liu ZJ, Velazquez OC. Mesenchymal stem cell-based therapy for non-healing wounds due to chronic limb-threatening ischemia: a review of preclinical and clinical studies. Frontiers in Cardiovascular Medicine. 2023;10:1113982. doi:10.3389/fcvm.2023.1113982
24. Chen J, Liu Y, Zhang J, et al. External Application of Human Umbilical Cord-Derived Mesenchymal Stem Cells in Hyaluronic Acid Gel Repairs Foot Wounds of Types I and II Dia betic Rats Through Paracrine Action Mode. Stem Cells Transl Med. 2023;12(10):689–706. doi:10.1093/stcltm/szad050
25. Wei L, Xu Y, Zhang L, Yang L, Zhao RC, Zhao D. Mesenchymal Stem Cells Promote Wound Healing and Effects on Expression of Matrix Metalloproteinases-8 and 9 in the Wound Tissue of Diabetic Rats. Stem Cells and Development. 2023;32(1–2):25–31. doi:10.1089/scd.2021.0218
26. Ha DH, Kim H-K, Lee J, et al. Mesenchymal Stem/Stromal Cell-Derived Exosomes for Immunomodulatory Th erapeutics and Skin Regeneration. Cells. 2020;9(5):1157. doi:10.3390/cells9051157
27. Wu J, Chen L-H, Sun S-Y, Li Y, Ran X-W. Mesenchymal stem cell-derived exosomes: the Dawn of diabetic wound hea ling. World Journal of Diabetes. 2022;13(12):1066–1095. doi:10.4239/wjd.v13.i12.1066
28. Las Heras K, Royo F, Garcia-Vallicrosa C, et al. Extracellular vesicles from hair follicle-derived mesenchymal stromal cells: isolation, characterization and therapeutic potential for chronic wound healing. Stem Cell Research & Therapy. 2022;13(1):147. doi:10.1186/s13287-022-02824-0
29. Sun B, Wu F, Wang X, et al. An Optimally Designed Engineering Exosome-Reductive COF Integrated Nanoagent for Synergistically Enhanced Diabetic Fester Wound Healing. Small. 2022;18(26):e2200895. doi:10.1002/smll.202200895
30. Ahmadieh-Yazdi A, Karimi M, Afkhami E, et al. Unveiling therapeutic potential: adipose tissue-derived mesenchymal st em cells and their exosomes in the management of diabetes mellitus, wo und healing, and chronic ulcers. Biochem Pharmacol. 2024;226:116399. doi:10.1016/j.bcp.2024.116399
31. Zhang X, Gan J, Fan L, Luo Z, Zhao Y. Bioinspired Adaptable Indwelling Microneedles for Treatment of Diabetic Ulcers. Adv Mater. 2023;35(23):e2210903. doi:10.1002/adma.202210903
32. Parveen K, Hussain MA, Anwar S, Elagib HM, Kausar MA. Comprehensive review on diabetic foot ulcers and neuropathy: treatment, prevention and management. World Journal of Diabetes. 2025;16(3):100329. doi:10.4239/wjd.v16.i3.100329
33. Reardon R, Simring D, Kim B, Mortensen J, Williams D, Leslie A. The diabetic foot ulcer. Aust J Gen Pract. 2020;49(5):250–255. doi:10.31128/AJGP-11-19-5161
34. Volmer-Thole M, Lobmann R. Neuropathy and Diabetic Foot Syndrome. International Journal of Molecular Sciences. 2016;17(6):917. doi:10.3390/ijms17060917
35. Fitridge R, Chuter V, Mills J, et al. The intersocietal IWGDF, ESVS, SVS guidelines on peripheral artery dis ease in people with diabetes and a foot ulcer. Diabetes Metab Res Rev. 2024;40(3):e3686. doi:10.1002/dmrr.3686
36. Li Y, Liu Y, Liu S, et al. Diabetic vascular diseases: molecular mechanisms and therapeutic strat egies. Signal Transduction and Targeted Therapy. 2023;8(1):152. doi:10.1038/s41392-023-01400-z
37. Gao JJ, Wu FY, Liu YJ, et al. Increase of PCSK9 expression in diabetes promotes VEGFR2 ubiquitination to inhibit endothelial function and skin wound healing. Science China Life Sciences. 2024;67(12):2635–2649. doi:10.1007/s11427-023-2688-8
38. Bandyk DF. The diabetic foot: pathophysiology, evaluation, and treatment. Seminars in Vascular Surgery. 2018;31(2–4):43–48. doi:10.1053/j.semvascsurg.2019.02.001
39. Deng L, Du C, Song P, et al. The Role of Oxidative Stress and Antioxidants in Diabetic Wound Healin g. Oxid Med Cell Longev. 2021;2021(1):8852759. doi:10.1155/2021/8852759
40. Lupachyk S, Shevalye H, Maksimchyk Y, Drel VR, Obrosova IG. PARP inhibition alleviates diabetes-induced systemic oxidative stress and neural tissue 4-hydroxynonenal adduct accumulation: correlation with peripheral nerve function. Free Radical Biology & Medicine. 2011;50(10):1400–1409. doi:10.1016/j.freeradbiomed.2011.01.037
41. Ma J, Pan P, Anyika M, Blagg BS, Dobrowsky RT. Modulating Molecular Chaperones Improves Mitochondrial Bioenergetics and Decreases the Inflammatory Transcriptome in Diabetic Sensory Neurons. ACS Chem Neurosci. 2015;6(9):1637–1648. doi:10.1021/acschemneuro.5b00165
42. André-Lévigne D, Pignel R, Boet S, Jaquet V, Kalbermatten DF, Madduri S. Role of Oxygen and Its Radicals in Peripheral Nerve Regeneration: from Hypoxia to Physoxia to Hyperoxia. International Journal of Molecular Sciences. 2024;25(4):2030. doi:10.3390/ijms25042030
43. Armstrong DG, Boulton AJM, Bus SA. Diabetic Foot Ulcers and Their Recurrence. The New England Journal of Medicine. 2017;376(24):2367–2375. doi:10.1056/NEJMra1615439
44. Miceli G, Basso MG, Pennacchio AR, et al. The Potential Impact of SGLT2-I in Diabetic Foot Prevention: promising Pathophysiologic Implications, State of the Art, and Future Perspecti ves-A Narrative Review. Medicina (Kaunas). 2024;60(11):1796. doi:10.3390/medicina60111796
45. Haris F, Jan Y-K, Liau B-Y, et al. Plantar pressure gradient and pressure gradient angle are affected by inner pressure of air insole. Front Bioeng Biotechnol. 2024;12:1353888. doi:10.3389/fbioe.2024.1353888
46. Ren Y, Wang H, Song X, Wu Y, Lyu Y, Zeng W. Advancements in diabetic foot insoles: a comprehensive review of design, manufacturing, and performance evaluation. Front Bioeng Biotechnol. 2024;12:1394758. doi:10.3389/fbioe.2024.1394758
47. Samakidou G, Eleftheriadou I, Tentolouris A, Papanas N, Tentolouris N. Rare diabetic neuropathies: it is not only distal symmetrical polyneur opathy. Diabetes Research and Clinical Practice. 2021;177:108932. doi:10.1016/j.diabres.2021.108932
48. Xu Z, Liu Y, Ma R, et al. Thermosensitive Hydrogel Incorporating Prussian Blue Nanoparticles Pro motes Diabetic Wound Healing via ROS Scavenging and Mitochondrial Func tion Restoration. ACS Appl Mater Interfaces. 2022;14(12):14059–14071. doi:10.1021/acsami.1c24569
49. Ding X, Jian T, Wu Y, et al. Ellagic acid ameliorates oxidative stress and insulin resistance in hi gh glucose-treated HepG2 cells via miR-223/keap1-Nrf2 pathway. Biomedicine & Pharmacotherapy = Biomedecine & Pharmacotherapie. 2019;110:85–94. doi:10.1016/j.biopha.2018.11.018
50. Lin S, Zhang Q, Li S, et al. Antioxidative and Angiogenesis-Promoting Effects of Tetrahedral Framew ork Nucleic Acids in Diabetic Wound Healing with Activation of the Akt/Nrf2/HO-1 Pathway. ACS Appl Mater Interfaces. 2020;12(10):11397–11408. doi:10.1021/acsami.0c00874
51. Xu C, Hu L, Zeng J, et al. Gynura divaricata (L.) DC. promotes diabetic wound healing by activati ng Nrf2 signaling in diabetic rats. J Ethnopharmacol. 2024;323:117638. doi:10.1016/j.jep.2023.117638
52. Akhter N, Wilson A, Arefanian H, et al. Endoplasmic Reticulum Stress Promotes the Expression of TNF-α in THP-1 Cells by Mechanisms Involving ROS/CHOP/HIF-1α and MAPK/NF-κB Pathways. International Journal of Molecular Sciences. 2023;24(20):15186. doi:10.3390/ijms242015186
53. Huang X, Liang P, Jiang B, et al. Hyperbaric oxygen potentiates diabetic wound healing by promoting fibr oblast cell proliferation and endothelial cell angiogenesis. Life Sciences. 2020;259:118246. doi:10.1016/j.lfs.2020.118246
54. Song J, Zeng J, Zheng S, et al. Sanguisorba officinalis L. promotes diabetic wound healing in rats through inflammation response mediated by macrophage. Phytotherapy Research: PTR. 2023;37(9):4265–4281. doi:10.1002/ptr.7906
55. Geng K, Ma X, Jiang Z, et al. WDR74 facilitates TGF-β/Smad pathway activation to promote M2 macropha ge polarization and diabetic foot ulcer wound healing in mice. Cell Biol Toxicol. 2023;39(4):1577–1591. doi:10.1007/s10565-022-09748-8
56. Zhao Q, Xu J, Han X, Zhang Z, Qu J, Cheng Z. Growth differentiation factor 10 induces angiogenesis to promote wound healing in rats with diabetic foot ulcers by activating TGF-β1/Smad3 signaling pathway. Frontiers in Endocrinology. 2023;13:1013018. doi:10.3389/fendo.2022.1013018
57. Cao M, Duan Z, Wang X, Gong P, Zhang L, Ruan B. Curcumin Promotes Diabetic Foot Ulcer Wound Healing by Inhibiting miR- 152-3p and Activating the FBN1/TGF-β Pathway. Mol Biotechnol. 2024;66(5):1266–1278. doi:10.1007/s12033-023-01027-z
58. Li J, Zhang J, Yang M, et al. Kirenol alleviates diabetic nephropathy via regulating TGF-β/Smads and the NF-κB signal pathway. Pharm Biol. 2022;60(1):1690–1700. doi:10.1080/13880209.2022.2112239
59. Zhang S, Xu Y, Zhang Junior C, Chen X, Zhu J. Dang-Gui-Si-Ni decoction facilitates wound healing in diabetic foot ul cers by regulating expression of AGEs/RAGE/TGF-β/Smad2/3. Arch Dermatol Res. 2024;316(7):338. doi:10.1007/s00403-024-03021-0
60. Jin Y, Huang Y, Zeng G, et al. Advanced glycation end products regulate macrophage apoptosis and infl uence the healing of diabetic foot wound through miR-361-3p/CSF1R and PI3K/AKT pathway. Heliyon. 2024;10(2):e24598. doi:10.1016/j.heliyon.2024.e24598
61. Zhang E, Gao B, Yang L, Wu X, Wang Z. Notoginsenoside Ft1 Promotes Fibroblast Proliferation via PI3K/Akt/mTOR Signaling Pathway and Benefits Wound Healing in Genetically Diabetic Mice. J Pharmacol Exp Ther. 2016;356(2):324–332. doi:10.1124/jpet.115.229369
62. Ciuffi S, Zonefrati R, Brandi ML. Adipose stem cells for bone tissue repair. Clin Cases Miner Bone Metab. 2017;14(2):217–226. doi:10.11138/ccmbm/2017.14.1.217
63. Naji A, Eitoku M, Favier B, Deschaseaux F, Rouas-Freiss N, Suganuma N. Biological functions of mesenchymal stem cells and clinical implicatio ns. Cell mol Life Sci. 2019;76(17):3323–3348. doi:10.1007/s00018-019-03125-1
64. Zhang Z, Yang X, Cao X, Qin A, Zhao J. Current applications of adipose-derived mesenchymal stem cells in bone repair and regeneration: a review of cell experiments, animal models, and clinical trials. Front Bioeng Biotechnol. 2022;10:942128. doi:10.3389/fbioe.2022.942128
65. Zhao L, Ni B, Li J, et al. Evaluation of the impact of customized serum-free culture medium on th e production of clinical-grade human umbilical cord mesenchymal stem c ells: insights for future clinical applications. Stem Cell Research & Therapy. 2024;15(1):327. doi:10.1186/s13287-024-03949-0
66. Dekoninck S, Blanpain C. Stem cell dynamics, migration and plasticity during wound healing. Nat Cell Biol. 2019;21(1):18–24. doi:10.1038/s41556-018-0237-6
67. Mazini L, Rochette L, Admou B, Amal S, Malka G. Hopes and Limits of Adipose-Derived Stem Cells (ADSCs) and Mesenchymal Stem Cells (MSCs) in Wound Healing. International Journal of Molecular Sciences. 2020;21(4):1306. doi:10.3390/ijms21041306
68. Liu L, Yao S, Mao X, Fang Z, Yang C, Zhang Y. Thermosensitive hydrogel coupled with sodium ascorbyl phosphate promot es human umbilical cord-derived mesenchymal stem cell-mediated skin wo und healing in mice. Scientific Reports. 2023;13(1):11909. doi:10.1038/s41598-023-38666-w
69. Azari Z, Nazarnezhad S, Webster TJ, et al. Stem cell-mediated angiogenesis in skin tissue engineering and wound healing. Wound Repair and Regeneration: Official Publication of the Wound Healing Society [And] the European Tissue Repair Society. 2022;30(4):421–435. doi:10.1111/wrr.13033
70. Yu X, Liu P, Li Z, Zhang Z. Function and mechanism of mesenchymal stem cells in the healing of diabetic foot wounds. Frontiers in Endocrinology. 2023;14:1099310. doi:10.3389/fendo.2023.1099310
71. Liu J, Zhang Y, Liu C, et al. A single dose of VEGF-A circular RNA sustains in situ long-term expres sion of protein to accelerate diabetic wound healing. Journal of Controlled Release: Official Journal of the Controlled Rel Ease Society. 2024;373:319–335. doi:10.1016/j.jconrel.2024.07.018
72. Zhang QZ, Su WR, Shi SH, et al. Human gingiva-derived mesenchymal stem cells elicit polarization of m2 macrophages and enhance cutaneous wound healing. Stem Cells. 2010;28(10):1856–1868. doi:10.1002/stem.503
73. Krepuska M, Mayer B, Vitale-Cross L, et al. Bone marrow stromal cell-derived hepcidin has antimicrobial and immunomodulatory activities. Scientific Reports. 2024;14(1):3986. doi:10.1038/s41598-024-54227-1
74. Liu Y, Zhang M, Liao Y, et al. Human umbilical cord mesenchymal stem cell-derived exosomes promote mu rine skin wound healing by neutrophil and macrophage modulations revea led by single-cell RNA sequencing. Frontiers in Immunology. 2023;14:1142088. doi:10.3389/fimmu.2023.1142088
75. Majidi M, Pakzad S, Salimi M, et al. Macrophage cell morphology-imprinted substrates can modulate mesenchymal stem cell behaviors and macrophage M1/M2 polarization for wound healing applications. Biotechnology and Bioengineering. 2023;120(12):3638–3654. doi:10.1002/bit.28546
76. Yu XT, Wang F, Ding JT, et al. Tandem mass tag-based serum proteomic profiling revealed diabetic foot ulcer pathogenesis and potential therapeutic targets. Bioengineered. 2022;13(2):3171–3182. doi:10.1080/21655979.2022.2027173
77. Lin CW, Hung CM, Chen WJ, et al. New Horizons of Macrophage Immunomodulation in the Healing of Diabetic Foot Ulcers. Pharmaceutics. 2022;14(10):2065. doi:10.3390/pharmaceutics14102065
78. Shi R, Lian W, Jin Y, et al. Role and effect of vein-transplanted human umbilical cord mesenchymal stem cells in the repair of diabetic foot ulcers in rats. Acta biochimica et biophysica Sinica. 2020;52(6):620–630. doi:10.1093/abbs/gmaa039
79. Chargaff E, West R. The biological significance of the thromboplastic protein of blood. The Journal of Biological Chemistry. 1946;166(1):189–197. doi:10.1016/S0021-9258(17)34997-9
80. Johnstone RM, Adam M, Hammond JR, Orr L, Turbide C. Vesicle formation during reticulocyte maturation. Association of plasma membrane activities with released vesicles (exosomes). The Journal of Biological Chemistry. 1987;262(19):9412–9420. doi:10.1016/S0021-9258(18)48095-7
81. Yi YW, Lee JH, Kim SY, et al. Advances in Analysis of Biodistribution of Exosomes by Molecular Imaging. International Journal of Molecular Sciences. 2020;21(2):665. doi:10.3390/ijms21020665
82. De Sousa KP, Rossi I, Abdullahi M, Ramirez MI, Stratton D, Inal JM. Isolation and characterization of extracellular vesicles and future di rections in diagnosis and therapy. Wiley Interdiscip Rev Nanomed Nanobiotechnol. 2023;15(1):e1835. doi:10.1002/wnan.1835
83. Al-Jipouri A, Eritja À, Bozic M. Unraveling the Multifaceted Roles of Extracellular Vesicles: insights into Biology, Pharmacology, and Pharmaceutical Applications for Drug Delivery. International Journal of Molecular Sciences. 2023;25(1). doi:10.3390/ijms25010485
84. Ibrahim SA, Khan YS. Histology, Extracellular Vesicles. StatPearls. Treasure Island (FL) Ineligible Companies. Disclosure: Yusuf Khan Declares No Relevant Financial relationships With Ineligible Companies. StatPearls Publishing; 2025.
85. Vidal M. Exosomes: revisiting their role as “garbage bags”. Traffic. 2019;20(11):815–828. doi:10.1111/tra.12687
86. Wu KY, Ahmad H, Lin G, Carbonneau M, Tran SD. Mesenchymal Stem Cell-Derived Exosomes in Ophthalmology: a Comprehensi ve Review. Pharmaceutics. 2023;15(4):1167. doi:10.3390/pharmaceutics15041167
87. Dai W, Hale SL, Kloner RA. Role of a paracrine action of mesenchymal stem cells in the improvement of left ventricular function after coronary artery occlusion in rats. Regenerative Medicine. 2007;2(1):63–68. doi:10.2217/17460751.2.1.63
88. Stremersch S, Vandenbroucke RE, Van Wonterghem E, Hendrix A, De Smedt SC, Raemdonck K. Comparing exosome-like vesicles with liposomes for the functional cellular delivery of small RNAs. Journal of Controlled Release: Official Journal of the Controlled Release Society. 2016;232:51–61. doi:10.1016/j.jconrel.2016.04.005
89. Wang J, Xia J, Huang R, et al. Mesenchymal stem cell-derived extracellular vesicles alter disease outcomes via endorsement of macrophage polarization. Stem Cell Research & Therapy. 2020;11(1):424. doi:10.1186/s13287-020-01937-8
90. Doyle LM, Wang MZ. Overview of Extracellular Vesicles, Their Origin, Composition, Purpose, and Methods for Exosome Isolation and Analysis. Cells. 2019;8(7):727. doi:10.3390/cells8070727
91. Isaac R, Reis FCG, Ying W, Olefsky JM. Exosomes as mediators of intercellular crosstalk in metabolism. Cell Metab. 2021;33(9):1744–1762. doi:10.1016/j.cmet.2021.08.006
92. Kalluri R, LeBleu VS. The biology, function, and biomedical applications of ex osomes. Science. 2020;367(6478):eaau6977. doi:10.1126/science.aau6977
93. Simpson RJ, Lim JW, Moritz RL, Mathivanan S. Exosomes: proteomic insights and diagnostic potential. Expert Review of Proteomics. 2009;6(3):267–283. doi:10.1586/epr.09.17
94. Kalani MYS, Alsop E, Meechoovet B, et al. Extracellular microRNAs in blood differentiate between ischaemic and haemorrhagic stroke subtypes. Journal of Extracellular Vesicles. 2020;9(1):1713540. doi:10.1080/20013078.2020.1713540
95. Hade MD, Suire CN, Suo Z. Mesenchymal Stem Cell-Derived Exosomes: applications in Regenerative Medicine. Cells. 2021;10(8):1.
96. Zhang Q, Jeppesen DK, Higginbotham JN, Franklin JL, Coffey RJ. Comprehensive isolation of extracellular vesicles and nanoparticles. Nat Protoc. 2023;18(5):1462–1487. doi:10.1038/s41596-023-00811-0
97. Veerman RE, Teeuwen L, Czarnewski P, et al. Molecular evaluation of five different isolation methods for extracell ular vesicles reveals different clinical applicability and subcellular origin. Journal of Extracellular Vesicles. 2021;10(9):e12128. doi:10.1002/jev2.12128
98. Wang W, Sun H, Duan H, et al. Isolation and usage of exosomes in central nervous system diseases. CNS Neurosci Ther. 2024;30(3):e14677. doi:10.1111/cns.14677
99. Lima Moura S, Martì M, Pividori MI. Matrix Effect in the Isolation of Breast Cancer-Derived Nanovesicles by Immunomagnetic Separation and Electrochemical Immunosensing-A Comparative Study. Sensors. 2020;20(4):965. doi:10.3390/s20040965
100. Lal A, Pike JFW, Polley EL, et al. Comparison of RNA content from hydrophobic interaction chromatography-isolated seminal plasma exosomes from intrauterine insemination (IUI) pregnancies. Andrologia. 2022;54(2):e14325. doi:10.1111/and.14325
101. Yang Z, Yang M, Rui S, et al. Exosome-based cell therapy for diabetic foot ulcers: present and prospect. Heliyon. 2024;10(20):e39251. doi:10.1016/j.heliyon.2024.e39251
102. Mondal J, Pillarisetti S, Junnuthula V, et al. Hybrid exosomes, exosome-like nanovesicles and engineered exosomes for therapeutic applications. Journal of Controlled Release: Official Journal of the Controlled Rel Ease Society. 2023;353:1127–1149. doi:10.1016/j.jconrel.2022.12.027
103. Wan T, Zhong J, Pan Q, Zhou T, Ping Y, Liu X. Exosome-mediated delivery of Cas9 ribonucleoprotein complexes for tissue-specific gene therapy of liver diseases. Science Advances. 2022;8(37):eabp9435. doi:10.1126/sciadv.abp9435
104. Kucharzewski M, Rojczyk E, Wilemska-Kucharzewska K, Wilk R, Hudecki J, Los MJ. Novel trends in application of stem cells in skin wound healing. Eur J Pharmacol. 2019;843:307–315. doi:10.1016/j.ejphar.2018.12.012
105. Jiang M, Jiang X, Li H, et al. The role of mesenchymal stem cell-derived EVs in diabetic wound healin g. Frontiers in Immunology. 2023;14:1136098. doi:10.3389/fimmu.2023.1136098
106. Than UTT, Guanzon D, Leavesley D, Parker T. Association of Extracellular Membrane Vesicles with Cutaneous Wound Healing. International Journal of Molecular Sciences. 2017;18(5):956. doi:10.3390/ijms18050956
107. Gurung S, Perocheau D, Touramanidou L, Baruteau J. The exosome journey: from biogenesis to uptake and intracellular signalling. Cell Communication and Signaling: CCS. 2021;19(1):47. doi:10.1186/s12964-021-00730-1
108. Matthay MA. Extracellular Vesicle Transfer from Mesenchymal Stromal Cells Modulates Macrophage Function in Acute Lung Injury. Basic Science and Clinical Implications. American Journal of Respiratory and Critical Care Medicine. 2017;196(10):1234–1236. doi:10.1164/rccm.201706-1122ED
109. Volarevic V, Markovic BS, Gazdic M, et al. Ethical and Safety Issues of Stem Cell-Based Therapy. International Journal of Medical Sciences. 2018;15(1):36–45. doi:10.7150/ijms.21666
110. Weiss DJ, English K, Krasnodembskaya A, Isaza-Correa JM, Hawthorne IJ, Mahon BP. The Necrobiology of Mesenchymal Stromal Cells Affects Therapeutic Efficacy. Frontiers in Immunology. 2019;10:1228. doi:10.3389/fimmu.2019.01228
111. Jiao Y-R, Chen K-X, Tang X, et al. Exosomes derived from mesenchymal stem cells in diabetes and diabetic complications. Cell Death & Disease. 2024;15(4):271. doi:10.1038/s41419-024-06659-w
112. Wen D, Peng Y, Liu D, Weizmann Y, Mahato RI. Mesenchymal stem cell and derived exosome as small RNA carrier and Imm unomodulator to improve islet transplantation. Journal of Controlled Release: Official Journal of the Controlled Rel Ease Society. 2016;238:166–175. doi:10.1016/j.jconrel.2016.07.044
113. Harrell CR, Jovicic N, Djonov V, Arsenijevic N, Volarevic V. Mesenchymal Stem Cell-Derived Exosomes and Other Extracellular Vesicle s as New Remedies in the Therapy of Inflammatory Diseases. Cells. 2019;8(12):1605. doi:10.3390/cells8121605
114. Nakamura Y, Kita S, Tanaka Y, et al. Adiponectin Stimulates Exosome Release to Enhance Mesenchymal Stem-Cel l-Driven Therapy of Heart Failure in Mice. Mol Ther. 2020;28(10):2203–2219. doi:10.1016/j.ymthe.2020.06.026
115. Weng Z, Zhang B, Wu C, et al. Therapeutic roles of mesenchymal stem cell-derived extracellular vesic les in cancer. J Hematol Oncol. 2021;14(1):136. doi:10.1186/s13045-021-01141-y
116. Xiong J, Hu H, Guo R, Wang H, Jiang H. Mesenchymal Stem Cell Exosomes as a New Strategy for the Treatment of Diabetes Complications. Frontiers in Endocrinology. 2021;12:646233. doi:10.3389/fendo.2021.646233
117. Guo J, Yang X, Chen J, et al. Exosomal miR-125b-5p derived from adipose-derived mesenchymal stem cells enhance diabetic hindlimb ischemia repair via targeting alkaline ceramidase 2. Journal of Nanobiotechnology. 2023;21(1):189. doi:10.1186/s12951-023-01954-8
118. Huang H, Zhu W, Huang Z, Zhao D, Cao L, Gao X. Adipose-derived stem cell exosome NFIC improves diabetic foot ulcers by regulating miR-204-3p/HIPK2. Journal of Orthopaedic Surgery and Research. 2023;18(1):687. doi:10.1186/s13018-023-04165-x
119. Li X, Xie X, Lian W, et al. Exosomes from adipose-derived stem cells overexpressing Nrf2 accelerate cutaneous wound healing by promoting vascularization in a diabetic foot ulcer rat model. Experimental & Molecular Medicine. 2018;50(4):1–14.
120. Li B, Luan S, Chen J, et al. The MSC-Derived Exosomal lncRNA H19 Promotes Wound Healing in Diabetic Foot Ulcers by Upregulating PTEN via MicroRNA-152-3p. Molecular Therapy Nucleic Acids. 2020;19:814–826. doi:10.1016/j.omtn.2019.11.034
121. Chen J, Li X, Liu H, et al. Bone marrow stromal cell-derived exosomal circular RNA improves diabetic foot ulcer wound healing by activating the nuclear factor erythroid 2-related factor 2 pathway and inhibiting ferroptosis. Diabetic Medicine: a Journal of the British Diabetic Association. 2023;40(7):e15031. doi:10.1111/dme.15031
122. Shi R, Jin Y, Zhao S, Yuan H, Shi J, Zhao H. Hypoxic ADSC-derived exosomes enhance wound healing in diabetic mice via delivery of circ-Snhg11 and induction of M2-like macrophage polarization. Biomedicine & Pharmacotherapy = Biomedecine & Pharmacotherapie. 2022;153:113463. doi:10.1016/j.biopha.2022.113463
123. Wu M, Tu J, Huang J, Wen H, Zeng Y, Lu Y. Exosomal IRF1-loaded rat adipose-derived stem cell sheet contributes to wound healing in the diabetic foot ulcers. Molecular Medicine. 2023;29(1):60. doi:10.1186/s10020-023-00617-6
124. Tang T, Chen L, Zhang M, et al. Exosomes derived from BMSCs enhance diabetic wound healing through circ-Snhg11 delivery. Diabetology & Metabolic Syndrome. 2024;16(1):37. doi:10.1186/s13098-023-01210-x
125. Yao X, Wei W, Wang X, Chenglin L, Björklund M, Ouyang H. Stem cell derived exosomes: microRNA therapy for age-related musculoskeletal disorders. Biomaterials. 2019;224:119492. doi:10.1016/j.biomaterials.2019.119492
126. Ti D, Hao H, Tong C, et al. LPS-preconditioned mesenchymal stromal cells modify macrophage polarization for resolution of chronic inflammation via exosome-shuttled let-7b. Journal of Translational Medicin. 2015;13(1):308. doi:10.1186/s12967-015-0642-6
127. Arabpour M, Saghazadeh A, Rezaei N. Anti-inflammatory and M2 macrophage polarization-promoting effect of mesenchymal stem cell-derived exosomes. International Immunopharmacology. 2021;97:107823. doi:10.1016/j.intimp.2021.107823
128. Su N, Hao Y, Wang F, Hou W, Chen H, Luo Y. Mesenchymal stromal exosome-functionalized scaffolds induce innate and adaptive immunomodulatory responses toward tissue repair. Science Advances. 2021;7(20). doi:10.1126/sciadv.abf7207
129. Bannon P, Wood S, Restivo T, Campbell L, Hardman MJ, Mace KA. Diabetes induces stable intrinsic changes to myeloid cells that contri bute to chronic inflammation during wound healing in mice. Dis Model Mech. 2013;6(6):1434–1447. doi:10.1242/dmm.012237
130. Sica A, Mantovani A. Macrophage plasticity and polarization: in vivo veritas. J Clin Invest. 2012;122(3):787–795. doi:10.1172/JCI59643
131. Venneri MA, Giannetta E, Panio G, et al. Chronic Inhibition of PDE5 Limits Pro-Inflammatory Monocyte-Macrophage Polarization in Streptozotocin-Induced Diabetic Mice. PLoS One. 2015;10(5):e0126580. doi:10.1371/journal.pone.0126580
132. Zhang XB, Chen XY, Qi J, et al. New Hope for Intervertebral Disc Degeneration: bone Marrow Mesenchymal Stem Cells and Exosomes Derived from Bone Marrow Mesenchymal Stem Cell Transplantation. Current Gene Therapy. 2022;22(4):291–302. doi:10.2174/1566523221666211012092855
133. Zhuang G, Meng C, Guo X, et al. A novel regulator of macrophage activation: miR-223 in obesity-associated adipose tissue inflammation. Circulation. 2012;125(23):2892–2903. doi:10.1161/CIRCULATIONAHA.111.087817
134. Essandoh K, Li Y, Huo J, Fan GC. MiRNA-Mediated Macrophage Polarization and its Potential Role in the Regulation of Inflammatory Response. Shock. 2016;46(2):122–131. doi:10.1097/SHK.0000000000000604
135. Xu J, Wu W, Zhang L, et al. The role of microRNA-146a in the pathogenesis of the diabetic wound-healing impairment: correction with mesenchymal stem cell treatment. Diabetes. 2012;61(11):2906–2912. doi:10.2337/db12-0145
136. Zhou X, Ye C, Jiang L, et al. The bone mesenchymal stem cell-derived exosomal miR-146a-5p promotes diabetic wound healing in mice via macrophage M1/M2 polarization. Molecular and Cellular Endocrinology. 2024;579:112089. doi:10.1016/j.mce.2023.112089
137. Zeuner M, Bieback K, Widera D. Controversial Role of Toll-like Receptor 4 in Adult Stem Cells. Stem Cell Reviews and Reports. 2015;11(4):621–634. doi:10.1007/s12015-015-9589-5
138. Liu Z, Hu L, Zhang T, et al. PKCβ increases ROS levels leading to vascular endothelial injury in diabetic foot ulcers. American Journal of Translational Research. 2020;12(10):6409–6421.
139. Dunnill C, Patton T, Brennan J, et al. Reactive oxygen species (ROS) and wound healing: the functional role o f ROS and emerging ROS-modulating technologies for augmentation of the healing process. Int Wound J. 2017;14(1):89–96. doi:10.1111/iwj.12557
140. Huang X, Shi L, Lin Y, et al. Pycnoporus sanguineus Polysaccharides as Reducing Agents: self- Assembled Composite Nanoparticles for Integrative Diabetic Wound Thera py. International Journal of Nanomedicine. 2023;18:6021–6035. doi:10.2147/IJN.S427055
141. Qi X, Cai E, Xiang Y, et al. An Immunomodulatory Hydrogel by Hyperthermia-Assisted Self-Cascade Glu cose Depletion and ROS Scavenging for Diabetic Foot Ulcer Wound Therap eutics. Adv Mater. 2023;35(48):e2306632. doi:10.1002/adma.202306632
142. Halim M, Halim A. The effects of inflammation, aging and oxidative stress on the pathoge nesis of diabetes mellitus (type 2 diabetes). Diabetes Metab Syndr. 2019;13(2):1165–1172. doi:10.1016/j.dsx.2019.01.040
143. Zhang Y, Bai X, Shen K, et al. Exosomes Derived from Adipose Mesenchymal Stem Cells Promote Diabetic Chronic Wound Healing through SIRT3/SOD2. Cells. 2022;11(16):1.
144. Kensler TW, Wakabayashi N, Biswal S. Cell survival responses to environmental stresses via the Keap1-Nrf2-ARE pathway. Annual Review of Pharmacology and Toxicology. 2007;47(1):89–116. doi:10.1146/annurev.pharmtox.46.120604.141046
145. Yagishita Y, Fukutomi T, Sugawara A, et al. Nrf2 protects pancreatic β-cells from oxidative and nitrosative stress in diabetic model mice. Diabetes. 2014;63(2):605–618. doi:10.2337/db13-0909
146. Ma T, Fu B, Yang X, Xiao Y, Pan M. Adipose mesenchymal stem cell-derived exosomes promote cell proliferation, migration, and inhibit cell apoptosis via Wnt/β-catenin signaling in cutaneous wound healing. Journal of Cellular Biochemistry. 2019;120(6):10847–10854. doi:10.1002/jcb.28376
147. Zhang Y, Han F, Gu L, et al. Adipose mesenchymal stem cell exosomes promote wound healing through accelerated keratinocyte migration and proliferation by activating the AKT/HIF-1α axis. Journal of Molecular Histology. 2020;51(4):375–383. doi:10.1007/s10735-020-09887-4
148. Ren S, Chen J, Guo J, et al. Exosomes from Adipose Stem Cells Promote Diabetic Wound Healing through the eHSP90/LRP1/AKT Axis. Cells. 2022;11(20):3229. doi:10.3390/cells11203229
149. Hsu HH, Wang AYL, Loh CYY, Pai AA, Kao HK. Therapeutic Potential of Exosomes Derived from Diabetic Adipose Stem Cells in Cutaneous Wound Healing of db/db Mice. Pharmaceutics. 2022;14(6):1206. doi:10.3390/pharmaceutics14061206
150. Carmeliet P. Angiogenesis in health and disease. Nature Medicine. 2003;9(6):653–660. doi:10.1038/nm0603-653
151. Chen CY, Rao SS, Ren L, et al. Exosomal DMBT1 from human urine-derived stem cells facilitates diabetic wound repair by promoting angiogenesis. Theranostics. 2018;8(6):1607–1623. doi:10.7150/thno.22958
152. Müller H, Nagel C, Weiss C, Mollenhauer J, Poeschl J. Deleted in malignant brain tumors 1 (DMBT1) elicits increased VEGF and decreased IL-6 production in type II lung epithelial cells. BMC Pulmonary Medicine. 2015;15(1):32. doi:10.1186/s12890-015-0027-x
153. Shen S, Liu H, Wang Y, et al. Long non-coding RNA CRNDE promotes gallbladder carcinoma carcinogenesis and as a scaffold of DMBT1 and C-IAP1 complexes to activating PI3K-AKT pathway. Oncotarget. 2016;7(45):72833–72844. doi:10.18632/oncotarget.12023
154. Yang J, Chen Z, Pan D, Li H, Shen J. Umbilical Cord-Derived Mesenchymal Stem Cell-Derived Exosomes Combined Pluronic F127 hydrogel Promote Chronic Diabetic Wound Healing and Complete Skin Regeneration. International Journal of Nanomedicine. 2020;15:5911–5926. doi:10.2147/IJN.S249129
155. Yu M, Liu W, Li J, et al. Exosomes derived from atorvastatin-pretreated MSC accelerate diabetic wound repair by enhancing angiogenesis via AKT/eNOS pathway. Stem Cell Research & Therapy. 2020;11(1):350. doi:10.1186/s13287-020-01824-2
156. Hu Y, Tao R, Chen L, et al. Exosomes derived from pioglitazone-pretreated MSCs accelerate diabetic wound healing through enhancing angiogenesis. Journal of Nanobiotechnology. 2021;19(1):150. doi:10.1186/s12951-021-00894-5
157. Shi R, Jin Y, Hu W, et al. Exosomes derived from mmu_circ_0000250-modified adipose-derived mesenchymal stem cells promote wound healing in diabetic mice by inducing miR-128-3p/SIRT1-mediated autophagy. American Journal of Physiology Cell Physiology. 2020;318(5):C848–c856. doi:10.1152/ajpcell.00041.2020
158. Wei P, Zhong C, Yang X, et al. Exosomes derived from human amniotic epithelial cells accelerate diabetic wound healing via PI3K-AKT-mTOR-mediated promotion in angiogenesis and fibroblast function. Burns & Trauma. 2020;8:tkaa020. doi:10.1093/burnst/tkaa020
159. Wang J, Wu H, Peng Y, et al. Hypoxia adipose stem cell-derived exosomes promote high-quality healing of diabetic wound involves activation of PI3K/Akt pathways. Journal of Nanobiotechnology. 2021;19(1):202. doi:10.1186/s12951-021-00942-0
160. Wang L, Hu L, Zhou X, et al. Exosomes secreted by human adipose mesenchymal stem cells promote scarless cutaneous repair by regulating extracellular matrix remodelling. Scientific Reports. 2017;7(1):13321. doi:10.1038/s41598-017-12919-x
161. Zhang J, Guan J, Niu X, et al. Exosomes released from human induced pluripotent stem cells-derived MSCs facilitate cutaneous wound healing by promoting collagen synthesis and angiogenesis. Journal of Translational Medicine. 2015;13(1):49. doi:10.1186/s12967-015-0417-0
162. Hu L, Wang J, Zhou X, et al. Exosomes derived from human adipose mensenchymal stem cells accelerates cutaneous wound healing via optimizing the characteristics of fibroblasts. Scientific Reports. 2016;6(1):32993. doi:10.1038/srep32993
163. Yang X, Wang J, Guo SL, et al. miR-21 promotes keratinocyte migration and re-epithelialization during wound healing. International Journal of Biological Sciences. 2011;7(5):685–690. doi:10.7150/ijbs.7.685
164. Lv Q, Deng J, Chen Y, Wang Y, Liu B, Liu J. Engineered Human Adipose Stem-Cell-Derived Exosomes Loaded with miR-21-5p to Promote Diabetic Cutaneous Wound Healing. Molecular Pharmaceutics. 2020;17(5):1723–1733. doi:10.1021/acs.molpharmaceut.0c00177
165. Ren S, Chen J, Duscher D, et al. Microvesicles from human adipose stem cells promote wound healing by optimizing cellular functions via AKT and ERK signaling pathways. Stem Cell Research & Therapy. 2019;10(1):47. doi:10.1186/s13287-019-1152-x
166. Teng L, Maqsood M, Zhu M, et al. Exosomes Derived from Human Umbilical Cord Mesenchymal Stem Cells Accelerate Diabetic Wound Healing via Promoting M2 Macrophage Polarization, Angiogenesis, and Collagen Deposition. International Journal of Molecular Sciences. 2022;23(18):10421. doi:10.3390/ijms231810421
167. Zhang B, Wang M, Gong A, et al. HucMSC-Exosome Mediated-Wnt4 Signaling Is Required for Cutaneous Wound Healing. Stem Cells. 2015;33(7):2158–2168. doi:10.1002/stem.1771
168. Zhang Y, Pan Y, Liu Y, et al. Exosomes derived from human umbilical cord blood mesenchymal stem cells stimulate regenerative wound healing via transforming growth factor-β receptor inhibition. Stem Cell Research & Therapy. 2021;12(1):434. doi:10.1186/s13287-021-02517-0
169. Safari B, Aghazadeh M, Davaran S, Roshangar L. Exosome-loaded hydrogels: a new cell-free therapeutic approach for skin regeneration. European Journal of Pharmaceutics and Biopharmaceutics: Official Journal of Arbeitsgemeinschaft Fur Pharmazeutische Verfahrenstechnik eV. 2022;171:50–59. doi:10.1016/j.ejpb.2021.11.002
170. Chen Y, Huang J, Chen R, et al. Sustained release of dermal papilla-derived extracellular vesicles from injectable microgel promotes hair growth. Theranostics. 2020;10(3):1454–1478. doi:10.7150/thno.39566
171. Brumberg V, Astrelina T, Malivanova T, Samoilov A. Modern Wound Dressings: hydrogel Dressings. Biomedicines. 2021;9(9):1235. doi:10.3390/biomedicines9091235
172. Li M, Ke QF, Tao SC, Guo SC, Rui BY, Guo YP. Fabrication of hydroxyapatite/chitosan composite hydrogels loaded with exosomes derived from miR-126-3p overexpressed synovial mesenchymal stem cells for diabetic chronic wound healing. Journal of Materials Chemistry B. 2016;4(42):6830–6841. doi:10.1039/C6TB01560C
173. Wang C, Liang C, Wang R, et al. The fabrication of a highly efficient self-healing hydrogel from natural biopolymers loaded with exosomes for the synergistic promotion of severe wound healing. Biomaterials Science. 2019;8(1):313–324. doi:10.1039/C9BM01207A
174. Shi Q, Qian Z, Liu D, et al. GMSC-Derived Exosomes Combined with a Chitosan/Silk Hydrogel Sponge Accelerates Wound Healing in a Diabetic Rat Skin Defect Model. Frontiers in Physiology. 2017;8:904. doi:10.3389/fphys.2017.00904
175. Chen Z, Zhang B, Shu J, et al. Human decellularized adipose matrix derived hydrogel assists mesenchymal stem cells delivery and accelerates chronic wound healing. Journal of Biomedical Materials Research Part A. 2021;109(8):1418–1428. doi:10.1002/jbm.a.37133
176. Zhao D, Yu Z, Li Y, Wang Y, Li Q, Han D. GelMA combined with sustained release of HUVECs derived exosomes for promoting cutaneous wound healing and facilitating skin regeneration. Journal of Molecular Histology. 2020;51(3):251–263. doi:10.1007/s10735-020-09877-6
177. Saldin LT, Cramer MC, Velankar SS, White LJ, Badylak SF. Extracellular matrix hydrogels from decellularized tissues: structure and function. Acta biomaterialia. 2017;49:1–15. doi:10.1016/j.actbio.2016.11.068
178. Verma D, Okhawilai M, Nangan S, et al. A sustainable and green approach towards the utilization of biopolymers for effective wound dressing applications: a detailed review. Nano-Structures & Nano-Objects. 2024;37:101086. doi:10.1016/j.nanoso.2023.101086
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms.php
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms.php
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Empagliflozin-Pretreated MSC-Derived Exosomes Enhance Angiogenesis and Wound Healing via PTEN/AKT/VEGF Pathway
Wang H, Bai Z, Qiu Y, Kou J, Zhu Y, Tan Q, Chen C, Mo R
International Journal of Nanomedicine 2025, 20:5119-5136
Published Date: 22 April 2025
Tailored Extracellular Vesicles from Dental Stem Cells: Advances in Specific Modifications for Enhanced Therapeutic Applications
Liu X, Li Z, Fu J, Wang R, He J, Yao J, Ye Q, He Y
International Journal of Nanomedicine 2025, 20:8327-8341
Published Date: 26 June 2025